6 KERUSAKAN BAKTERI OLEH SENYAWA ANTIBAKTERI DARI
EKSTRAK Chaetoceros gracilis
6.1 Pendahuluan 6.1.1 Latar belakang
Mikroalga merupakan biota perairan yang selama ini pemanfaatannya di Indonesia masih terbatas untuk pakan larva. Sesungguhnya mikroalga mempunyai potensi untuk dikembangkan karena dapat menghasilkan komponen aktif dan kandungan kimia yang cukup potensial. Rosa et al. (2005) menyatakan bahwa mikroalga telah lama dikenal karena memiliki aktivitas biologikal seperti pigmen, vitamin, lemak, sterol dan protein, selain itu juga menjadi sumber yang potensial untuk produk komersial di bidang akuakultur dan kosmetika.
Salah satu jenis mikroalga yang memiliki aktifitas biologikal adalah Chaetoceros. Chaetoceros gracilis merupakan salah satu mikroalga laut yang menghasilkan komponen aktif seperti antibakteri yang mana merupakan antibakteri alami yang aman penggunaannya. Hasil penelitian Pribadi (1998) menunjukkan bahwa Chaetoceros gracilis memiliki aktivitas antibakteri terhadap Bacillus subtilis, Escherichia coli dan Pseudomonas sp.
Komponen yang mempunyai aktivitas antibakterial dalam Chaetoceros tergolong asam lemak (Metting dan Pyne 1986; Wang 1999. Komponen aktif pada Chaetoceros dapat menghambat bakteri Gram negatif dan positif (Wang 1999). Penelitian sebelumnya menunjukkan bahwa Chaetoceros gracilis yang ditumbuhkan dalam medium NPSi memiliki aktivitas antibakteri yang dapat menghambat pertumbuhan bakteri Gram postif Staphylococcus aureus dan Bacillus cereus serta bakteri Gram negatif Vibrio harveyi dan Escherichia coli.
Efektivitas antibakteri untuk setiap bakteri tidak sama, karena masing-masing bakteri memiliki struktur dinding sel yang berbeda. Struktur dinding sel bakteri Gram positif berbeda dengan bakteri Gram negatif. Pada bakteri Gram positif mengandung 90% peptidoglikan serta lapisan tipis asam teikoat dan teikuronat. Bakteri Gram negatif memiliki lapisan di luar dinding sel yang mengandung 5 -10% peptidoglikan, selain itu juga terdiri dari protein, lipopolisakarida dan lipoprotein. Bakteri Gram negatif mempunyai dua lapisan lipid (bilayer lipid) yang disebut lapisan lipopolisakarida (LPS). Lapisan ini tersusun atas fosfolipid, polisakarida dan protein (Madigan et al. 2003). Polisakarida dalam dinding sel biasanya mengandung asam amino
N-asetilglukosamin dan asam N-asetilmuramat. Pada gula amino ini terikat rantai-rantai peptida pendek. Lapisan peptidoglikan lebih tebal (40 lapisan) pada dinding sel bakteri Gram positif daripada dinding sel bakteri Gram negatif (1-5 lapisan) (Lewis et al. 2007). Bakteri Gram negatif memiliki dua lapisan lipid yang dipisahkan oleh peptidoglikan. Ada juga outer membrane yang menempel pada lapisan lipopolisakarida memperkuat sel dan melindungi dari lingkungan luar. Pada membran ini ada porin dengan diameter 1-2 mm yang mengatur akses larutan ke membran sitoplasma (Moat et al. 2002).
Antimikroba dapat merusak membran sitoplasma dan mempengaruhi integritasnya. Kerusakan pada membran dapat menyebabkan terjadinya peningkatan permeabilitas dan terjadi kebocoran sel, yang diikuti dengan keluarnya materi intraselular. Minyak atsiri dapat bereaksi dengan fosfolipid dari membran sel yang menyebabkan permeabilitas meningkat dan unsur pokok penyusun sel hilang (Kim et al. 1995). Setiap zat yang mampu merusak dinding sel atau mencegah sintetisnya akan menyebabkan sel peka terhadap tekanan osmotik. Adanya tekanan osmotik dalam sel bakteri akan menyebabkan terjadinya lisis (Setyabudi dan Gan 1995). Asam lemak dapat menghambat pertumbuhan sel bakteri Gram positif Staphylococcus aureus dan S. pyogenes, serta bakteri Gram negatif Escherichia coli dan Pseudomonas aeruginosa. Mekanisme penghambatan antibakteri asam lemak ini belum jelas tetapi diduga mengganggu sintesis asam lemak (Zheng et al. 2005).
Setiap jenis bakteri memiliki sensitifitas yang berbeda terhadap komponen aktif atau zat antimikroba. Mekanisme hambatan senyawa aktif terhadap bakteri juga berbeda-beda. Pada penelitian ini dilakukan kajian mekanisme kerusakan sel bakteri patogen setelah dikontakkan dengan ekstrak C. gracilis yang ditumbuhkan dalam medium NPSi.
6.1.2 Tujuan
Tujuan dari penelitian ini adalah menganalisis kerusakan bakteri yang meliputi kebocoran sel (protein dan asam nukleat), gangguan dinding sel, serta morfologi sel bakteri setelah kontak dengan ekstrak Chaetoceros gracilis.
6.2 Bahan dan Metode 6.2.1 Bahan dan alat (1) Bahan baku
Chaetoceros gracilis yang telah diekstraksi dan beberapa bakteri uji yang meliputi bakteri Gram positif (Bacillus cereus ATCC 13091, Staphylococcus aureus ATCC 25923) serta bakteri Gram negatif (Escherichia coli ATCC 25922 dan Vibrio harveyi).
(2) Alat
Alat-alat yang digunakan pada tahap percobaan ini juga sama dengan alat-alat yang digunakan pada tahap sebelumnya. Untuk analisis mekanisme hambatan digunakan alat-alat seperti water bath shaker, spektrofotometer, sentrifus, mikroskop, mikroskop elektron (JEOL JIM 5310nLV), dan alat gelas yang digunakan di laboratorium.
6.2.2 Metode penelitian
Mekanisme hambatan atau kerusakan sel bakteri akibat kontak dengan ekstrak Chaetoceros gracilis dilakukan terhadap bakteri Bacillus cereus dan Vibrio harveyi yang memiliki aktivitas antibakteri paling besar (daerah hambatan paling besar). Pengamatan mekanisme kerja ekstrak dilakukan dengan cara menganalisis kerusakan dinding sel bakteri dengan cara mengukur zat pembentuk dinding sel dan menganalisis kerusakan membran sel dengan cara mengamati kebocoran sel, serta mengamati morfologi sel sebelum dan setelah kontak dengan ekstrak Chaetoceros gracilis.
6.2.3 Prosedur analisis
Metode untuk mengamati kerusakan tersebut antara lain dengan mengukur pra zat penyusun dinding sel (N-asetil glukosamin), menganalisis kebocoran sel bakteri dan menganalisis morfologi sel bakteri menggunakan scanning electron microscopy (SEM).
(1) Analisis N-asetil-glukosamin (Reissig 1955 yang diacu Bintang 1993) Percobaan ini dilakukan dengan tujuan untuk menentukan pengaruh senyawa antibakteri (ekstrak Chaetoceros gracilis) terhadap dinding sel bakteri dengan cara mengukur kadar N-asetil-glukosamin sebagai prazat mukopeptida penyusun dinding sel.
Sebanyak 250 µg bakteri uji dicampur dengan 3 ml larutan antibakteri (ekstrak Chaetoceros gracilis), dalam air suling steril dengan kadar 40 µg/ml dan
diinkubasi pada suhu 37 oC selama 1 jam, lalu disentrifugasi 7000 rpm pada 4 oC
selama 10 menit. Kemudian sel bakteri tersebut dibilas dengan air suling steril
dan disentrifugasi 7000 rpm pada 4 oC selama 10 menit. Sebagai pembanding
digunakan sel bakteri sama tanpa antibakteri (ekstrak C. gracilis) dan langsung dibilas dengan air suling steril. Masing-masing perlakuan ditambahkan 0,5 ml
TCA (Trichloro Acid) 10 % suhu 4 oC dan diinkubasi pada suhu 4 oC selama 1
jam, lalu disentrifugasi 7000 rpm selama 10 menit.
Fase cair ditambahkan eter dengan volume yang sama untuk mengeluarkan TCA, dengan cara mengocok campuran ini pada vorteks dan dibiarkan supaya eter terpisah, lalu eter dibuang. Larutan bebas TCA ditambahkan 75 µl HCl 0,25 N dan dimasukkan ke dalam penangas air mendidih
selama 5 menit. Lalu ditambah 150 µl NaOH 0,125 N dalam Na2B4O7 2% dan
dipanaskan pada penangas air mendidih selama 7 menit. Kemudian dicampur dengan 1350 µl dimetil aminobenzaldehida 1 % dalam campuran asam asetat dan asam klorida dengan perbandingan 95 : 5, lalu dibiarkan selama 20 menit
pada suhu 37 oC, selanjutnya dibaca serapan optiknya pada panjang gelombang
550 nm. Bila terjadi kekeruhan, artinya terjadi penimbunan N-asetil glukosamin. (2) Analisis kebocoran sel bakteri (Bunduki et al. 1995)
Pengamatan kebocoran sel dilakukan untuk mempelajari bagaimana ekstrak mengganggu permeabilitas membran sel. Mekanisme perusakan membran sel merupakan salah satu tanda tidak normalnya sel setelah ada perlakuan ekstrak. Analisa kebocoran sel dilakukan dengan menggunakan alat Spektro UV-VIS RS Digital Spectrophotometer LaboMed, Inc. pada panjang gelombang 280 nm dan 260 nm. Panjang gelombang 280 nm digunakan untuk mengukur kadar nitrogen dari protein sel, sedangkan panjang gelombang 260 nm untuk mengukur kadar nitrogen dari nukleus sel.
Sebanyak 10 ml kultur murni disentrifugasi selama 10 menit. Filtrat dibuang lalu ditambahkan 5 ml larutan garam fisiologis (0,85% NaCl) dalam endapan sel pada tabung reaksi, kemudian diaduk menggunakan vorteks agar sel homogen dalam larutan fisiologis. Selanjutnya ditambahkan ekstrak dan dibiarkan selama 24 jam. Sebagai pembanding digunakan sel bakteri sama tanpa penambahan ekstrak. Selanjutnya suspensi disentrifugasi pada 10 000 rpm selama 10 menit dan supernatan disaring dengan kertas saring untuk
memisahkan selnya. Analisis dilakukan dengan mengamati OD dari cairan supernatan, menggunakan spektrofotometer (Spectro UV-Vis RS) pada panjang gelombang 280 dan 260 nm.
(3) Analisis perubahan morfologi sel bakteri (Bozolla dan Russel 1992)
Analisis perubahan morfologi sel dilakukan untuk mempelajari perubahan morfologi terhadap struktur sel akibat penggunaan ekstrak yang mengandung senyawa antibakteri, yang meliputi kerusakan morfologi sel, struktur bakteri, serta kerusakan dinding sel. Mula-mula bakteri dibuat tersuspensi dalam ekstrak, kemudian diinkubasi pada inkubator goyang dengan kecepatan 100 rpm. Selanjutnya cairan disentrifugasi dan dibuang supernatannya, lalu ditambahkan glutaraldehida 2% dan direndam. Kemudian disentrifugasi lagi, dibuang larutan fiksatif, lalu ditambahkan bufer caccodylate dan dibiarkan beberapa menit, disentrifugasi lagi, dibuang bufernya lalu ditambahkan osmium tetra oksida. Selanjutnya disentrifugasi lagi, dibuang larutannya, ditambahkan alkohol 50%, lalu ditambahkan alkohol lagi, disentrifugasi lagi, ditambahkan butanol. Kemudian dibuat ulasan suspeni pada cover slip, lalu dikeringkan. Selanjutnya spesimen yang sudah jadi dilihat menggunakan mikroskop elektron (SEM) JEOL,
JIM-5310 LV
.
6. 3 Hasil dan Pembahasan
6.3.1 Pengaruh ekstrak Chaetoceros gracilis terhadap kebocoran sel
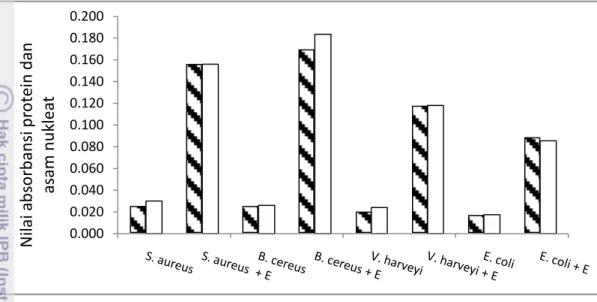
Kebocoran sel bakteri pada penelitian ini dimaksudkan untuk melihat kerusakan atau gangguan permeabilitas pada membran sel bakteri. Analisis kebocoran akibat pemberian ekstrak dilakukan dengan mengukur kekeruhan medium pertumbuhan bakteri yang telah diberi ekstrak dibandingkan tanpa ekstrak dengan menggunakan spektrofotometer. Kerusakan membran diukur dari bahan-bahan yang dilepaskan oleh sel bakteri yang dapat diserap pada panjang gelombang 260 nm (N nitrogen dalam asam nukleat) dan 280 nm (N nitrogen dalam protein). Mekanisme perusakan membran sel merupakan salah satu tanda tidak normalnya sel setelah ada perlakuan ekstrak. Hasil analisis kebocoran sel dapat dilihat pada Gambar 13.
Hasil penelitian (Gambar 13) menunjukkan bahwa nilai OD 260 nm dan OD280
nm pada semua bakteri uji dipengaruhi oleh penggunaan ekstrak C.gracilis.
Bakteri yang dikontakkan dengan ekstrak memiliki nilai OD lebih besar daripada tanpa ekstrak. Hal ini menunjukkan terjadinya pelepasan asam nukleat dan
protein ke dalam medium pertumbuhannya. Berdasarkan analisis ini dapat dikatakan bahwa sel bakteri uji mengalami kerusakan atau kebocoran akibat adanya ekstrak Chaetoceros gracilis.
Gambar 13 Pengaruh ekstrak C. gracilis terhadap kebocoran asam nukleat ( = OD 260 nm) dan kebocoran protein sel ( = OD 280 nm) Kebocoran sel bakteri terjadi diduga karena rusaknya ikatan hidrofobik komponen penyusun membran. Kim et al. (1995) menyatakan bahwa kebocoran sel terjadi karena ikatan hidrofobik yang terdiri dari komponen penyususn membran seperti protein dan fosfolipid rusak, serta larutnya komponen-komponen lain yang berikatan secara hidrofilik dan hidrofobik. Lin et al. (2000) juga menyatakan bahwa kondisi ini dapat meningkatkan permeabilitas membran sel, sehingga memudahkan masuknya komponen antibakteri ke dalam sel serta mengakibatkan keluarnya substansi sel seperti protein dan asam nukleat yang menyebabkan terjadinya kerusakan sel. Menurut Ultee et al. (1998) senyawa aktif dapat menyerang membran sitoplasma dan mempengaruhi integritas membran sitoplasma sehingga mengakibatkan kebocoran materi intraselular. Adanya gugus hidrofobik pada senyawa antimikroba menyebabkan perubahan komposisi dan pelarutan pada membran sel yang akhirnya membran mengalami kerusakan.
Pada penelitian ini terjadi kebocoran sel bakteri uji, yang menunjukkan terjadinya kerusakan membran sel bakteri. Bahan aktif dari C. gracilis yang berperan dalam penghambatan bakteri diduga asam lemak. Karena asam lemak dapat mengganggu membran bakteri. Zheng et al. (2005) melaporkan bahwa asam lemak dapat menghambat pertumbuhan sel bakteri Gram positif
0.000 0.020 0.040 0.060 0.080 0.100 0.120 0.140 0.160 0.180 0.200
N
ilai
ab
sor
b
an
si
p
ro
tein
d
an
asam
n
u
klea
t
Staphylococcus aureus dan S. pyogenes, serta bakteri Gram negatif Escherichia coli dan Pseudomonas aeruginosa. Mekanisme penghambatan antibakteri asam lemak belum jelas. Heat et al. (2001) menyatakan bahwa biosintesa lipid menjadi target untuk bahan antibakteri. Lipid merupakan komponen utama untuk pertumbuhan sel, sehingga biosintesis lipid merupakan target yang baik untuk intervensi terapeutik dalam penyakit yang disebabkan oleh bakteri Gram negatif. Komposisi lipid pada bakteri lebih sederhana dibanding manusia, oleh karena itu sangat ideal untuk pengembangan obat baru.
Ekstrak C. gracilis pada penelitian ini mengandung asam lemak jenuh seperti kaprilat, miristat, palmitat, laurat, miristoleat, pentadekanoat, stearat, heneikosanoat, behenat, serta asam lemak tidak jenuh seperti palmitoleat, heptadekanoat, elaidat, oleat, linoleat, arakhidonat, linolenat, dokosadienoat, eikosapentaenoat dan dokosaheksaenoat. Menurut Zheng et al. (2005) asam lemak tidak jenuh seperti asam palmitoleat, asam oleat, asam linolenat dan asam arakhidonat, serta asam lemak jenuh stearat memiliki aktivitas antibakteri. Hal ini sesuai dengan yang dinyatakan Metting dan Pyne (1986) serta Wang (1999) dimana komponen aktif yang dimiliki Chaetoceros adalah golongan asam lemak. Zheng et al. (2005) menyatakan bahwa mekanisme aktivitas antibakteri jenis
asam lemak belum jelas. Asam lemak tidak jenuh rantai panjang (C16-C20)
memiliki aktivitas antibakteri terhadap Staphylococcus, Streptococcus,
Mycobacterium, Helicobacter, dan Bacilli. Dilika et al. (2000) melaporkan bahwa asam lemak linoleat dan oleat memiliki aktivitas antibakteri yang dapat melawan Bacillus megaterium dengan MIC 0,2 dan 0,05 mM. Kedua asam lemak ini juga menghambat pertumbuhan Pseudomonas phaseolicola. Selain itu asam lemak linoleat juga mempunyai aktivitas penghambatan terhadap Streptococcus mutans dan B. larvae. Kedua asam lemak ini mempunyai aktivitas sinergistik.
Chaetoceros gracilis mengandung asam lemak antara lain asam palmitoleat, asam oleat, asam linoleat, asam linolenat, asam arakhidonat, asam stearat yang diduga memiliki aktivitas antibakteri. Menurut Zheng et al. (2005) asam lemak tidak jenuh menunjukkan aktivitas penghambatan lebih besar dibanding asam lemak jenuh. Asam linoleat menunjukkan aktivitas antibakterial yang merupakan antimikroba pada bahan pangan tambahan dan antibakteri dalam herbal. Asam linoleat ini juga diduga menghambat pertumbuhan dengan cara meningkatkan permeabilitas membran bakteri, tetapi reaksi mekanisme hambatannya belum jelas. Senyawa aktif dalam triclosan adalah asam linoleat
yang telah ditargetkan sebagai biocide yang memiliki spektrum luas, dimana digunakan sebagai bahan tambahan antibakteri yang berperan sebagai biocide non spesifik (Zheng et al. 2005). Adanya kandungan asam lemak yang memiliki aktivitas antibakteri dalam Chaetoceros gracilis memerlukan penelitian lanjutan untuk pengembangan bidang farmasetika dan pangan.
6.3.2 Pengaruh ekstrak Chaetoceros gracilis terhadap dinding sel bakteri
Unit dasar dari dinding sel bakteri tersusun atas peptidoglikan, dimana memberikan kekuatan pada sel bakteri, selain itu berperan juga sebagai dasar membran sitoplasma. Peptidoglikan tersusun atas asetilglukosamin dan N-asetilmuramat serta beberapa asam amino seperti L-alanin, D-alanin, D-glutamat dan lisin. N-asetilglukosamin merupakan prazat mukopeptida pembentuk dinding sel bakteri, yang mana dapat terganggu oleh adanya antibiotik.
Kerusakan dinding sel bakteri dapat dilihat dengan mengukur prazat mukopeptida penyusun dinding sel yang ditandai dengan kekeruhan pada media. Penelitian ini bertujuan menentukan pengaruh penggunaan ekstrak Chaetoceros gracilis yang mempunyai aktivitas antibakteri terhadap kerusakan dinding sel bakteri. Hasil analisis prazat disajikan pada Gambar 14. Bakteri yang medium pertumbuhanya ditambah ekstrak Chaetoceros gracilis menghasilkan absorbansi (optical density) lebih besar dibanding tanpa penambahan ekstrak, artinya di dalam medium ada penimbunan N-asetil glukosamin sebagai prazat mukopetida penyusun dinding sel bakteri. Hal ini menunjukkan terjadinya gangguan atau kerusakan dalam dinding sel bakteri.
Gambar 14 Pengaruh ekstrak C. gracilis terhadap kandungan N-asetil glukosamin ( = tanpa ekstrak, = penambahan ekstrak).
0.000 0.010 0.020 0.030 0.040 0.050 0.060
S. aureus
B. cereus
V. harveyi
E. coli
Ab so rb an si N -aset il glu ko sam in
Struktur dinding sel bakteri Gram positif tidak sama dengan bakteri Gram negatif. Dinding sel bakteri Gram negatif memiliki dua lapisan lipid (bilayer lipid), yang disebut lapisan lipopolisakarida (LPS). Lapisan ini tersusun atas fosfolipid, polisakarida dan protein (Madigan et al. 2003). Hasil analisis prazat mukopeptida pembentuk dinding sel bakteri yang diduga N-asetil glukosamin menunjukkan bahwa ekstrak menyebabkan kerusakan dinding sel bakteri. Penelitian serupa telah dilakukan Bintang (1993) yang melaporkan bahwa senyawa aktif yang dihasilkan oleh Streptococcus lactis dapat menghambat pembentukan dinding sel bakteri Eschericha coli, mekanisme penghambatannya adalah menghambat kerja enzim fosfatase alkalis pada tahap awal pembentukan dinding sel bakteri, sehingga terjadi penimbunan pra zat pembentuk dinding sel.
Kandungan pra zat pembentuk dinding sel bakteri seperti N-asetilglukosamin yang ditunjukkan dengan hasil serapan optik pada bakteri S. aureus dan B. cereus lebih besar dibanding bakteri E. coli. Hal ini terjadi karena struktur dinding sel bakteri tersebut berbeda, sehingga efek antibakteri terhadap bakteri juga berbeda. Escherichia coli merupakan bakteri Gram negatif yang mempunyai dua lapisan lipid, sedangkan bakteri Gram positif seperti Stapylococcus aureus dan Bacillus cereus hanya memiliki satu lapisan, sehingga antibiotik lebih mudah menembus ke dalam sel bakteri Stapylococcus aureus dan Bacillus cereus.
Pada penelitian ini kontak ekstrak dengan bakteri dapat menyebabkan kerusakan dinding sel bakteri. Rusaknya dinding sel bakteri diduga karena adanya reaksi antara senyawa aktif dari ekstrak dengan dinding sel bakteri . Menurut Kabara et al. (1972) cara kerja obat antara lain merubah permeabilitas dari dinding sel. Hal ini dapat terjadi karena keluarnya nutrien atau terjadinya difusi metabolit esensial. Ultee et al. (1998) melaporkan bahwa mekanisme kerja antimikroba ada yang mempunyai spektrum luas, sempit dan ada yang hanya efektif terhadap mikroorganisme tertentu. Pengaruh antibiotik terhadap dinding sel dapat terjadi akibat akumulasi asam lemak maupun asam organik dari bahan (antimikroba) dalam bentuk tidak terdisosiasi akan menyebabkan perubahan terhadap komposisi penyusun dinding sel. Senyawa aktif dapat bereaksi dengan dinding sel bakteri dan membran sel. Selain itu kerusakan pada dinding sel bakteri juga dapat disebabkan oleh terjadinya tekanan osmotik.
6.3.3 Pengaruh ekstrak C gracilis terhadap morfologi sel bakteri (1) Bacillus cereus
Bacillus cereus adalah bakteri patogen, Gram positif berbentuk batang berspora, banyak ditemukan air, debu maupun tanah, yang mana sporanya tahan panas. Bakteri ini menghasilkan ekstraselular toksin dan enzim. Eksotoksin B. cereus dapat menyebabkan diare. Bahan pangan yang sering ditumbuhi bakteri ini antara lain nasi, susu, jagung, sayuran, daging, sosis, puding. Bakteri ini sensitif terhadap Butylated hydoxyanisole (BHA), pertumbuhannya dapat dihambat pada konsentrasi <500 ppm (Jay 2000).
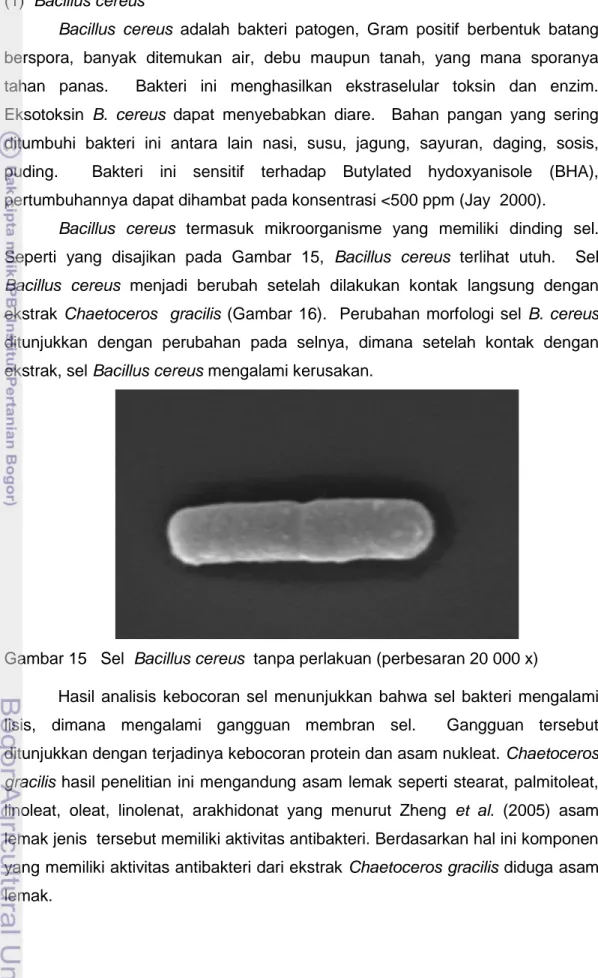
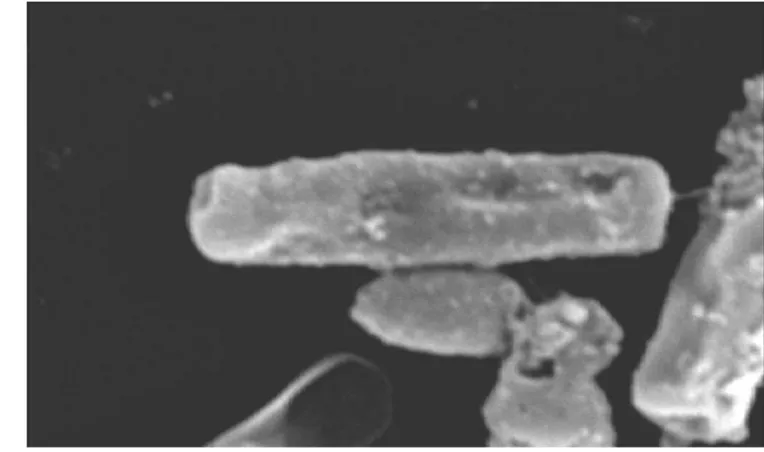
Bacillus cereus termasuk mikroorganisme yang memiliki dinding sel. Seperti yang disajikan pada Gambar 15, Bacillus cereus terlihat utuh. Sel Bacillus cereus menjadi berubah setelah dilakukan kontak langsung dengan ekstrak Chaetoceros gracilis (Gambar 16). Perubahan morfologi sel B. cereus ditunjukkan dengan perubahan pada selnya, dimana setelah kontak dengan ekstrak, sel Bacillus cereus mengalami kerusakan.
Gambar 15 Sel Bacillus cereus tanpa perlakuan (perbesaran 20 000 x)
Hasil analisis kebocoran sel menunjukkan bahwa sel bakteri mengalami lisis, dimana mengalami gangguan membran sel. Gangguan tersebut ditunjukkan dengan terjadinya kebocoran protein dan asam nukleat. Chaetoceros gracilis hasil penelitian ini mengandung asam lemak seperti stearat, palmitoleat, linoleat, oleat, linolenat, arakhidonat yang menurut Zheng et al. (2005) asam lemak jenis tersebut memiliki aktivitas antibakteri. Berdasarkan hal ini komponen yang memiliki aktivitas antibakteri dari ekstrak Chaetoceros gracilis diduga asam lemak.
Gambar 16 Sel Bacillus cereus yang dikontakkan dengan ekstrak C. gracilis
(perbesaran 20 000 x) (2) Vibrio harveyi
Vibrio harveyi merupakan bakteri Gram negatif yang sering menyebabkan gangguan kesehatan pada larva udang. Tingginya mortalitas larva di panti benih udang kebanyakan dikarenakan Luminescent vibriosis yang disebabkan oleh Vibrio harveyi atau Vibrio splendidus.
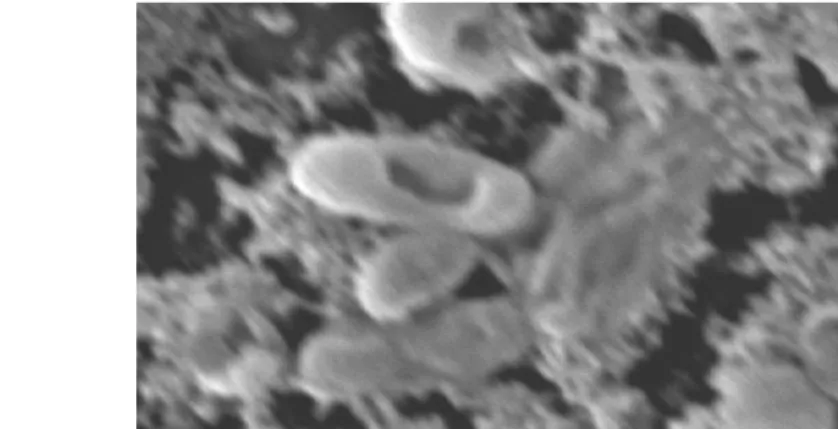
Hasil analisis kebocoran menunjukkan bahwa Vibrio harveyi mengalami kebocoran akibat kontak dengan ekstrak C. gracilis. Demikian juga hasil analisis prazat yang menunjukkan bahwa bakteri ini mengalami lisis. Hasil analisis biokimia ini didukung dengan hasil pengamatan menggunakan SEM. Sel bakteri yang tidak dikontakkan dengan ekstrak Chaetoceros gracilis terlihat utuh (Gambar 17), sedangkan yang dikontakkan dengan ekstrak Chaetoceros gracilis terlihat mengalami kerusakan (Gambar 18).
Gambar 17 Sel Vibrio harveyi tanpa perlakuan (perbesaran 20 000 x)
Beberapa peneliti telah melaporkan bahwa Chaetoceros memiliki antibakteri yang termasuk dalam golongan asam lemak. Chaetoceros gracilis pada penelitian ini juga mengandung asam lemak. Kabara et al. (1972)
menyatakan bahwa asam-asam lemak terutama asam laurat dapat menghambat enzim yang terlibat pada produksi energi dan pembentukan komponen struktural sehingga dapat mengganggu pembentukan dinding selnya. Mekanisme kerusakan dinding sel dapat disebabkan oleh adanya akumulasi komponen lipofilik yang terdapat pada dinding sel atau membran sel.
Gambar 18 Sel Vibrio harveyi yang dikontakkan dengan ekstrak Chaetoceros gracilis (perbesaran 20 000 x)
6.4 Kesimpulan
Berdasarkan hasil penelitian yang diperoleh dapat disimpulkan :
(1) Bakteri uji setelah kontak dengan ekstrak mengalami kebocoran sel.
(2) Kontak antara bakteri uji dengan ekstrak Chaetoceros gracilis
mengakibatkan kebocoran sel bakteri.
(3) Sel bakteri uji (B. cereus dan V. harveyi) mengalami perubahan (kerusakan)
morfologi setelah kontak dengan ekstrak Chaetoceros gracilis.
Berdasarkan hasil penelitian yang diperoleh, dapat disarankan untuk dilakukan penelitian lebih lanjut tentang reaksi mekanisme hambatan antibakteri.