BAB VII
A.
KOMPOSISI STRUKTUR TULANG DAN OTOT
Tulang merupakan jaringan ikat termineralisasi. Tulang terdiri atas bahan organik (protein) & anorganik. Bahan organik yaitu protein antara lain kolagen Tipe I (90-95%) dan Tipe V (sedikit) dan bahan anorganik/mineral terdiri atas kristalin hidrosiapatit Ca10[PO4]6[OH]2), dimana mengandung natrium, magnesium,karbonat, fluorida, 99% kalsium tubuh. Kristalin hidrosiapatit akan memberi kekuatan dan kelenturan.
Protein utama tulang berupa kolagen tipe I, terdiri motif protein yaitu poly-Asp dan poly-Glu yang mengikat kalsium dan memberi kerangka bagi mineralisasi. Struktur makromolekul berupa proteoglikan.
Pada remodeling tulang, dimana struktur dinamik diresorpsi pengendapan jaringan tulang yang baru, yang beradaptasi dengan sinyal fisik ( menahan beban tubuh) dan hormon. Sel yang berperan sebagai sel osteoklas (resorpsi) dan osteoblas (pengendapan tulang) . Osteoblas mengendalikan mineralisasi dengan mengatur pelintasan ion kalsium dan fosfat, enzim alkali fosfatase untuk menghasilkan ion fosfat . Peranan osteoklas dalam resorpsi tulang, enzim-enzim lisosomal hidorolase & ion H di lepas dalam ling mikro (pH rendah/asam), di matriks tulang osteoklas, disolusi kalsium fosfat, produk resorpsi tulang ke sitoplasma melalui kapiler darah.
Kelainan tulang yaitu osteogenesis Imperfekta ( tulang rapuh), terjadi defisiensi jaringan ikat, menyebabkan mutasi gen COL1A1 & COL1A2. Osteopetrosis ( penyakit tulang pualam) terjadi peningkatan densitas tulang, akibat ketidakmampuan resorpsi tulang menyebabkan mutasi gen yang mengkode karbonik anhidrase.
Komposisi biokimia jaringan otot terdiri atas protein otot yang berfungsi struktural untuk sistem biologis. Otot merupakan transducer esi ioki ia uta a ya g merubah
energi potensial (kimia) menjadi energi kinetik (mekanisme). Jaringan tunggal terbesar : 20% massa tubuh baru lahir, 40% massa tubuh dewasa dan 30% massa tubuh tua. Otot sebagai transducer kimia mekanis memiliki syarat yaitu ada suplai energi kimia yang konstan (ATP, kreatin fosfat), ada pengaturan aktivitas mekanis (kecepatan, lama dan kekuatan kontraksi otot), mesin berhubungan dengan operator melalui sistem syaraf, bisa digunakan lebih dari satu kali dan dapat dikembalikan ke keadaan semula.
Syarat diatas dipenuhi oleh 3 tipe otot terdiri atas otot rangka, otot jantung/lurik, otot polos . Umumnya otot berfungsi sebagai penarik bukan pendorong. Otot rangka bersifat volunter syaraf. Otot polos dan jantung involunter.
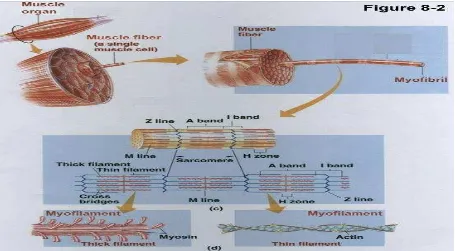
Komposisi biokimia jaringan otot yaitu otot lurik terdiri atas serabut sel otot berinti banyak, dikelilingi membran yang peka terhadap listrik membentuk sarkolema yang direntangkan terdiri miofibril dalam sarkoplasma terdapat dalam cairan intrasel, serta mengandung ATP dan fosfokreatin.
Gambar 7.1 Struktur otot
Aktin merupakan monomer G-aktin, struktur protein globuler, BM 43.000, 25% berat protein otot, terdapat magnesium serta berpolimerisasi membentuk G-aktin berpolimerisasi. Aktin F (6-7 nm) mempunyai struktur berulang setiap 35,5 nm. Ada 4 Protein lain bagian kecil dari massa otot lurik yaitu tropomiosin yang terdiri dari molekul fibrosa 2 rantai dan pada celah aktin-F terdapat disemua otot, troponin yang merupakan unit untuk otot lurik terdiri 3 protein berupa troponin T (TpT) terikat pada tropomiosin, troponin I (TpI) menghambat interaksi aktif F-miosin dan troponin C (TpC) protein pengikat 4 kalsium ((mirip kalmodulin) dan filamen tipis otot lurik Aktin-F, tropomiosin dan 3 komponen troponin.
Gambar 7.2 Aktin dan miosin
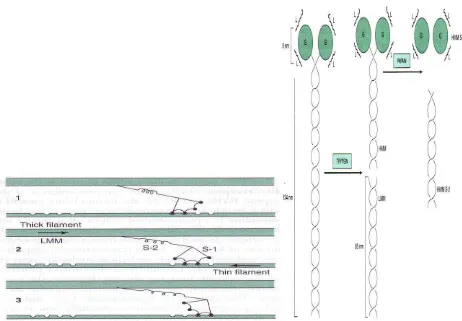
Miosin memiliki 55% berat protein otot dan membentuk filamen tebal berupa heksamer, BM 460.000, mempunyai bagian fibrosa 2 heliks membelit dengan kepala globuler pada ujung heliks, BM sepasang heksamer rantai berat 200.000, BM sepasang heksamer rantai ringan 15.000-27.000. Miosin otot beraktivitas menghidrolisis ATP (ATP-ase) yang terikat pada aktinF. Miosin dipecah oleh tripsin menjadi 2 fragmen yosin yaitu :
1. Meromiosin ringan (LMM), light meromiosin, terdiri dari kumpulan serabut -heliks tidak larut dan tidak ada aktivitas ATPase dan tidak terikat aktin-F
2. Meromiosin berat (HMM), heavy meromiosin merupakan protein larut, BM 34.000, mempunyai bagian fibrosa dan globuler dan ada aktivitas ATPase dan terikat pada aktin F
Gambar 7.4 meromiosin ringan dan berat Ada 2 Subfragmen HMM
S1 : BM 115.000, fibrosa, ada aktitvitas ATP-ase
Gambar 7.5 Aktivitas subfragmen meromiosin berat
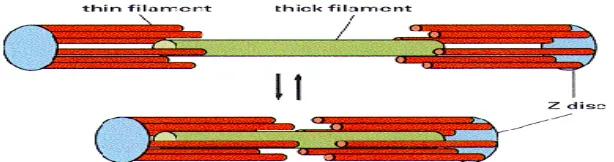
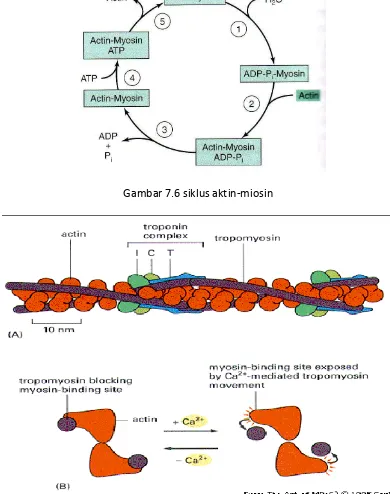
Siklus biokimia kontraksi otot kepala iosi e ghidrolisis ATP → ADP + Pi, kepala miosin yang mengandung ADP dan Pi berotasi mengikat aktin-F → 900 C, ADP dan Pi dilepas dari
aktin-miosin 450 C dengan menarik aktin ke pusat sarkomer , ATP baru terikat pada aktin-F
Gambar 7.6 siklus aktin-miosin
Gambar 7.7 Peranan Ca dalam pengikatan aktin-miosin
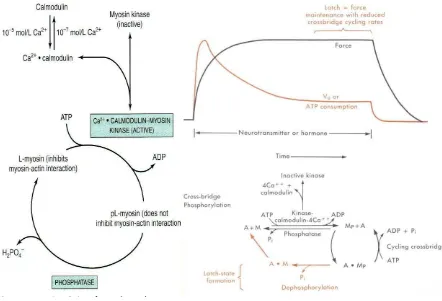
Pengaturan kontraksi dan Relaksasi Otot, dimana Ca2+ memegang peranan pengaturan
mencegah pengikatan kepala miosin ke tempat pengikatan aktin-F dengan merubah konformasi aktin-F bertujuan mencegah percepatan ATPase myosin.
Gambar 7.8.Filamen aktin dengan subfragmen S1
Relaksasi, Ca2+ sarkoplasma turun < 10-7 mol/L akibat Ca2+ kembali ke retikulum
sarkoplasma melalui pompa Ca2+ yang membutuhkan energi. TpC 4Ca2+ kehilangan Ca2+ .
Troponin berinteraksi dengan tropomiosin menghambat interaksi kepala miosin dengan aktin-F. Terlepas kepala miosin dari aktin F pada relaksasi. Ca2+ mengatur kontraksi otot dengan
mekanisme alosterik yang diperantarai TpC, TpI, TpT, tropomiosin dan aktin F.
Otot jantung terdiri atas cairan ekstrseluler sumber utama Ca2+ bila cairan ekstraseluler
tidak membasahi otot jantung kontraksi berhenti 1 menit dan pada otot rangka dapat bergerak beberapa jam tanpa Ca2+ ekstraseluler.
Gambar 7.9 Peranan Ca dalam kontraksi otot
Gambar 7.9 Mekanisme depolarisasi otot
Akibat hilangnya ATP dalam sarkoplasma, pada pompa Ca2+ dalam retikulum
sarkoplasma berhenti mempertahankan konsentrasi Ca2+ sarkoplasma yang rendah terjadi
interaksi miosin-Aktin F dipermudah. Pelepasan miosin dari aktin F yang memerlukan ATP tidak terjadi menyebabkan kaku mayat (rigor mortis). Pengaturan kontraksi berdasarkan myosin. Semua otot mempunyai : aktin, miosin dan tropomiosin keculai otot lurik vertebrata mempunyai troponin. Otot polos mempunyai molekul -aktinin dan tropomiosin, tetapi tidak mempunyai troponin. Kontraksi otot polos diatur oleh Ca2+ . Miosin otot polos terikat aktin-F
tidak ada tropomiosin serta tidak ada aktivitas ATPase. Mekanisme otot polos mencegah pengikatan kepala miosin ke aktin-F fosforilasi rantai ringan memulai siklus kontraksi pengikatan pelepasan dari otot polos.
Pengaturan kontraksi berdasarkan myosin, pada sarkoplasma otot polos terdapat kinase
ra tai ri ga iosi → aktivitas terga tu g kalsiu . Aktivitas C2+ dari kinase rantai ringan
miosin berikatan dengan kalmadulin. Ada 4Ca2+ ke subunit kinasenya (BM 105.000) kinase
rantai ringan akan diaktifkan oleh kalmadulin 4Ca2+ menfosforilasi rantai ringan-p yang
menghambat interaksi aktin-F miosin pada siklus kontraksi dimulai.
Relaksasi Otot Polos 1, Ca2+ sarkoplasma <10-7 mol/L menyebabkan Ca2+ berdisosiasi dari
baru pada rantai ringan-p terjadi fosfatesi rantai ringan protein aktif terus tanpa kalsium yang menyingkirkan fosfat dari rantai ringan-p. Terhambatnya pengikatan kepala miosin ke aktin dan aktifitas ATPase . Kepala miosin terlepas dari aktin-F dengan adanya ATP , tetapi tidak dapat terikat kembali karena rantai ringan-p defosforilasi terjadi relaksasi.
Relaksasi Otot Polos 2, protein kinase diaktifkan oleh cAMP menyebabkan mengfosforilasi kinase rantai ringan miosin terjadi afinitas kalmadulin C2+ menurun. cAMP
meningkat serta mengurangi respon kontraksi otot polos terhadap pengikatan C2+
sarkoplasma. Inhibitor fenotiazin dan obat antipsikotik akan terikat di kalmadulin dapat mencegah pengikatan kalmadulin pada enzim-enzim yang memerlukan kalsium.
Gambar 7.9 Peranan kalmadulin dalam fosforilasi protein otot
Fosforilasi protein Otot, fosforilasi rantai ringan miosin otot polos dapat mengurangi hambatan interaksi aktin-miosin, mulai siklus kontraksi, fosforilasi dibutuhkan untuk interaksi aktin-miosin otot polos. Fosfat rantai ringan miosin akan e e tuk helate de ga Ca2+
pada kompleks tropomiosin-TpC aktin menyebabkan peningkatan kecepatan pembentukan jembatan silang (cross bridges) miosin-aktin. TpI dan komponen peptida pompa C2+ retikulum
4. 2 molekul ADP yang dikatalisis enzim adenilil siklase
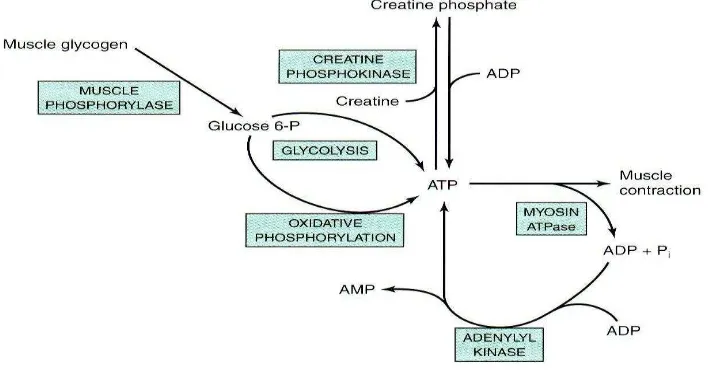
Cadangan ATP di otot tidak bertahan lama (1-2 detik), harus diperbaharui, enzim yang mengkatalisis fosforilasi kreatin yaitu CPK= kreatin fosfokinase, sarkoplasma otot rangka dalam tempat cadangan glikogen pada granula-granula dekat pita I. Kelainan otot spesifik (penyakit McArdle) akibatnya glikogen fosforilase inaktif dan penimbunan glikogen (glikogen storage disease).
Gambar 7.10 Sumber ATP dalam Otot
Fosforilasi oksidatif/ aerob, kontraksi otot misalnya sikap mempertahankan badan, menyimpan oksigen dalam mioglobin, di mana hem / tempat oksigen terikat disebut mioglobin pada otot berwarna merah. Mioadenilat kinase merupakan enzim otot mengkatalisis pembentuk molekul 1 ATP, 2 AMP dan 2 ADP yang digabung dengan hidrolisis ATP oleh ATPase miosin selama kontraksi otot.
Degradasi protein aktin-miosin, protein otot rangka sebagai sumber utama energi cadangan (bukan lemak) yang berfungsi degradasi asam amino hasil penelitian berupa aktin dan miosin dimetilisasi dengan ikatan peptide, metilhistidin (3Methis) dan diekskresikan di dalam urin akibat dari pemecahan protein. Otot mengoksidasi leusin menjadi CO2 dan rangka karbon dari aspartat, asparagin, glutamat, isoleusin dan valin menjadi asam trikarboksilat dan didegradasi otot meningkat 3-5 kaki selama puasa dan DM
Degradasi rangka Karbon aktin-miosin, rangka karbon asam-asam amino yang didegradasi dalam otot masuk dalam siklus asam sitrat , yaitu glutamin dan piruvat membentuk laktat. Keadaan puasa pada otot membebaskan semua asam-asam amino yang berasal dari pemecahan protein kecuali isoleusin, valin, glutamat, aspartat, asparagin akan digunakan untuk sintesis glutamin untuk jaringan lain serta otot membebaskan amoniak.
Gambar 7.11 Siklus Alanin