Diseminasi dan Mutasi Gen ENV GP70 Simian retrovirus Serotipe-2
(SRV-2) pada Jaringan Macaca fascicularis
Dissemination and Mutation of Simian retrovirus
serotype-2 (SRV-2) env gp70 Gene in Macaca fascicularis Tissues
Kristiana V1, Saepuloh U2*, Iskandriati D1,2, Pamungkas J2,3
1 Fakultas Teknobiologi, Universitas Katolik Indonesia Atma Jaya 2 Laboratorium Mikrobiologi dan Imunologi, Pusat Studi Satwa Primata, Lembaga Penelitian dan Pengabdian kepada Masyarakat, Institut Pertanian Bogor
3Fakultas Kedokteran Hewan, Institut Pertanian Bogor *Korespondensi: [email protected]
Abstract. Simian type D retroviruses (SRVs) cause simian acquired immune deficiency syndrome (SAIDS) in macaques with similar clinical signs to human AIDS. Thus, SRV can be used as a model of retrovirus infection in human. In this study, it was reported that tissues dissemination of the SRV serotype-2 naturally infect Macaca fascicularis from Indonesia. The presence of virus was detected from several tissues by polymerase chain reaction with specific primers that partially recognized env gp70 of SRV-2. Proviral SRV-2 was identified in peripheral blood mononuclear cell (PBMCs) and tissues, including lymphoid and non-lymphoid tissues. This showed broad distribution of SRV-2 in naturally infected macaques. The sequences of env gp70 were then aligned to determine variation among isolates that had been distributed in tissues. Nucleotide sequences alignment of env gp70 among isolates indicated high homology at the identities of 97.1 - 99.2% compared to PBMCs positive control (Mf ET1006). All sequences had close genetic relationship as indicated by the same cluster with SRV-2 reference sequences.
Key words : dissemination, env gp70 gene , Macaca fascicularis, SRV-2 Pendahuluan
Simian retrovirus Tipe D (SRV) adalah Betaretrovirus yang banyak ditemukan menginfeksi satwa primata genus Macaca. SRV merupakan patogen yang fatal bagi primata dan dilaporkan menyebabkan infeksi endemik di beberapa pusat penelitian primata di Amerika Serikat pada sekitar tahun 1980-an. Hingga kini, SRV telah ditemukan sebanyak tujuh serotipe.
SRV seringkali dikaitkan sebagai penyebab SAIDS (Simian Acquired Immuno Deficiency Syndrome), suatu sindrom penurunan sistem kekebalan tubuh atau imunodefisiensi pada simian (Marx et al. 1985, Gardner et al. 19888). SAIDS memiliki gejala klinis yang mirip dengan AIDS pada manusia (Daniel et al. 1984; Lerche et al.1995). Beberapa kesamaan gejala klinis ini menyebabkan SRV digunakan sebagai model untuk melihat aspek patogenitas retrovirus pada manusia.
Salah satu aspek patogenitas yang pen-ting adalah diseminasi virus dalam jaringan tu-buh. Beberapa jenis virus dapat menyebabkan infeksi yang terlokalisasi pada organ tertentu,
sementara beberapa lainnya dapat menyebab-kan infeksi yang tergeneralisasi (Guzman et al. 1999; Chopra & Mason, 1970). Untuk melihat diseminasi virus, pengamatan langsung pada berbagai organ manusia tidak memungkinkan untuk dilakukan. Penggunaan satwa primata, terutama genus Macaca, menjadi alternatif yang baik, karena kemiripan anatomi, fungsi fi-siologis, dan genetika dengan manusia.
Studi mengenai diseminasi SRV pada jaringan Macaca merupakan salah satu model untuk melihat penyebaran retrovirus pada manusia (Lackner et al. 1988). Pada studi yang dilakukan Wilkinson et al. (2003), sampel darah M. fascicularis yang berasal dari koloni endemik terinfeksi SRV-2 sengaja diinokulasikan melalui pembuluh darah (intravena) ke M. fascicularis yang bebas infeksi SRV untuk melihat penyebaran virus tersebut dalam jaringan. Hasilnya memperlihatkan adanya variasi penyebaran SRV ke beberapa jaringan tubuh, baik karena perbedaan respon antibodi tubuh maupun kemampuan transmisi virus. Pada penelitian ini, dilakukan analisis diseminasi SRV-2 pada M. fascicularis yang terinfeksi secara alami di habitat aslinya (Indonesia).
Dalam penyebarannya pada jaringan tubuh, SRV perlu mengenali reseptor pada sel inang untuk dapat masuk ke dalam sel dan melakukan proses replikasi. Bagian virus yang berfungsi untuk mengenali reseptor pada sel inang adalah selubung (envelope) virus. Selubung virus ini disandikan gen env pada genom SRV. Selubung virus menjadi faktor yang penting dalam tropis virus pada jaringan. Potensi patogenik virus salah satunya dipengaruhi variasi sekuen gen env (Philipp-Staheli et al. 2006). Data mengenai pengaruh diseminasi SRV terhadap terjadinya mutasi pada gen env belum tersedia. Oleh karena itu, analisis sekuen pun dilakukan pada gen env SRV yang telah terdistribusi pada jaringan.
Penelitian ini dilakukan untuk mengetahui diseminasi SRV-2 pada beberapa jaringan M. fascicularis asal Indonesia yang terinfeksi secara alami dengan mendeteksi keberadaan gen env gp70 SRV-2 menggunakan teknik PCR. Selain itu, ingin diketahui pula kemungkinan terjadinya mutasi pada gen env gp70 dari SRV-2 yang telah terdistribusi pada jaringan M. fascicularis dengan membandingkan sekuen gen env gp70 SRV-2 tiap jaringan yang terinfeksi dengan sekuen kontrol.
Materi dan Metode Sampel jaringan
Sampel yang digunakan adalah sampel jaringan (ginjal, hati, paru-paru, timus, yeyunum, kelenjar getah bening, limpa, otak, jantung) dan sel darah tepi berinti tunggal (peripheral blood mononuclear cells/PBMCs) M. fascicularis yang telah dibuktikan positif terinfeksi SRV-2 secara alami dengan uji PCR (data sekunder). Sampel merupakan koleksi Pusat Studi Satwa Primata, Lembaga Penelitian dan Pengabdian kepada Masyarakat-Institut Pertanian Bogor (PSSP, LPPM-IPB).
Ekstraksi DNA
Ekstraksi DNA dilakukan dengan menggunakan QIAamp DNA Blood Mini Kit (Qiagen) dengan mengikuti cara kerja yang diberikan dalam manual.
Amplifikasi DNA dengan PCR
Daerah target yang akan diamplifikasi adalah sebagian daerah env gp70 dari SRV-2. Primer yang digunakan adalah SRV-1/3 5737U21 (5’-CCAGATGGCTACCAGAACGAY-3’)
dan SRV-2 6243L22
(5’-CAAAACTGTGGGGACATTGCTA-3’) (Iskandriati et al. 2010). Master mix yang digunakan GoTaq Green Master Mix (Promega).
Kondisi PCR diatur dengan tahapan awal berupa pre-denaturasi pada suhu 94oC selama 10 menit. Tahap selanjutnya, adalah tahap amplifikasi dengan 40 siklus pengulangan, meliputi denaturasi pada suhu 94oC selama 30 detik, penempelan primer pada suhu 62oC selama 30 detik, dan ekstensi pada suhu 72oC selama 30 detik. Tahapan terakhir, pasca ekstensi pada suhu 72oC selama 7 menit dan dilanjutkan dengan penyimpanan pada suhu 4oC.
Elektroforesis dan visualisasi DNA
Hasil PCR lalu divisualisasi dengan elektroforesis horisontal (100 Volt, 45 menit) pada gel agarosa 1,5% dengan penambahan 10 µL etidium bromida (10 mg/ml) dan diamati pada UV-transiluminator (GelDoc, Bio-Rad). Penanda DNA yang digunakan 100 bp DNA ladder (Invitrogen). Hasil PCR yang positif ditunjukkan adanya pita fragmen DNA yang berukuran 506 pasang basa.
Purifikasi DNA gel agarosa
Pita DNA positif yang dihasilkan
dipurifikasi gel agarosa menggunakan QIAquick Gel Extraction Kit (Qiagen) dengan mengikuti cara kerja yang diberikan dalam manual.
Desekuenan (Perunutan)
DNA yang telah dipurifikasi dari gel agarosa, kemudian dirunut dengan menggunakan primer SRV-1/3 5737U. Sampel dikirim untuk disekuen di P.T. Charoen Phokpand Indonesia.
Analisis bioinformatika
Runutan yang diperoleh diolah lebih lanjut dengan menggunakan program BioEdit (Ibis Biosciences). Runutan tiap sampel lalu disejajarkan menggunakan program ClustalW (European Bioinformatics Institute/EBI) dengan kontrol runutan isolat SRV-2 asal M. fascicularis asli Indonesia (ET1006) (Iskandriati et al. 2010) dan runutan SRV-2 galur D2/MNE/ WA yang dikulturkan pada sel A549 (A549/ SRV2). Penyejajaran juga dilakukan dengan beberapa runutan pembanding dari basis data GenBank, yaitu SRV-2 galur D2/RHE/OR/ V1 (AF126468.1), SRV-2 galur D2/RHE/ OR (AF126467.1), SRV-2 complete genome (M16605.1), SRV-1 complete genome (M11841), SRV-3/Mason Pfizer Monkey Virus, MPMV (AF033815.1), 4 (NC_014474.1), SRV-5 (AB611707.1) dan SRV-8 (NC_031326.1). Hasil penyejajaran digunakan untuk membuat pohon filogenetika dengan program MEGA 6.0. Pohon filogenetika dibangun menggunakan metode neighbor-joining dengan uji bootstrap pada 1000 kali pengulangan.
Hasil dan Pembahasan
Amplifikasi dengan teknik PCR dilakukan untuk mengidentifikasikan keberadaan DNA provirus SRV-2 pada sampel yang dianalisis. Hasil positif ditunjukkan adanya pita berukuran sekitar 506 pasang basa. Hasil amplifikasi menunjukkan terdapat tujuh sampel jaringan yang positif, yaitu paru-paru, yeyunum, kelenjar getah bening, limpa, otak, jantung, dan PBMCs (Gambar 1). Ini menunjukkan infeksi SRV-2 secara alami dapat berdiseminasi hingga ke jaringan-jaringan tersebut.
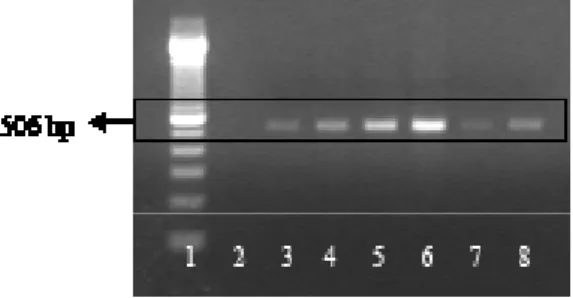
Purifikasi fragmen DNA dilakukan pada hasil PCR yang positif, kecuali hasil PCR sampel jaringan otak, karena hasilnya terlalu sedikit. Hasil purifikasi, seperti yang dapat dilihat pada Gambar 2, menunjukkan pita dengan variasi ketebalan pita yang mirip dengan hasil PCR. Ini menunjukkan keberhasilan proses purifikasi fragmen DNA yang dilakukan.
Diseminasi virus pada jaringan berhasil dilihat dengan mendeteksi keberadaan gen env gp70 SRV-2 pada sampel jaringan menggunakan teknik PCR. Ketebalan pita yang menunjukkan proviral SRV-2 paling besar teridentifikasi pada PBMCs dan jaringan limfoid lain seperti kelenjar getah bening dan limpa. Hal ini diperkirakan karena jalur penyebaran virus dalam tubuh melalui darah, disamping sel target dari retrovirus adalah sel-sel kekebalan tubuh. Akibatnya gejala utama infeksi retrovirus adalah penurunan sistem kekebalan tubuh hewan yang terinfeksi. SRV-2 juga menunjukkan diseminasi yang cukup luas ke beberapa organ non-limfoid, misalnya yeyunum dan paru-paru. Fenomena ini mendukung adanya infeksi oportunistik SAIDS yaitu diare yang persisten dan radang paru-paru (pneumonia). Diseminasi yang meluas ke beberapa beberapa jaringan ini dapat disebabkan rendahnya respon antibodi tubuh
yang dapat menetralisir virus pada M. fascicularis tersebut (Wilkinson et al. 2003).
Keenam fragmen DNA hasil purifikasi kemudian dirunut dengan menggunakan primer SRV-1/3 5737U. Runutan gen env SRV-2 dari enam jaringan kemudian disejajarkan secara bertingkat (multiple alignment) dengan runutan pembanding. Penyejajaran yang dilakukan adalah penyejajaran nukleotida gen env gp70 antar isolat asal jaringan dengan runutan kontrol ET1006 dan runutan A549/SRV-2.
Dari hasil penyejajaran yang dilakukan, dilihat persentase kemiripan antar sekuen isolat asal jaringan dengan isolat PBMCs asal Mf ET1006 sebgai kontrol positif, seperti yang dapat dilihat pada Tabel 1. Pada tabel tersebut dapat dilihat bahwa kemiripan (homology) antar sekuen isolat jaringan tersebut menunjukkan persen identitas nukleotida antara 97,4% sampai dengan 99,2%.
Penyejajaran nukleotida juga dilakukan antar isolat asal jaringan dengan isolat retrovirus referensi baik antar SRV-2 maupun SRV-1, SRV-3, SRV-4 dan SRV-5. Hasil penyejajaran kemudian digunakan untuk membangun pohon filogenetika. Pohon filogenetika yang dibentuk berdasarkan hasil penyejajaran nukleotida gen env gp70 antar isolat asal jaringan dengan sekuen kontrol ET1006 dan A549/SRV-2 maupun SRV 1-5 asal Genbank dapat dilihat pada Gambar 3.
Analisis penyejajaran nukleotida gen env gp70 antar sekuen isolat asal jaringan yang dibandingkan dengan isolat kontrol (PBMCs asal ET1006) menunjukkan kesamaan identitas nukleotida pada kisaran antar 96,0% sampai dengan 99,1%. Persentase kemiripan tertinggi adalah pada isolat PBMCs yang mencapai 99,1%, paru-paru 98,3% dan jejunum 97,6%; selanjutnya jaringan limpa 96,9% dan cairan getah bening sebesar 96,5%. Dari gambaran
Gambar 1 Hasil visualisasi PCR gen, env gp70 SRV-2, dari sampel beberapa jaringan asal M. fascicularis yang terinfeksi SRV-2. Marker 100 bp ladder (Invitrogen) (1), ginjal (3), paru-paru (4), hati (5), timus (6), yeyunum (7), kelenjar getah bening (8), limpa (9), otak (10), jantung (11), PBMCs (12), kontrol SRV-2 (13)
Gambar 2 Hasil purifikasi fragmen gen, env gp70 SRV-2. Marker 100 bp ladder (1), ginjal (3), paru-paru (3), yeyunum (4), kelenjar getah bening (5), limpa (6), jantung (7), PBMCs (8)
Tabel 1 Persentase kemiripan hasil penyejajaran runutan nukleotida di antara keenam isolat asal jaringan dengan kontrol isolat SRV asal Indonesia (ET1006) dan dibandingkan dengan SRV-2 referensi Genbank
Gambar 3 Pohon filo-genetik hasil penyejajaran nukleotida gen env gp70 SRV-2 isolat asal jaringan dengan isolat SRV-2 asal referensi dan SRV-1, 3, 4, dan 5
data tersebut dapat dilihat bahwa persentase identitas homologi pada jaringan non-limfoid (PBMCs, yeyunum dan paru-paru) lebih besar dibandingkan dengan jaringan jaringan limfoid (limpa dan cairan getah bening). Ini berarti mutasi nukleotida virus SRV-2 pada jaringan non-limfoid lebih kecil dibandingkan dengan
mutasi pada jaringan asal limfoid, walaupun mutasi yang terjadi sangat kecil, sehingga masih berada dalam kluster yang sama dalam pohon filogenetik. Hal ini kemungkinan berhubungan dengan kemampuan virus untuk menghindar dari sistem kekebalan tubuh pada organ-organ limfoid melalui mekanisme mutasi.
1 2 3 4 5 6 7 8 9 10 1. Jantung - 95,7 96,4 95,0 95,5 97,8 96,0 94,8 95,5 92,0 2. Jejunum - 97,6 95,8 96,2 98,6 97,6 95,1 95,8 93,0 3. Paru-paru - 96,5 96,9 99,2 98,3 95,8 96,5 93,7 4. Kelenjar getah bening - 95,1 97,4 96,5 94,2 94,8 92,1 5. Limpa - 97,8 96,9 94,4 95,1 92,3 6. PBMCs - 99,1 96,7 97,4 94,6 7. ET1006 - 97,4 98,1 94,2 8. SRV2_AF126467 - 97,7 93,1 9. SRV2_AF126468 - 92,9 10. SRV2_M16605 -
Pada pohon filogenetika hasil penyejajaran nukleotida env gp70 isolat asal jaringan (Gambar 3), isolat dapat dilihat membentuk satu kluster bersama-sama dengan kontrol positif ET1006 dan SRV-2 referensi yaitu AF126467, AF126468, dan M16605. Variasi nukleotida env gp70 antar isolat jaringan sangat kecil dan memiliki kemiripan yang tinggi dengan isolat lapang ET1006. Hal ini diperkirakan bahwa proviral SRV-2 pada jaringan bersifat stabil dan hanya sedikit mengalami mutasi padahal daerah gen yang dideteksi adalah daerah envelop yang memiliki variabilitas cukup tinggi dibanding daerah gen lainnya (Philipp-Staheli et al. 2006). Adapun sekuen kontrol yang digunakan yaitu SRV-2 ET1006 yang merupakan isolat lapang asal Indonesia menunjukkan adanya kedekatan antara isolat SRV-2 asal Indonesia dengan isolat SRV-2 strain D2/CEL/OR, vD2/RHE/OR dan D2/RHE/ORVI. Hal ini memperkuat asumsi bahwa isolat ET1006 merupakan isolat lapang SRV-2 yang diisolasi di pusat penelitian primata Amerika Serikat (Iskandriati et al. 2010).
SRV-1 dan SRV-3 membentuk kelompok sendiri yang terpisah dengan kelompok SRV-2. Ini menunjukkan kekerabatan yang lebih dekat antara SRV-1 dengan SRV-3, dibandingkan dengan SRV-2. Demikian juga dengan SRV-4 yang memiliki kedekatan dengan SRV-8 dan membentuk kluster tersendiri terpisah dari kelompok SRV-2 (Zao et al. 2010; Zao et al. 2016). SRV-5 juga membentuk kluster yang terpisah dengan SRV-2 demikian juga terhadap SRV-1/SRV-3 dan SRV-4/SRV-8 (Takano et al. 2013).
Secara umum, gen env gp70 dari SRV-2 menunjukkan adanya sedikit variasi. Tingkat variasi yang rendah mengindikasikan genom SRV-2 yang cukup stabil. Hasil ini sesuai dengan penelitian sebelumnya yang menunjukkan adanya variasi pada gen env gp70 dengan tingkat variasi yang tidak terlalu signifikan (Rosenblum et al. 2000).
Simpulan
Diseminasi SRV-2 yang menginfeksi M. fascicularis secara alami berhasil diamati dengan mengamplifikasi daerah gen env gp70 SRV-2 asal beberapa jaringan. SRV-2 menunjukkan diseminasi yang cukup luas ke beberapa jaringan, yaitu jaringan paru-paru, yeynum, kelenjar getah bening, limpa, otak, jantung, dan PBMCs. Analisis bioinformatika menunjukkan adanya homologi yang tinggi pada gen env gp70 SRV-2 yang terdistribusi pada jaringan tersebut.
Untuk menggambarkan dengan jelas diseminasi virus dan pengaruhnya pada patogenitas virus, pengamatan diseminasi perlu dilakukan secara menyeluruh pada seluruh jaringan tubuh.
Ucapan Terimakasih
Penelitian ini didanai Pusat Studi Satwa Primata, Lembaga Penelitian dan Pengabdian kepada Masyarakat–Institut Pertanian Bogor (PSSP, LPPM-IPB).
Daftar Pustaka
Chopra HC, Mason MM. 1970. A new virus
in a spontaneous mammary tumor of a rhesus monkey. J Cancer Res 30:2081-2086.
Daniel MD, King NW, Letvin NL, Hunt RD, Seghal PK, Desrosiers RC. 1984. A new
type D retrovirus isolated from macaques with an immunodeficiency syndrome. J Science 223:602-605
Gardner MB, Luciw P, Lerche N, Marx P. 1988. Nonhuman primate retrovirus
isolates and AIDS. J Adv Vet Sci and Comp Med 32:171-190
Guzman RE, Kerlin RL, Zimmerman TE.
1999. Histologic lesions in cynomolgus
monkeys (Macaca fasccularis) naturally infected with simian retrovirus type D: Comparison of seropositive, virus-positive, and uninfected animals. J Toxicol Pathol 27(6):672-677.
Iskandriati D, Saepuloh U, Mariya S, Grant RF, Solihin DD, Sajuthi D, Pamungkas J. 2010. Isolation and characterization
of simian retrovirus type D from Macaca fascicularis and M. nemestrina in Indonesia. J Microbiol Indones 4(3): 132-136
Lackner AA, Rodriguez MH, Bush CE, Munn RJ, Kwang HS, Moore PF, Osborn KG, Marx PA, Gardner MB, Lowenstine LJ. 1988. Distribution of
a macaque immunosuppressive type D retrovirus in neural, lymphoid, and salivary tissues. J Virol 62(6): 2134-2142
Lerche NW, Heneine W, Kaplan JE, Spira T, Yee JL, Khabbaz RF. 1995. An
expanded search for human infection with simian type D retrovirus. AIDS Res Hum Retroviruses 11(4):527-529
Marx PA, Bryant ML, Osborn KG, Maul DH, Lerche NW, Lowenstine LJ, Kluge JD, Zaiss CP, Henrickson RV, Shiigi SM. 1985. Isolation of a new serotype
of simian acquired immune deficiency syndrome type D retrovirus from Celebes black macaques (Macaca nigra) with immune deficiency and retroperitoneal fibromastosis. J Virol 56(2):571-578.
Philipp-Staheli J, Marquardt T, Thouless ME, Bruce AG, Grant RF, Tsai CC, Rose TM. 2006. Genetic variability of
the envelope gene of type D simian retrovirus-2 (SRV-2) subtypes associated with SAIDS-related retroperitonial fibromatosis in different macaque species. J Virol 3:11.
Rosenblum LS, Weiss RA, McClure MO.
2000. Virus load and sequence variation
in simian retrovirus type 2 infection. J Virol 74(8):3449-3454.
Takano JI, Leon A, Kato M, Abe Y, Fujimoto K. 2013. Isolation and DNA
characterization of a simian retrovirus 5 from a Japanese monkey (Macaca fuscata). J Gen Virol 94: 955–959.
Wilkinson RC, Murrell CK, Guy R, Davis G, Hall JM, North DC, Rose NJ, Almond N. 2003. Persistence and dissemination of
simian retrovirus type 2 DNA in relation to viremia, seroresponse, and experimental transmissibility in Macaca fascicularis. J Virol 77 (20):10751-10759.
Zao CL, Armstrong K , Tomanek L, Cooke A, Berger R, Estep JS, Marx PA, Trask JS, Smith DG, Yee J, Lerche NW. 2010. The Complete Genome and
Genetic Characteristics of SRV-4 Isolated from Cynomolgus Monkeys (Macaca fascicularis. J Virol 405(2): 390–396.
Zao CL, Tomanek L,Cooke A, Berger R, Yang L, Xie C, Chen S, Shi C, Rong R.
2016. A novel simian retrovirus subtype
discovered in cynomolgus monkeys (Macaca fascicularis). J Gen Virol (manuscript).