1
Gambaran Histopatologi Tiroid dan Ekspresi Interleukin 1 (IL-1) pada Hewan Model Tikus (Rattus norvegicus) Autoimmune Thyroiditis Hasil Injeksi
Tiroglobulin Kambing (cTG)
Histopathology of Thyroid Gland and Expression of Interleukine 1 (IL-1) in Animal Model Rattus norvegicus Autoimmune Thyroiditis Injected by
Caprine Thyroglobulin (cTG)
Prima Santi[1], Agung Pramana Warih M.[2], Dyah A.Oktaviane A.P[1] [1]
Program Studi Pendidikan Dokter Hewan Program Kedokteran Hewan, [2]
Jurusan Biologi Fakultas Matematika dan Ilmu Pengetahuan Alam Universitas Brawijaya
[email protected] ABSTRAK
Autoimmune thyroiditis (AITD) merupakan salah satu penyakit autoimun yang terjadi pada organ spesifik yaitu kelenjar tiroid. Deteksi dini keadaan subklinis penyakit ini belum terjangkau sehingga kondisi pasien tidak segera dapat diketahui. Hewan model AITD yang dikenal dengan experimental autoimmune thyroiditis (EAT) perlu dibuat untuk menghasilkan penanda spesifik AITD untuk pembuatan detection kit. Optimasi dosis dalam pembuatan EAT perlu dilakukan untuk menghasilkan EAT yang mempresentasikan kejadian AITD pada hewan model secara signifikan. Tujuan penelitian ini adalah untuk mengetahui tingkat keparahan AITD dengan variasi dosis tiroglobulin kambing (cTG) yang berbeda pada tikus (Rattus norvegicus) model AITD berdasarkan gambaran histopatologi tiroid dan ekspresi IL-1. Hewan model AITD dibuat dengan cara diinjeksi cTG yang diemulsikan dengan CFA atau IFA (perbandingan 1:1) sebanyak 0,2 mL secara subkutan cervical pada hari ke-0, 14 dan 28 dengan dosis 100µg/µL dan 200µg/µL. Hasil penelitian menunjukkan bahwa injeksi cTG menyebabkan kerusakan struktur dan bentuk folikel serta kenaikan infiltrasi sel-sel mononuklear pada gambaran histopatologi tiroid kelompok tikus perlakuan secara signifikan. Injeksi cTG juga menunjukkan peningkatan ekspresi IL-1 secara signifkan pada kelompok tikus perlakuan (p<0.05). Peningkatan dosis cTG berbanding lurus terhadap peningkatan ekspresi IL-1 dan kerusakan jaringan tiroid pada tikus perlakuan. Perlakuan yang signifikan berbeda nyata meningkatkan kerusakan struktur dan bentuk folikel, infiltrasi sel-sel mononuklear dan ekspresi IL-1 yaitu kelompok tikus yang diinjeksi 200µg/µL cTG.
Kata kunci : Autoimmune thyroiditis, Experimental autoimmune thyroiditis, Tiroglobulin kambing (cTG), Histopatologi jaringan tiroid, Interleukin-1
ABSTRACT
Autoimmune thyroiditis (AITD) is an autoimmune disease specifically affecting thyroid gland. Detection kit of AITD patients was expensive so that the patient's condition was not immediately known. Animal models of AITD known as experimental autoimmune thyroiditis (EAT) should be made to produce a specific marker for AITD detection. Dose optimization in the manufacture of animal models is needed to produce the present EAT incidence in animal models of AITD significantly. The purpose of this study was to know the severity of AITD on EAT using caprine thyroglobulin (cTG), based thyroid histopathology of thyroid gland and expression of IL-1.The EAT induced by 100µg/µL and 200µg/µ cTG was injected subcutaneously cervical on day 0, 14 and 28. The results showed that the injection of cTG
2
significantly damage to the structure and form follicles, and increase infiltration of mononuclear cells on thyroid histopathology of treated group. cTG injection also showed increased expression of IL-1 are significantly in treated group (p <0:05). Increasing dose of cTG cause increasing expression of IL-1 and tissue damage in rat thyroid treatment. Groups of rat were injected cTG 200μg/μL significantly increase the structure and form of follicle damage, infiltration of mononuclear cells and expression of IL-1
Keywords: Autoimmune thyroiditis, Experimental autoimmune thyroiditis, Caprine thyroglobulin (cTG), Histopathology of thyroid gland,, Interleukine-1
PENDAHULUAN
Autoimmune thyroiditis (AITD) merupa-kan salah satu penyakit autoimun yang ter-jadi pada organ spesifik yaitu kelenjar tiroid. Penyakit ini biasanya berupa hashimoto thy-roiditis (hipotithy-roiditis) dan grave’s disease (hipertiroiditis) (Chistiakov dan Turakulov, 2003). Penyakit ini ditandai dengan infiltrasi limfosit dan autoreaktif terhadap tiroid seba-gai mekanisme respon imun (Quaratino, 2004). Infiltrasi sel limfosit pada penyakit ini memediasi kerusakan sel-sel pada tiroid sehingga pada gambaran histopatologi tiroid yang mengalami AITD menunjukkan adanya infiltrasi sel mononuklear, perubahan struk-tur dan bentuk jaringan tiroid (Chistiakov dan Turakulov, 2003; Quarantino, 2004). Respon imun juga disertai dengan adanya aktivitas sitokin proinflamasi antara lain IL-1, IL-6 dan IL-8 pada kelenjar tiroid (Qua-rantino, 2004; Chistiakov, 2005).
Penyakit autoimun termasuk AITD memi-liki angka prevalensi mencapai 5-10% pada populasi di dunia dengan jumlah tercatat terbanyak di negara Amerika Serikat dan Jepang lebih sering terjadi pada wanita dibandingkan pria dan yang paling umum terjadi ada usia 45-65 tahun (Akin, 2011; Norris, 2007; Vanderpump, 2009). Penyakit ini secara klinis mirip dengan canine lym-phocytic thyroiditis (CLT) pada anjing (Gos-selin, 1982). Beberapa ras anjing yang rentan terhadap AITD yaitu irish setter, golden retriever, cocker spaniel, shetland sheepdog, beagle, doberman, giant schnauzer, hova-wart, pointer, skye terrier dan boxer dengan prevalensi lebih banyak terjadi pada anjing betina usia 2-4 tahun (Dodds, 2001; Nach-reiner, et al., 2002; NachNach-reiner, et al., 2002).
Gangguan pada kelenjar tiroid menjadi sangat penting karena hormon tiroid memi-liki pengaruh pada reproduksi, pertumbuhan dan diferensiasi serta metabolisme tubuh (Norris, 2007). Deteksi dini untuk AITD subklinis saat ini belum terjangkau sehingga kondisi pasien tidak dapat segera didiagnosa (Akin, 2011). Pengembangan detection kit dan pola terapi yang tepat mulai banyak dila-kukan.
Hewan model AITD disebut dengan experimental autoimmune thyroiditis (EAT) yang dapat dibuat metode induksi non-self-antigen atau self-non-self-antigen. Self-non-self-antigen yang dapat digunakan sebagai inducer EAT yaitu tiroglobulin (TG), thyroid stimulating hor-mone receptor (TSH-R) dan thyroid pero-xidase (TPO) (Weetman, 2004). Jumlah TG mencapai 75% pada jaringan tiroid lebih berpeluang menjadi autoantigen dibanding-kan TPO dan TSHR, selain TG merupadibanding-kan protein spesifik organ tiroid yang secara signifikan dapat menginduksi AITD pada EAT (Zhongtian, et al., 2004, Zhou dan Gill, 2005; Xiao-hong, et al., 2011). Tingkat keparahan yang dihasilkan TG mampu menginfiltrasi sel imun seluler spesifik pada jaringan tiroid yaitu sel TCD4+, sel TCD8+ dan sel B secara maksimal dibandingkan TPO dan TSHR (Ng, et al. 2004).
Hewan model AITD dengan menggunakan TG yang sudah dikembangkan dari tiro-globulin babi (pTG), tirotiro-globulin tikus (mTG), tiroglobulin sapi (bTG), tiroglobulin rodent (rTG), dan tiroglobulin manusia (hTG) namun hasil EAT berbasis TG sebe-lumnya kurang mampu menyebabkan infiltrasi sel mononuklear secara merata
3 (Karras, et al. 2005; Zhou dan Gill, 2005; Arata, et al., 2006; Xiao-hong, et al., 2011) Tiroglobulin dari berbagai spesies-spesies dapat digunakan dalam pengembangan EAT karena memiliki kesamaan fisik, biokimia dan struktur molekul sehingga penelitian ini menggunakan tiroglobulin kambing (cTG) dengan harapan dapat mempresentasekan ke-jadian AITD pada EAT secara signifikan
(Zhou dan Gill, 2005; Karras, et al. 2005; Arata, et al. 2006).
Penelitian ini bertujuan untuk mengetahui terjadinya reaksi inflamasi dan tingkat ke-parahan AITD pada tiroid tikus (Rattus norvegicus) model AITD yang diinjeksi va-riasi dosis cTG berdasarkan ekspresi IL-1 dan gambaran histopatologi tiroid.
MATERI DAN METODE Alat dan Bahan
Bahan yang digunakan adalah tikus putih (Rattus norvegicus) betina strain Wistar, tiro-globulin kambing (cTG), complete freud’s adjuvant (F5881 - Sigma Aldrich - USA), incomplete freud’s adjuvant (F5506-Sigma Aldrich-USA), Phospate Buffer Saline (PBS), PBS Asida, NaCl fisiologi, Tris-HCl, perekasi biuret, Phosphate Buffer Saline Tween-Phenyl Metane Sulfonyl Fluoride (PBST-PMSF), pasir kuarsa, etanol ber-tingkat (70%, 80%, 90%, 100%), parafor-maldehyde (PFA) 10%, xylol, alkohol, parafin cair, peroxidase block-H2O2 (K0679-Dako-USA), bovine serum albumin-BSA, mouse anti IL-1β (F162-Bioworld Technology-China), rabbit anti-mouse IgG-biotin link (K0679-Dako-USA), strep avidin horse radish peroxidase-SAHRP Dako-USA), DAB+ cromagen (K0679-Dako-USA), Mayer's Hematoxylin (S3309-Dako-USA) Eosin (CS701-DAKO-USA), entellan, dan aquadest.
Peralatan yang digunakan antara lain disetting set, blue and yellow tips, mikropi-pet, mortar dan pestel, spuit, sentrifugator, spektofotometer, vorteks, sonikator, inku-bator, magnetic stirrer, microtube, labu ukur, object glass, cover glass, centrifuge tube, autoclave, neraca analitik, disposable sy-ringe, oven, refrigerator dan mikroskop ca-haya.
Perlakuan Hewan Coba
Tikus (Rattus norvegicus) dibagi dalam 3 kelompok perlakuan masing-masing terdiri dari 6 ekor tikus yaitu kelompok normal, kelompok perlakuan 1 (dosis 100µg/µL) dan
kelompok perlakuan 2 (dosis 200µg/µL). Penggunaan hewan coba dalam penelitian ini telah mendapatkan sertifikat laik etik oleh Komisi Etik Penelitian Universitas Brawi-jaya No:140-KEP-UB
Isolasi dan Perhitungan Kadar cTG
Organ tiroid kambing dicuci dengan PBS sebanyak 3x. Organ sebanyak 1 gram dipotong-potong kecil kemudian diletakkan pada mortar dingin dan ditambahkan 1 mL PBST-PMSF serta pasir kuarsa secukupnya. Organ digerus dengan pestel dan diratakan. Homogenat ditambahkan 4 mL PBST-PMST kemudian dipindahkan ke dalam microtube dan digetarkan dengan vorteks selama 10 menit. Homogenat disonifikasi selama 10 menit lalu disentrifus (6.000 rpm selama 15 menit). Supernatan kemudian dipindahkan ke dalam microtube baru. Supernatan kemudian ditambahkan etanol absolut dingin (1:1) dan dibiarkan selama 24 jam dalam freezer. Su-pernatan setelah 24 jam disentrifus (10.000 rpm selama 15 menit). Etanol lalu dibuang dan sisa endapan dikeringkan. Endapan pro-tein ditambahkan buffer Tris-HCl dingin 20mM (1:1). Ekstrak protein cTG divorteks selama 5 menit dan disimpan pada freezer. Kadar cTG diukur menggunakan metode uji biuret dengan menggunakan spektrofoto-meter (540nm) (Amin, dkk, 2009).
Perhitungan Dosis dan Metode Injeksi (cTG) Dosis cTG kelompok perlakuan merupa-kan modifikasi yang didasarmerupa-kan pada Zhou dan Gill (2005) dan Xiao-hong, et al. (2011). Dosis cTG pada kelompok B yaitu 100 µg/µL cTG diemulsikan dengan CFA atau IFA (1:1). Pada kelompok C digunakan dosis
4 200 µg/µL cTG diemulsikan dengan CFA atau IFA (1:1). Volume injeksi cTG pada tikus sebanyak 0,2 mL/ekor.
Pembuatan Hewan Coba Model AITD Injeksi cTG
Injeksi dilakukan paada bagian subkutan cervical pada hari ke-0 dan booster pada hari ke-14 dan ke-28 pada kelompok perlakuan 1 dan 2. Kelompok perlakuan 1 diinjeksi 100 μg/µL cTG (dalam CFA 1:1 kemudian dilakukan booster dengan dosis 100 μg/µL cTG (dalam IFA 1:1) pada hari 14 dan ke-28. Kelompok perlakuan 2 diinjeksi 200 μg/µL cTG (dalam CFA 1:1) pada hari ke-0 dan booster dengan dosis 200 μg/µL cTG (dalam IFA 1:1) pada hari ke-14 dan ke-28. Pengambilan Organ Tiroid Tikus
Tikus dieutanasia dengan dislokasi leher kemudian dibedah dalam posisi ventrodorsal pada daerah cervical. Tiroid dipreparasi dari kedua sisi laring lalu dicuci dengan NaCl fisiologis 0.9% lalu dan dimasukkan pada paraformaldehide (PFA) 10%.
Pembuatan Preparat Histopatologi
Proses pembuatan preparat histopatologi terdiri dari fiksasi, dehidrasi dan infiltrasi, penjernihan, infiltrasi parafin, embedding, sectioning, penempelan di gelas objek (Amin, dkk, 2009).
Pewarnaan Preparat dengan Hematoxylin-Eosin
Preparat jaringan tiroid dimasukkan da-lam xylol (5 menit), lalu dimasukkan dada-lam alkohol absolut, alkohol 100%, 90%, 80% dan 70% (3 menit). Jaringan dicuci dengan aquades 1x dan dengan PBS pH 7,4 (3x5 menit). Jaringan diwarnai dengan Mayer’s Hematoxylin-Eosin (5 menit) dan dicuci aquadest (3x5menit). Preparat dikering anginkan, mounting dan ditutup dengan cover glass (Ramos-Vara, 2005).
Pewarnaan Preparat dengan Imunohisto-kimia - Interleukin 1 (IL-1)
Preparat dimasukkan xylol (5 menit) kemudian dalam alkohol absolut, alkohol
100%, 90%, 80% dan 70% (3 menit). Jaringan dicuci dengan aquades dan PBS pH 7,4 (3x5 menit). Jaringan lalu direndam dalam H2O2 (45 menit) lalu dicuci dengan PBS pH 7,4 (3x5 menit). Jaringan diblocking dengan BSA 1% (45 menit) lalu jaringan dicuci dengan PBS pH 7,4 (3x5 menit). Jaringan ditetesi antibodi primer mouse anti-IL-1 dan diinkubasi selama semalam pada suhu 4oC lalu dicuci dengan PBS pH 7,4 (3x5 menit). Jaringan ditetesi antibodi sekunder rabbit anti-mouse IgG berlabel biotin dan diinkubasi selama 1 jam. Jaringan lalu dicuci dengan PBS pH 7,4 (3x5 menit). Jaringan ditetesi SAHRP (45 menit). Jaringan lalu dicuci dengan PBS pH 7,4 (3x5 menit). Jaringan ditetesi DAB (7 menit). Preparat dicuci dengan aquades selama 3x5 menit. Preparat dicounterstaining dengan Mayer’s Hematoxylin selama 5 menit lalu dicuci dengan stilled water 3x5menit. Preparat kemudian dikeringanginkan dilan-jutkan dimounting dan ditutup dengan cover glass (Ramos-Vara, 2005).
Pengamatan Ekpresi Interleukin 1 (IL-1) Preparat histopatologi jaringan tiroid hasil pewarnaan imunohistokimia diamati meng-gunakan mikroskop cahaya Olympus BX51 perbesaran 40x-1000x sebanyak 5 lapang pandang. Perhitungan presentase area eks-presi IL-1 menggunakan gambaran histo-patologi jaring-an tiroid perbesaran 400x dan dianalisa dengan software Axio Vision. Perhitungan ekspresi IL-1 terhadap kontrol dikonversikan dalam persentase meng-gunakan rumus rata-rata persentase area kelompok perlakuan dikurangi dengan rata-rata persentase area kelompok kontrol dibagi dengan rata-rata kontrol dikalikan dengan 100% (Mandella, 2013).
Pengamatan Preparat Histopatologi
Preparat histopatologi jaringan tiroid diamati menggunakan mikroskop cahaya Olympus BX51 mulai perbesaran 40x-1000x sebanyak 5 lapang pandang untuk melihat adanya perubahan struktur dan bentuk folikel serta infiltrasi sel mononuklear. Gambaran histopatologi jaringan tiroid perbesaran
5 100x, 400x dan 1000x dianalisa secara deskriptif. Perhitungan presentase area infiltrasi sel mononuklear dilakukan pada gambaran histopatologi jaringan tiroid perbesaran 400x dengan software ImageJ. Infiltrasi sel mononuklear kemudian
diklasi-fikasikan berdasarkan indeks patologi tiroid AITD (Tabel 1). Perhitungan infiltrasi sel mononuklear terhadap kontrol dikonversikan dalam persentase menggunakan rumus seperti pada pengamatan ekspresi interleukin (IL-1).
Tabel 1. Indeks Patologi Tiroid AITD
Indeks Patologi Perubahan Histopatologi
0 Tidak ada infiltrasi, kelenjar yang normal menunjukkan folikel utuh dengan
lapisan sel epitel
1 Terjadi infiltrasi sel mononuklear dengan tingkat rendah, tidak jelas, sel
mononuklear didistribusikan perivaskular
2 Infiltrasi sel mononuklear terlihat dengan jelas, mempengaruhi 10 sampai
40% kerusakan pada jaringan
3 Infiltrasi melibatkan 40 sampai 80% jaringan
4 Infiltrasi melibatkan lebih dari 80% jaringan
Sumber. Karras et al., (2005)
Analisis Data
Data penelitian ini berupa data kualitatif dan data kuantitatif. Data kualitatif berupa pengamatan histopatologi jaringan tiroid yang disajikan dan analisis secara deskriptif. Data kuantitatif berupa data presentase area infiltrasi sel mononuklear dan presentase
area ekspresi IL-1 yang ditabulasi meng-gunakan Microsoft Office Excel kemudian dianalisa menggunakan analysis of variance (ANOVA) dengan software SPPS 16 for Windows. Apabila terdapat perbedaan nyata dilanjutkan uji Beda Nyata Jujur (Tukey) dengan α= 0.05%.
HASIL DAN PEMBAHASAN Ekspresi Interleukin 1 (IL-1)
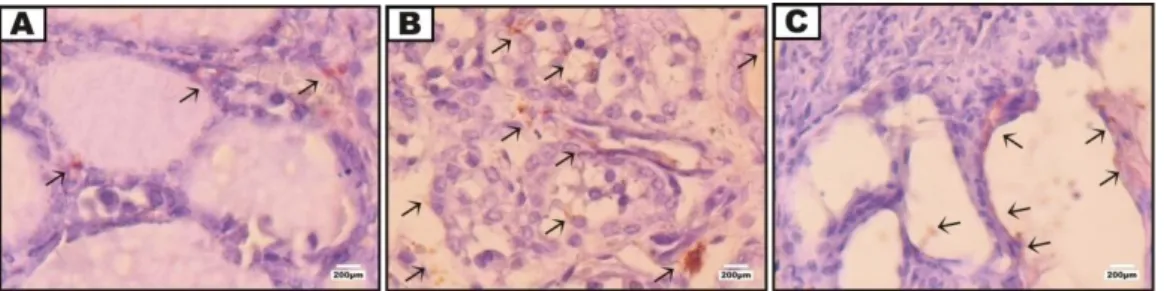
Hasil penelitian menunjukkan injeksi cTG meningkatkan ekspresi Interleukin 1 (IL-1) (Gambar 1). Rata-rata persentase area eks-presi IL-1 pada jaringan tiroid tikus per-lakuan mengalami peningkatan dibandingkan pada jaringan tiroid tikus normal (Tabel 2). Rata-rata persentase area ekspresi IL-1 pada tikus normal tanpa perlakuan adalah 0.354%. Ekspresi IL-1 meningkat secara signifikan pada tikus perlakuan. Tikus perlakuan 1 rata-rata persentase area ekspresi IL-1 mencapai 2.57% sedangkan pada tikus perlakuan 2 mencapai 8.708%. Rata-rata persentase area ekspresi IL-1 tikus perlakuan 1 dan 2 mengalami peningkatan masing-masing ber-turut 625% dan 2.359% dibandingkan pada jaringan tiroid tikus normal.
Interleukin 1 merupakan sitokin penting yang memediatori respon inflamasi dan
autoimun (Dinarello, 2005). Produsen utama dari IL-1 adalah monosit dan kadangkala makrofag, sel dendritik dan sel-sel lain (Limei, et al., 2008). Jaringan tiroid tikus normal menunjukkan adanya ekspresi IL-1 dengan intensitas rendah yaitu 0.345%, ekspresi tersebut dihasilkan oleh monosit pada bagian pembuluh darah pada jaringan tiroid untuk mengatur tirosit (sel-sel tiroid) (Gambar 5.1.A). Interleukin 1 memodulasi pertumbuhan tirosit untuk menjalankan fungsi memproduksi hormon-hormon tiroid serta memodulasi apoptosis sel-sel tirosit. Chistiakov (2005) bahwa keberadaan IL-1 pada jaringan tiroid normal berfungsi dalam menjaga pertumbuhan dan fungsi dari tirosit. Simons, et al. (1998) juga menyatakan bahwa IL-1 pada tiroid normal berfungsi menginduksi ekspresi Fas untuk memodulasi apoptosis tirosit.
6
Gambar 1. Ekspresi Interleukin 1 (IL-1) pada jaringan tiroid.
Keterangan: (A) jaringan tiroid tikus normal (tanpa perlakuan); (B) jaringan tiroid tikus perlakuan 1 (dosis 100µg/µL) dan (C) jaringan tiroid tikus perlakuan 2 (dosis 200µg/µL). Panah=ekspresi IL-1 (warna coklat muda). Perbesaran 1000x. Tabel 2. Rata-Rata Persentase Area Ekspresi Interleukin 1 (IL-1)
Kelompok Perlakuan Ekspresi IL-1 (%) (Mean ± SD)
Peningkatan Ekspresi IL-1 terhadap kontrol (%)
A (Normal tanpa perlakuan) 0.354 ± 0.1678 a 0
B (Perlakuan 1 dosis 100µg/µL) 2.57 ± 0.3860 b 625 C (Perlakuan 2 dosis 200µg/µL) 8.708 ± 0.2626 c 2.359
Keterangan : Perbedaan notasi (a,b,c) menunjukkan adanya perbedaan yang signifikan antar perlakuan terhadap ekspresi IL-1 (p<0.05)
Jaringan tiroid tikus perlakuan menun-jukkan peningkatan ekspresi IL-1. Interleukin 1 pada tikus perlakuan 1 dan 2 terlihat terekspresi disekitar sel-sel mono-nuklear. Sitokin proinflamasi ini memang diproduksi oleh sel-sel mononuklear pada jaringan tiroid akibat injeksi cTG sesuai dengan pernyataan Weetman (2003) bahwa IL-1 diproduksi oleh sel-sel mononuklear yang terinfiltrasi pada jaringan tiroid. Ekspresi IL-1 terlihat tersebar merata pada tikus perlakuan 1 karena infiltrasi sel mono-nuklear menyebar merata pada jaringan tiroid yang merupakan produsen sitokin tersebut. Interleukin 1 pada tikus perlakuan 2 terlihat berkumpul di jaringan tiroid yang tersisa disekitar tumpukan sel mononuklear (Gambar 1). Hal tersebut menyebabkan eks-presi IL-1 pada perlakuan 1 terlihat seakan-akan intensitasnya lebih tinggi dari perlakuan 2, namun rata-rata persentase ekspresi IL-1 pada jaringan tiroid perlakuan 2 lebih tinggi dibandingkan perlakuan 1 (Tabel 2).
Perbedaan intensitas ekspresi IL-1 pada perlakuan 2 dibandingkan perlakuan 1 kare-na dosis cTG pada perlakuan 2 lebih tinggi dari dosis cTG pada perlakuan 1. Perlakuan 2 yang injeksi cTG dengan dosis 200µg/µL
menyebabkan antigen yang masuk ke dalam tubuh tikus lebih banyak daripada perlakuan 1 dengan dosis 100µg/µL cTG. Jumlah antigen (cTG) yang lebih banyak akan mengakibatkan produksi antibodi (Ab-cTG) yang lebih banyak. Ikatan antigen-antibodi yang lebih banyak akan menyebabkan infil-trasi sel mononuklear pada jaringan tiroid juga lebih banyak. Infiltrasi sel mononuklear yang lebih banyak maka akan mengeks-presikan IL-1 dengan intensitas lebih tinggi dibanding pada perlakuan 1.
Ekspresi interleukin 1 pada jaringan tiroid perlakuan berfungsi untuk menginisiasi sel T CD4+, makrofag dan sel B merespon cTG seperti pernyataan Dinarello (2005) bahwa IL-1 berfungsi untuk menginisasi sel-sel imunokompeten untuk menyerang antigen. Ikatan Ab-cTG-TG memicu monosit (makrofag) pada jaringan tiroid mem-produksi IL-1 yang menginisiasi sel-sel mononuklear bermigrasi ke jaringan tiroid tikus perlakuan 1 dan 2. Migrasi sel-sel mononuklear pada jaringan tiroid diinisiasi oleh IL-1 sesuai dengan Weetman (2003) yang menyatakan bahwa ekspresi IL-1 menstimulus sel-sel mononuklear dan limfosit menginvasi jaringan tiroid. Simons,
7 et al. (1998) menyatakaan bahwa kenaikan ekspresi IL-1 menginduksi monocyte chemo-attractant protein-1 (MCP-1) dan granulocyte-macrophage colony-stimulating factor (GM-CSF) dalam pematangan dan infiltrasi monosit pada jaringan tiroid (dendritik sel) sehingga peningkatan ekspresi IL-1 memberikan respon terjadinya infiltrasi sel-sel mononuklear pada jaringan tiroid tikus perlakuan. Ekspresi IL-1 pada tikus perlakuan dengan intensitas tinggi akan menyebabkan mekanisme imunitas pada jaringan tiroid meningkat dan memberikan efek selanjutnya (Gambar 1). Peningkatan ekspresi IL-1 menunjukkan jaringan tiroid tikus perlakuan mengalami reaksi inflamasi
seperti yang dinyatakan Limei, et al. (2008) bahwa peningkatan ekspresi IL-1 terjadi secara signifikan pada penyakit autoimun seperti rheumatoid arthritis dan autoimmune thyroiditis.
Gambaran Histopatologi
Gambaran histopatologi jaringan tiroid hewan model tikus hasil injeksi cTG dapat diketahui melalui pewarnaan Hematoksilin-Eosin (HE). Pengamatan histopatologi ja-ringan tiroid meliputi perubahan struktur dan bentuk folikel serta infiltrasi sel-sel mono-nuklear. Histopatologi jaringan tiroid tikus perlakuan terlihat mengalami kerusakan (Gambar 2).
Gambar 2. Histopatologi pada jaringan tiroid (pewarnaan hematoksilin-eosin).
Keterangan: (A) jaringan tiroid tikus normal (a. folikel utuh, b.lumen utuh, c.sel epitel tersusun rapi); (B) jaringan tiroid tikus perlakuan 1 (dosis 100µg/µL) (a. bekas folikel terlihat, b. lumen tidak utuh, c dan panah. sel mononuklear); (C) jaringan tiroid tikus perlakuan 2 (dosis 200µg/µL) (c dan panah. sel mononuklear). Perbesaran 100x. Insert perbesaran 400x.
Hasil penelitian menunjukkan jaringan tiroid pada tikus normal terlihat folikel berbentuk bulat dengan struktur penyusun berupa sel epitel yang mengelilingi tertata rapi (Gambar 2.A). Jaringan tiroid pada tikus perlakuan yang diinjeksi cTG terlihat terjadi perubahan bentuk dan struktur dibandingkan jaringan tiroid normal (Gambar 2.B dan Gambar 2.C). Jaringan tiroid pada tikus perlakuan 1 terlihat bentuk folikelnya sudah tidak ter-lihat jelas pada sebagian besar jaringan tiroid. Struktur folikel pada tikus perlakuan 1 mengalami kerusakan dimana sel epitel yang mengelilinginya terlihat tidak tertata rapi seperti pada jaringan tiroid normal. Folikel pada jaringan tiroid terlihat tidak utuh. Lumen folikel dipenuhi oleh sel-sel mono-nuklear (Gambar 2.B). Jaringan tiroid tikus perlakuan 2 mengalami peningkatan kerusakan pada struktur maupun
bentuk folikel. Sel epitel yang menyusun folikel tidak ditemukan pada jaringan tiroid sehingga bentuk folikel tidak terlihat. Pada jaringan tiroid tikus perlakuan 2 ditemukan adanya infiltrasi sel-sel mononuklear. Sel-sel mononuklear tersebut menumpuk pada jaringan yang tersisa (Gambar 2.C).
Gambaran histopatologi pada jaringan tiroid tikus perlakuan hasil injeksi cTG memiliki kesamaan dengan hasil penelitian Zhou dan Gill (2005) menggunakan mTG, Xiao-hong, et al. (2011) menggunakan pTG, menunjukkan bahwa jaringan tiroid menga-lami kerusakan sehingga terlihat adanya perubahan struktur dan bentuk folikel.. Kerusakan pada jaringan tiroid tikus per-lakuan yang ditandai dengan perubahan struktur dan bentuk folikel disebabkan adanya aktivitas sel-sel mononuklear yang terinfiltrasi pada jaringan tiroid.
8 Jaringan tiroid pada tikus perlakuan selain terjadi perubahan struktur dan bentuk folikel juga terjadi infiltrasi sel-sel mononuklear (Gambar 3). Sel mononuklear tersebut terlihat memiliki satu buah inti berbentuk bulat dengan sitoplasma lebih besar dari inti seperti pada Junqueira (2007) bahwa sel mononuklear berbentuk bulat dengan satu inti berada ditengah. Gambaran histologi jaringan tiroid tikus normal tanpa perlakuan
tidak nampak adanya keberadaan sel-sel mononuklear (Gambar 3.A) hal tersebut sesuai dengan pernyataan Chistiakov (2005) dan Norris (2007) bahwa keberadaan sel mononuklear (monosit) pada tiroid tikus normal berada di dalam pembuluh darah dalam jaringan tiroid yang berfungsi mengekspresikan IL-1 untuk mengatur per-tumbuhan dan fungsi tirosit.
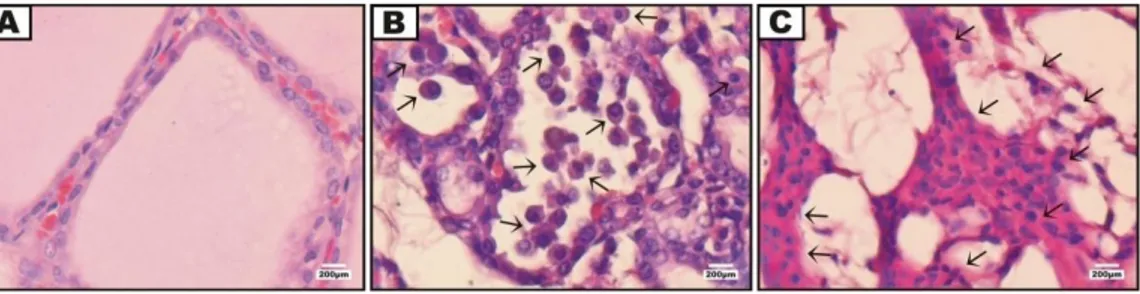
Gambar 3. Infiltrasi sel mononuklear pada jaringan tiroid (pewarnaan hematoksilin-eosin).
Keterangan: (A) jaringan tiroid tikus normal; (B) jaringan tiroid tikus perlakuan 1 (dosis 100µg/µL); (C) jaringan tiroid tikus perlakuan 2 (dosis 200µg/µL). Panah=sel mononuklear. Perbesaran 1000x.
Tabel 3. Rata-Rata Persentase Area Infiltrasi Sel Mononuklear
Kelompok Perlakuan Infiltrasi sel Mononuklear
(%) (Mean ± SD) Indeks Patologi Perubahan Histopatologi terhadap Kontrol (%)
A (Normal tanpa perlakuan) 0.06 ± 0.03 a 0 0
B (Perlakuan 1 dosis 100µg/µL) 12.87 ± 4.08 b 2 21219
C (Perlakuan 2 dosis 200µg/µL) 80.55 ± 5.67 c 4 133270
Keterangan : Perbedaan notasi (a,b,c) menunjukkan adanya perbedaan yang signifikan antar perlakuan terhadap infiltrasi sel mononuklear (p<0.05)
Sel-sel mononuklear terinfiltrasi pada jaringan tiroid tikus perlakuan 1 dan 2 (Gambar 3.B dan Gambar 3.C). Sel-sel mononuklear pada jaringan tiroid tikus perlakuan 1 terlihat jelas bentuknya dan mampu dibedakan dengan sel epitel. Dis-tribusi sel mononuklear pada jaringan tiroid perlakuan 1 menyebar merata pada lumen folikel. Terlihat jelas bahwa lumen-lumen folikel tidak lagi utuh dan berisi koloid namun digantikan oleh sel-sel mono-nuklear (Gambar 3.B). Histopatologi ja-ringan tiroid perlakuan 2 terlihat bahwa sel-sel mononuklear menumpuk pada jaringan tiroid yang tersisa. Sel-sel mononuklear pada jaringan tiroid tikus perlakuan 2 tidak dapat terlihat dengan jelas bentuknya. Pada gam-baran histopatologi tikus perlakuan 2
infiltrasi sel mononuklear terlihat menumpuk sehingga tidak dapat dipisahkan dengan sel epitel (Gambar 3.C).
Penelitian Zhou dan Gill (2005) yang diinjeksi mTG, Karras, et al. (2005) dan Arata, et al. (2006) yang diinjeksi hTg mau-pun Xiao-hong, et al. (2011) yang diinjeksi pTG menunjukkan bahwa sel-sel mono-nuklear terinfiltrasi pada jaringan tiroid per-lakuan. Infiltrasi sel-sel mononuklear meru-pakan akibat yang ditimbulkan dari ikatan antigen-antibodi pada jaringan tiroid dikare-nakan antibodi tidak mampu membedakan cTG dengan TG.
Pemberian perlakuan mengakibatkan pe-ningkatan rata-rata persentase area infiltrasi sel mononuklear (Tabel 3). Rata-rata per-sentase area infiltrasi sel mononuklear pada
9 tikus normal tanpa perlakuan adalah 0.06%. Rata-rata persentase area infiltrasi sel mono-nuklear meningkat secara signifikan pada kelompok tikus perlakuan. Infiltrasi pada tikus perlakuan 1 mencapai 12.87% sedang-kan pada tikus perlakuan 2 mencapai 80.55%. Peningkatan infiltrasi sel mono-nuklear pada tikus perlakuan 1 dan 2 berturut-turut mencapai 21219% dan 133270% dari jaringan tiroid normal.
Grading kerusakan tiroid AITD ber-dasarkan infiltrasi sel-sel mononuklear hasil penelitian menggunakan indeks patologi tiroid AITD Karras et al., (2005) (Tabel 1). Grading menunjukkan rata-rata persentase area infiltrasi sel-sel mononuklear masuk dalam grade 0, 2 dan 4 (Tabel 3). Jaringan tiroid normal merupakan grade 0 (tidak ada infiltrasi), jaringan tiroid perlakuan 1 meru-pakan grade 2 (sel mononuklear jelas dan mempengaruhi 10% sampai 40% kerusakan pada jaringan) dan jaringan tiroid perlakuan 2 merupakan grade 4 (infiltrasi melibatkan lebih dari 80% jaringan). Kerusakan jaringan tiroid penelitian ini mampu mengakibatkan infiltrasi lebih tinggi dari injeksi 0,8µg/µL mTG pada Zhou dan Gill (2005) yang hanya mampu menghasilkan infiltrasi sel mono-nuklear grade 0-0.5. Karras, et al. (2005) yang diinjeksi 200µg/µL hTg menunjukkan bahwa injeksi yang dilakukan hanya meng-hasilkan kerusakan jaringan tiroid dengan grade 0-0.5.
Injeksi cTG pada tikus dikenali sebagai antigen oleh APC. Pengenalan awal cTG pada APC akan mengakibatkan pengaktifan sel T yang kemudian akan berdiferensiasi menjadi sel Th1 dan Th2. Pembentukan sel Th2 meng-inisiasi sel B menghasilkan sel memori dan sel plasma yang memproduksi antibodi cTG (Ab-cTG). Antibodi cTG akan mengikat cTG dan menghancurkannya. Pelepasan antigen cTG ke dalam darah seca-ra bertahap akan mengakibatkan pengenalan ulang APC. Pengenalan ulang ini akan dires-pon oleh sel memori. Resdires-pon sel memori berupa inisiasi terhadap sel plasma akan mengakibatkan memproduksi Ab-cTG. Pro-ses ini akan terjadi berulang kali setiap kali antigen dilepas ke pembuluh darah.
Pembentukkan Ab-cTG terus diproduksi dan jumlahnya terus meningkat.
Antibodi (Ab-cTG) kemudian mengikat dan menghancurkan cTG. Antigen cTG memiliki kesamaan dengan TG pada tiroid tikus perlakuan. Kesamaan tersebut meng-akibatkan Ab-cTG mengenali TG pada tiroid tikus sebagai antigen. Pengenalan tersebut mengakibatkan terjadinya reaksi autoimun pada tiroid tikus. Antibodi Ab-cTG kemu-dian mengikat TG pada tiroid tikus per-lakuan. Ikatan tersebut menginisiasi sel mononuklear (monosit dan makrofag) meng-ekspresikan IL-1. Ekspresi IL-1 menginisiasi migrasi sel mononuklear dari pembuluh darah ke jaringan tiroid. Migrasi tersebut menyebabkan infiltrasi sel-sel mononuklear pada jaringan tiroid tikus. Infiltrasi sel-sel mononuklear mengakibatkan TG pada bagian lumen folikel maupun yang masih diproduksi pada sel epitel tiroid tikus difagosit dan dihancurkan. Mekanisme terse-but menyebabkan kerusakan pada jaringan tiroid tikus perlakuan (Gambar 3). Kerusakan dan infiltrasi sel mononuklear jaringan tiroid pada tikus perlakuan tersebut menunjukkan salah satu ciri dari penyakit AITD. Kerusakan dan infiltrasi sel mononuklear pada jaringan tiroid tikus pada EAT hasil injeksi tiroglobulin juga ditunjukkan pada Zhou dan Gill (2005); Karras, et al. (2005); Arata, et al., (2006) dan Xiao-hong, et al., (2011).
Perbedaan tingkat kerusakan dan infiltrasi sel mononuklear pada perlakuan 2 diban-dingkan perlakuan 1 dikarena dosis cTG pada perlakuan 2 lebih tinggi (dua kali) dari dosis cTG pada perlakuan 1. Perlakuan 2 yang injeksi cTG dengan dosis 200µg/µL menyebabkan antigen yang masuk ke dalam tubuh tikus lebih banyak daripada perlakuan 1 dengan dosis 100µg/µL cTG. Jumlah antigen (cTG) yang lebih banyak akan mengakibatkan produksi antibodi (Ab-cTG) yang lebih banyak. Ikatan antigen-antibodi yang lebih banyak akan menyebabkan in-filtrasi sel mononuklear pada jaringan tiroid juga lebih banyak. Infiltrasi sel mononuklear yang lebih banyak maka akan menyebabkan
10 kerusakan yang lebih tinggi dibanding pada perlakuan 1.
Variasi dosis injeksi cTG pada tikus perlakuan signifikan berbeda nyata dalam peningkatan kerusakan histopatologi tiroid
dan ekspresi IL-1. Peningkatan dosis cTG berbanding lurus dengan peningkatan keru-sakan histopatologi tiroid dan ekspresi IL-1 pada jaringan tiroid tikus perlakuan.
KESIMPULAN
Berdasarkan hasil penelitian yang telah dilakukan dapat diambil kesimpulan bahwa: 1. Injeksi variasi dosis cTG secara signifikan
meningkatkan ekspresi interleukin 1 (IL-1) pada tikus (Rattus norvegicus) AITD (p<0.05) yang merupakan penanda terjadinya reaksi inflamasi. Peningkatan dosis cTG berbanding lurus dengan ekspresi interleukin 1 (IL-1) pada tikus AITD.
2. Injeksi variasi dosis cTG mengakibatkan kerusakan secara signifikan (p<0.05) pada jaringan tiroid tikus (Rattus norvegicus) AITD yang ditandai dengan perubahan struktur dan bentuk folikel serta meningkatkan infiltrasi sel mononuklear yang merupakan parameter terjadinya reaksi autoimun. Peningkatan dosis cTG berbanding lurus dengan peningkatan kerusakan jaringan tiroid AITD. UCAPAN TERIMAKASIH
Peneliti mengucapkan terima kasih kepada analis dan staf Laboratorium Biokimia, Laboratorium Molekuler dan Laboratorium Fisiologi Hewan FMIPA Universitas Brawijaya serta Laboratorium
Patologi RS. Dr. Soetomo Surabaya yang telah membantu dalam penelitian ini dan kepada DIKTI yang telah mendanai sebagian dari penelitian ini melalui kegiatan PKM.
DAFTAR PUSTAKA
Akin, F. (ed.), 2011. Basic and Clinical Endocrinology Up-to-Date, InTech, Rijeka,
Amin, M.H.F, A.P.W. Marhendra dan
Aulanni’am. 2009. Pengaruh Paparan
Lipopolisakarida pada Rongga Mulut dan Assisted Drainage Therapy (Adt) terhadap Kadar S-Ige dan Profil Radikal Bebas Pada Tikus Asma, Paper Presentasi pada Seminar Nasional Biologi XX dan Kongres PBI XIV UIN Maliki, Malang, 24-25 Juli.
Arata, N., T. Ando, P.Unger and T. F. Davies. 2006. By-Stander Activation in Autoimmune
Thyroiditis: Studies on Experimental
Autoimmune Thyroiditis in The GFP+ Fluorescent Mouse. Clinical Immunology. 2006:121, pp. 108-117
Baratawidjaja, K.G. dan I. Rengganis. 2010. Imunologi Dasar. Jakarta. Balai Penerbit FKUI.
Chistiakov, D.A. and R.I. Turakulov. 2003. CTLA-4 and its Role in Autoimmune Thyroid Disease. J. Mol Endocrinol Vol 31 pp.21–36 Chistiakov, D.A. 2005. Immunogenetics of
Hashimoto's Thyroiditis. J. Autoimmune Diseases Vol.2 No.1
Dinarello, C.A. 2005. The IL-1 Family and Inflammatory Diseases. AMGEN-suppl. 27 17-08-2005 12:13 Page S-1
Dodds, W.J. 2001. Vaccination Protocols for Dogs Predisposed to Vaccine Reactions. J. Americ. Anim. Hosp. Assoc. Vol 37 No.3 pp.211-214
Gosselin, S.J, C.C. Capen, S.L. Martin and S. Krakowka. 1982. Autoimmune Lymphocytic Thyroiditis in Dogs. Vet. Immunol and Immunopathol Vol.3 pp.185-201
Junqueira, L.C. 2007. Histology Dasar: Teks dan Atlas Edisi 10. Jakarta : EGC.
Karras, E., H. Yang, P. Lymberi and P. Christadoss. 2005. Human Thyroglobulin
Peptide p2340 Induces Autoimmune
Thyroiditis in HLA-DR3 Transgenic Mice. Journal of Autoimmunity 24:2005, pp. 291-296
Limei L., Zhaoliang F., Jianke R., Ruilin S., Zhihui L., Zhejin S., Long W., Xia S., Jun Y., Zhugang W. and Jian F. 2008. Functional Imaging of Interleukin 1 Beta Expression in Inflammatory Process Using Bioluminescence
Imaging in Transgenic Mice. BMC
11
Mandella, N.I. 2013. Ekspresi Tumor Necrosis Factor (TNF-α) Dan Gambaran Histopatologi Sendi Tikus Artritis (Rattus norvegicus) yang Mendapatkan Terapi Ekstrak Buah Kesemek Junggo (Diospyros kaki L,f). [Skripsi]. Program Kedokteran Hewan. Universitas Brawijaya.
Nachreiner, R.F., K.R. Refsal, P.A. Graham and M.M. Bowman. 2002. Prevalence of Serum Thyroid Hormone Autoantibodies in Dogs with Clinical Signs of Hypothyroidism. J. AmericVet. Med. Assoc. (Abstr). Vol. 220, No. 4, pp.466-471
Ng, H.P., J.P. Banga and A.W. Kung. 2004. Development of A Murine Model Of
Autoimmune Thyroiditis Induced with
Homologous Mouse Thyroid Peroxidase, Endocrinology, vol.145, no.2, pp. 809–816 Norris, D.O. 2007. Vertebrate Endocrinology 4th
ed. Burlington. Elsevier Academic Press. Quaratino, S. 2004. Drug Discovery Today:
Disease Models | Autoimmune Diseases and Inflammation Vol. 1, No. 4
Ramos-Vara, J.A. 2005. Technical Aspects of
Immunohistochemistry. Veterinary
Pathology, vol.42, no.4, pp. 405–426.
Simons P.J., F.G. Delamarre and H.A. Drexhage:
Antigen-Presenting Dendritic Cells as
Regulators of The Growth of Thyrocytes: A
Role for Interleukin-1 Beta and Interleukin-6. Endocrinology 1998, 139:3148-3156.
Vanderpump, M.P.J. 2009. 'Epidemiology of Thyroid Dysfunction-Hypothyroidism and Hyperthyroidism' dalam Smyth, PPA (ed), Thyroid Internasional 2-2009. Merck KGaA, Darmstadt, pp.1-11
Weetman, A.P. 2004. Autoimmune Thyroid Disease. Autoimmunity, vol.37, no.4, pp. 337– 40.
Xiao-hong S., Ri-zeng Z., Chen-huan Y. and Fang W. 2011. Effects of Modified Haizao
Yuhu Decoction in Experimental
Autoimmune Thyroiditis Rats.
J.Ethnopharmacol, vol.135, no.2, pp. 321-324.
Zhongtian J., Kouki M., Keisei F., Saeko H., Jun-ichi T., Jo S. Sadayoshi I. Susumu S. and Katsumi Y. 2004. Experimental Autoimmune Thyroiditis in Nonobese Diabetic Mice Lacking Interferon Regulatory Factor-1. J. Clin Immunol Vol.113 pp. 187– 192
Zhou, J.S. and Gill, H.S. 2005.
Immunostimulatory Probiotic Lactobacillus rhamnosus HN001 and Bifidobacterium lactis
HN019 Do Not Induce Pathological
Inflammation in Mouse Model of
Experimental Autoimmune Thyroiditis. Intern J.Food Microbiol, vol.103, no.1, pp. 97–104.