KARAKTERISASI REPRODUKSI MUNCAK,
Muntiacus muntjak muntjak JANTAN: KAJIAN ANATOMI,
PROFIL METABOLIT TESTOSTERON, DAN SPERMATOGENESIS
SELAMA PERIODE PERTUMBUHAN RANGGAH
SRI WAHYUNI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2012
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi Karakterisasi Reproduksi Muncak, Muntiacus muntjak muntjak Jantan: Kajian Anatomi, Profil Metabolit Testosteron, dan Spermatogenesis selama Periode Pertumbuhan Ranggah adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Mei 2012
Sri Wahyuni NIM B362080011
SRI WAHYUNI. Reproductive Characterization of Male Muntjak, Muntiacus muntjak muntjak: Study of Anatomy, Testosterone Metabolite Profile, and Spermatogenesis during Antler Growth Periods. Under direction of TUTY L. YUSUF, SRIHADI AGUNGPRIYONO, and MUHAMMAD AGIL.
Muntjaks (Muntiacus muntjak muntjak) are belonging to Cervidae which distributed in Java Island and Southern part of Sumatera. Muntjaks have been protected by Indonesian Government since 1999. In order to support breeding program of the species and to avoid from extinction, several activities in this study were conducted. Therefore, the objective of this study was to investigate the reproductive capacity of male muntjaks in captivity. Anatomical procedure and histological preparation of reproductive organ (testis, ductus epididymidis, accessory sex glands, and penis) from an adult male muntjak (♂#1) in hard antler period was carried out to collect data of their morphology and histology structure. Immunoreactive testosterone (iT) level of fecal samples was measured by the enzymeimmunoassay method using testosterone assay (Möstl) from two adult male muntjaks with individual code as ♂#2 and ♂#3. Furthermore, core needle biopsy and electroejaculation methods were applied for obtained testicular tissues and semen for study spermatogenesis. The entire samples were collected in hard antler (RK), casting (C), and velvet antler (RV) periods with different sampling frequency. The result showed that in general, the anatomy of reproductive organ in male muntjak was somewhat similar with other small ruminant included Cervids (ram and goat, rusa deer, pampas deer, and reeves muntjak). As dominant male, ♂#2 had sturdier and bigger hard antlers (RK1 and RK2) compare to ♂#3. The duration of antler cycle of ♂#2 was also longer (459 days) than ♂#3 (381 days) in the first antler cycle. Duration of second antler cycle of ♂#3 was longer (20.65%) than in previous cycle (485 days) Based on the hormonal data, iT level of ♂#2 was different among RK, C, and RV periods where iT level during RK 1 period was higher than C (p = 0.003) and RV (p = 0.02). In
addition, iT level during RK 2 was also significantly higher than those C (p = 0.009) and RV (p = 0.06). In the first antler cycle, the pattern of T secretion
in ♂#3 was slightly different if compared to the ♂#2. ♂#3 showed iT level was significantly lower in RK1 than RK2 (p = 0.0003), RK3 (p= 0.0034), C1 (p = 0.05), and C2 (p = 0.0001). However, the pattern of testosterone secretion was changed when ♂#3 in RK2 at the second of antler cycle coincided with increase of ♂#3’s age and death of ♂#2. Interestingly, iT level still detected during C and RV periods in both of muntjaks and showed a positive correlation with spermatogenesis which was proven by presence of spermatozoa in semen with significant concentration during those periods. The highest sperm concentration, however, was found in RK, and slightly decreased in C and RV. These findings provide that the specific pattern of testosterone metabolite profile that exhibited by ♂#2 and ♂#3 had closed correlation with their age, body size, antler size, and antler cycles length. In conclusion, testosterone that synthesized during an antler cycle plays a central role in continuing reproductive activities in male muntjaks. Therefore, muntjaks could provide reproductive function throughout the year of reproductive aseasonality which is similar to the reeves and formosan muntjaks. Keywords: muntjak, testosterone metabolite profile, antler growth period,
SRI WAHYUNI. Karakterisasi Reproduksi Muncak, Muntiacus muntjak muntjak Jantan: Kajian Anatomi, Profil Metabolit Testosteron, dan Spermatogenesis selama Periode Pertumbuhan Ranggah. Dibimbing oleh TUTY L. YUSUF, SRIHADI AGUNGPRIYONO, dan MUHAMMAD AGIL.
Muncak (Muntiacus muntjak muntjak) termasuk ke dalam famili Cervidae yang tersebar di Pulau Jawa dan Pulau Sumatera bagian selatan. Sejak tahun 1999, muncak ditetapkan sebagai satwa yang dilindungi di Indonesia. Perburuan liar dan eksploitasi habitat muncak dan satwa lainnya di alam menjadi penyebab utama penurunan populasi muncak. Sampai saat ini belum ada data pasti mengenai populasi muncak di habitat asli maupun di area konservasi. Kekhawatiran akan punahnya muncak di Indonesia menjadi alasan kuat dilakukannya penelitian ini. Selain itu karakteristik reproduksi muncak Indonesia sejauh ini belum dilaporkan, sehingga upaya pengembangbiakan muncak di penangkaran tidak dapat berjalan dengan baik. Pada penelitian ini digunakan tiga ekor muncak jantan dewasa sesuai dengan Surat Keputusan Menteri Kehutanan Republik Indonesia nomor SK. 23/Menhut-II/2011. Muncak diperoleh dari hasil tangkapan masyarakat di Kabupaten Cilacap, Jawa Tengah.
Regulasi hormonal terhadap pertumbuhan ranggah berkaitan erat dengan aktivitas reproduksi selama satu siklus ranggah, meliputi: periode ranggah keras (RK), lepas ranggah atau casting (C), dan ranggah velvet (RV). Sejauh mana keterkaitan antara pertumbuhan ranggah dengan aktivitas steroidogenesis dan spermatogenesis untuk menghasilkan spermatozoa selama satu siklus ranggah pada muncak dapat diketahui dari penelitian ini. Oleh karena itu, tujuan dari penelitian ini adalah untuk mengetahui dan mempelajari karakteristik reproduksi muncak jantan selama periode pertumbuhan ranggah yang pada akhirnya dapat menjelaskan gambaran fertilitas muncak dan pola reproduksinya. Ada empat kegiatan penelitian yang dilakukan dengan metode spesifik, yaitu: 1) anatomi dan histologi organ reproduksi muncak jantan, 2) karakteristik pertumbuhan ranggah muncak, 3) profil metabolit testosteron selama periode pertumbuhan ranggah, dan 4) spermatogenesis dan kualitas semen muncak selama periode pertumbuhan ranggah. Hasil pengamatan selanjutnya dibahas pada bagian akhir dari penelitian ini. Data yang diperoleh dapat digunakan sebagai bahan pertimbangan dalam manajemen pemeliharaan dan pengembangbiakan muncak di penangkaran.
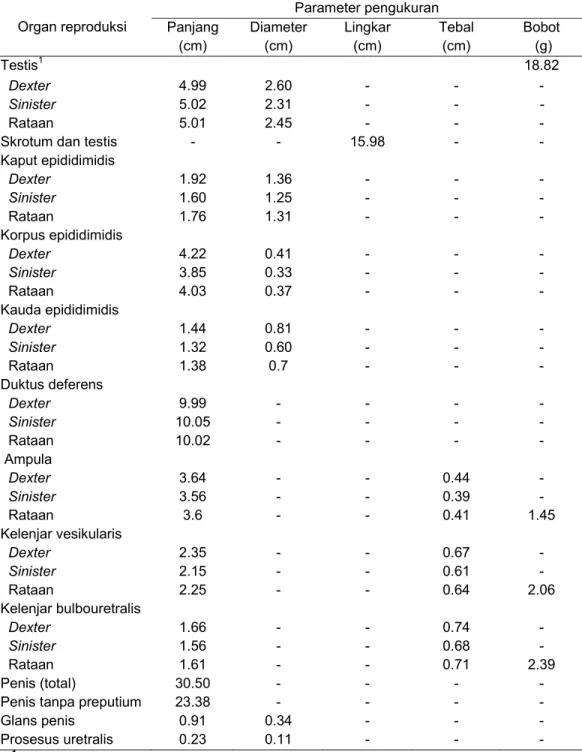
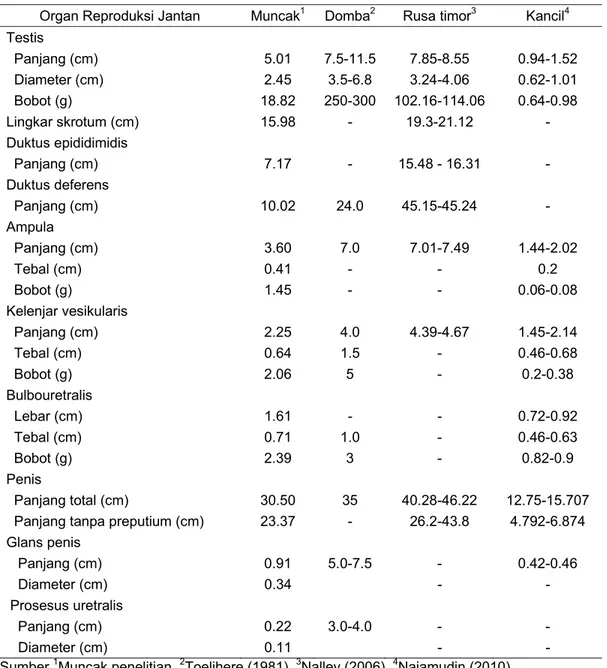
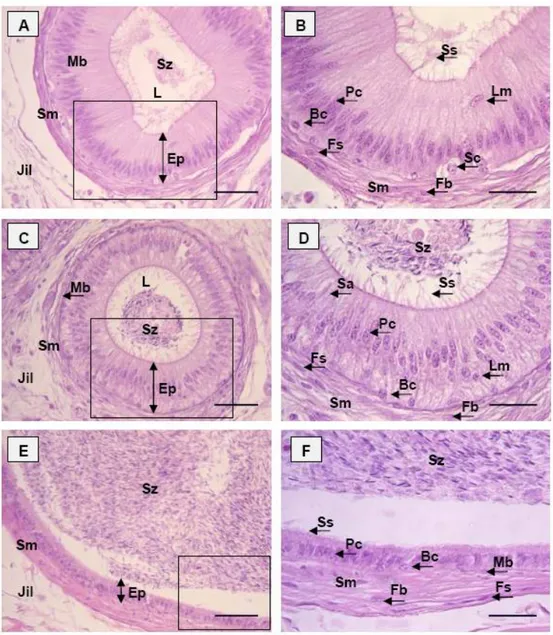
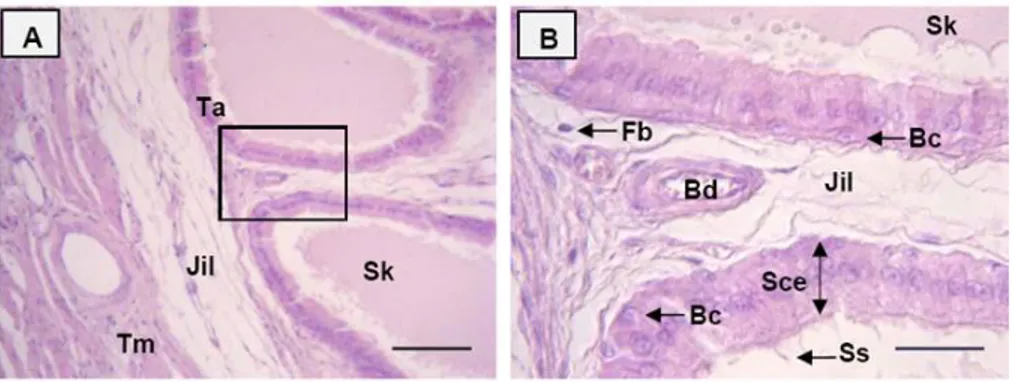
Posedur anatomi dan histologi dilakukan untuk memperoleh data tentang anatomi dan histologi organ reproduksi dari seekor muncak jantan dewasa dengan kode ♂#1 pada periode RK. Pengamatan morfologi, morfometri, struktur histologi organ reproduksi muncak jantan meliputi testis, duktus epididimidis, kelenjar asesoris kelamin dan penis. Karakteristik organ reproduksi muncak secara umum mirip dengan ruminansia kecil dan spesies rusa lainnya. Muncak memiliki testis berbentuk bulat lonjong dengan kauda epididimidis yang teramati dengan jelas walaupun bagian tersebut masih terbungkus skrotum. Penis muncak bertipe fibroelastik dilengkapi dengan fleksura sigmoidea di bagian korpus penis. Glans penis muncak berukuran kecil dengan prosesus uretralis yang pendek. Kelenjar prostat secara makroskopis tidak ditemukan, namun secara mikroskopis kelenjar tersebut ditemukan di sekeliling uretra pars pelvina dalam bentuk pars diseminata. Ciri lainnya adalah kelenjar bulbouretralis yang berukuran besar, seperti yang ditemukan pada reeves muntjak. Secara morfometri organ reproduksi muncak lebih kecil dibandingkan domba dan rusa
lainnya tidak dapat dilakukan, karena gambaran histologi seluruh organ reproduksi kecuali testis dan duktus epididimidis belum dilaporkan pada Cervidae tersebut. Data anatomi dan histologi organ reproduksi muncak jantan tersebut dapat digunakan untuk mengetahui fungsi fisiologi masing-masing organ dalam aktivitas reproduksi. Gambaran histologi testis pada periode ranggah keras dapat digunakan untuk menentukan tahapan epitel tubuli seminiferi dan durasinya selama spermatogenesis berlangsung. Selain itu, ukuran panjang penis muncak dapat digunakan untuk memprediksi ukuran panjang saluran reproduksi muncak betina yang penting diketahui untuk aplikasi teknik inseminasi buatan.
Informasi mengenai karakteristik pertumbuhan ranggah muncak diperoleh dari dua ekor muncak jantan dewasa yang diberi kode ♂#2 dan ♂#3. Pengamatan dilakukan terhadap morfologi dan morfometri pertumbuhan RV dan RK post casting, serta durasi setiap periode ranggah. Pengamatan dilakukan selama mendekati satu siklus ranggah pada ♂#2, dan dua siklus ranggah pada ♂#3. Pada kegiatan ini juga diamati kaitan antara periode pertumbuhan ranggah dengan pemunculan perilaku spesifik, yaitu perilaku reproduksi dan agresif. Pertumbuhan RV dimulai setelah casting dengan urutan pertumbuhan ranggah velvet utama (RVU) dan diikuti ranggah velvet cabang (RVC). Kedua muncak mengalami urutan pertumbuhan RVU dan RVC yang sama, namun morfometrinya berbeda. Morfologi dan morfometri RK1 dan RK2 post casting pada ♂#2 berukuran lebih besar dan sudah memiliki cabang rangah dibandingkan RK1 dan RK2 pada ♂#3. Durasi masing-masing periode ranggah ♂#2 juga lebih panjang dibandingkan ♂#3 pada siklus ranggah pertama. Memasuki siklus ranggah kedua, terjadi perubahan morfologi dan peningkatan morfometri RK pada ♂#3. Hasil menunjukkan bahwa periode RK merupakan periode terpanjang dibandingkan periode C dan RV, dengan total durasi satu siklus ranggah muncak ♂#2 lebih lama (459 hari) dibandingkan ♂#3 pada siklus rangah pertama, yaitu 319 hari. Peningkatan umur dan ukuran tubuh ♂#3 berimplikasi terhadap perubahan morfologi dan peningkatan morfometri dan durasi satu siklus ranggah (489 hari). Oleh karena itu prediksi umur ♂#2 di awal penelitian adalah 5 tahun, sedangkan ♂#3 berumur 3 tahun. Prediksi umur ♂#2 dan ♂#3 tersebut diperlukan untuk mengetahui seberapa besar pengaruh status sosial, seperti peringkat dominasi dan pengaruh stimulo-social dari muncak betina fertil terhadap profil metabolit testosteron, aktivitas spermatogenesis, dan kualitas spermatozoa pada ♂#2 dan ♂#3, selama periode pertumbuhan ranggah. Perilaku reproduksi dan perilaku agresif ditemukan saat kedua muncak berada pada periode RK, dan jarang ditemukan pada periode C dan RV.
Aplikasi metode enzymeimmunoassay (EIA) menggunakan asai testosteron Möstl telah memperlihatkan profil testosteron kedua muncak selama periode pertumbuhan ranggah. Penggunaan asai testosteron telah memberikan hasil yang dapat menerangkan adanya keterkaitan antara status sosial seperti peringkat dominasi antara ♂#2 dan ♂#3 dengan pola sekresi testosteron selama periode pertumbuhan ranggah. Pola sekresi testosteron diketahui dari hasil pengukuran konsentrasi testosteron imunoreaktif (iT) selama periode RK, C, dan RV. Konsentrasi iT ♂#2 pada periode RK lebih tinggi dibandingkan saat muncak tersebut berada pada periode C (p = 0.009) dan RV (p = 0.06). Tingginya konsentrasi iT pada periode RK juga dilaporkan pada Cervidae lainnya yang ditandai dengan peningkatan aktivitas reproduksi. Pola sekresi testosteron ♂#3 pada siklus ranggah pertama sedikit berbeda dengan ♂#2, yaitu konsentrasi iT pada periode RK1 lebih rendah dibandingkan periode C1 (p = 0.05), C2
RV2. Pada saat ♂#3 memasuki siklus ranggah kedua, pola sekresi testosteron mulai menyerupai pola sekresi testosteron ♂#2, walaupun secara statistik tidak berbeda. Perubahan pola sekresi tersebut terjadi bersamaan dengan meningkatnya umur ♂#3 dan kematian ♂#2 pada saat ♂#3 berada pada awal periode RK2 dari siklus ranggah kedua. Pada periode awal pertumbuhan RV, konsentrasi iT relatif rendah dibandingkan pada periode RK. Pola ini dibuktikan dengan adanya korelasi negatif (pearson correlation, r = -0.785) antara laju pertumbuhan RV dengan konsentrasi iT pada ♂#2 dan secara statistik berbeda (p = 0.021), dan periode RV2 dari siklus ranggah kedua pada ♂#3 (pearson correlation, r = -0.574), namun secara statistik tidak berbeda (p = 0.311). Hasil lain memperlihatkan bahwa testosteron masih terdeteksi dalam konsentrasi yang lebih rendah walaupun muncak berada pada periode C dan RV. Hal ini mengindikasikan bahwa testosteron tetap diperlukan untuk kelangsungan aktivitas spermatogenesis selama satu siklus ranggah, meskipun aktivitas tersebut sedikit menurun dibandingkan saat muncak berada pada periode RK.
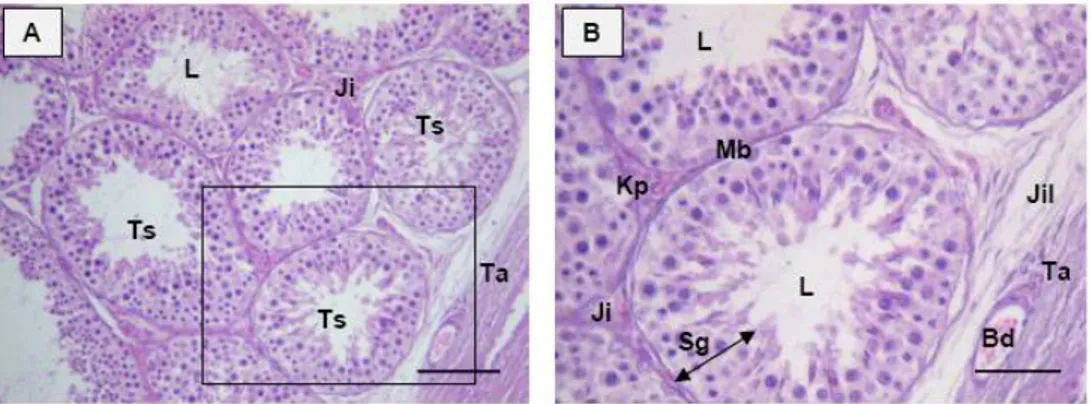
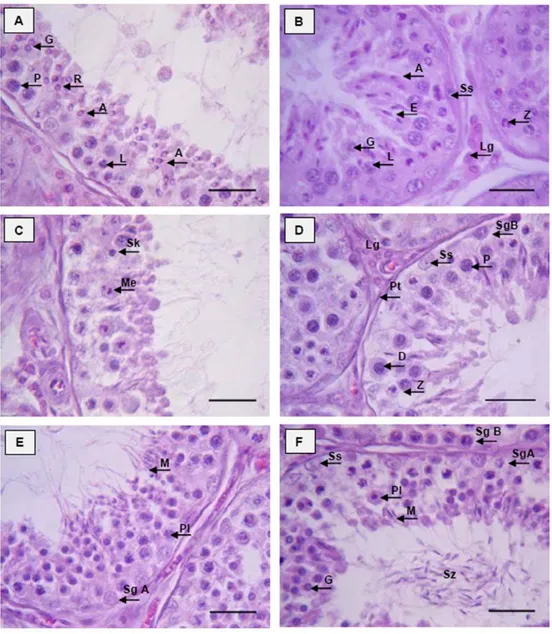
Berdasarkan metode morfologi tubular, ditemukan delapan tahapan epitel tubuli seminiferi (tahapan spermatogenik) pada muncak. Frekuensi dan durasi masing-masing tahapan yang dikelompokkan atas tahap pre meiosis (tahap I-III), tahap meiosis (tahap IV), dan tahap post meiosis (tahap V-VIII) bervariasi. Frekuensi tahap pre meiosis 47.75% lebih tinggi dibandingkan tahap meiosis 6.87%, dan post meiosis 43.37%, dengan durasi setiap tahapan adalah 5.07 hari, 0.73 hari, dan 4.81 hari. Identifikasi terhadap perkembangan spermatid pada periode RK, C, dan RV memperlihatkan gambaran diferensiasi round spermatid menjadi elongated spermatid. Hal ini mengindikasikan bahwa selama periode C dan RV, aktivitas spermatogenesis masih berlangsung dan dibuktikan dengan masih tingginya konsentrasi spermatozoa pada kedua periode tersebut, yaitu 262.5 juta/ml pada periode C, dan 362.60 ± 17.68 pada periode RV. Konsentrasi spermatozoa tertinggi, yaitu 506.25 ± 61.87 juta/ml ditemukan pada periode RK. Hasil tersebut paralel dengan konsentrasi testosteron yang terdeteksi pada setiap periode ranggah, namun berbeda dengan rusa timor dan spesies rusa lainnya. Periode aktif reproduksi pada rusa timor dan spesies rusa di wilayah temperate berlangsung selama periode RK. Hal berbeda ditemukan pada formosan muntjak dan axis deer, kedua spesies tersebut aktif bereproduksi sepanjang siklus pertumbuhan ranggahnya seperti yang diperlihatkan oleh muncak penelitian.
© Hak Cipta milik IPB, tahun 2012 Hak Cipta dilindungi Undang-undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah
b. Pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
2. Dilarang mengumumkan dan memperbanyak sebagian atau seluruh Karya tulis dalam bentuk laporan apa pun tanpa izin IPB
Muntiacus muntjak muntjak JANTAN: KAJIAN ANATOMI,
PROFIL METABOLIT TESTOSTERON, DAN SPERMATOGENESIS
SELAMA PERIODE PERTUMBUHAN RANGGAH
SRI WAHYUNI
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Biologi Reproduksi
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2012
Penguji pada Ujian Tertutup : Dr. Drh. Amrozi
Dr. Drh. Chairun Nisa’, M.Si, PAVet
Penguji pada Ujian Terbuka : Dr. Ir. Novianto Bambang Wawandono, M.Si. Dr. Drh. Bambang Purwantara, M.Sc.
muntjak Jantan: Kajian Anatomi, Profil Metabolit Testosteron, dan Spermatogenesis selama Periode Pertumbuhan Ranggah.
Nama : Sri Wahyuni
NIM : B362080011
Disetujui Komisi Pembimbing
Prof. Dr.Drh. Tuty L. Yusuf, M.S. Ketua
Drh. Srihadi Agungpriyono, Ph.D, PAVet (K). Dr. Drh. Muhammad Agil, M.Sc. Agr.
Anggota Anggota
Mengetahui
Ketua Program Studi Dekan Sekolah Pascasarjana
Biologi Reproduksi
Prof. Dr. Drh. M. Agus Setiadi Dr. Ir. Dahrul Syah, M.Sc. Agr.
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Disertasi yang berjudul Karakterisasi Reproduksi Muncak, Muntiacus muntjak muntjak Jantan: Kajian Anatomi, Metabolit Testosteron, dan Spermatogenesis selama Periode Pertumbuhan Ranggah, diajukan untuk memperoleh gelar doktor pada Sekolah Pascasarjana Institut Pertanian Bogor.
Terima kasih dan penghargaan yang tulus penulis sampaikan kepada Ibu Prof. Dr. Drh. Tuty L. Yusuf, M.S., Bapak Drh. Srihadi Agungpriyono, Ph.D, PAVet (K), dan Bapak Dr. Drh. Muhammad Agil, M.Sc. Agr., selaku Ketua dan Anggota Komisi Pembimbing atas bimbingan, arahan, dan saran yang diberikan selama penulis menjalani penelitian menempuh pendidikan doktor pada Program Studi Biologi Reproduksi, Sekolah Pascasarjana Institut Pertanian Bogor.
Ucapan terima kasih juga penulis sampaikan kepada:
1. Rektor Universitas Syiah Kuala dan Dekan Fakultas Kedokteran Hewan Universitas Syiah Kuala, Banda Aceh atas izin yang diberikan kepada penulis untuk melanjutkan studi doktor di PS BRP, SPs IPB.
2. Rektor Institut Pertanian Bogor dan Dekan Sekolah Pascasarjana (SPs IPB) yang telah menerima penulis sebagai mahasiswa di SPs IPB angkatan 2008.
3. Direktorat Jenderal Pendidikan Tinggi (DIKTI), Kementerian Pendidikan Nasional atas pemberian dana BPPS, beasiswa Sandwich-like 2010, dan Hibah Penelitian Disertasi Doktor tahun 2011.
4. Pemerintah Provinsi Aceh atas bantuan beasiswa yang diberikan selama penulis menempuh pendidikan.
5. Kementerian Kehutanan Republik Indonesia dan instansi terkait: Balai Konservasi Sumber Daya Alam (BKSDA) Jawa Tengah dan BKSDA Jawa Barat atas proses perizinan penggunaan muncak pada penelitian ini. 6. Pusat Penelitian Biologi LIPI atas rekomendasi yang diberikan terkait
penggunaan muncak penelitian.
7. Ibu Dr. Drh. Chairun Nisa’, M.Si, PAVet dan Bapak Dr. Drh. Amrozi, selaku penguji luar komisi pada ujian tertutup.
8. Bapak Dr. Ir. Novianto Bambang Wawandono, M.Si. dan Bapak Dr. Drh. Bambang Purwantara, M.Sc. selaku penguji luar komisi pada
ujian terbuka.
9. Bapak Prof. Dr. Drh. M. Agus Setiadi selaku Ketua PS BRP, SPs IPB beserta seluruh staf pendidik dan kependidikan atas bimbingan dan bantuan yang diberikan selama menjalankan studi di PS BRP.
10. Bapak Dr. Drh. Amrozi selaku Kepala Unit Rehabilitasi Reproduksi (URR) FKH IPB beserta staf atas izin penggunaan fasilitas kandang dan laboratorium serta bantuan teknis selama penelitian berlangsung.
11. Dr. Drh. Nurhidayat, PAVet, selaku Kepala Laboratorium Anatomi beserta staf: Prof. Dr. Drh. Koeswinarning Sigit, M.S., Dr. Drh. Heru Setijanto, PAVet., Dr. Drh. Chairun Nisa’, M.Si, PAVet., Dr. Drh. Savitri Novelina, M.Si, PAVet, dan Drh. Supratikno, M.Si, PAVet., yang telah memberikan
12. Prof. J. Keith Hodges dan Dr. Michael Heistermann dari Reproductive Biology Unit, German Primate Center, Goettingen, Germany beserta staf (Petra Kiesl dan Andrea Heistermann) atas bimbingan pada kegiatan analisis hormon muncak dalam Program Sandwich-like 2010.
13. Drh. I Ketut Mudite Adnyane, M.Si, Ph.D, PAVet., Ir. Asri Pudjirahaju, M.Si, dan Gholib, S.Pt, M.Si, atas segala bantuan yang diberikan selama penelitian.
14. Rumah Sakit Hewan IPB atas bantuan teknis saat proses pengambilan sampel penelitian.
15. Teman-teman PS. BRP angkatan 2008: Ir. Ekayanti M. Kaiin, M. Si., Ir. Tatan Kostaman, MP, dan Harry Murti, S.Si atas persahabatan yang baik, dukungan dan bantuan selama menjalankan studi di BRP.
16. Juliper Silalahi, SKH., Lidya M.Manik, SKH., Rissar Siringo Ringo, SKH, Danang Dwi Cahyadi, SKH., Drh. Dedi R.S, Bondan Achmadi, SE., Drh. Wahono Esthi Prasetyaningtyas, M.Si, PAVet., Drh. Kusdiantoro Mohammad, M.Si, PAVet., Forum Mahasiswa Pascasarjana Biologi Reproduksi (Forum Wacana BRP), Madia, Rudi, serta seluruh pihak yang tidak dapat disebutkan satu per satu, atas segala bantuan yang diberikan. Akhirnya ucapan terima kasih dan penghargaan yang tulus disampaikan kepada suami tercinta Said Ashim, SE dan ananda tersayang Said Muhammad Muafi, serta ayahanda dr. H. Anwar Jakfar, M.S. dan ibunda Hj. Trimurti Chaidir, kakanda Sri Wartini dan Sri Maryam, serta adinda Sri Haryani, Rahmat Hidayat, dan Firman Hidayat, keluarga besar Walid (alm) H. Said Ismail dan Ummi Hj. Syarifah Nurbasty atas segala dukungan dan doa yang dipanjatkan. Semoga karya ilmiah ini bermanfaat.
Bogor, Mei 2012 Sri Wahyuni
Penulis dilahirkan di Medan pada tanggal 19 November 1969 dari Ayahanda dr. H. Anwar Jakfar, M.S dan Ibunda Hj. Trimurti Chaidir sebagai anak ketiga dari enam bersaudara. Penulis menikah dengan Said Ashim, SE pada tahun 1996 dan dikarunai seorang putra, Said Muhammad Muafi. Saat ini penulis merupakan staf pengajar pada Fakultas Kedokteran Hewan Universitas Syiah Kuala, Banda Aceh.
Pendidikan dokter hewan diselesaikan pada tahun 1994 dari Fakultas Kedokteran Hewan Universitas Syiah Kuala, Banda Aceh. Pada tahun 2005 penulis memulai pendidikan magister (S2) di Sekolah Pascasarjana Institut Pertanian Bogor dan lulus pada tahun 2008. Pada tahun dan di perguruan tinggi yang sama, penulis melanjutkan pendidikan program doktor (S3) pada Program Studi Biologi Reproduksi (BRP).
Penelitian yang dilakukan merupakan penelitian eksplorasi mengenai karakteristik reproduksi muncak jantan yang tersebar di Indonesia. Satwa tersebut merupakan satwa yang dilindungi oleh Pemerintah Republik Indonesia sejak tahun 1999. Data yang diperoleh dari penelitian ini diharapkan dapat memberikan rekomendasi bagi program pengembangbiakan muncak di Indonesia. Sehubungan hal tersebut, penulis telah mensosialisasikan penelitian ini pada pertemuan Southeast Asia Veterinary Association (SEAVSA) di Bogor tahun 2010 dengan judul Testicular Needle Biopsy as an Alternative Biopsy Method for Assessment of Spermatogenesis in the Kijang Muntjak (Muntiacus muntjak muntjak). Publikasi pertama berjudul: Morfologi dan Morfometri Pertumbuhan Ranggah Velvet Muncak Jantan (Muntiacus muntjak muntjak), telah dimuat di Jurnal Kedokteran Hewan Universitas Syiah Kuala tahun 2011. Publikasi berikutnya berjudul Histologi dan Histomorfometri Testis dan Epididimis Muncak, Muntiacus muntjak muntjak pada Periode Ranggah Keras, akan dimuat pada Jurnal Veteriner Universitas Udayana pada bulan September 2012.
Halaman
DAFTAR TABEL ………. xv
DAFTAR GAMBAR ……… xvi
DAFTAR LAMPIRAN ………. xvii
DAFTAR SINGKATAN ……… xviii
PENDAHULUAN ………. 1 Latar Belakang ……… 1 Perumusan Masalah ……….. 4 Tujuan Penelitian ………. 4 Manfaat Penelitian ……….. 5 Kerangka Pemikiran ……… 5 TINJAUAN PUSTAKA ……… 9
Muncak dan Status Konservasinya ………. 9
Morfologi dan karakteristik Reproduksi Muncak Jantan……… 10
Pertumbuhan dan Siklus Ranggah ……….. 13
Biosintesis dan Metabolisme Androgen ……….. 15
Metode Pengukuran Kadar Metabolit Steroid ……… 17
Anatomi Organ Reproduksi Jantan ……….. 19
Spermatogenesis dan Tahapan Epitel Tubuli Seminiferi Testis……….. 21
Morfologi dan Kualitas Semen ……….. 24
Peluang Pemanfaatan Ejakulat Muncak ………. 25
ANATOMI DAN HISTOLOGI ORGAN REPRODUKSI MUNCAK JANTAN PADA PERIODE RANGGAH KERAS ………. 29 Abstrak ………. 29
Abstract ………... 29
Pendahuluan ……… 30
Bahan dan Metode ………. 32
Hasil dan Pembahasan ………. 37
Simpulan ……….. 67
Daftar Pustaka ……… 67
KARAKTERISTIK PERTUMBUHAN RANGGAH MUNCAK……….. 73
Abstrak ………. 73
Abstract ……… 73
Pendahuluan ……… 74
Bahan dan Metode ………. 76
Hasil dan Pembahasan ……….. 81
Simpulan ……….. 97
PERIODE PERTUMBUHAN RANGGAH ………
Abstrak ………. 101
Abstract ……….. 101
Pendahuluan ……….. 102
Bahan dan Metode ……… 104
Hasil dan Pembahasan ………. 110
Simpulan ………. 122
Daftar Pustaka ……… 122
SPERMATOGENESIS DAN KUALITAS SEMEN MUNCAK SELAMA PERIODE PERTUMBUHAN RANGGAH………. 127 Abstrak ……… 127
Abstract ……….. 127
Pendahuluan ……….. 128
Bahan dan Metode ………. 130
Hasil dan Pembahasan ……… 137
Simpulan ………. 161
Daftar Pustaka ……… 161
PEMBAHASAN UMUM ……….. 167
SIMPULAN DAN SARAN UMUM ………. 175
DAFTAR PUSTAKA ………... 176
LAMPIRAN ……….. 187 .
Halaman
1 Kelenjar asesoris pada beberapa spesies hewan ………. 20 2 Morfometri organ reproduksi muncak jantan ……….. 39 3 Perbandingan morfometri organ reproduksi jantan ……….. 40 4 Karakteristik sel epitel germinal tubuli seminiferi muncak ……… 46 5 Morfometri duktus epididimidis muncak ……….. 52 6 Keberadaan kelenjar asesoris kelamin muncak ………. 56 7 Morfometri ranggah keras post casting pada muncak ... 89 8 Durasi siklus ranggah muncak …..……… 90 9 Kriteria penentuan umur muncak pada periode awal penelitian ………. 91 10 Perilaku percumbuan (courtship) pada muncak ……… 93 11 Perilaku agresif pada muncak ……….. 95 12 Pemunculan perilaku reproduksi dan agresif pada muncak ……… 96 13 Frekuensi dan durasi delapan tahapan epitel tubuli seminiferi ………. 143 14 Ukuran sel germinal tubuli seminiferi testis muncak ………. 146 15 Tahapan epitel tubuli seminiferi testis pada setiap periode ………. 147 16 Tipe sel germinal tubuli seminiferi testis ……….……… 149 17 Morfometri testis dan skrotum muncak selama periode ……... 151 18 Morfometri spermatozoa muncak ... 153 19 Kualitas semen muncak selama periode pertumbuhan ranggah………. 156
Halaman
1 Bagan alir kerangka penelitian ……… 7 2 Muncak (Muntiacus muntjak muntjak, Zimmermann 1780) jantan …… 9 3 Lokasi penyebaran muncak di Indonesia ………. 10 4 Morfologi tulang kepala muncak jantan (Muntiacus sp) ………. 11 5 Regulasi hormonal pertumbuhan ranggah Cervidae ………. 14 6 Regulasi steroidogenesis androgen ……….. 16
7 Anatomi testis ……… 21
8 Spermatogenesis pada mamalia ……… 22 9 Morfologi spermatozoa mamalia ……… 25 10 Bagan alir disain penelitian I ……… 36 11 Orientasi eksternal testis muncak ………. 37 12 Anatomi organ reproduksi muncak jantan ……… 38 13 Morfologi testis dan duktus epididimidis ……… 42 14 Struktur histologi testis muncak jantan ………. 43 15 Tipe sel epitel germinal tubuli seminiferi testis muncak ……….. 45 16 Struktur histologi duktus epididimidis ………. 51 17 Duktus deferens muncak ………. 54 18 Morfologi kelenjar asesoris kelamin muncak ……… 55 19 Morfologi komparatif kelenjar asesoris kelamin muncak ……… 56 20 Struktur histologi ampula muncak ………. 58 21 Struktur histologi kelenjar vesikularis muncak ………. 59 22 Struktur histologi pars diseminata prostat muncak ………. 61 23 Struktur histologi kelenjar bulbouretralis muncak ……… 64 24 Morfologi penis muncak ……….. 65 25 Struktur histologi penis muncak ………. 66 26 Muncak (Muntiacus muntjak muntjak) jantan ……… 77 27 Pengukuran panjang dan diameter ranggah keras …………... 78 28 Bagan alir disain penelitian II ……….. 80 29 Pertumbuhan ranggah velvet muncak ………... 83 30 Morfometri pertumbuhan ranggah velvet ♂#2………... 85 31 Morfometri pertumbuhan ranggah velvet ♂#3……….………. 86
33 Perilaku percumbuan (courtship) pada muncak ……….. 94 34 Perilaku agresif pada muncak ………. 95 35 Uji paralel dari asai testosteron (Möstl)...……….. 107 36 Bagan alir disain penelitian III……….………. 109 37 Konsentrasi testosteron imunoreaktif (iT) ♂#2 ……… 111 38 Perbedaan konsentrasi testosteron imunoreaktif (iT) ♂#2 ...…………. 112 39 Konsentrasi testosteron imunoreaktif (iT) ♂#3 ……… 113 40 Perbedaan konsentrasi testosteron imunoreaktif (iT) ♂#3 ………. 114 41 Hubungan pertumbuhan ranggah dan konsentrasi iT ♂#2 ……… 118 42 Hubungan pertumbuhan ranggah dan konsentrasi iT ♂#3 siklus I ….. 120 43 Hubungan pertumbuhan ranggah dan konsentrasi iT ♂#3 siklus II .... 120 44 Bagan alir disain penelitian IV ………. 136 45 Delapan tahapan epitel tubuli seminiferi muncak ……… 139 46 Skema delapan tahapan epitel tubuli seminiferi testis muncak ………. 140 47 Perkembangan sel germinal, durasi spermatogenesis ………... 143 48 Spermatogenesis pada tubuli seminiferi testis ... 148 49 Morfologi spermatozoa muncak ………. 152 50 Ultrastruktur spermatozoa muncak ……… 152 51 Morfologi spermatozoa abnormal pada muncak ………. 159
Halaman
1 Prosedur pewarnaan hematoksilin-eosin (HE) ………. 187 2 Prosedur pewarnaan periodic acid Schiff (PAS) ………. 188 3 Preparasi spermatozoa untuk SEM ……… 189 4 Prosedur pewarnaan William ……….. 190
Singkatan Kata 5α-DHT 5α-dehidrotestosteron
17β-HSD 17β-hidroksisteroid dehidrogenase
♂#1 Muncak jantan nomor 1
♂#2 Muncak jantan nomor 2
♂#3 Muncak jantan nomor 3
ABP Androgen binding protein
ACTH Adrenocorticothropic hormone
C Casting
C1 Casting 1
C2 Casting 2
CNB Core needle biopsy
D Diplotene
DHEA Dehidroepiandrosteron DHT Dehidrotestosteron
DRKC Diameter ranggah keras cabang
DRKU Diameter ranggah keras utama
DRVC Diameter ranggah velvet cabang
DRVU Diameter ranggah velvet utama
E Elongated spermatid
EIA Enzymeimmunoassay
GnRH Gonadothropin releasing hormone
HE Hematoksilin-eosin HPG Hipotalamus-hipofise-gonad IB Inseminasi buatan iT Testosteron imunoreaktif L Leptotene LC Least concern LH Luteinizing hormone P Pachytene
PAS Periodic acid Schiff
PC Principle cells
Pl Preleptotene
PRKC Panjang ranggah keras cabang
PRKU Panjang ranggah keras utama
PRVC Panjang ranggah velvet cabang
PRVU Panjang ranggah velvet utama
R Round spermatid RIA Radioimmunoassay RK Ranggah keras RK1 Ranggah keras 1 RK2 Ranggah keras 2 RK3 Ranggah keras 3
RKU Ranggah keras utama
RV Ranggah velvet
RV1 Ranggah velvet 1
RV2 Ranggah velvet 2
RVC Ranggah velvet cabang
RVU Ranggah velvet utama
SEM Scanning electron microscope
SHBG Steroid hormone-binding globulin
Ss Spermatosit sekunder
Sz Spermatozoa
VNO Vomeronasal organ
Z Zygotene
PENDAHULUAN UMUM
Latar Belakang
Pengetahuan dan informasi mengenai biologi reproduksi suatu spesies satwa khususnya satwa liar sangat diperlukan untuk mensukseskan program konservasi satwa tersebut. Biologi reproduksi satwa liar bersifat spesifik spesies, dengan variasi kompleks yang meliputi struktur anatomi dan fungsi organ reproduksi, profil hormon reproduksi, siklus estrus pada betina, fisiologi gamet serta deposisi semen pada organ reproduksi betina. Data biologi reproduksi tersebut sangat penting untuk mendukung program pemeliharaan, pengembangbiakan maupun penerapan teknologi reproduksi seperti inseminasi buatan, embrio transfer, fertilisasi in vitro, bank plasma nutfah (genome resource bank), mikromanipulasi gamet, semen sexing dan transfer inti sel somatis pada satwa liar (Comizolli et al. 2000; Andrabi dan Maxwell 2007). Selain itu informasi dasar mengenai biologi reproduksi satwa liar, juga bermanfaat sebagai bahan pertimbangan dalam penentuan kebijakan dan strategi pengelolaan satwa liar di penangkaran. Keberhasilan dari strategi tersebut akan menjamin dan meningkatkan populasi satwa yang dilindungi dan terancam punah.
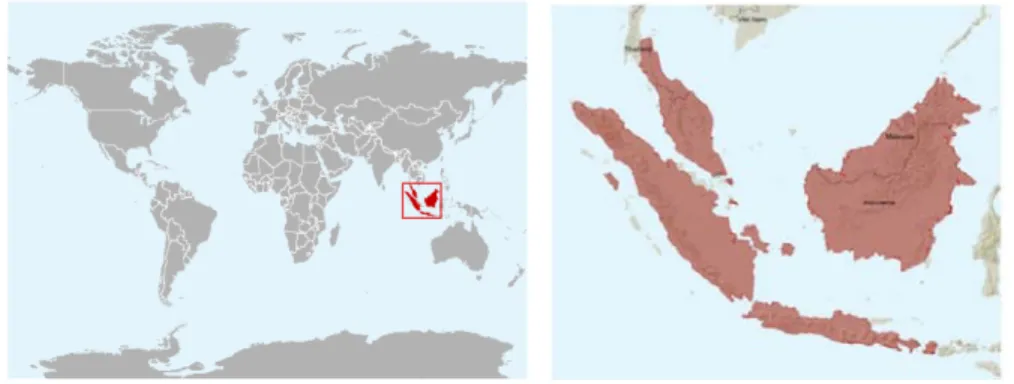
Muncak (Muntiacus muntjak muntjak) tersebar di Pulau Jawa dan Pulau Sumatera bagian selatan. Menurut International Union for Conservation of Nature and Natural Resources (IUCN), status konservasi muncak (kecuali M. criniforns) adalah tergolong least concern (LC) atau berisiko rendah untuk punah (IUCN 2011). Walaupun status muncak masih tergolong LC, namun seluruh sub spesies muncak yang tersebar di Indonesia telah dilindungi seperti yang tercantum dalam Daftar Lampiran Peraturan Pemerintah Republik Indonesia Nomor 7 Tahun 1999 sejak tanggal 27 Januari 1999 (PHKA 2004). Penetapan status perlindungan muncak di Indonesia, disebabkan telah terjadinya penurunan populasi muncak di habitat alami. Laporan tentang populasi muncak di habitat alami maupun di area konservasi hingga saat ini belum tersedia. Kekhawatiran akan punahnya muncak di Indonesia menjadi alasan penting dilakukannya upaya pengembangbiakan muncak di luar kawasan konservasi. Namun demikian, upaya tersebut harus didukung dengan ketersediaan data aspek reproduksi yang dapat digunakan untuk menentukan aktivitas dan pola reproduksi muncak Indonesia yang sampai saat ini belum dilaporkan.
Cervidae jantan dicirikan dengan keberadaan ranggah (Bubenik 2006). Pada beberapa spesies rusa, pertumbuhan ranggah berkorelasi erat dengan aktivitas organ reproduksinya, seperti sintesis testosteron, spermatogenesis untuk menghasilkan spermatozoa dan aktivitas kelenjar asesoris kelamin dalam menghasilkan plasma semen. Hal tersebut telah dilaporkan pada rusa timor jantan, yang memperlihatkan adanya keterkaitan antara aktivitas reproduksi dengan pertumbuhan ranggahnya (Handarini et al. 2004; Handarini dan Nalley 2008). Demikian pula pada spesies rusa lainnya seperti axis deer (Axis axis) (Loudon dan Curlewis 1988), pampas deer (Ozotoceros bezoarticus
bezoarticus) (Pereira et al. 2005) dan red deer (Cervus elaphus) (Bartos et al. 2009). Namun demikian, pola reproduksi seperti yang ditemukan
pada beberapa spesies rusa tersebut sedikit berbeda pada formosan muntjak. Walaupun pertumbuhan ranggah berkorelasi erat dengan konsentrasi testosteron, namun aktivitas reproduksi formosan muntjak tetap berjalan sepanjang tahun (Pei et al. 2009) yang ditandai dengan ditemukannya spermatozoa motil pada berbagai stadia ranggah (Liu et al. 2004). Pola reproduksi muncak yang tersebar di Indonesia hingga saat ini belum dilaporkan, apakah terdapat kemiripan dengan pola reproduksi formosan muntjak atau menyerupai pola reproduksi rusa yang hidup di berbagai negara termasuk Indonesia.
Sejauh ini informasi mengenai pola reproduksi genus Muntiacus masih terbatas pada spesies reeves muntjak (Muntiacus reevesi) (Chapman dan Harris
1991) yang diintroduksi ke Inggris, dan sub spesies formosan muntjak (M. reevesi micrurus) yang berasal dari Taiwan (Pei et al. 2009). Di sisi lain,
penelitian pada beberapa spesies rusa famili Cervidae di Indonesia telah dilakukan, diantaranya tentang karakteristik ranggah rusa timor (Cervus timorensis) (Semiadi 1997) dan pertumbuhan ranggah pada rusa bawean (Axis kuhlii) (Semiadi et al. 2003). Selain itu aplikasi teknik inseminasi buatan (IB) juga telah dilakukan pada rusa timor, rusa sambar (Cervus unicolor) dan rusa bawean (Dradjat 2002).
Sehubungan hal tersebut, ada beberapa metode pengamatan yang dapat digunakan untuk mengetahui karakteristik dan pola reproduksi muncak selama satu siklus ranggah. Pengamatan tersebut dilakukan terhadap: 1) profil metabolit testosteron yang dilakukan secara non invasif, 2) pertumbuhan ranggah, dan 3) spermatogenesis dan kualitas semen selama periode pertumbuhan ranggah.
Pengamatan terhadap ketiga aspek reproduksi tersebut terlebih dahulu diawali dengan pengamatan terhadap anatomi dan histologi organ reproduksi muncak jantan yang hingga saat ini datanya belum tersedia.
Pada penelitian ini, penentuan karakteristik dan pola reproduksi muncak jantan berdasarkan profil metabolit testosteron terkait aktivitas steroidogenesis selama periode pertumbuhan ranggah, dilakukan dengan teknik non invasif menggunakan metode enzymeimmunoassay (EIA). Melalui teknik dan metode tersebut, aktivitas steroidogenesis selama periode pertumbuhan ranggah dapat dideteksi berdasarkan konsentrasi testosteron imunoreaktif (iT). Teknik non invasif telah dikenal luas sebagai teknik yang praktis, murah, dan cocok digunakan pada berbagai spesies satwa liar, baik yang berada di habitat ex situ maupun in situ (Mauget et al. 2007). Aplikasi metode EIA pada badak sumatera (Dicerorhinus sumatrensis) (Heistermann et al. 1998; Agil 2007) telah memberikan hasil dengan tingkat validitas yang baik. Pada muncak, teknik analisis tersebut baru dilakukan pada formosan muntjak dengan metode radioimmunoassay (RIA) (Pei et al. 2009). Analisis dengan metode EIA sejauh ini belum dilakukan, sehingga metode EIA sangat sesuai diaplikasikan pada penelitian ini.
Karakteristik dan pola reproduksi muncak jantan selama periode pertumbuhan ranggah dapat pula diketahui dari aktivitas spermatogenesis pada tubuli seminiferi testis muncak. Kemungkinan adanya perbedaan aktivitas spermatogenesis selama periode ranggah keras, casting, dan ranggah velvet, dapat diketahui berdasarkan proses diferensiasi spermatid untuk menghasilkan spermatozoa pada ketiga periode pertumbuhan ranggah tersebut. Diferensiasi spermatid dapat dideteksi menggunakan metode pewarnaan periodic acid Schiff (PAS).
Evaluasi terhadap semen muncak dapat memberi gambaran tentang aktivitas spermatogenesis dan fertilitas muncak selama satu siklus ranggah. Motilitas, konsentrasi, jumlah spermatozoa hidup dan tingkat abnormalitasnya menjadi karakteristik penting dalam menentukan fertilitas satwa tersebut. Selain itu, data tentang kualitas spermatozoa selama periode pertumbuhan ranggah dapat digunakan untuk menentukan waktu yang tepat untuk penampungan semen. Manfaat lainnya adalah sebagai data pendukung pada teknik preservasi spermatozoa muncak yang dapat digunakan untuk IB dan disimpan sebagai materi genetik.
Perumusan Masalah
Informasi berupa data lengkap yang berhubungan dengan karakteristik reproduksi pada muncak jantan belum dilaporkan, yaitu:
1. Anatomi dan histologi organ reproduksi.
2. Pertumbuhan ranggah, durasi siklus ranggah dan perilaku spesifik selama periode ranggah.
3. Profil hormon reproduksi selama periode pertumbuhan ranggah.
4. Aktivitas reproduksi selama periode pertumbuhan ranggah yang dikaitkan dengan spermatogenesis dan kualitas semen.
5. Pola reproduksi selama periode pertumbuhan ranggah.
Tujuan Penelitian
Secara umum, penelitian ini bertujuan untuk mendukung program konservasi muncak melalui penyediaan data yang berhubungan dengan aspek reproduksi, baik yang dilakukan dengan perkawinan alami maupun penerapan teknologi reproduksi. Adapun tujuan khusus dari penelitian ini adalah mempelajari karakteristik reproduksi muncak jantan selama periode pertumbuhan ranggah dengan cara menganalisis dan menggali informasi mengenai:
1. Anatomi dan histologi organ reproduksi.
2. Pertumbuhan ranggah dan durasi siklus ranggah, serta perilaku spesifik selam periode pertumbuhan ranggah.
3. Profil metabolit testosteron dan korelasinya dengan pertumbuhan ranggah.
4. Spermatogenesis dan kualitas semen.
Manfaat Penelitian
Seluruh data yang diperoleh dari penelitian ini diharapkan dapat:
1. Memberikan informasi mengenai karakteristik reproduksi muncak jantan yang dapat digunakan sebagai bahan rekomendasi pada program pengembangbiakan muncak di penangkaran.
2. Sebagai data dasar untuk aplikasi teknologi reproduksi pada muncak. 3. Mendukung program konservasi plasma nutfah melalui preservasi semen
4. Mendukung program budidaya muncak yang dapat digunakan sebagai satwa harapan penghasil protein alternatif asal hewan.
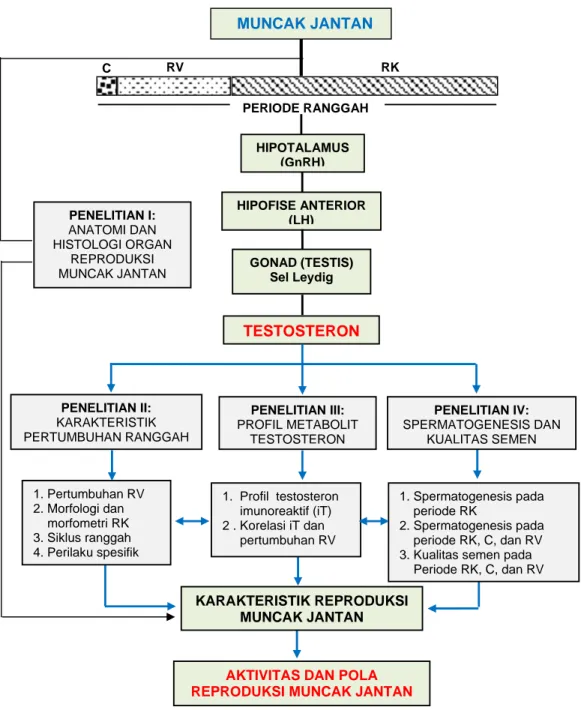
Kerangka Pemikiran
Ranggah merupakan karakteristik reproduksi sekunder yang dimiliki oleh sebagian besar Cervidae jantan, termasuk muncak. Pertumbuhan ranggah bersifat tahunan yang disebut dengan siklus pertumbuhan ranggah. Satu siklus pertumbuhan ranggah terbagi atas periode ranggah keras (RK), tanpa ranggah atau casting (C), dan ranggah lunak atau ranggah velvet (RV). Tumbuh kembangnya ranggah, berada dibawah kontrol poros hipotalamus-hipofise-gonad yang melibatkan GnRH dari hipotalamus, LH dari hipofise anterior dan testosteron dari gonad (testis). Tingginya konsentrasi LH yang terjadi pada periode ranggah keras (RK) akan menstimulasi sel-sel Leydig testis untuk mensintesis testosteron dalam konsentrasi tinggi. Testosteron diperlukan untuk membantu spermatogenesis (efek parakrin) khususnya pada proses spermiogenesis. Tingginya konsentrasi testosteron selama periode RK akan meningkatkan aktivitas spermatogenesis yang dapat diketahui dari konsentrasi spermatozoa, meningkatkan perilaku agresif dan tingginya libido. Pada akhir periode RK dan saat memasuki periode C, konsentrasi testosteron menurun drastis. Kondisi ini terjadi karena testosteron dalam konsentrasi tinggi di sirkulasi darah memberikan umpan balik negatif ke hipotalamus dan hipofise, sehingga sekresi GnRH dan LH menurun. Akibatnya konsentrasi testosteron juga menurun dan berefek pada penurunan aktivitas reproduksi. Setelah periode C, muncak memasuki periode RV yang ditandai dengan masih rendahnya testosteron akibat belum optimalnya rangsangan LH pada sel Leydig testis. Akhir periode RV (shedding) merupakan awal periode RK baru dengan konsentrasi testosteron yang tinggi dan muncak memasuki periode aktif reproduksi.
Dari uraian tersebut, dapat dijelaskan bahwa konsentrasi testosteron berpengaruh terhadap periode pertumbuhan ranggah, spermatogenesis dan kualitas spermatozoa dan pemunculan perilaku spesifik seperti perilaku reproduksi dan agresif. Sejauh mana keterkaitan antara testosteron dan beberapa aktivitas reproduksi pada muncak jantan, dapat diketahui dari kegiatan penelitian yang dibagi atas empat tahapan penelitian. Masing-masing penelitian dilakukan dengan metode spesifik yang hasil dan pembahasannya disampaikan pada bagian tersendiri dari disertasi ini, dengan judul:
1. Anatomi dan histologi organ reproduksi muncak jantan. 2. Karakteristik pertumbuhan ranggah muncak jantan.
3. Profil metabolit testosteron muncak jantan selama periode pertumbuhan ranggah.
4. Spermatogenesis dan kualitas semen muncak selama periode pertumbuhan ranggah.
Keterkaitan antara data yang diperoleh setiap penelitian tersebut khususnya data penelitian II, III, dan IV, akan dibahas di bagian pembahasan umum dari disertasi ini. Dari pembahasan tersebut dapat diketahui fertilitas dan status reproduksi muncak jantan selama satu siklus ranggah.
Gambar 1 Bagan alir kerangka pemikiran. GnRH: gonadotrophin releasing hormone, LH: luteinizing hormone, C: casting (lepas ranggah), RV: ranggah velvet, RK: ranggah keras. TESTOSTERON PERIODE RANGGAH C RV RK MUNCAK JANTAN HIPOTALAMUS (GnRH) HIPOFISE ANTERIOR (LH) PENELITIAN III: PROFIL METABOLIT TESTOSTERON PENELITIAN IV: SPERMATOGENESIS DAN KUALITAS SEMEN PENELITIAN II: KARAKTERISTIK PERTUMBUHAN RANGGAH 1. Pertumbuhan RV 2. Morfologi dan morfometri RK 3. Siklus ranggah 4. Perilaku spesifik KARAKTERISTIK REPRODUKSI MUNCAK JANTAN PENELITIAN I: ANATOMI DAN HISTOLOGI ORGAN REPRODUKSI MUNCAK JANTAN 1. Spermatogenesis pada periode RK 2. Spermatogenesis pada periode RK, C, dan RV 3. Kualitas semen pada
Periode RK, C, dan RV 1. Profil testosteron
imunoreaktif (iT) 2 . Korelasi iT dan pertumbuhan RV
AKTIVITAS DAN POLA REPRODUKSI MUNCAK JANTAN
GONAD (TESTIS) Sel Leydig
y s M S T d d M S d j m o f s g s s Muncak dan Mun yang terseb sepuluh spe M. rooseve Spesies M 15 sub spe Thailand, M di Indonesia dan Nias, M. m. banc Selatan dan dan Pulau (Maryanto e juga munca muntjak. De ordo famili sub famili genus spesies sub spesies Gambar 2 n Status Ko cak (Muntia ar luas di As esies, namu ltorum, M. M. muntjak (i esies yang t alaysia dan a adalah: M. M. m. r canus di Pu n Pulau Jawa Jawa; da et al. 2008). ak jawa me ensie (1970) : Artiodac : Cervida : Cervina : Muntiac : Muntiac : Muntiac Muncak (M periode ra onservasiny acus sp) ter sia (Gamba un saat ini h feae, M. c indian muntj tersebar mu Indonesia. m. montanu robinsoni d ulau Bangka a; M. m. ple an M. m. Sub spesie emiliki gamb mengklasifi ctyla ae ae cus cus muntjak cus muntjak Muntiacus mu anggah velve ya rgolong artio r 2 dan 3). P hanya tersis rinifrons da tjak) sering ulai dari Ind Beberapa s us di Pulau S di Kepulau a dan Belitu echaricus di nainggolan es M. m. Mu baran morfo kasikan mun muntjak, Zim untjak muntja t. odactyla (he Pada awalny a lima spes n M. muntj disebut bar dochina, Ind ub spesies Sumatera (b an Riau ung, M. m. Pulau Kalim ni di Pulau untjak Zimm ologis yang ncak sebaga mmermann 1 ak, Zimmerm ewan berku ya, muncak sies, yaitu: M jak (Ma et rking deer, t dia, Srilanka muncak yan bagian utara (Bintan) da muntjak di matan, Bawa u Bali dan mermann 178 mirip deng ai berikut: 1780 ann 1780) j uku genap) terdiri atas M. reevesi, al. 1986). terdiri atas a, Vietnam, ng terdapat dan barat) an Linga; Sumatera al, Matasiri, n Lombok 80, disebut gan indian antan pada
Muncak (M. m. muntjak) dan sub spesies muncak lainnya yang tersebar di Indonesia merupakan satwa yang dilindungi di Indonesia, seperti yang tercantum di dalam Daftar Lampiran Peraturan Pemerintah Republik Indonesia Nomor 7 tahun 1999, sejak tanggal 27 Januari 1999 (PHKA 2004). Namun demikian, satwa ini belum tercantum di dalam Daftar Appendix CITES (Convention on International Trade in Endangered Species of Fauna and Flora). Berdasarkan data International Union for Conservation of Nature and Natural Resources (IUCN), terdapat satu spesies muncak dengan status vulnerable di dunia, yaitu Muntiacus crinifrons (Black Muntjak) yang berasal dari China (Wemmer 1998). Sedangkan spesies muncak lainnya termasuk bahan penelitian ini (M. m. muntjak) masih memiliki status least concern (IUCN 2011). Populasi muncak di habitat asli diduga semakin menurun, seperti yang terjadi pada populasi spesies satwa liar Indonesia lainnya. Penyebab utama dari penurunan populasi muncak di Indonesia adalah hilangnya habitat satwa seiring dengan alih fungsi hutan sebagai lahan pertanian, dan maraknya perburuan liar.
Gambar 3 Lokasi penyebaran muncak di Indonesia yang meliputi Pulau Sumatera, Pulau Jawa, Pulau Kalimantan, Pulau Bali dan Lombok (Sumber: IUCN 2011). Morfologi dan Karakteristik Reproduksi Muncak Jantan
Secara umum, tubuh seluruh spesies muncak (Muntiacus sp) ditutupi oleh rambut pendek, lembut, berwarna kuning hingga coklat tua serta abu-abu kecoklatan. Rambut di bagian perut berwarna lebih terang (Worlddeer 2005; Maryanto et al. 2008). Muncak jantan berukuran lebih besar dari betina dengan berat badan 22 kg (jantan) dan 20 kg (betina) (Philips 1984), memiliki ranggah, taring dan kelenjar preorbital di bagian frontal wajah. Kelenjar ini berfungsi sebagai penghasil sekreta yang berfungsi sebagai marker untuk menentukan daerah teritorialnya (Jackson 2002). Ukuran tubuh M. muntjak sedikit lebih besar
dibandingkan dengan M. crinifrons, yaitu dengan rataan panjang tubuh 1043 mm, tinggi bahu 611 mm, panjang kepala 209 mm dengan ukuran fossa lacrimalis lebih kecil dari pada fossa orbitalis (Ma et al. 1986).
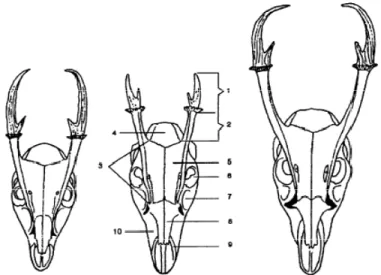
Ranggah merupakan organ asesoris reproduksi sekunder yang tumbuh apabila seekor rusa jantan telah mencapai pubertas (Wallace dan Birtles 1985). Selain sebagai simbol keindahan, ranggah juga digunakan sebagai organ pertahanan saat terjadi perkelahian dengan jantan lainnya (Price et al. 2005). Dari seluruh Cervidae jantan di dunia, hanya ada beberapa spesies yang tidak memiliki ranggah, yaitu chinese water deer (Hydropotes inermis), rusa musk (Moschus chrysogaster) dan mouse deer (Moschiola mimenoides). Selain pada rusa jantan, ranggah juga ditemukan pada rusa betina, yaitu pada rheindeer (Rangifer tarandus), namun ukuran ranggahnya lebih kecil dibandingkan rheindeer jantan (Rajaram 2004). Muncak yang tergolong Cervidae memiliki bentuk ranggah yang berbeda dengan ranggah Cervidae lainnya, yaitu berukuran pendek dengan ujung tanduk cenderung melengkung ke dalam pada muncak berumur tua dan memiliki pedikel (dasar ranggah) yang panjang (Gambar 4). Panjang ranggah pada beberapa spesies muncak bervariasi, yaitu 29-47 mm pada M. reevesi, 38-47 mm pada M. rooseveltorum, 36-49 mm pada M. feae, 48-55 mm pada M. crinifrons dan 48-84 mm pada M. Muntjak (Ma et al. 1986).
Gambar 4 Morfologi tulang kepala muncak jantan (Muntiacus sp). Muntiacus reevesi (kiri), Muntiacus feae (tengah) dan Muntiacus muntjak (kanan). Ranggah (1), pedicle (2), frontal ridge (3), os temporale (4), os frontale (5), foramen orbitalis (6), fossa lacrimalis (7), os nasale (8), os premaxillaris (9), dan os maxillaris (10) (Sumber: Ma et al. 1986).
Ciri khas muncak jantan lainnya adalah sepasang gigi taring atas yang tumbuh dari os maxillaris dengan ukuran sekitar 2 cm yang tetap terlihat saat mulutnya tertutup. Pada saat melakukan aktivitas reproduksi, taring lebih berperan sebagai alat pertahanan saat terjadi kompetisi dengan jantan lainnya dari pada ranggah (Chapman 1997). Selain itu, muncak memiliki vokalisasi spesifik berupa suara gongongan sebagai panggilan atau peringatan kepada muncak lainnya agar menghindar dari predator. Namun vokalisasi tersebut tidak digunakan pada saat aktivitas reproduksi dan untuk penentuan teritorialnya (Oli dan Jacobson 1995).
Perilaku reproduksi M. muntjak secara umum mirip dengan Cervidae lainnya, yaitu poligami pada jantan dan poliestrus pada betina. Aktivitas organ reproduksi jantan dan betina berlangsung setelah muncak berumur di atas satu tahun. Siklus estrus muncak betina berkisar antara 14-21 hari dengan durasi estrus selama dua hari. Sedangkan periode gestasi berlangsung selama 210 hari dengan jumlah anak 1-2 ekor per kelahiran (Jackson 2002) dengan jarak per kelahiran sekitar tujuh bulan (Barrette 2008). Aktivitas reproduksi formosan muntjak (Muntiacus reeves micrurus) menunjukkan pola yang tidak berhubungan dengan periode pertumbuhan ranggah dan konsentrasi testosteron. Walaupun diketahui testosteron dibutuhkan dalam konsentrasi tinggi untuk pertumbuhan ranggah, namun pada saat konsentrasi testosteron rendah, spermatogenesis tetap berlangsung (Pei et al. 2009). Hal ini dibuktikan dengan ditemukannya spermatozoa selama satu siklus ranggah (Liu et al. 2004).
Aktivitas reproduksi muncak seperti tersebut diatas berbeda dengan Cervidae yang hidup di negara empat musim yang dipengaruhi oleh panjang-pendeknya hari (photoperiod). Pada saat intensitas pencahayaan sedikit, terjadi peningkatan sekresi hormon gonadotropin, aktivitas testikular dan aktivitas reproduksi lainnya (Lincoln 1985; Semiadi 1995). Ukuran testis, kelenjar asesoris kelamin, sekresi hormon dan spermatogenesis mengalami perubahan mengikuti siklus ranggah. Pada saat ranggah keras aktivitas reproduksi semakin meningkat, yang ditandai dengan terjadinya perkawinan antara rusa jantan dan betina (Lincoln 1985). Namun demikian pada beberapa spesies rusa lainnya ditemukan adanya variasi pada aktivitas perkawinan. Pada red deer (Cervus elaphus), aktivitas reproduksi dimulai dua bulan sebelum musim gugur berlangsung (Suttie dan Kay 1985). Sedangkan pada roe deer (Capreolus
capreolus), perkawinan lebih sering terjadi pada musim panas antara bulan Mei sampai Agustus (Sempere 1990; Blottner et al. 1996).
Pertumbuhan dan Siklus Ranggah
Ranggah merupakan satu-satunya organ pada mamalia yang mengalami perpanjangan dan mampu beregenerasi secara berulang. Setiap tahun ranggah lepas dan tumbuh kembali dari blastema dan selanjutnya bercabang. Ranggah terdiri atas kulit, syaraf, buluh darah, jaringan fibrosa, tulang rawan (kartilago) dan tulang keras. Struktur ranggah tersebut berbeda dengan tanduk pada ruminansia lain seperti kerbau dan kambing. Tanduk ruminansia merupakan jaringan yang mengalami keratinisasi yang tumbuh dibawah kontrol sel-sel mesenkim pada lapisan basal tanduk. Pada beberapa spesies rusa, ranggah dapat tumbuh sekitar 2 cm per hari, sehingga ranggah disebut sebagai organ yang mengalami pertumbuhan tercepat dibandingkan organ lainnya (Price dan Allen 2004; Price et al. 2005). Regenerasi ranggah bersifat tahunan dan terjadi secara berulang disepanjang hidup Cervidae jantan. Fenomena tersebut dipengaruhi oleh keberadaan testosteron dengan
konsentrasi berbeda sesuai dengan tahapan regenerasi ranggah (Suttie et al. 1992). Pada roe deer (Capreolus capreolus), pertumbuhan ranggah
berlangsung setelah bulan Januari di saat periode pencahayaan dan konsentrasi testosteron mulai meningkat. Namun pada sebagian besar Cervidae lainnya, proses tersebut terjadi pada musim panas (Sempere dan Boissin 1981).
Pertumbuhan ranggah pada Cervidae yang hidup di daerah beriklim tropis, sub tropis, dan beriklim sedang, terbagi atas empat tahapan pertumbuhan, yaitu tahap pedicle, tahap ranggah lunak (velvet), tahap ranggah keras (hard antler) dan tahap lepas ranggah (casting) (Fennessy dan Suttie 1985). Suatu siklus ranggah diawali dengan inisiasi pertumbuhan pedicle pada Cervidae jantan yang memasuki usia pubertas. Pada red deer (Cervus elaphus), inisiasi tersebut berhubungan erat dengan peningkatan konsentrasi testosteron plasma sebagai respon terhadap perubahan sekresi LH. Pertumbuhan ranggah pada jantan pubertas dan dewasa terjadi pada saat konsentrasi testosteron plasma menurun atau rendah yang disebabkan adanya mekanisme umpan balik negatif terhadap hipotalamus dan hipofise karena tingginya konsentrasi testosteron selama periode ranggah keras. Akibatnya sekresi GnRH dan LH berkurang (Gambar 5) (Fennessy dan Suttie 1985). Peningkatan testosteron plasma secara
signifik pada pada yang d Gamba pedicle usia p osifika diferen trabec transis Tahap antlero selanju osifika pedicle kan terjadi tahap castin beberapa sp dilaporkan p ar 5 Regula releasi velvet (RK). ( Analisis h e dan awal pubertas dije asi intramem nsiasi sel-se cular bone sional yang p ini ditan ogenik yang utnya berd asi endokon e. Sedangka pada saat p ing, konsent pesies rusa ada red dee
si hormonal ing hormone (RV), pengel (Modifikasi da histologi terh pertumbuha elaskan oleh mbran, dita el antlerogen di dalam p berlangsun dai dengan terdapat pa iferensiasi ndral yang an tahap ke proses peng trasi hormo temperate er (Price et a pertumbuha (GnRH), lute upasan kulit v ari Fennessy hadap form an ranggah Li dan Sutt andai denga nik menjadi periosteum. g pada saa n terbentuk ada permuka menjadi ko ditandai de empat adala gelupasan k n tersebut konsentrasi al. 2005). an ranggah einizing horm velvet atau sh dan Suttie 19 masi dan id pada Cerv tie (1994). T an dimulain osteoblas ya Tahap ked at panjang p knya jaring aan apikal. S ondrosit. Ta engan berla ah tahap os kulit velvet r sangat rend nya tidak te pada Cervid one (LH), cas hedding (S), d 985). dentifikasi ta idae jantan Tahap pertam nya proses ang selanjut dua adalah pedicle men gan kartilag Sel-sel antle ahap ketiga angsungnya ifikasi endok ranggah. Na dah dan ba erdeteksi, se ae. Gonadot sting (C), ran dan ranggah ahapan osif yang mema ma adalah t proliferasi tnya membe tahap osif ncapai 5-10 go oleh se erogenik ters a adalah t a kondrogen kondral rang amun hkan eperti tropin nggah keras fikasi asuki ahap dan entuk fikasi mm. el-sel sebut ahap nesis ggah.
Tahap ini merupakan tahap akhir yang ditandai dengan berlanjutnya proses osifikasi kondrogenik hingga terbentuk ranggah. Onset formasi ranggah ditandai dengan munculnya selaput tipis (velvet skin) yang menutupi bagian ujung distal pedicle.
Biosintesis dan Metabolisme Androgen
Pada hewan jantan terdapat tiga jenis hormon steroid yang berperan penting dalam sistem reproduksi, yaitu testosteron, dihidrotestosteron dan estradiol. Dari ketiga jenis hormon steroid tersebut, testosteron memiliki peran dominan. Sekitar 95% testosteron disintesis dan disekresikan oleh sel-sel Leydig tubuli semeniferi testis, sedangkan sebagian kecil lainnya disekresikan oleh kelenjar adrenal. Selain testosteron, sel-sel Leydig testis juga mensekresikan androgen lainnya, yaitu dehidrostestosteron (DHT), dehidroepiandrosteron (DHEA) dan androstenedion yang kurang memiliki efek androgen dibandingkan testosteron. Sel-sel Leydig juga menghasilkan beberapa hormon steroid lainnya dalam jumlah kecil, seperti estradiol, estron, pregnenolon, 17α-hidroksipregnenolon dan 17α-hidroksiprogesteron (Braunstein 1997).
Pada vertebrata, biosintesis hormon steroid seperti testosteron dan metabolitnya berasal dari kolesterol sebagai prekursor hormon (Koolman dan Röhm 2001). Biosintesis testosteron oleh sel-sel Leydig dapat berlangsung akibat stimulus luteinizing hormone (LH) yang dihasilkan oleh sel-sel LH di hipofise anterior. Hormon ini bekerja secara sinergis dengan follicle stimulating hormone (FSH) dan prolaktin. Peran prolaktin pada proses sintesis testosteron adalah dengan meningkatkan jumlah dan afinitas reseptor LH di membran sel Leydig. Adanya interaksi antara LH dan reseptornya akan mengaktifkan sistem adenilat siklase, termasuk aktivasi protein kinase dan sintesis RNA (Pineda 2003), sehingga terjadi peningkatan sintesis pregnenolon dengan jumlah rantai karbon 21 (C21) dari kolesterol (C27) oleh mitokondria sel-sel Leydig. Hasil akhir dari proses tersebut adalah testosteron (C19) (Koolman dan Röhm 2001; Pineda 2003).
Jalur biosintesis hormon androgen (Gambar 6) diawali dengan proses konversi kolesterol menjadi pregnenolon oleh enzim cytochrome P450 side-chain cleavage (P450scc) yang berlangsung di mitokondria sel Leydig. Pregnenolon selanjutnya berdifusi ke sitoplasma dan dikonversikan melalui jalur ∆5 atau ∆4 dengan produk akhir testosteron. Pada jalur ∆5, pregnenolon diubah menjadi 17α-pregnenolon oleh P450c17/C17 hidroksilase dan akan berubah menjadi
DHEA domin menja dehidr Jalur pregne terjadi andros testos Gamba A oleh P450c an dibandin di testost rogenase (1 ∆4 lebih d enolon men nya konve stenedion o teron sebag ar 6 Regulas hitam m testis, manusia Hikim c17/C17,20 l ngkan denga teron mela 17β-HSD) d ominan dite njadi proge ersi proge leh P450c17 gai produk ak si steroidoge menunjukkan garis hitam a. STAR = et al. 2005). iase/desmol an jalur ∆4. alui andro dan 3β-hid emukan pad steron oleh esteron m , sedangkan khir oleh 17β nesis androg proses yan tebal menun = steroidoge lase. Pada t Walaupun ostenedion roksisteroid da testis ro h 3β-HSD. enjadi 17α n androsten β-HSD (Hikim
gem oleh lut ng berlangsu njukkan jalur enic acute testis manus DHEA dap oleh 17 dehidrogen odensia den Tahap sela α-hidroksipro nedion akan m et al. 2005 teinizing horm ng di mitoko steroidogen regulatory p sia, jalur ∆5 at dikonvers β-hidroksist nase (3β-H ngan meng anjutnya ad ogesteron diubah me 5). mone (LH). K ondria sel L nik dominan protein (Sum lebih sikan eroid HSD). ubah dalah dan enjadi Kotak eydig pada mber:
Testosteron yang memasuki sirkulasi darah terikat pada albumin serum dan steroid hormone-binding globulin (SHBG). Senyawa ini merupakan suatu glikosilasi protein dimer yang terutama diekspresikan di hati dan selanjutnya memasuki sel secara difusi. Testosteron dapat dikonversikan menjadi 5α-DHT oleh dua tipe isoenzim mikrosomal, yaitu 5α-reduktase-1 dan 5α-reduktase-2. Enzim 5α-reduktase-2 akan mengkonversi testosteron menjadi 5α-DHT pada organ genital. Enzim ini dikode oleh gen SRD5A2 yang terdapat di kromosom 2 (2p23) dan hanya diekspresikan oleh jaringan genital yang bergantung pada androgen. Selain itu, testosteron juga dapat diaromatisasi menjadi estradiol-17β oleh enzim aromatase sitokrom P450 (P450arom) di jaringan perifer. Walaupun estradiol-17β merupakan hormon steroid utama pada betina, namun pada jantan hormon ini diperlukan dalam konsentrasi kecil pada proses spermatogenesis, libido, dan beberapa fungsi reproduksi lainnya (Meaden dan Chedrese 2009).
Menurut Hodges dan Heistermann (2003); Heisterman (2010), hormon steroid yang beredar di sirkulasi darah akan mengalami perubahan metabolik di hati atau ginjal sebelum dieliminasi oleh tubuh. Perubahan yang terjadi secara alami bervariasi di antara spesies hewan, baik dari jenis metabolit maupun jalur ekskresinya. Informasi yang diperoleh dari variasi tersebut, pada tahap berikutnya akan bermanfaat untuk memilih teknik pengukuran metabolit hormon steroid dan interpretasi hasil yang akurat. Metabolisme hormon steroid seperti androgen secara mayoritas berlangsung di hati, namun beberapa aktivitas katabolik juga berlangsung di ginjal. Metabolit steroid yang dihasilkan akan diekskresikan oleh ginjal melalui urin, sedangkan sistem empedu mengekskresikan metabolit melalui feses (Amaral et al. 2009).
Hormon androgen yang beredar di sirkulasi darah akan dikonversikan oleh hati menjadi beberapa metabolit androgen, seperti androsteron dan etiokolanolon. Metabolit androgen tersebut selanjutnya akan terkonjugasi dengan glukoronik dan asam sulfat sebelum diekskresikan melalui urin sebagai 17-ketosteroid (sekitar 20-30%), sedangkan sisanya merupakan hasil metabolisme steroid kelenjar adrenal (Meaden dan Chedrese 2009).
Metode Pengukuran Konsentrasi Metabolit Steroid
Penentuan konsentrasi hormon reproduksi jantan (androgen) dapat dilakukan dengan berbagai cara, seperti pemeriksaan hormon dari plasma (Morais et al. 2002), urin dan feses (Busso et al. 2004). Teknik pemeriksaan rutin melalui plasma yang diperoleh dari darah juga dapat dilakukan pada hewan
ternak, namun teknik tersebut sedikit berisiko bila dilakukan pada satwa liar, karena harus melewati prosedur imobilisasi dengan bahan kimia dan restrain fisik (Hamasaki et al. 2001). Oleh karena itu perlu dilakukan teknik pemeriksaan lainnya yaitu dengan teknik non invasif. Teknik ini sangat sesuai bila diaplikasikan pada satwa liar untuk tujuan penelitian jangka panjang (longitudinal study), baik di lingkungan in situ maupun ex situ. Dengan pendekatan non invasif, hewan terhindar dari stres akibat proses restrain yang biasa dilakukan pada teknik invasif. Teknik non invasif dapat diaplikasikan untuk menganalisis metabolit hormon yang terdapat pada feses dan urin (Mateo dan Cavigelli 2005; Washburn et al. 2004). Poole et al. (1984), menggunakan teknik tersebut untuk menentukan konsentrasi testosteron dari sampel feses dan urin gajah liar afrika (Loxodonta africana), Hamasaki et al. (2001) pada sika deer (Cervus nippon), Morato et al. (2004) pada jaguar (Panthera onca), dan Mauget et al. (2007) pada chinese water deer (Hydropotes inermis)
Pada tahap berikutnya penelitian mengenai teknik ini terus berkembang, terutama yang berhubungan dengan improvisasi metode analisis, transportasi dan purifikasi sampel (Ziegler dan Wittwer 2005). Proses validasi monitoring non invasif terhadap variasi metabolit androgen yang terdapat di dalam ekstrak feses juga telah dilakukan pada beberapa spesies satwa liar, seperti spotted hyena (Crocuta crocuta) (Dloniak et al. 2004) dan Gajah Afrika (Loxodonta africana) (Ganswindt et al. 2005). Pada satwa betina aplikasi teknik ini telah dilaporkan untuk menentukan siklus ovariumnya, seperti pada badak putih (Ceratotherium simum simum) (Patton et al. 1999; Schwarzenberger et al. 1998), schimitar horned-oryx (Oryx dammah) (Morrow dan Monfort 1998) dan okapi (Okapia jhonstoni) (Kusuda et al. 2007).
Ada beberapa tahapan yang dilakukan untuk menganalisis profil hormon reproduksi dengan memanfaatkan sampel feses, yaitu: 1) koleksi sampel feses dari lapangan, 2) penanganan dan persiapan sampel feses, dan 3) analisis laboratorium. Sebelum sampling dilakukan, perlu diketahui kondisi lapangan tempat sampel berada dan jumlah feses yang akan dikoleksi. Hal tersebut berkaitan dengan proses transportasi sampel dari lapangan hingga tahap analisis laboratorium dilakukan (Ziegler dan Wittwer 2005). Proses penyimpanan sampel sangat menentukan kualitas hormon yang akan dianalisis. Metabolisme bakteri gastrointestinal yang terdapat di feses secara cepat akan merusak hormon yang diekskresikan (Heistermann 2010). Metode terdahulu yang dilakukan untuk
sampling feses di lapangan adalah menyimpan sampel menggunakan alkohol sebagai bahan preservatif hingga tahap analisis dilakukan. Namun metode ini memiliki kelemahan, yaitu dapat menyebabkan terjadinya perubahan struktur
steroid hormon akibat terbatasnya udara pada saat transportasi (Khan et al. 2002). Untuk itu, telah dilakukan beberapa metode penyimpanan
yang dapat mempertahahankan kondisi sampel yang dikoleksi, seperti pembekuan sampel (freezing samples), dan pengeringan sampel (drying samples). Proses pengeringan (lyophilization) sampel feses merupakan tahapan penting untuk mengawetkan metabolit hormon steroid agar dapat disimpan dalam waktu lama, sehingga konsentrasi metabolit steroid yang akan diperiksa tidak berubah secara signifikan (Hunt dan Wasser 2003). Tahap berikutnya adalah ekstraksi sampel di yang dilanjutkan dengan tahap analisis laboratorium. Sampel feses yang telah diekstraksi dapat dianalisis secara langsung dengan teknik enzymeimmunoassay menggunakan antibodi spesifik terhadap hormon yang akan dideteksi (Ziegler dan Wittwer 2005).
Pengukuran hormon dan metabolit hormon biasanya dilakukan dengan prosedur imunologi menggunakan antibodi hormon atau antibodi grup spesifik hormon. Ada dua metode immunoassay yang tersedia, yaitu radioimmunoassay (RIA) dan enzyme immunoassay (EIA). Metode RIA dilakukan dengan melabel hormon dengan marker radioaktif tertentu, sedangkan EIA menggunakan preparasi enzim atau enzim yang dilabel dengan biotin. Metode EIA lebih dipilih karena tidak menimbulkan efek radioaktif seperti RIA. Selain itu biaya asai lebih murah dibandingkan dengan RIA (Hodges dan Heistermann 2003). Aplikasi metode EIA terhadap metabolit androgen feses dari spesies rusa telah dilaporkan pada pampas deer (Ozotoceros bezoarticus bezoarticus) (Pereira et al. 2005), dan red brocket buck (Mazama americana) (Versiani et al. 2009). Meode tersebut telah terbukti sebagai metode yang tepat
dan praktis untuk mengetahui status reproduksi pada kedua spesies rusa tersebut.
Anatomi Organ Reproduksi Jantan
Anatomi organ reproduksi jantan bervariasi pada berbagai spesies hewan. Variasi tersebut berhubungan erat dengan fungsi reproduksinya serta penyesuaian terhadap anatomi organ reproduksi betina. Secara umum, organ reproduksi jantan terdiri atas: sepasang testis dan saluran-salurannya: duktus epididimidis dan duktus deferens yang menyalurkan produk eksokrin testis
menuju uretra (Colville dan Bassert 2002). Selain itu terdapat pula kelenjar asesoris kelamin yang terdiri atas: glandula vesikularis, glandula prostata dan glandula bulbouretralis (Tabel 1). Kelenjar asesoris kelamin tersebut berperan sebagai organ yang menghasilkan bahan pembentuk plasma semen (Haron et al. 1999). Uretra adalah saluran reproduksi jantan yang berjalan di sepanjang penis dan berfungsi untuk menyalurkan urin dan semen. Sedangkan penis merupakan organ kopulasi jantan yang menyalurkan semen ke dalam organ reproduksi betina saat terjadi ejakulasi (Senger 2005).
Tabel 1 Kelenjar asesoris kelamin pada beberapa spesies hewan.
Spesies Glandula Vesikularis Glandula Prostata Glandula Bulbourethralis
Sapi + + + Anjing - + - Kuda + + + Kucing - + + Llamas - + + Domba + + + Babi + + + (Sumber: Pineda 2003).
Secara anatomi testis berada di luar tubuh dan dilindungi oleh kulit tebal yang disebut skrotum. Testis merupakan organ elipsoidal padat dengan ukuran bervariasi pada setiap spesies hewan. Pada kucing, ukuran testis relatif lebih kecil bila dibandingkan domba dan kambing. Selain itu orientasi testis pada tubuh hewan juga bersifat spesifik spesies, seperti testis sapi yang menjulur kearah vertikal dari sumbu tubuh, testis kuda dan anjing pada posisi horizontal, sedangkan testis babi dan kucing berada di bagian ventral anus (Senger 2005). Testis dexter et sinister digantung dan dipisahkan di dalam skrotum oleh funikulus spermatikus yang terdiri atas duktus deferens, buluh darah (arteri dan vena) dan syaraf serta dibungkus oleh peritoneum. Dari arah superfisial ke profundal, testis dilapisi oleh skrotum, tunika dartos, fasia spermatikus eksterna, tunika vaginalis lamina parietalis dan tunika vaginalis lamina viseralis, serta tunika albuginea (Dyce et al. 1996). Mayoritas jaringan parenkim testis ditempati oleh tubuli seminiferi. Komposisi jaringan parenkim testis babi terdiri atas 87% tubuli seminiferi dan 7% sel sel Leydig dengan jumlah sel sertoli sekitar 42 juta per gram testis (Almeida et al. 2006).