The developmental transition to flowering represses ascorbate
peroxidase activity and induces enzymatic lipid peroxidation in
leaf tissue in
Arabidopsis thaliana
Zhenzhen Ye, Roxana Rodriguez, Augustine Tran, Hoang Hoang,
Darryn de los Santos, Shawn Brown, Robert Luis Vellanoweth*
Department of Chemistry and Biochemistry,California State Uni6ersity Los Angeles,Los Angeles,CA90032-8202,USA
Received 7 February 2000; received in revised form 31 May 2000; accepted 3 June 2000
Abstract
Leaf senescence in many plant species is associated with increased oxidative damage to cellular macromolecules by reactive oxygen species (ROS). Since ROS levels and their damage products in many plants are known to increase during senescence, it is possible that these changes are due to a decline in the levels of certain antioxidant enzymes. Using specific assays, we find that the developmental transition to bolting and flowering is associated with up to a 5-fold decline in ascorbate peroxidase activity and an increase in chloroplastid superoxide dismutase. As expected, these changes are associated with a measured increase in lipid peroxidation products. By HPLC separation of the products, we identified the different positional isomers and find that stereospecific lipid peroxidation occurs after the bolting transition. The product distribution suggests that enzyme-mediated lipid peroxidation, via a lipoxygenase, is responsible for the observed increase. Surprisingly, though consistent with the known induction of antioxidant defenses by hydrogen peroxide, the activity of APX rebounds with further development (reproduction and seed setting) and this increase (up to 5-fold) is associated with declines in lipid peroxidation and with the onset of visible senescence symptoms. Thus, in Arabidopsis, ROS increases are associated with the developmental transition to flowering, perhaps due to programmed declines in APX activity, and apparently lead to the oxidative activation of lipoxygenase and subsequent lipid peroxidation. The reactivation of APX at later stages appears to help reduce the lipid peroxidation rate, although the senescence program continues unabated. © 2000 Elsevier Science Ireland Ltd. All rights reserved.
Keywords:Ascorbate peroxidase; Lipid peroxidation; Reactive oxygen species; Senescence; Arabidopsis
www.elsevier.com/locate/plantsci
1. Introduction
During the final developmental phase in the life of an annual plant, reproduction and seed devel-opment are accompanied by senescence of the leaves. This genetically programmed aging event is an active process characterized by the degradation of leaf cell components. Metabolism in the leaves is drastically altered. During vegetative growth, the primary function of the leaf is to provide fixed carbon to the rest of the plant through the process
of photosynthesis. However, as senescence pro-ceeds, leaf photosynthetic capacity is diminished through a destruction of chlorophyll, a proteolytic attack on the abundant CO2-fixing enzyme
ribu-lose bisphosphate-1, 5-carboxylase, and a decline in de novo synthesis of the photosynthetic appara-tus [1]. Protein and nucleic acid levels decline through enhanced proteolysis and nuclease attack and membrane integrity is lost. A critical role for reactive oxygen species (ROS) such as superoxide anion (O2−), hydrogen peroxide (H2O2), and
hy-droxyl radical (HO) throughout all stages of senescence has been implicated in many plants, with the primary target of damage being the mem-brane lipids [2]. Although apparently a destructive
* Corresponding author. Tel: +1-323-3432148; fax: + 1-323-3436490.
E-mail address:[email protected] (R.L. Vellanoweth).
process, the onset of senescence requires the active transcription and translation of new gene products and is thus under direct genetic control. It is definitely not a simple case of down-regulation of genes that define a juvenile state, though such repression forms part of the process, but rather progression into this final life phase requires the expression of new genes [3 – 5]. Some of these senescence-associated genes have begun to be elu-cidated, and the known products of the more abundantly expressed genes are those expected: nucleases [6,7], proteases [3,4], and nitrogen reas-similation products [8], as well as some surprises [9]. The primary adaptive reason for the induced degradation is to mobilize leaf nitrogen for trans-port to the developing seeds. With the induction of the senescence program, the leaf no longer pro-vides new fixed carbon to the rest of the plant and thus senescence involves a redistribution of re-sources from somatic tissue to the developing progeny, a perfect example of parental altruism. Several models have been put forth to explain the highly ordered and controlled nature of leaf senescence. In many leguminous plants, the aging program appears to be controlled by the develop-ing seeds [10]. Removal of floral structures extends the longevity of the leaves, suggesting the involve-ment of a diffusible substance that initiates leaf senescence. The nature of this diffusible regulator is still unknown. Plant-specific hormones also ap-pear to institute senescent phenotypes in many systems. Since most studies using plant hormones involve their exogenous addition to various or-gans, attached or detached, in many instances their endogenous role remains unclear. A common molecular theme in plant senescence is evident in more downstream events, however. In most spe-cies examined, ROS and their damage products increase during leaf aging. Thompson has sug-gested that O2− levels rise in senescing plant
tis-sues due to an increase in lipoxygenase activity, placing ROS production and damage at the same location, cellular membranes [2]. Others have sug-gested, both in plants [11] and in animals [12] that age-related ROS increases result from leaks in the normal electron transfer reactions of metabolic pathways. Whatever their source, ROS are inti-mately associated with the degradative aspects of senescence physiology in all plants that have been studied.
In this report, we test the hypothesis that age-dependent increases in ROS are the result of a programmed downregulation of antioxidant en-zymes in Arabidopsis. Given the increasingly im-portant role of H2O2as a signaling molecule, both
in plant and animal systems, we decided to focus on those enzymes that can modulate H2O2 levels.
We show that the transition from vegetative growth to reproductive growth (bolting) is related to the longevity of rosette leaves as measured by chlorophyll level and that this transition is associ-ated with ascorbate peroxidase (APX) decline and lipoxygenase (LOX) activation.
2. Materials and methods
2.1. Plant material and growth conditions
Arabidopsis thaliana (Columbia) seeds (Lehle Seeds, Inc) were sown in 2:1:1 potting soil:perlite:vermiculite following a 5-day imbibed incubation in 0.1% agarose at 4°C. Nearly syn-chronous germination was encouraged by a 3-day incubation of the pots at 4°C before transfer to growth chambers. The plants were irrigated weekly with a nutrient solution [13] and kept in a growth chamber or environmental room at 23°C under a 12 h photoperiod with a light fluence of 250 mmole/m2 per s. Plants were evenly spaced
every 4 cm to encourage similar growth rates and sizes.
2.2. Extract preparations and enzyme acti6ity
assays
Extracts were prepared by freezing leaf tissue in liquid N2and pulverizing to a fine powder before
phosphate buffer (pH 7.0) with a constant current of 23 mA per gel. SOD activity was determined by the negative staining procedure using nitroblue tetrazolium. For SOD isozyme identification, the gels were incubated prior to activity staining in 5 mM H2O2 or 2 mM KCN. Of the three isozymes
that may be present, the Mn form is insensitive to either treatment, while the CuZn isozyme is sensi-tive to CN−. The Fe isozyme is sensitive to H
2O2
[16]. APX activity was determined as described [14]. This assay is based on the ability of ascorbate to reduce NBT, in the presence of tetramethylethylene-diamine to formazan. Ascorbate peroxidase pre-vents the formation of formazan in the presence of H2O2, and thus the ascorbate peroxidase activity is
seen as an achromatic band on a purple-blue background. Activities were quantified using Molecular Analyst software (BioRad, Inc.) on dig-ital images recovered from a CCD camera driven GelDoc 1000 system. The linear range of activity was determined by loading increasing amounts of cell extract on the gels and comparing the pixel density of the bands with the amounts loaded.
2.3. Lipid peroxide and chlorophyll measurements
Lipids were extracted from growing plant rosette leaves at various stages following Bligh and Dyer’s method [17]. Lipid peroxides were quantified by the FOX method [18] using the PeroXoquant™ Quan-titative Peroxide Assay from Pierce Chemical Co. Samples (50 ml) were incubated with 500 ml of
working reagent which consists of 1 volume of 25 mM of Fe2+ and 2.5 M H
2SO4, and 100 volumes
of 4 mM BHT and 125 mM xylenol orange in
methanol. After a 20 min incubation at room temperature, the absorbances at 560 nm were read on an HP 8452A diode array spectrophotometer. Freshly prepared hydrogen peroxide solutions were used as standard.
A portion of total lipid extracts was used for chlorophyll determination. The lipid extracts were diluted withN,N-dimethylformamide (DMF). Ab-sorbances at 648 and 664 nm were recorded using DMF as blank. The total chlorophyll content was calculated as described [19].
2.4. Lipid peroxide characterization
Standards were prepared by autoxidation of linoleic and linolenic acids and subsequent
reduc-tion to hydroxyl forms. In 10 ml of 0.2 mM tBuOOH in 0.5% ethanol, linoleic or linolenic acid was dissolved at 4 mM final concentration. The autoxidation was initiated by adding FeSO4to 0.2
mM. The reaction proceeded for 10 min at room temperature. Reduction was fulfilled by introducing a molar excess of NaBH4 in 5 N NaOH. The
reaction mixture was acidified to pH 4 with 5 N HCl and extracted by 1 ml hexane: diethyl ether (70:30, v/v). After centrifugation, the upper layer was collected and used for HPLC injection.
Hydroperoxy fatty acids were isolated and re-duced to their corresponding hydroxyl forms ac-cording to Degousee et al. [20]. Briefly, fatty acids from Arabidopsis were prepared by homogeniza-tion of 0.5 g of rosette leaves in 1 ml 5%(w/v) NaBH4 in 0.2 N NaOH. After acidifying the
ho-mogenate to pH 4 with 70% HClO4, 2.2 ml of
CHCl3: CH3OH (50:50, v/v) were used to separate
aqueous and organic phase. The lower phase was collected after centrifugation for 5 min at 11 000 rpm 4°C. The upper phase was further extracted by 1.1 ml chloroform. All lower phases were combined following a 5 min 11 000 rpm 4°C centrifugation. Solvent was removed and the residue dissolved in 0.5 ml of absolute ethanol and 0.5 ml of 3.5 N NaOH. Following a 15 min reflux and cooling, 0.5 ml 3.5 N HClO4was used to acidify sample to pH
4. Phases were then separated with 0.3 ml hexane: diethyl ether (70:30, v/v). The upper layer was collected after a 10 min 10 000 rpm centrifugation at 4°C. After filtering through a 0.45mm NY PP
filter, the sample was used for HPLC analysis. Isolated leaf fatty acids were characterized by analytical HPLC using a Rainin Dynamax instru-ment equipped with a 4.6×250 mm Microsorb-MV column packed with 5 mm silica gel. Mobile phase
solvent consisted of hexane: diethyl ether: acetic acid (70:30:0.5, v/v/v). Hydroxyl fatty acid stereoisomers were separated by isocratic elution at a flow rate of 0.5 ml/min. Absorbance at 234 nm was monitored with a photodiode array PDA-2 detector.
3. Results
3.1. Antioxidant enzyme acti6ities during the
Arabidopsis life span
the progress of leaf senescence [2]. Their buildup is purported to lead to membrane leakage and
organellar disintegration during late stages of senescence. It is possible that a programmed downregulation of antioxidant capacity might contribute to the observed ROS increase. We focused our studies first on enzyme activity, as opposed to mRNA levels, since it is at this level that changes would affect ROS metabolism. We thus prepared enzyme extracts from Arabidopsis leaves at each important stage in the plant’s life history and measured the activities of two antioxidant enzymes. The stages included pre-bolting, i.e. before the transition from the vegetative (juvenile) phase to the reproductive stage, short bolt (B1 cm), medium bolt (\4 cm),
long bolt (\10 cm), flowering, seeding, and
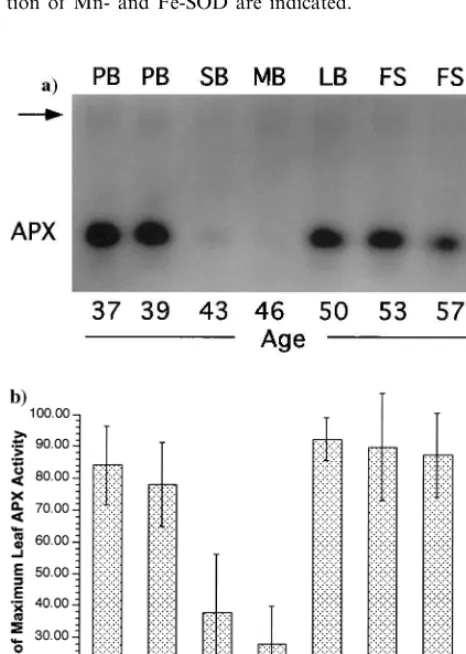
senescing (with rosette leaves at least 25% yellow). Equal amounts of protein isolated from the various stages were loaded onto a native PAGE gel and stained for SOD activity. An inverted image of one such gel is shown in Fig. 1. Slight variations in Mn-SOD activity as the life span progressed were evident in this example, while Fe-SOD consistently displayed an increase at the mid-bolt stage followed by a return to pre-bolt levels prior to senescence. The situation was even more dramatic for ascorbate peroxidase (APX). Fig. 2a shows the results of a similar experiment for APX activity. The major band for APX activity declined precipitously as the plants entered the reproductive phase of life. Thus, from pre-bolt (39 days after planting) to short bolt (43 dap) APX levels declined more than fivefold. The decline was only transient, however, as the APX level rebounded to pre-bolt levels at the long bolt stage (50 dap). APX levels were then found to vary only slightly as the plants continued their life span through senescence. Data from six separate experiments were compiled and are shown in Fig. 2b. The APX activity at each stage is reported as the six-experiment average of the % of the highest activity measured within any series. The standard deviation of the data is shown as error bars in the figure. The transition to bolting at the meristem was associated with an average two to threefold decrease in leaf APX activity. A very careful selection of plants at identical developmental stages showed that this decline can reach fivefold (Fig. 2). In all cases, the APX level subsequently increased to the highest levels measured as the bolts reached lengths of 10 cm. This dip and subsequent rebound of APX activity occurred
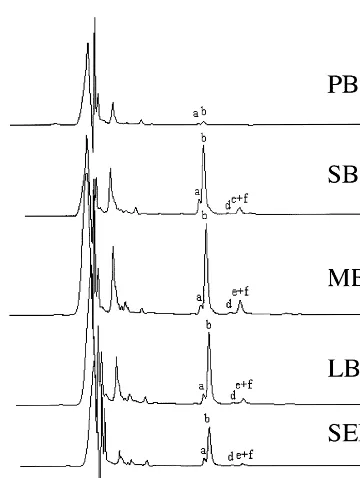
Fig. 1. Leaf superoxide dismutase activity as a function of life history stage. Inverted image of a native PAGE gel stained for SOD activity. Extracts were prepared from plants of the indicated age in days and the following stages: PB, pre-bolt; SB, short bolt; MB, medium bolt; LB, long bolt; FS, flower-ing and seedflower-ing; and SN, senescflower-ing. The positions of migra-tion of Mn- and Fe-SOD are indicated.
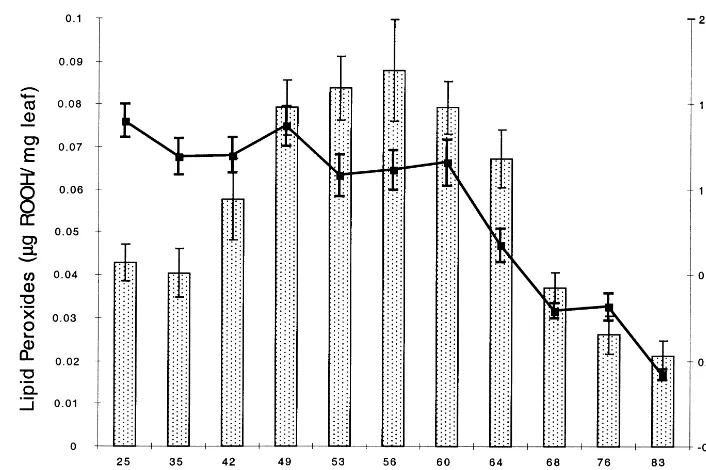
Fig. 3. Leaf lipid peroxide level as a function of life history stage. Lipid peroxides (columns) were measured using the FOX assay and are expressed asmg of ROOH per mg of leaf tissue. The curve indicates the level of chlorophyll at the indicated stages. SN1,
leaf was approximately 20% yellow; SN2, 40% yellow; SN3, 60% yellow; SN4 80% yellow; and SN5, 100% yellow.
within a span of 10 days, or roughly one-eighth the total life span. As the plants entered senes-cence, only a slight decrease in APX was mea-sured. Thus, in Arabidopsis it appears that APX activity does decline in an apparently programmed manner, but its timing precedes considerably the onset of visible symptoms of senescence.
3.2. Leaf lipid peroxide le6els as a function of life
span
The primary purpose of leaf APX is to scavenge hydrogen peroxide before it can react with cellular biomolecules and cause damage. Our observations that leaf APX declines as Arabidopsis plants enter the reproductive phase suggests that H2O2 levels
might increase during the 10-day period of low activity and lead to biomolecular damage, a hall-mark of senescence in virtually all species [21]. To test this possibility, we measured the levels of whole-cell lipid peroxides at the same stages used above. Total lipid was extracted and lipid perox-ides were determined using the ferrous oxidation-xylenol orange (FOX) method [18]. This assay method was used due to its higher specificity to-wards lipid peroxides than the TBARS test. For each stage, only the 5th and 6th leaves to emerge from the rosette were used for extract prepara-tions. Twenty samples were collected for each
stage in order to study the deviation in the data. The data in Fig. 3 show that lipid peroxides began to increase at the bolting transition, as expected from the observed decline in APX, and reached a maximum twofold higher level by the flowering stage. As the plants entered senescence though, indicated by the decline in leaf chlorophyll at 64 dap, lipid peroxide levels also declined. By the final stage of senescence (83 dap, leaves completely yellow but not dry), lipid peroxides were even lower than at the young, pre-bolt stage. Since the data are reported as mg lipid peroxide per mg of
leaf tissue, the level of lipid peroxides on a per cell basis would be even lower since leaf mass had declined due to senescence processes. The APX decline was therefore associated with an increase in lipid peroxidation during the 10-day period of low APX. Lipid peroxides remained elevated, however, for a further 10 – 15 days, suggesting that some mechanism other than simple chemical at-tack of H2O2 on membrane lipid double bonds
was responsible for the elevated lipid peroxide levels.
3.3. Isomeric composition of leaf lipid peroxides
The reaction of H2O2 with plant membrane
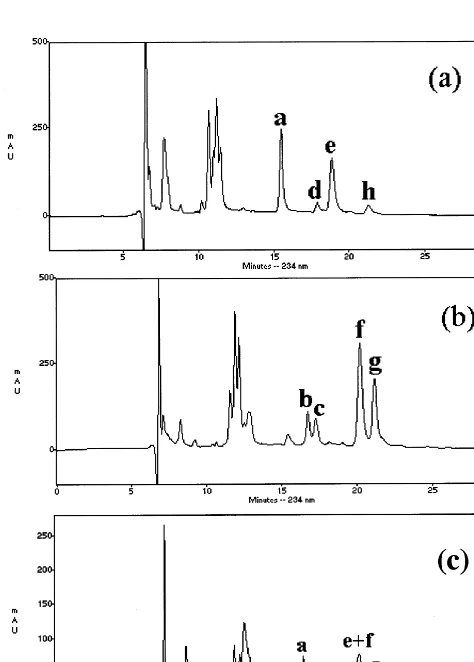
added peroxy group on the fatty acid chain and in the number of double bonds and chain length reflecting the natural diversity of cellular lipids. The major lipid fraction in Arabidopsis leaves is composed of linoleic and linolenic acids, however [22]. Thus, a determination of the peroxy position in cellular lipids is manageable by HPLC isomeric composition analysis. A set of standards was first prepared by autoxidation and enzymatic genera-tion of lipid hydroperoxides to identify the peaks on the chromatogram. Fig. 4a and b show results from t-butly hydroperoxide-catalyzed autoxida-tion of linoleic and linolenic acid, respectively,
while a mixture of the two is shown in Fig. 4. The conjugated diene of the reduced products was followed by A234. Compared with published work
[20,23] under similar HPLC conditions, the peaks were identified and are listed in the Fig. 4 legend. The amounts of the positional isomers were not equal. Thus, 13-HODE and 9-HODE represented about 85% of the four identified isomers from linoleic acid while 16-HOTE and 9-HOTE ac-counted for about 75% of the four isomers from linolenic acid. Other peaks that eluted prior to 12 min were not identified but were also present in the pre-autoxidation material. Standards were also prepared from enzymatic peroxidation of linoleic acid using soybean lipoxygenase (LOX) and sub-jected to HPLC separation. Unlike autoxidation, 13-HODE was the predominant product with more than 90% of the total as shown in Fig. 5. Next, lipid peroxides from different-aged Ara-bidopsis leaves were prepared and analyzed by HPLC following reduction to their corresponding hydroxyl forms. The chromatographic profile (Fig. 6) between 15 and 25 min showed considerably less complexity in the leaf-derived sample than the autoxidation standards (Fig. 4a and b) and was rather similar to the enzymatically generated stan-dard (Fig. 5). In the natural sample, the second peak, 13-HOTE, showed the greatest increase in amount following the transition from a pre-bolted stage to the reproductive phase. The level of 13-HOTE slowly began to decrease as the plants entered senescence, paralleling the kinetics of lipid peroxide changes found by the FOX assay (Fig. 3). In addition to the major increase in 13-HOTE, smaller increases were observed for 13-HODE, 13-HODE trans, 16-HOTE, and 9-HODE. These isomers followed the same kinetics as 13-HOTE and thus contributed to the overall pattern of lipid peroxidation changes during the life span.
3.4. Leaf longe6ity and flowering
We have made casual observations in our lab that the timing of the developmental transition to the reproductive state was associated with the timing of visible senescence symptoms in leaves. We have observed this phenomenon in the Colum-bia ecotype, whereas others have reported that late-flowering mutants exhibit wild-type leaf senes-cence timing in the Landsberg ecotype, suggesting a lack of association between flowering and
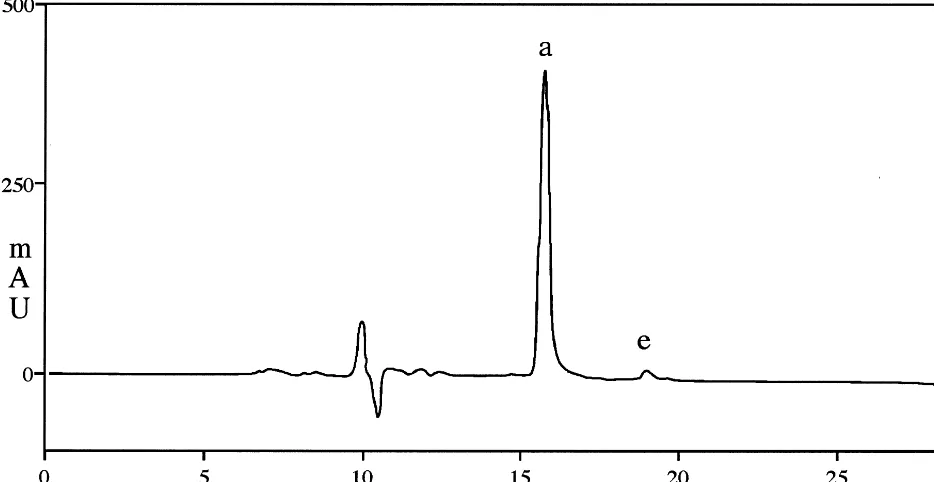
Fig. 5. HPLC separation of products generated by incubation of linoleic acid with soybean lipoxygenase and reduced to their hydroxy derivatives. The products indicated by letter are: a, 13-HODE; and e, 9-HODE.
4. Discussion
In the present investigation of antioxidant de-fense enzyme activities during the Arabidopsis life span, we have evidence for a developmentally regulated depression of leaf APX activity as plants enter the reproductive state (Fig. 2). The results show that APX activity is dramatically decreased in activity just after bolting. At least seven APX isozymes are known in Arabidopsis [25]. Because of our extraction method, we expect that the one band we detect on the APX activity gel is at least one of the cytoplasmic types, of which there are two. The other forms, two chloroplastic and three membrane bound, would be expected to be found in the membrane fraction, although we cannot rule out the possibility that the chloroplast forms con-tribute to the observed band. The mitochondrially localized Mn-SOD, exhibits only minor changes in activity throughout the life span, displaying a con-sistent slight increase at later stages. The chloro-plastid Fe-SOD, on the other hand, increases several-fold at the mid-bolt stage and declines prior to senescence. Interestingly, although cer-tainly expected considering the role of H2O2in the
induction of antioxidant defense genes [26], the activity of APX rebounds with further develop-ment (reproduction and seed setting) and this in-crease (up to fivefold) precedes the onset of visible senescence symptoms. The mechanism of APX decline is unknown, but considering that APX mRNA levels are known to be induced by oxida-tive stress, a transcriptional mode is likely. We carried out an add-back experiment looking for
evidence of an APX inhibitor in the short bolt extracts, but found none (data not shown). Of course, many assumptions are made in such an experiment and a negative result does not rule out the possibility of a specific APX inhibitor. An action directly on the protein is possible, though, as at least two reports have suggested that APX activity can be altered without affecting APX mRNA levels [27,28] and the APX mRNA is
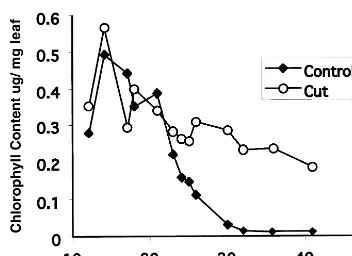
Fig. 7. Leaf chlorophyll level versus days after 5th leaf emergence. Bolting occurred at 15 days. Chlorophyll was measured spectrophotometrically following DMF extraction of leaf material. Control plants were allowed to flower and set seed normally while ‘Cut’ plants had their bolts continuously removed as they appeared. The standard deviation of the data was less than 10%.
tosolic APX levels remain fairly high, perhaps keeping H2O2 levels in check. The situation may
differ in the mitochondria, however, as it was recently reported that mitochondrial APX levels decline during pea leaf senescence [30]. In our study, the maximum lipid peroxidation rate is achieved by the long bolt stage when flowers have formed and seeds are setting. This stage is also when APX activity has returned to a level similar to that of the pre-bolt stage. For at least six more days, total lipid peroxide is elevated before it begins to fall. The delay suggests that H2O2, which
presumably goes unscavenged during the APX activity dip, is not the sole source of lipid peroxi-dation. The complete characterization of the lipid peroxides by HPLC (Fig. 6) revealed that the predominant product during this period is 13-HOTE, not the racemic mixture of all isomers expected from random, nonspecific chemical at-tack by H2O2. This suggests that a specific enzyme,
a 13-LOX specific for linolenic acid, is activated at the same time that APX levels decline and cata-lyzes the peroxidation observed. Thus, in Ara-bidopsis rosette leaves, ROS increases are associated with the developmental transition to flowering, due to programmed declines in APX activity, and may lead to the oxidative activation of a linolenic acid-specific 13-LOX and subsequent lipid peroxidation. The reactivation of APX at later stages coincides with a reduced lipid peroxi-dation rate, although the senescence program con-tinues unabated.
A fundamental feature of plant lipoxygenases is their conversion from an inactive form, where the non-heme Fe center is in the high-spin ferrous state, to an active form, where the Fe is oxidized to the ferric state. This Fe oxidation can occur by reaction with H2O2or organic peroxides [31,32]. It
thus seems plausible that the decline in APX al-lows an increase in H2O2 concentration that
acti-vates a pre-existing LOX to attack membrane-resident fatty acids, generating the ob-served increase in 13-HOTE. We quantified all hydroperoxy isomers over three generations of plants and considered 13-HODE and 13-HOTE to be enzymatically derived (although some small fraction is undoubtedly of non-enzymatic origin) and all other isomers to be non-enzymatically derived. As a function of life history stage, the ratio of enzymatic to non-enzymatic lipid peroxi-dation changed considerably. In the pre-bolt stage, somehow prevented from being translated during
the hypersensitive response [29].
Since the substrate for APX is H2O2, we
ex-pected that peroxidation reactions would increase from substrate buildup during this transitory, low APX period. This assumes, of course, a limited role for catalase in controlling H2O2 levels in the
cytoplasm. Most of the catalase activity is local-ized to glyoxysomes and peroxisomes, while APX is the major H2O2 scavenger in the cytosol and
cy-the ratio was approximately one. However, just after the bolting transition, the ratio increased to 4 – 6. As development proceeded, the ratio declined to 2 – 3. The even distribution of isomers in the pre-bolt stage suggests that a substantial propor-tion of lipid peroxidapropor-tion during vegetative growth is from random chemical attack. LOX-catalyzed peroxidation predominates after the bolting transi-tion and remains the major source of peroxides throughout the subsequent life of the plant. This regulated biochemical event may initiate the syn-thesis of a lipid derived secondary messenger. Jas-monic acid is one such wound-response messenger that is purported to be involved in senescence and its synthesis via the octadecanoate pathway is commenced with 13-lipoxygenase, apparently the same that is activated in response to bolting [33]. Other LOX-derived volatiles have also been demonstrated, including a number of six-carbon compounds that appear to mediate a wound re-sponse [34]. We do not believe that the peroxida-tion observed in this study is a simple, artifactual wound response resulting from the preparation of the extracts. The leaves were immediately trans-ferred to liquid N2 after harvesting, pulverized,
and extracted with organic solvents. Thus any wound response would be terminated or prevented from initiating. In addition, all extracts were pre-pared in exactly the same manner, but peroxida-tion was only observed at those stages where APX activity had declined. Thus, it appears from the presence of 13-HOTE that a LOX enzyme is acti-vated in a developmental manner, coincident with the programmed APX inactivation, and leads to lipid peroxidation as the plant enters the reproduc-tive phase of life.
Like most organisms, the controlling factors determining longevity in Arabidopsis are un-known. A number of studies suggest that the primary regulator of Arabidopsis leaf senescence is something other than ethylene or seed develop-ment. The senescence-promoting hormone ethyl-ene does not appear to be the dominant factor in leaf longevity since an analysis of ethylene-insensi-tive mutants reveals that senescence still proceeds, though slightly delayed [35 – 37]. While late-flower-ing mutants have been suggested to exhibit in-creased longevity of rosette leaves, Hensel et al. [3] have reported that in Landsberg erecta this is simply due to an increased number of leaves in such mutants and that the longevity of individual
by time and development. As previously suggested by others [3,36], an endogenous, age-related signal appears to initiate the natural senescence program in Arabidopsis leaves. Our results in this paper further suggest that a particular developmental stage, the floral transition, contributes to senes-cence initiation. Considering the involvement of ROS in the generation of senescence physiology in many other species, it is possible that the increase in ROS, caused by the bolting transition-induced depression of APX activity, serves as the develop-mental signal to initiate the expression of the senescence program. It is interesting to note that the late-flowering Arabidopsis mutant Gigantea
exhibits both increased leaf longevity and in-creased tolerance to oxidative stressors [39], due apparently to elevated levels of antioxidant en-zymes. The increased antioxidant capacity of this mutant may interfere with the programmed de-pression of APX activity we find with wild-type plants.
Precedence for ROS control of gene expression is found in the plant hypersensitive response (HR) to pathogenic attack. An incompatible interaction of an avirulent pathogen (bacterial or fungal) with a resistant host plant involves a highly localized necrosis of the infected tissue. An elicitor molecule produced as a result of the infection is thought to initiate a G protein-coupled signal transduction pathway that activates a plasma membrane-local-ized oxidase generating O2− and H2O2 in an
NADPH-dependent reaction [40 – 42]. An
addi-tional contributor to the elevated H2O2appears to
be a programmed decrease in cytosolic APX protein in response to pathogen infection, as re-ported in tobacco [29]. This result was further substantiated in experiments where APX activity was decreased using antisense RNA; such plants are hypersensitive to pathogen infection [43]. Thus, programmed declines in APX activity, as we report in this study, may constitute a general mechanism for ensuring the production of H2O2.
It is thought that the locally high levels of ROS produced during the HR catalyze a strengthening of the cell wall of the infected cells by forming crosslinks, apparently to prevent the spread of the pathogen. High local concentrations of H2O2 also
initiate the programmed cell death pathway that requires the expression of new genes for the de-struction of the affected cells [42]. In adjacent cells, the levels of ROS are lower and thus do not reach the threshold level for programmed cell death. However, the lower levels are sufficient to activate the expression of a battery of genes, in-cluding superoxide dismutase (SOD) and glu-tathione S-transferase, that appear to provide protection from the ROS [26].
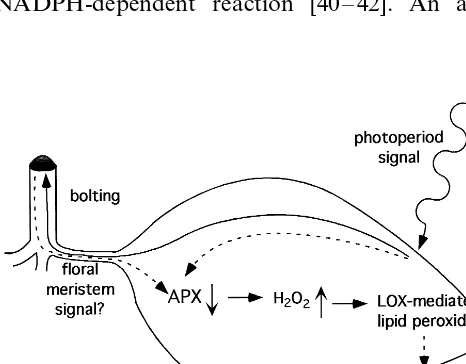
From our results and the above discussion, we propose the following model for the coupling of floral initiation and senescence (Fig. 8). The transi-tion of the vegetative meristem to a reproductive meristem is a consequence of an unknown plant age-dependent, photoperiod signal [44]. Either that same signal, or a different diffusible substance from the floral meristem, results in a downregula-tion of APX activity. Loss of APX would cause an accumulation of H2O2, which may be exacerbated
by the observed increase in Fe-SOD shortly there-after. H2O2is known to activate LOX in vitro, and
an in vivo activation may lead to the observed accumulation of 13-HOTE. This may represent the first step in the synthesis of a lipid-derived second messenger that leads to the activation of the early senescence-associated genes (SAGs). Subsequent SAGs (middle and late) would be activated in a hierarchical manner via transcription factor mem-bers of the early SAG classes. Thus, ROS need not be present throughout all stages leading to senes-cence but only as early players in the initiation of the gene expression cascade leading to the eventual regulated dismantling of the leaves during senescence.
Acknowledgements
This work was supported by grants from the National Science Foundation (DBI-9710796 and HRB-9628725) and from the National Institutes of Health Minority Biomedical Research Support Program (GM-08101-25S3).
References
[1] S. Gepstein, Photosynthesis, in: L.D. Nooden, A.C. Leopold (Eds.), Senescence and Aging in Plants, Aca-demic Press, 1988, pp. 85 – 109.
[2] J.E. Thompson, The molecular basis for mebrane deteri-oration during senescence, in: L.D. Nooden, A.C. Leopold (Eds.), Senescence and Aging in Plants, Aca-demic Press, 1988, pp. 51 – 83.
[3] L.L. H. ensel, V. Grbic, D.A. Baumgarten, A.B. Bleecker, Developmental and age-related processes that influence the longevity and senescence of photosynthetic tissues in Arabidopsis, Plant cell 5 (1993) 553 – 564. [4] K.N. Lohman, S. Gan, M.C. John, R.M. Amasino,
Molecular analysis of natural leaf senescence in Ara-bidopsis thaliana, Physiol. Plant 92 (1994) 322 – 328. [5] L.M. Weaver, S. Gan, B. Quirino, R.M. Amasino, A
comparison of the expression patterns of several senes-cence-associated genes in response to stress and hor-mone treatment, Plant Mol. Biol. 37 (1998) 455 – 469. [6] A. Blank, T.A. McKeon, Expression of three RNase
activities during natural and dark-induced senescence of wheat leaves, Plant Physiol. 97 (1991) 1409 – 1413. [7] C.B. Taylor, P.A. Bariola, S.B. Delcardayre, R.T.
Raines, P.J. Green, RNS2-a senescence-associated RNase or Arabidopsis that diverged from the S – RNases before speciation, Proc. Natl. Acad. Sci. USA 90 (1993) 5119 – 5122.
[8] K. Kamachi, T. Yamaya, T. Hayakawa, T. Mae, K. Ojima, Changes in cytosolic glutamine synthetase and its messanger RNA in a leaf blade of rice plants during natural senescence, Plant Physiol. 98 (1992) 1323 – 1329. [9] E. Himelblau, H. Mira, S.J. Lin, V.C. Culotta, L. Penar-rubia, R.M. Amasino, Identification of a functional homolog of the yeast copper homeostasis gene ATX1 fromArabidopsis, Plant Physiol. 117 (1998) 1227 – 1234. [10] A.C. Leopold, Senescence in plant development, Science
134 (1961) 1727 – 1732.
[11] P.R. Rich, W.B. Bonner, The sites of superoxide anion generation inhigher plant mitochondria, Arch. Biochem. Biophys. 188 (1978) 206 – 213.
[12] B.N. Ames, M.K. Shinenaga, T.M. Hagen, Oxidants, antioxidants, and the degenerative diseases of aging, Proc. Natl. Acad. Sci. USA 90 (1993) 7915 – 7922. [13] L.J. DeKok, M. Graham, Levels of pigments, soluble
proteins, amino acids and sulfhydryl compounds in fo-liar tissue of Arabidopsis thaliana during dark-induced and natural senescence, Plant Physiol. Biochem. 27 (1989) 203 – 209.
[14] M.V. Rao, B.A. Hale, D.P. Ormrod, Amelioriation of ozone-induced oxidative damage in wheat plants grown under high carbon dioxide, Plant Physiol. 109 (1995) 421 – 432.
[15] M.M. Bradford, A rapid and sensitive method for the quatitation of microgram quantities of protein utilizing the principle of protein-dye bunding, Anal. Biochem. 72 (1976) 248 – 254.
[16] K. Asada, K. Yoshikawa, M. Takahashi, Y. Maeda, K. Enmanji, Superoxide dismutases from blue-green alga, Plectonema boryanum, J. Biol. Chem. 250 (1975) 2801 – 2807.
[17] E.G. Bligh, W.J. Dyer, A rapid method of total lipid extraction and purification, Can. J. Biochem. Physiol. 37 (1959) 911 – 917.
[18] Z.Y. Jiang, A.C. Woodlard, S.P. Wolff, Lipid hydroper-oxide measurement by oxidation of Fe2+ in the pres-ence of xylenol orange. Comparison with the TBA assay andan iodometric method, Lipids 26 (1991) 853 – 856. [19] J.F. Wintermans, A.D. Mots, Spectrophotmetric
charac-teristicsof chlorophylls a and b and their pheophytins in ethanol, Biochem. Biophys. Acta 109 (1965) 448 – 453. [20] N. Degousee, C. Triantaphylides, S. Starek, G. Iacazio,
D. Martini, C. Bladier, R. Voisine, J.-L. Montillet, Measurement of thermally produced volatile alkanes: an assay for plant hydroperoxy fatty acid evaluation, Anal. Biochem. 224 (1995) 524 – 531.
[21] C.E. Finch, Longevity, senescence and the genome. The University of Chicago Press: Chicago, (1990).
[22] J. Browse, C.R. Somerville, Glycerolipids, in: E.M. Meyerowitz, C.R. Somerville (Eds.), Arabidopsis, 1994, pp. 881 – 912.
[23] H.W. Chan, G. Levett, Autoxidation of methyl-linoleate. Separationand analysis of isomerice mixtures of methyl-linoleate-hydroperoxidases and methyl hy-droxylinoleates, Lipids 12 (1977) 99 – 104.
[24] L.D. Nooden, A.C. Leopold, Senescence and aging in plants. Academic Press Inc., San Diego (1988).
[25] H.M. Jespersen, I.V. Kjaersgard, L. Ostergaard, K.G. Welinder, From sequence analysis of three novel ascor-bate peroxidases from Arabidopsis thaliana to structure, function and evolution of seven types of ascorbate per-oxidase, Biochem. J. 326 (1997) 305 – 310.
[26] I. Crute, J. Benyon, J. Dangl, E. Holub, B. Mauchi-Mani, A. Slusarenko, B. Staskawicz, F. Ausubel, Micro-bial pathogenesis in Arabidopsis, in: E.M. Meyerowitz, C.R. Somerville (Eds.), Arabidopsis, CSHL Press, 1994, pp. 705 – 747.
[27] R. Mittler, B. Zilinskas, Regulation of pea cytosolic ascorbate peroxidase and other antioxidant enzymes during the progression of drought stress and following recovery from drought, Plant J. 5 (1994) 397 – 405. [28] F. Lopez, G. Vansuyt, F. Casse-Delbart, P. Fourcroy,
Ascorbate peroxidase activity, not the mRNA level, is enhanced in salt-stressed Raphanus sati6us plants, Plant
Physiol. 97 (1996) 13 – 20.
[30] A. Jimenez, J.A. Hernandez, G. Pastori, L.A. delRio, F. Sevilla, Role of the ascorbate – glutathione cycle of mito-chondria and peroxisomes in the senescence of peas leaves, Plant Physiol. 118 (1998) 1327 – 1335.
[31] J. deGroot, G.A. Veldink, J.F.G. Vliegenthart, J. Bold-ingh, R. Wever, B. vanGelder, Demonstration by EPR spectroscopy of the functional role of iron in soyabean lipoxygenase – 1, Biochim. Biophys. Acta 377 (1976) 71 – 79.
[32] F. Macri, E. Braidot, E. Petrussa, A. Vianello, Lipoxy-genase activity associated to isolated soybean plasma membranes, Biochim. Biophys. Acta 1215 (1994) 109 – 114.
[33] B. McGurl, G. Pearce, C. Ryan, Polypeptide signalling for plant defence gene, Biochem. Soc. Symp. 60 (1994) 149 – 154.
[34] N.J. Bate, S.J. Rothstein, C6-volatiles derived from the lipoxygenase pathway a subset of defense-related genes, Plant J. 16 (1998) 561 – 569.
[35] A.B. Bleecker, M.A. Estelle, C. Somerville, H. Kende, Insensitivity to ethylene conferred by a dominant muta-tion in Arabidopsis thaliana, Science 241 (1988) 1086 – 1089.
[36] V. Grbic, A.B. Bleecker, Ethylene regulates the timing of leaf senescence in Arabidopsis, Plant J. 8 (1995) 595 – 602.
[37] S.A. Oh, J.H. Park, G.I. Lee, K.H. Paek, S.K. Park, H.G. Nam, Identification of three genetic loci
con-trolling leaf senescence in Arabidopsis thaliana, Plant J. 12 (1997) 527 – 535.
[38] L.D. Noode´n, J.W. Hillsberg, M.J. Schneider, Induction of leaf senscence in Arabidopsis thaliana by long days through a light-dosage effect, Physiol. Plant 96 (1996) 491 – 495.
[39] J. Kurepa, J. Smalle, M. VanMontagu, D. Inze, Oxida-tive stress tolerance and longevity in Arabidopsis: the late flowering mutant gigantea is tolerant to paraqaut, Plant J. 14 (1998) 759 – 764.
[40] C.J. Lamb, Plant disease resistance genes in signal per-ception and transduction, Cell 76 (1994) 419 – 422. [41] L. Legendre, P.F. Heinstein, P.S. Low, Evidence for
participation of GTP – binding proteins in elucidationof the rapid oxidation burst in cultured soybean cells, J. Biol. Chem. 267 (1992) 20140 – 20147.
[42] R. Tenhaken, A. Levine, L.F. Brisson, R.A. Dixon, C.J. Lamb, Function of the oxidative burst in hypersenstive disease resistance, Proc. Natl. Acad. Sci. USA 92 (1995) 4158 – 4163.
[43] R. Mittler, E.H. Herr, B.L. Orvar, W. vanCamp, H. Willekens, D. Inze, B.E. Ellis, Transgenic tobacco plants with reduced capability to detoxify reactive oxygen inter-mediates are hyperresponsive to pathogen infection, Proc. Natl. Acad. Sci. USA 96 (1999) 14165 – 14170. [44] S.D. O’Neill, The photoperiodic control of flowering
progress toward understanding the mechanism of induc-tion, Photochem. Photobiol. 56 (1992) 789 – 801.