ABSTRACT
Blast is a devastating disease in rice caused by the filamentous Ascomycetes Pyricularia oryzae. The disease showed a high level of variability and can be classified into various pathotypes or races based on the infection pattern observed on a set of differential rice cultivars. Information on the genetic variability as well as structure of the fungal population in the field are essential to determine the suitable host resistance. The research was conducted to study the genetic variability of blast pathogen originated from five endemic areas of blast disease, namely North Sumatra, West Sumatra, Bogor (West Java), Lampung, and Sukabumi (West Java). Sequence-specific DNA that confers to pathogenesis of the pathogen, i.e. host-species specificity (Pwl2) gene, sterol delta 8 to delta 7 isomerase (Erg2) gene, and Cutinase (Cut1) gene. The sequence-specific DNA of P. oryzae isolates was generated using a pair of specific primers corresponding to
Pwl2, Erg2, Cut1 gene. Eight haplotypes were detected in all of 230 isolates, where 2.2% is haplotype A-000, 6.1% haplotype B-001, 6.1% haplotype C-011, 60.9% haplotype D-111, 1.3% haplotype E-010, 5.2% haplotype F-110, 1.7% haplotype G-100, and 16.1% haplotype H-101. Heplotype D-111 was dominantly found in each isolate location. [Keyword: Pyricularia oryzae, genetic diversity, sequence-specific DNA, primers, haplotype, host resistance]
ABSTRAK
Penyakit blas yang disebabkan oleh cendawan Pyricularia oryzae merupakan salah satu penyakit penting pada padi. Penyakit ini memiliki keragaman yang tinggi dan ter-kelompok dalam beberapa patotipe atau ras berdasarkan pola infeksi pada satu set varietas diferensial padi. Keragaman genetik dan patotipe serta struktur populasi P. oryzae di lapangan perlu diketahui terutama untuk menentukan tanaman inang yang memiliki gen ketahanan yang sesuai. Penelitian ini dilakukan untuk mengetahui keragaman genetik P. oryzae
pada tingkat molekuler dengan menganalisis DNA cendawan
P. oryzae yang berasal dari lima daerah endemis blas, yaitu Sumatera Utara, Sumatera Barat, Bogor, Lampung, dan Sukabumi. Analisis dilakukan dengan menggunakan primer spesifik yang berkaitan dengan patogenisitas patogen, yaitu gen Pwl2 (gen avirulen yang memiliki inang spesifik), gen
Erg2 (metabolit sekunder yang mengkatalisis perubahan sterol delta 8 menjadi delta 7 isomerase), dan gen Cut1 (gen cutinase yang menghidrolisis kutin pada dinding sel tanaman). Hasil analisis PCR dengan tiga primer spesifik pada 230 DNA isolat
P. oryzae menunjukkan bahwa keragaman genetik cendawan tersebut terkelompok menjadi delapan haplotip, yaitu haplotip A-000 sebanyak 2,2%, B-001 6,1%, C-011 6,1%, D-111 60,9%, E-010 1,3%, F-110 5,2%, G-100 1,7%, dan H-101 16,1%. Haplotip D-111 merupakan kelompok yang dominan pada setiap lokasi asal isolat.
[Kata kunci: Pyricularia oryzae, keragaman genetik, DNA sekuen spesifik, primer, haplotipe, ketahanan inang]
PENDAHULUAN
Penyakit blas yang disebabkan oleh cendawan
Pyricularia oryzae merupakan salah satu penyakit penting pada tanaman padi. Di Asia Selatan dan Asia Tenggara, kerugian akibat penyakit ini diperkirakan mencapai US$55 juta/tahun (Herdt 1991). Di Indonesia penyakit ini juga mendapat perhatian besar terutama pada pertanaman padi di lahan kering (gogo). Oleh karena itu, program pemuliaan yang mengarah pada pembentukan varietas yang tahan ataupun toleran terhadap penyakit blas terus dikembangkan.
Cendawan P. oryzae dapat menyerang daun (leaf blast), buku (node blast), leher malai (neck blast), bulir padi (spikelet blast) (Chen 1993; Scardaci et al. 1997), dan kolar daun (collar rot) (Scardaci et al. 1997). Bentuk dan ukuran bercak dipengaruhi oleh perbedaan kultivar dan kondisi lingkungan. Selain menyerang padi, penyakit blas juga dapat menimbulkan kerusakan pada tanaman serealia seperti gandum dan sorgum (Kahmann dan Basse 1997) serta 40 spesies gulma rumput-rumputan dan gulma lainnya (Ou 1985).
Tingkat keragaman P. oryzae cukup tinggi karena cendawan ini mampu melakukan perkawinan antar-haploid hifa yang berlainan material genetiknya (parasexual exchanged DNA) (Zeigler et al. 1997) serta adanya elemen transposon POT2 (Kachroo et al.
Keragaman genetik cendawan
Pyricularia oryzae
berdasarkan primer spesifik gen virulensi
Genetic variability of the rice blast fungus Pyricularia oryzae
based on sequence-specific primers linked to pathogen virulence genes
Reflinur1, Masdiar Bustamam1, Utut Widyastuti2, dan H. Aswidinnoor2
1Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian,
Jalan Tentara Pelajar No. 3A, Bogor 16111, Indonesia
1994) dan MAGGY (Leong et al. 1994) dalam genom yang memungkinkan terjadinya segregasi dan rekom-binasi antar-ras P. oryzae sehingga tingkat mutasinya cukup tinggi (Kiyosawa 1976).
Informasi mengenai gen ketahanan tanaman dan jenis isolat (ras) merupakan dasar dalam perakitan piramiding gen-gen Pi. Pada tanaman padi telah di-ketahui terdapat 20 gen major yang mengendalikan ketahanan terhadap penyakit blas (McCouch et al. 1994).
Gen-gen yang mengendalikan perkembangan blas selama infeksi telah berhasil diisolasi dan dikarak-terisasi. Beberapa gen mempengaruhi pembentukan apresorium sedangkan yang lainnya penting untuk fungsi penetrasi apresorium. Gen-gen yang terlibat dalam diferensiasi apresorium dikelompokkan menjadi gen biosintesis melanin, gen yang mempengaruhi pembentukan konidia atau morfologi konidia, dan gen yang terlibat dalam pengenalan permukaan inang dan jalur penerusan sinyal (Tenjo dan Hamer 2002).
Penelitian sifat genetik isolat P. oryzae dilakukan dengan menggunakan tiga macam primer yang mem-punyai peran spesifik dalam pengenalan inang P. oryzae. Ketiga primer tersebut adalah: (1) Cut1 yang merupakan lokus DNA blas yang menjadikan gen cutinase berfungsi sebagai pendegradasi lapisan kitikula tanaman (Sweigard et al. 1992), (2) Erg2 merupakan lokus DNA yang berperan sebagai pe-nyandi metabolit sekunder pada cendawan yang menjadi target antifungal pada sel tanaman (Keon et al. 1994), dan (3) Pwl2 sebagai gen avirulen yang ber-sifat spesifik inang (Valent and Chumley 1994). Ketiga gen tersebut telah dikembangkan menjadi primer
sequence characterized amplified region markers
(SCAR) yang dapat digunakan untuk memonitor rekombinasi dan perubahan struktur populasi P. oryzae (Soubarbere et al. 2001). Penelitian ini dilakukan untuk mengetahui keragaman genetik dan mengelompokkan isolat-isolat P. oryzae berdasarkan lokus Cut1, Erg2 dan Pwl2, serta memonitor rekom-binasi dan dinamika populasi P. oryzae di daerah endemis blas.
BAHAN DAN METODE
Penelitian dilaksanakan di Laboratorium Biologi Molekuler dan Rekayasa Genetik serta rumah kaca Balai Besar Penelitian dan Pengembangan Biotek-nologi dan Sumberdaya Genetik Pertanian (BB Biogen), Bogor mulai bulan Januari sampai September 2004. Isolat P. oryzae yang digunakan meliputi 230
isolat dari lima ras P. oryzae standar (Mogi et al. 1991) serta 225 isolat P. oryzae koleksi BB Biogen yang berasal dari Lampung, Sukabumi, Bogor, Sumatera Utara, dan Sumatera Barat.
Isolasi cendawan
Cendawan P. grisea diisolasi mengikuti metode isolasi monokonidia oleh Bonman et al. (1986). Satu konidia diidentifikasi dengan stereomikroskop dan ditransfer secara aseptik ke media prune agar slants (3 buah prune/l, laktosa 5 g/l, yeast ekstrak 1 g/l, agar bacto 17 g/l) dan diinkubasi selama 4 hari pada suhu ruang. Biakan yang tumbuh pada media prune agar slants
digunakan sebagai sumber isolat dalam isolasi DNA.
Isolasi DNA genom
Isolasi DNA genom cendawan P. oryzae dilakukan dari kultur miselia cendawan. Penyiapan kultur miselia cendawan mengikuti metode Chen (1993). Miselia dipanen dari kultur cair menggunakan kertas saring Whatman No. 1 yang diletakkan pada alat filtrasi Buchner. Miselia selanjutnya diliofilisasi dengan vakum dan dikeringbekukan selama 24 jam pada suhu 50oC dan tekanan 50-70 atm. Miselia dapat disimpan pada suhu -20oC sampai saat akan dipergunakan.
DNA genom total cendawan P. oryzae diisolasi menggunakan larutan CTAB/NaCl mengikuti metode IRRI (1994) yang dimodifikasi melalui proses dena-turasi makromolekul (seperti protein) dengan larutan fenol:khloroform:isoamilalkohol (25:24:1 v/v/v) 1x volume larutan. Selanjutnya supernatan dimurnikan dengan penambahan larutan khloroform:isoamil-alkohol (24:1) 0,5x volume larutan sebanyak dua kali.
Analisis PCR
Keragaman genetik cendawan P. oryzae untuk sifat virulensi diidentifikasi menggunakan markah Erg2,
Pwl2, dan Cut1, yaitu markah yang mempunyai hu-bungan dengan virulensi (Soubabere et al. 2001). Identifikasi dilakukan dengan PCR menggunakan DNA genom sebanyak 25 ng serta sepasang primer spesifik untuk gen Erg2 (1.440 bp), Pwl2 (900 bp), dan
Cut1 (1.730bp). Analisis PCR dilakukan pada total reaksi 20 µl yang mengandung 25 ng DNA genom cetakan, dNTP 0.1 µM (dATP, dCTP, dGTP, dan dTTP), primer (Cut1, Erg2, dan Pwl2) 1 pmol, enzim Taq DNA polymerase 1 unit dalam larutan bufer 1x. Urutan basa dari primer-primer tersebut adalah:
a) Cut1 (F:5’-TATAGCGTTGACCTTGTGGA3’; R:5’TAAGCATCTCAGACCGAACC-3’)
b) Erg2 (F:5’-GCAGGGCTCATTCTTTTCT3’; R:5’CCGACTGGAAGGTTTCTTTA-3’) c) Pwl2 (F:5’-TCCGCCACTTTTCTCATTCC-3’;
R:5’- GCCCTCTTCTCGCTGTTCAC-3’). Reaksi amplifikasi dilakukan berdasarkan metode Soubabere et al. (2001). Sebanyak 10 µl produk PCR diseparasi dengan elektroforesis pada gel agarose 1% (w/v). Pengamatan dilakukan terhadap pola pita DNA hasil amplifikasi DNA pada ukuran 1.730 bp untuk primer Cut1, 1.440 bp untuk primer Erg2, dan 900 bp untuk primer Pwl2 (Soubabere et al. 2001). Pita DNA hasil amplifikasi yang muncul untuk setiap primer dari masing-masing isolat diskor dengan nilai 1 (ada) dan 0 (tidak ada). Selanjutnya data ini digunakan untuk mengelompokkan isolat-isolat uji menjadi beberapa haplotip, dengan asumsi diperoleh delapan jenis haplotip berdasarkan kombinasi ketiga jenis gen (Pwl2, Erg2, dan Cut1) pada isolat-isolat yang diuji. Penentuan haplotip tersebut disajikan pada Tabel 1.
HASIL DAN PEMBAHASAN
Hasil elektroforesis produk PCR yang merupakan representasi dari masing-masing haplotip disajikan pada Gambar 1. Hasil penelitian menunjukkan bahwa berdasarkan keberadaan gen Pwl2, Erg2, dan Cut1 pada 230 isolat P. oryzae yang dianalisis diperoleh delapan jenis haplotip, yaitu haplotip A-000 sebanyak 5 isolat (2,2%), B-001 14 isolat (6,1%), C-011 14 isolat (6,1%), D-111 141 isolat (61,3%), E-010 3 isolat (1,3%), F-110 12 isolat (5,2%), G-100 4 isolat (1,7%), dan
haplotip H-101 37 isolat (16,1%) (Tabel 2 ). Dari delapan jenis haplotip tersebut, haplotip D-111 paling banyak ditemukan, diikuti oleh haplotip F-110, sedangkan E-010 paling sedikit ditemukan.
Dilihat dari tiga daerah asal isolat P. oryzae, isolat asal Lampung menunjukkan keragaman genetik yang tinggi (8 haplotip), diikuti isolat dari Sukabumi (7 haplotip) dan daerah lain (Sumatera Utara, Sumatera Barat, dan Bogor) sebanyak 6 haplotip. Hal ini me-nunjukkan bahwa umumnya cendawan P. oryzae yang diuji memiliki ketiga gen yang terlibat dalam proses patogenisitas.
Tabel 1. Pengelompokan haplotip cendawan Pyricularia oryzae berdasarkan keberadaan tiga gen virulensi.
Tabel 1. Classification of Pyricularia oryzae haplotypes based on three virulence genes.
Haplotip
Gen penyandi virulensi
Haplotype
Virulence encoding gene Pwl2a Erg2b Cut1c A - 0 0 0 01 0 0 B - 0 0 1 0 0 1 C - 0 1 1 0 12 1 D - 1 1 1 1 1 1 E - 0 1 0 0 1 0 F - 1 1 0 1 1 0 G-100 1 0 0 H - 1 0 1 1 0 1
Keterangan: a = 900 bp, b = 1440 bp, c = 1730 bp, 1 = tidak ada
pita DNA, 2 = ada pita DNA.
Notes: a = 900 bp, b = 1440 bp, c = 1730 bp, 1 = no DNA band, 2 = DNA band.
Gambar 1. Profil DNA genom cendawan Pyricularia oryzae yang mewakili masing-masing haplotip berdasarkan hasil amplifikasi dengan primer Cut1, Erg2 dan Pwl2.
Fig. 1. Genom DNA profile of Pyricularia oryzae representing haplotype based on amplification result using Cut1, Erg2 and
Pwl2 primers. 1.730 bp (Cut1) ▲ ▲ ▲ 1.730 bp (Erg2) 1.730 bp (Pwl2) A-000 1kb B-001 C-011 D-111 E-010 F-110 G-100 H-101 1kb
Haplotip G-100 tidak ditemukan pada isolat asal Sukabumi, sedangkan haplotip A-000 dan E-010 tidak dijumpai pada isolat dari Sumatera Utara, Sumatera Barat, dan Bogor. Perbedaan jenis haplotip pada satu lokasi asal isolat dan antar-ketiga lokasi asal isolat menunjukkan adanya keragaman genetik P. oryzae. Keragaman haplotip P. oryzae antara lain dipengaruhi oleh faktor lingkungan seperti suhu dan kelembapan, baik pada lokasi yang sama maupun berbeda. Chen et al. (1995) melaporkan bahwa perbedaan agroekologi lokasi asal P. oryzae menyebabkan diferensiasi yang besar dalam sidik jari DNA genom cendawan dengan pelacak MGR586.
Dalam kaitannya dengan keragaman, salah satu komponen lingkungan yang berpengaruh terhadap epidemi penyakit tanaman (piramida penyakit) adalah faktor spasial, yang mencakup penyebaran suatu populasi patogen dalam suatu area yang luas. Peran faktor spasial dalam membentuk keragaman genetik populasi P. oryzae juga berkaitan dengan keberadaan inang atau varietas yang ditanam pada suatu lokasi. Menurut Agrios (1998), lingkungan dapat mempenga-ruhi ketersediaan inokulum, tingkat pertumbuhan patogen, daya tahan hidup patogen, kerentanan ge-netik inang, serta arah dan jarak penyebaran patogen. Ditinjau dari penyebaran haplotip P. oryzae pada tiap lokasi (Tabel 2), haplotip D-111 merupakan kelom-pok yang paling banyak ditemukan di semua lokasi asal isolat (Lampung, Sukabumi, Sumatera Utara, Sumatera Barat, dan Bogor). Dari 114 isolat P. oryzae
asal Lampung, 64 isolat (54,4%) merupakan haplotip D-111, diikuti dengan haplotip H-101 sebanyak 26 isolat (22,8%). Sebanyak 59 isolat (70,7%) dari 82 isolat asal Sukabumi termasuk dalam haplotip D-111, diikuti oleh haplotip F-110 sebanyak 8 isolat (9.8%). Pada lokasi lain (Sumatera Utara, Sumatera Barat, dan Bogor), dari 34 isolat, 17 isolat (50%) adalah haplotip D-111 dan 5 isolat (14,7%) haplotip H-101.
Berdasarkan proporsi haplotip, dalam populasi genom P. oryzae ditemukan 89,1% gen Cut1, 84,8% gen Pwl2, dan 73.1% gen Erg2. Hal ini menunjukkan bahwa umumnya genom P. oryzae memiliki ketiga gen yang terlibat dalam patogenisitas blas. Tenjo dan Hamer (2002) melaporkan bahwa perkembangan P. oryzae selama proses infeksi dikendalikan oleh banyak gen dan beberapa gen di antaranya sudah berhasil diisolasi.
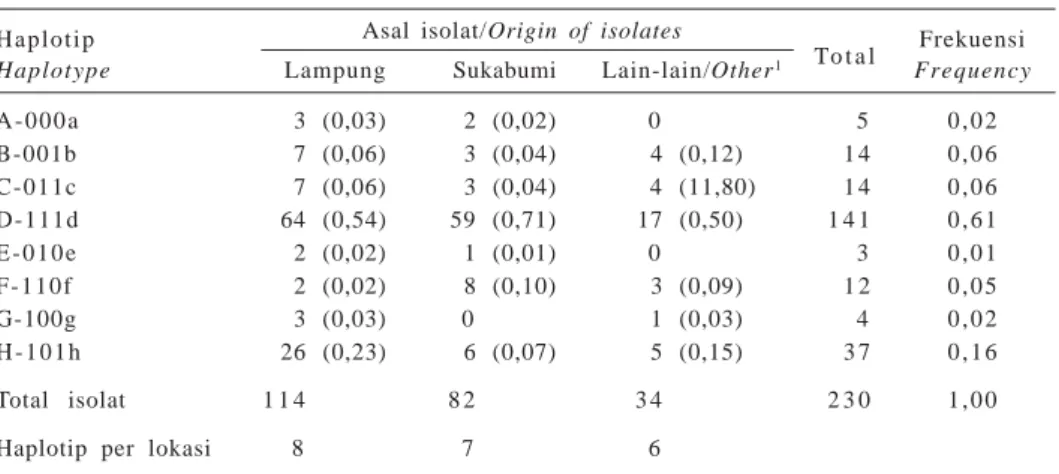
Proporsi ketiga gen berdasarkan asal isolat cen-dawan disajikan pada Gambar 2. Ketiga gen penyandi virulensi blas ini ditemukan pada sebagian besar isolat dari ketiga lokasi. Di Lampung, gen yang paling banyak ditemukan adalah Cut1 (91,2%), diikuti oleh
Pwl2 dan Erg2 masing-masing 83,3% dan 65,8%. Di Sukabumi, gen yang paling banyak diperoleh adalah
Pwl2 (89%), sementara gen Cut1 dan Erg2 masing-masing 86,6%.
Gen dengan virulensi tertinggi yang ditemukan pada lokasi lain (Sumatera Utara, Sumatera Barat, dan Bogor) adalah Cut1 (88,2%), diikuti oleh dan Erg2 (82,4%) dan Pwl4 (76,5%). Untuk menentukan
inter-Tabel 2 Frekuensi masing-masing haplotip 230 isolat cendawan Pyricularia oryzae.
Table 2. Frequency of 230 haplotype of Pyricularia oryzae isolates.
Haplotip Asal isolat/Origin of isolates
To t a l Frekuensi
Haplotype Lampung Sukabumi Lain-lain/Other1 Frequency
A - 0 0 0 a 3 (0,03) 2 (0,02) 0 5 0 , 0 2 B-001b 7 (0,06) 3 (0,04) 4 (0,12) 1 4 0 , 0 6 C-011c 7 (0,06) 3 (0,04) 4 (11,80) 1 4 0 , 0 6 D-111d 64 (0,54) 59 (0,71) 17 (0,50) 1 4 1 0 , 6 1 E - 0 1 0 e 2 (0,02) 1 (0,01) 0 3 0 , 0 1 F - 1 1 0 f 2 (0,02) 8 (0,10) 3 (0,09) 1 2 0 , 0 5 G-100g 3 (0,03) 0 1 (0,03) 4 0 , 0 2 H - 1 0 1 h 26 (0,23) 6 (0,07) 5 (0,15) 3 7 0 , 1 6 Total isolat 1 1 4 8 2 3 4 2 3 0 1 , 0 0
Haplotip per lokasi 8 7 6
Keterangan/Notes:
1Isolat berasal dari Sumatera Utara, Sumatera Barat, dan Bogor.
1Isolat originated from North Sumatra, West Sumatra, and Bogor.
a = Pwl2 (0), Erg2 (0), Cut1(0), b = Pwl2 (0), Erg2 (0), Cut1(1), c = Pwl2 (0), Erg2 (1), Cut1(1), d = Pwl2 (1), Erg2 (1), Cut1(1), e = Pwl2 (0), Erg2 (1), Cut1(0), f = Pwl2 (1), Erg2 (1), Cut1(0), g = Pwl2 (1), Erg2 (0), Cut1(0), h = Pwl2 (1), Erg2 (0), Cut1(1).
aksi gen-gen tersebut dalam menimbulkan penyakit ataupun gen yang paling dominan berperan dalam proses patogenisitas perlu dilakukan penelitian lebih lanjut.
KESIMPULAN
Amplifikasi DNA 230 isolat cendawan P. oryzae dari lima lokasi, yaitu Lampung, Sukabumi, Sumatera Utara, Sumatera Barat, dan Bogor dengan menggunakan primer spesifik gen virulensi, menghasilkan delapan haplotip, yaitu haplotip A-000 5 isolat (2,2%), B-001 14 isolat (6,1%), C-011 14 isolat (6,1%), D-111 141 isolat (61,3%), E-010 3 isolat (1,3%), F-110 12 isolat (5,2%), G-100 4 isolat (1,7%), dan H-101 37 isolat (16,1%). Haplotip D-111 paling banyak ditemukan pada setiap lokasi asal isolat P. grisea.
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Mahrup yang telah banyak membantu dalam isolasi mono-konidia isolat P. oryzae dan Ma’sumah dalam isolasi DNA cendawan tersebut. Penulis juga mengucapkan terima kasih kepada Dr. Mukelar Amir yang telah membantu penulis dalam memperoleh lima ras standar Indonesia.
DAFTAR PUSTAKA
Agrios, N.G. 1988. Plant Pathology. Academic Press, University of Florida. p. 198-235.
Bonman, J.M, T.I Vergel De Dios, and M.M. Khin.1986. Physiologic specialization of P. oryzae in the Philippines. Plant Dis. 70: 767-769.
Chen, D. 1993. Population structure of Pyricularia grisea
(Cooke) Sacc. in two screening sites and quantitative characterization of major and minor resistance genes. PhD Thesis, University of the Philippines at Los Banos. 161 pp. Chen, D., R.S. Zeigler, H. Leung, and R.J. Nelson. 1995. Population structure of Pyricularia grisea at two screening sites in the Philippines. Phytopathology 85: 1011-1020. Herdt, R.W. 1991. Research priorities for rice biotechnology. p. 35-37. In G.S Khush and G.H. Toennissen (Eds). Rice Biotechnology. Alden Press Ltd., London.
Kachroo, P., S.A. Leong, and B.B. Chattoo. 1994. Pot2, an inverted repeat transposon from the rice blast fungus
Magnaporthe grisea. Mol. Gene Genet 245: 39-348. Kahmann, R. and C. Basse. 1997. Signaling and development
in pathogenic fungi, new strategies for plant protection. Trends Plant Sci. 2: 366-368.
Keon J.P., C.S. James, and S. Court. 1994. Isolation of Erg2
gene, encoding sterol delta 8 to delta isomerase, from the rice blast fungus Magnaporthe grisea and its expression in the maize smut pathogen Ustilago maydis. Curr. Genet. 25: 5 3 1 - 5 3 7 .
Kiyosawa, S. 1976. Pathogenic variation of Pyricularia oryzae
and their use in genetic and breeding studies. Sabrao J. 8: 53-67. Leong, S.A., M. Farman, J. Smoith, A. Budde, Y. Tosa, and N. Nita. 1994. Molecular genetic approach to the study of cultivar specificity in the rice blast fungus. p. 87-110. In
Gambar 2. Frekuensi gen yang ditemukan pada 230 isolat Pyricularia oryzae berdasarkan lokasi asal isolat.
Fig. 2. Frequency of gen Pwl2, Erg2, and Cut1 in 230 isolates of Pyricularia oryzae.
Frekuensi isolat Frequency of isolates ( % ) 8 3 , 3 3 8 9 , 0 7 6 , 5 6 5 , 7 9 8 6 , 6 8 2 , 4 9 1 , 2 3 8 6 , 6 8 8 , 2 1 0 0 9 0 8 0 7 0 6 0 5 0 3 0 4 0 2 0 0 Pwl2 Erg2 Cut1 1 0 Gen
Lampung Sukabumi Lain-lain/Other
1 2 0 1 1 0
R.S. Zeigler, S.A. Leong, and P.S. Teng (Eds.). Rice Blast Disease. CAB International, Wallingford (UK) and IRRI, Manila, Philippines.
McCouch, S.R., R.J. Nelson, J. Tohme, and R.S. Zeigler. 1994. Mapping of blast resistance genes in rice. p. 167-186. In
R.S. Zeigler, S.A. Leong, and P.S. Teng (Eds.). Rice Blast Disease. CAB International, Wallingford (UK) and IRRI, Manila, Philippines.
Mogi, S. 1991. Establishment of the differential variety series for pathogenic race identification of rice blast fungus and the distribution of race based on the new differentials in Indonesia. Rice Disease Study Group Karawang, Jatisari. Sukamandi Research Institute for Food Crops, Subang. 30 pp. Ou, SH. 1985. Rice Disease. Commonw. Mycol. Inst. 2nd ed.
Kew. Surrey, England. 380 pp.
Scardaci, S.C., R.K. Webster, C.A. Greer, J.E Hill, J.G. Williams, R.G. Mutters, D.M. Brandon, K.S. McKenzie, and J.J. Oster. 1997. Rice blast: a new disease in California. Agronomy Fact Sheet Series 1997-2. Department of Agronomy and Range Science, University of California, Davis. 3 pp.
Soubabere, O., V. Jorge, J.L. Notteghem, M.H. Lebrun, and D. Tharreau. 2001. Sequence characterized amplified region markers for the rice blast fungus, Magnaphorthe grisea. Molecular Ecology Notes 1(Issue 1-2): 11-12.
Sweigard, J.A., F.G. Chumley, and B. Valent. 1992. Disruption of Magnaporthe grisea cutinase gene. Mol. Gene Genet. 232: 183-190.
Tenjo, F.A. and J.E. Hamer. 2002. Pathogenic development in Magnaporthe grisea. p. 399-418. In J.W. Goethe (Ed.). Molecular Biology of Fungal Development. Universitat Frankfurt, Germany.
Valent, B. and F.G. Chumley. 1994. Avirulence genes and mechanisms of genetic instability in rice blast fungus. p. 111-134. In R.S. Zeigler, S.A. Leong, and P.S. Teng (Eds.). Rice Blast Disease. Wallingford (UK): CAB International-IRRI, Manila, Philippines.
Zeigler, R.S., R.P. Scott, H. Leung, A.A. Bordeos, J. Kumar, and R.J. Nelson 1997. Evidence of parasexual exchange of DNA in the rice blast fungus: challenges its exclusive clonality. Phytopathology 87: 284-294.