ASOSIASI POLIMORFISME GEN KCNJ11 DAN ABCC8
DENGAN DIABETES MELLITUS TIPE 2 PADA
POPULASI MASYARAKAT BALI
AGENG WIYATNO
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
ASOSIASI POLIMORFISME GEN KCNJ11 DAN ABCC8
DENGAN DIABETES MELLITUS TIPE 2 PADA
POPULASI MASYARAKAT BALI
AGENG WIYATNO
Skripsi
sebagai salah satu syarat untuk memperoleh gelar
Sarjana Sains pada
Departemen Biokimia
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
Judul Skripsi : Asosiasi Polimorfisme Gen KCNJ11 dan ABCC8 dengan
Diabetes Mellitus Tipe 2 pada Populasi Masyarakat Bali
Nama
: Ageng Wiyatno
NRP
: G84070047
Disetujui
Komisi pembimbing
Dr. Ir. I Made Artika, M.App.Sc.
Dr. drh. Safarina G. Malik, M.S.
Ketua
Anggota
Diketahui
Dr. Ir. I Made Artika, M.App.Sc.
Ketua Departemen Biokimia
PRAKATA
Assalamualaikum wr.wb
Penulis panjatkan puji syukur ke hadirat Allah SWT. Atas rahmat-Nya
penelitian ini telah selesai. Salawat dan salam mudah-mudahan selalu tercurah
kepada junjungan Nabi Besar Muhammad SAW. Laporan hasil penelitian ini
berjudul “
Asosiasi Polimorfisme Gen
KCNJ11 dan
ABCC8 dengan
Diabetes
Mellitus Tipe 2 pada Populasi Masyarakat Bali
”. Penelitian telah
dilakukan di
Lembaga Biologi Molekuler Eijkman pada bulan Maret hingga bulan September
2011, dibimbing oleh Dr. Ir. I Made Artika, M.App.Sc. dan Dr. drh. Safarina G.
Malik, M.S.
Penulis sampaikan terima kasih kepada Dr. Ir. I Made Artika, M.App.Sc.
dan Dr. drh. Safarina G. Malik, M.S. atas kesediaan untuk membimbing penulis.
Terima kasih untuk Prof. dr. Sangkot Marzuki, AM., PhD., DSc. dan Prof. dr.
Herawati Sudoyo, PhD. yang telah memberikan kesempatan kepada penulis untuk
melakukan penelitian di Lembaga Eijkman. Tidak lupa terima kasih kepada
Sukma Oktavianthi, Clarissa Asha Febinia, Tri Cita Hutama, Pradiptajati Kusuma,
Yumni Khairina Ghassani dan Leli Nurfitriyani yang membantu penulis dalam
melakukan penelitian ini. Terima kasih untuk Bapak, Ibu dan adik-adikku yang
terus memberikan dukungan dan motivasi kepada penulis. Mudah-mudahan
penelitian ini berguna bagi perkembangan ilmu pengetahuan.
Bogor, Oktober 2011
Ageng Wiyatno
DAFTAR ISI
Halaman
DAFTAR GAMBAR ... ix
DAFTAR TABEL ... ix
DAFTAR LAMPIRAN ... ix
PENDAHULUAN ... 1
TINJAUAN PUSTAKA ... 2
Protein Integral K
ATPchannels dan Mekanisme Sekresi Insulin ... 2
Analisis SNP Menggunakan Metode PCR-RFLP ... 3
Gen KCNJ11 Penyandi Kir6.2 ... 4
Gen ABCC8 Penyandi SUR1 ... 4
Desa dan Penduduk Nusa Ceningan, Legian, Penglipuran dan Pedawa Bali .. 5
BAHAN DAN METODE ... 5
Bahan dan Alat ... 5
Metode Penelitian ... 6
HASIL DAN PEMBAHASAN... ... 9
Karakteristik Klinis dan Sebaran Sampel Secara Umum ... 9
Asosiasi Jenis Kelamin dengan kriteria DMT2 ... 9
Asosiasi Kelompok Usia Tertentu dengan kriteria DMT2 ... 10
Deteksi SNP E23K Gen KCNJ11 dan SNP A1369S Gen ABCC8 ... 12
Asosiasi SNP E23K Gen KCNJ11 dan SNP A1369S Gen ABCC8 dengan
kriteria DMT2 ... 13
Haplotipe Gen KCNJ11 dan ABCC8 ... 15
Asosiasi Haplotipe Gen KCNJ11 dan ABCC8 dengan kriteria DMT2 ... 15
SIMPULAN DAN SARAN ... 16
Simpulan ... 16
Saran ... 16
DAFTAR PUSTAKA ... 16
DAFTAR GAMBAR
Halaman
1 Struktur K
ATPchannel pada membran sel pankreas
... 2
2 Mekanis
me sekresi insulin pada sel
pankreas ... 3
3 Letak empat desa terpilih di Bali ... 5
4 Pengaturan suhu dan waktu yang digunakan pada reaksi PCR gen KCNJ11
dan ABCC8 ... 8
5 Asosiasi jenis kelamin dengan kriteria DMT2 berdasarkan konsentrasi FPG
pada masing-masing desa ... 9
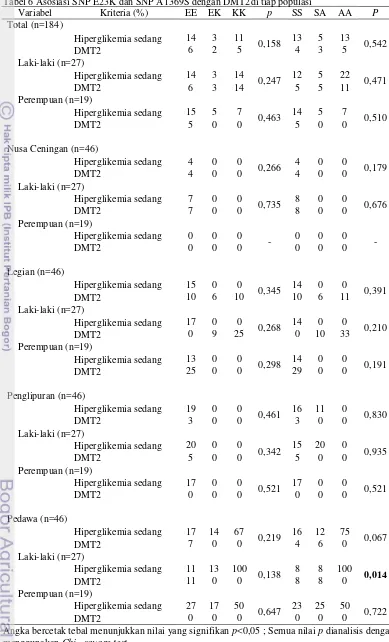
6 Representasi deteksi polimorfisme SNP E23K gen dan A1369S gen ABCC8
yang dilakukan dengan teknik PCR-RFLP ... 12
DAFTAR TABEL
Halaman
1 Kriteria individu berdasarkan konsentrasi FPG ... 1
2 Karakteristik klinis dan sebaran sampel secara umum ... 10
3 Asosiasi kelompok usia dan jenis kelamin dengan kriteria DMT2 pada masing-
masing desa ... 11
4 Nilai Frekuensi Alel Minor (MAF) gen KCNJ11 pada masing-masing
populasi ... 12
5 Nilai Frekuensi Alel Minor (MAF) gen ABCC8 pada masing-masing populasi
... 13
6 Asosiasi SNP E23K dan SNP A1369S dengan DMT2 di tiap populasi ... 14
7 Asosiasi haplotipe gen KCNJ11 dan ABCC8 dengan kriteria DMT2 ... 15
DAFTAR LAMPIRAN
Halaman
1 Alur penelitian
... 21
2 Tahap-tahap perancangan primer ... 22
3 Primer untuk deteksi polimorfisme SNP E23K dan A1369S ... 23
ABSTRAK
AGENG WIYATNO. Asosiasi Polimorfisme Gen
KCNJ11 dan
ABCC8 dengan
Diabetes Mellitus Tipe 2 pada Populasi Masyarakat Bali. Dibimbing oleh I
MADE ARTIKA dan SAFARINA GOLFIANI MALIK.
PENDAHULUAN
Diabetes mellitus Tipe 2 (DMT2) merupakan penyakit endemik di dunia (Riedel et al. 2004). Pada tahun 2010, penderita DMT2 di dunia telah mencapai 300 juta jiwa dengan peningkatan sebanyak 7 juta jiwa per
tahun (International Diabetes Federation
/IDF 2011). Badan Kesehatan Dunia (WHO)
memperkirakan penderita DMT2 akan
meningkat dua kali lipat antara tahun 2005 hingga 2030. Lebih dari 80% penderita DMT2 terdapat di negara berkembang dengan angka pertumbuhan yang lebih tinggi daripada di negara maju (WHO 2011).
Indonesia merupakan negara dengan jumlah penderita DMT2 terbanyak keempat setelah India, Cina dan Amerika Serikat (WHO 2011). Berdasarkan hasil Riset Kesehatan Dasar (Riskesdas) tahun 2007, proporsi penyebab kematian akibat DMT2 pada usia 45-54 tahun di daerah perkotaan menduduki peringkat ke-2, yaitu 14,7% dan di daerah pedesaan menduduki peringkat ke-6, yaitu 5,8% dari seluruh penyebab kematian di
Indonesia (Depkes 2010, Mihardja et al.
2009).
Diabetes mellitus Tipe 2 merupakan penyakit kronis yang hingga saat ini belum dapat diobati. Penyakit ini muncul akibat gangguan mekanisme pengaturan glukosa darah berupa ketidakmampuan pankreas memproduksi insulin atau sel target tidak mampu merespon insulin yang diproduksi (IDF 2011). Manifestasi klinis DMT2 dipengaruhi oleh faktor lingkungan dan genetik (O’Rahilly et al. 2005; Gloyn dan McCarthy 2001). Kurangnya aktivitas fisik dan tingginya kadar nutrisi yang dikonsumsi adalah pencetus utama DMT2 (McCarthy 2010; Misra dan Khurana 2008). Secara genetik, DMT2 dipengaruhi oleh interaksi kompleks beberapa gen yang mengatur
metabolisme energi di dalam tubuh.
Polimorfisme yang terjadi pada banyak gen penyandi komponen sel pengatur metabolisme
glukosa berimplikasi secara signifikan
terhadap timbulnya DMT2 (Riedel 2004). Genome Wide Association Study (GWAS) melaporkan lebih dari 20 gen sebagai faktor
resiko penyakit DMT2, diantaranya ABCC8,
KCNJ11, PPARγ, UCP2, TCF7L2, CDKAL1, CDKN2A/B, IGF2BP2, HHEX/IDE, FTO, dan SLC30A8 (Jablonski et al. 2010; Pirie et al. 2010; Zhao et al. 2010; Feng et al. 2008). Mutasi pada gen-gen tersebut berpengaruh terhadap aktivitas komponen sel yang berperan dalam metabolisme glukosa. Salah
satu komponen sel yang penting dalam
metabolisme glukosa adalah KATP channel
pada sel pankreas. Protein integral ini
berfungsi mengatur sekresi insulin (Nichols dan Koster 2002). Kerusakan pada protein ini dapat mengakibatkan hiperinsulinemia dan hiperglikemia. Protein integral KATP channel
disandi oleh dua gen, yaitu KCNJ11 yang
menyandi subdomain Kir6.2 dan ABCC8 yang
menyandi subdomain SUR1 (Florez et al.
2004; Schwanstecher et al. 2002). Studi
genetika pada berbagai populasi menunjukkan bahwa kedua gen tersebut berasosiasi dengan DMT2 (Florez et al. 2004; Gloyn et al. 2004; Nielsen et al. 2003; Sakura et al. 1996).
Analisis genetik dibutuhkan untuk
mengetahui keterlibatan latar belakang
genetik sebagai faktor resiko penyakit DMT2
dalam merancang strategi pencegahan,
penanganan dan pengobatan DMT2. Analisis genetik juga membuka jalan bagi terciptanya metode pengobatan yang spesifik terhadap pasien dan klasifikasi subtipe penyakit yang lebih baik (Vejrazkova dan Bendlova 2005).
Salah satu kriteria diagnosis diabetes mellitus yakni berdasarkan konsentrasi
glukosa darah puasa (Fasting Plasma
Glucose/FPG) (Tabel 1). Pada kondisi normal, konsentrasi FPG berada di bawah 110 mg/dL. Penderita diabetes akan memiliki konsentrasi FPG di atas 126 mg/dL setelah berpuasa selama 8 jam. Individu dengan konsentrasi FPG diantara 110-126 mg/dL perlu
berhati-hati karena termasuk dalam kriteria
hiperglikemia sedang (WHO-IDF 2006). Tujuan penelitian ini adalah mengetahui prevalensi DMT2 pada populasi empat desa terpilih di Bali, yaitu populasi Nusa Ceningan, Legian, Penglipuran dan Pedawa. Penelitian ini juga bertujuan menganalisis asosiasi
polimorfisme E23K pada gen KCNJ11 dan
polimorfisme A1369S pada gen ABCC8
dengan DMT2 berdasarkan keberadaan Single
Nucleotide Polymorphisms (SNPs). Tujuan
selanjutnya dari penelitian ini adalah
menganalisis haplotipe yang terbentuk dari kedua gen tersebut dan asosiasinya dengan DMT2.
Tabel 1 Kriteria individu berdasarkan konsentrasi FPG.
Kelompok Konsentrasi FPG
(mg/dL)
Normal < 110
Hiperglikemia sedang 110-126
Hipotesis penelitian ini adalah prevalensi penyakit DMT2 pada populasi empat desa di
Bali, yaitu Nusa Ceningan, Pedawa,
Penglipuran dan Legian mirip satu sama lain karena latar belakang genetik yang relatif homogen. Nilai prevalensi yang diperoleh berasosiasi dengan polimorfisme E23K pada gen KCNJ11 dan A1369S pada gen ABCC8 sebagai penyandi subdomain KATP channel.
DMT2 di keempat desa berasosiasi dengan
haplotipe tertentu yang dibentuk oleh
polimorfisme kedua gen tersebut karena fungsi kedua gen berkaitan erat satu sama
lain. Penelitian ini bermanfaat dalam
merancang upaya penanganan dan
pencegahan DMT2 di masa depan, khususnya di daerah Bali.
TINJAUAN PUSTAKA
Protein Integral KATP channel dan Mekanisme Sekresi Insulin
Protein integral KATP channel merupakan
protein kanal yang terletak pada sel jantung, sel pankreas, sel syaraf, jaringan otot halus dan ginjal (Minami 2004). Protein ini memiliki fungsi yang spesifik pada tiap
jaringan. KATP channel berperan sebagai
penghubung antara keadaan metabolik dengan
keseimbangan elektrik sel. KATP channel
merupakan protein heterooktamer yang
bersifat uniport terletak di membran sel. Protein ini terdiri atas subunit potassium inwardly rectifying channel (Kir) dan subunit
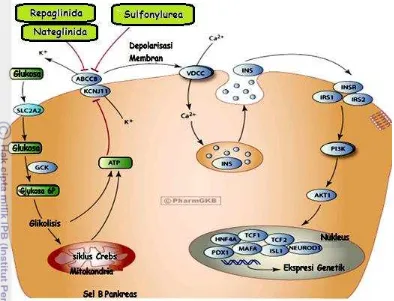
Sulfonylurea Receptor (SUR) seperti yang terlihat pada Gambar 1 (Inagaki et al. 1995).
Protein integral KATP channel berperan
penting dalam berbagai mekanisme
biokimiawi, diantaranya sekresi hormon, penghantaran sinyal neuron, respon sel otot halus dan perlindungan sel jantung dan otak terhadap iskemia (Ashcroft et al. 1998). KATP
channel pada sel pankreas berperan dalam
mengatur jumlah insulin. Adenosin
Triphosphat (ATP) yang dihasilkan dari metabolisme glukosa akan berikatan dengan reseptor yang terdapat pada subunit Kir6.2 menyebabkan penutupan channel dan terjadi depolarisasi membran plasma. Terjadinya
depolarisasi mengaktifkan pembukaan
Voltage Dependent Calcium Channel (VDCC)
yang menyebabkan masuknya Ca2+ melalui
membran sel. Peningkatan konsentrasi Ca2+ di dalam sel akan memicu sekresi granula-granula insulin ke aliran darah.
Produksi insulin akan memicu respon reseptor insulin pada sel target untuk meningkatkan penyerapan glukosa darah (Gong 2001). Sebaliknya, pada saat tidak ada ATP yang tersedia, protein channel ini akan
memompa K+ keluar sel sehingga terjadi
hiperpolarisasi. Pada keadaan tersebut,
penyerapan glukosa berjalan lambat sehingga dapat melindungi sel dari kerusakan akibat kurangnya energi seperti yang terjadi pada iskemik otak dan jantung (Fischer et al. 2008). Mekanisme sekresi insulin dapat diamati pada Gambar 2.
Gambar 1 Struktur KATP channel pada membran sel pankreas.
(A) Letak KATP channel pada membran sel dengan beberapa senyawa aktivator dan
inhibitornya; (B) dan (C)KATP channel merupakan protein herterooktamer yang terdiri atas dua
Kerja KATP channel dikendalikan oleh
perbandingan ATP/ADP dan konsentrasi Mg-ADP di dalam sel (Wasada 2002). Protein KATP channel pada sel pankreas terdiri atas
delapan buah subunit yang dapat dibagi menjadi dua jenis, yaitu Kir6.2 yang berperan sebagai kanal selektif K+ dan SUR1 yang berperan sebagai pengatur kerja protein channel (Sakamoto et al. 2007). Subunit
SUR1 merupakan komponen Sulfonylurea
receptor (SUR) yang menyebabkan protein integral KATP channel dapat dikendalikan
dengan penambahan sulfonylurea (contohnya: tolbutamida dan glibenklamida) (Hansen et al. 1998). Protein-protein subunit KATP channel
(Kir6.2 dan SUR1) disandi oleh gen KCNJ11 dan ABCC8 (Aguilar-Bryan dan Bryan 1999). Keberadaan polimorfisme pada kedua gen dapat mempengaruhi aktivitas KATP channel.
Polimorfisme tersebut berupa Single
Nucleotide Polymorphisms (SNPs) yang dapat
dianalisis dengan berbagai metode,
diantaranya dengan menggunakan metode Polymerase Chain Reaction-Restriction Fragment Length Polymorphisms (PCR-RFLP) (Yokes MB et al. 2001).
Analisis SNPs Menggunakan Metode PCR-RFLP
Single Nucleotide Polymorphisms (SNPs) adalah perbedaan satu basa pada urutan nukleotida tertentu yang muncul secara signifikan (lebih dari 1%) pada populasi (Ke et al. 2008). Polimorfisme tersebut dapat muncul pada daerah ekson maupun intron dengan frekuensi 1/100 basa hingga 1/300
basa (Human Genome Project Information
2011). SNPs yang muncul di daerah ekson dapat mempengaruhi protein yang disandi oleh gen sehingga mengakibatkan perubahan
karakteristik protein yang disandinya.
Perubahan karakteristik ini dilaporkan
berkaitan dengan berbagai penyakit.
Polymerase Chain Reaction (PCR) merupakan metode sintesis asam nukleat secara in vitro untuk mengamplifikasi segmen
DNA secara spesifik berdasarkan
pengulangan siklus termal. Tahap-tahap yang terjadi pada proses PCR yaitu, denaturasi, annealing dan ekstensi. Pada tahap denaturasi, terjadi pemisahan utas ganda DNA menjadi satu utas DNA. Pada tahap annealing primer kemudian menempel pada masing-masing
utas DNA. Dengan bantuan enzim Taq
Gambar 2 Mekanisme sekresi insulin pada sel pankreas.
Glukosa yang masuk ke dalam sel dipecah menghasilkan ATP yang akan berikatan dengan KATP
-channel. Akibatnya, KATP channel menutup dan menyebabkan depolarisasi membran. Depolarisasi
polimerase, DNA akan tersusun di sepanjang utas tunggal sehingga membentuk DNA utas ganda baru (Sambrook et al. 2001).
Restriction Fragment Length Polymorphisms (RFLP) adalah teknik analisis DNA untuk membedakan variasi yang terdapat pada sekuen-sekuen homolog. Prinsip metode ini adalah pemotongan DNA menjadi beberapa bagian oleh enzim restriksi. Letak situs pemotongan enzim restriksi bersifat spesifik terhadap urutan basa nukleotida tertentu yang disebut palindrom.
Potongan-potongan DNA akan terpisah ketika
dielektroforesis berdasarkan ukurannya. Hasil visualisasi potongan-potongan DNA tersebut berupa pita-pita DNA yang dapat diukur
panjangnya menggunakan marka DNA
(Sambrook et al. 2001).
Perbedaan basa pada SNPs menentukan urutan nukleotida yang dapat dikenali oleh enzim restriksi. Jika urutan basa membentuk urutan yang sama dengan situs pengenalan enzim restriksi tertentu, maka nukleotida tersebut akan dipotong oleh enzim restriksi tersebut. Sebaliknya, jika tidak sesuai dengan situs pengenalannya, maka urutan basa tersebut tidak akan dipotong. Akibatnya visualisasi dengan elektroforesis dua jenis alel yang berbeda akan menghasilkan pita dengan ukuran yang berbeda (Sambrook et al. 2001).
Gen KCNJ11 Penyandi Kir6.2
Salah satu gen yang banyak dilaporkan berasosiasi dengan DMT2 adalah gen
penyandi potassium inwardly rectifying
channel (KCNJ11). Gen ini terletak di kromosom 11p.15.1. dengan panjang 4.083 bp dari urutan basa ke 17.406.795 hingga 17.410.878, menyandi 390 asam amino dengan ukuran protein sebesar 43.541 Da. Gen ini menyandi Kir6.2, yaitu subdomain
pada protein integral KATP channel sel
pankreas yang berfungsi sebagai membran selektif K+ (Vejrazkova dan Bendlova 2005).
Salah satu SNPs yang ditemukan pada gen ini adalah polimorfisme E23K (rs5219). Pada polimorfisme tersebut terjadi perubahan basa adenin menjadi guanin di nukleotida ke 1222 yang menyebabkan perubahan asam amino asam glutamat (E) (asam amino negatif) menjadi lisin (K) (asam amino positif) pada kodon ke 23. Perubahan tersebut terjadi di ujung sitosolik N-terminal Kir6.2 yang
menyebabkan penurunan afinitas KATP
channel terhadap ATP sehingga channel terbuka lebih lama dan tidak terjadi depolarisasi membran. Akibatnya, VDCC tidak aktif dan ion kalsium (Ca2+) tidak dapat
masuk ke dalam sel. Tidak adanya kalsium sebagai pemicu sekresi granula insulin
menyebabkan gangguan sekresi insulin
(Riedel et al. 2004, Schwantecher et al. 2002).
Gen ABCC8 Penyandi SUR1
Gen ABCC8 merupakan anggota dari subfamili protein ATP Binding Cassette yang memiliki ukuran DNA sepanjang 4.980 basa dari urutan basa ke 17.414.431 hingga 17.498.448 menyandi 1.581 asam amino dengan ukuran 176.992 Da. Pada gen ini ditemukan sekitar 150 SNPs yang telah dianalisis di berbagai populasi (Campbell et al. 2003). Salah satu SNPs yang terbentuk
pada gen ABCC8 adalah SNP A1369S
(rs757110). Polimorfisme tersebut mengubah basa timin menjadi guanin yang terletak pada nukleotida ke 4321. Perubahan asam amino yang terjadi adalah serin (S) yang bersifat polar menjadi alanin (A) yang bersifat non polar pada kodon 1369 (Feng et al. 2008). Perubahan tersebut menyebabkan penurunan
afinitas SUR1 terhadap Magnesium Adenosin
Diphosphat (MgADP) (Hansen et al. 1998). Meskipun ada penelitian menunjukkan bahwa SNP A1369S tidak berasosiasi dengan DMT2 (Sakamoto et al. 2007), namun SNP A1369S dilaporkan memiliki asosiasi yang kuat
dengan SNP E23K pada gen KCNJ11 dengan
nilai OR= 1,17 (p=0,003) >90% (Florez et al. 2004). Beberapa penelitian melaporkan bahwa pada populasi Kaukasia polimorfisme pada SUR1 berasosiasi dengan neonatal diabetes dan penyakit jantung (Lefer et al. 2009; Giurgea et al 2006; Hansen et al 1997).
Protein subunit yang disandi oleh gen ABCC8 adalah SUR1yang berfungsi mengatur
kinerja KATP channel, dikendalikan oleh
keberadaan MgADP yang akan meningkatkan aktivitas KATP channel. Ikatan antara MgADP
dengan subunit SUR1 menyebabkan K+
terpompa keluar sel dan menurunkan jumlah produksi Ca2+ sehingga mengurangi sekresi insulin (Campbell 2003). Sebaliknya, senyawa sulfonylurea (contohnya: tolbutamida dan
glibenklamida) dan nonsulfonylurea
(contohnya nateglinida dan repaglinida) dapat mengaktifkan SUR1 melalui pembentukan ikatan dengan reseptor sulfonylurea. Ikatan tersebut menyebabkan protein KATP channel
Desa dan Penduduk Nusa Ceningan, Legian, Penglipuran dan Pedawa Bali
Bali merupakan provinsi yang terletak di sebelah selatan Indonesia, tepatnya di 8°3'40"-8°50'48" Lintang Selatan dan 114°25'53"-115°42'40" Bujur Timur. Provinsi Bali terbagi menjadi 9 kabupaten dan 55 kecamatan. Berdasarkan relif dan topografi, bagian utara Bali terdiri atas gunung-gunung dan dataran tinggi, sedangkan bagian selatan berupa dataran rendah yang dialiri sungai-sungai dan pulau-pulau kecil. Sekitar 92,3% penduduk Bali memeluk agama Hindu yang membuat penduduk Bali cenderung menikah dengan orang sesama Bali. Oleh karena itu, diduga bahwa populasi Bali cenderung bersifat homogen secara genetik. Namun, khusus untuk daerah pesisir, lebih besar kemungkinan untuk terpapar pengaruh genetik lain karena letaknya yang di pesisir.
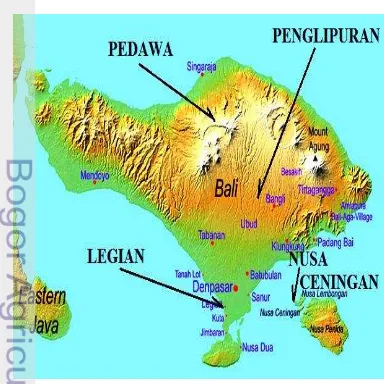
Nusa Ceningan, Legian, Pedawa dan Penglipuran merupakan desa yang terletak di Provinsi Bali, Indonesia (Gambar 3). Keempat desa ini memiliki karakteristik yang berbeda dari segi topografi maupun kebudayaannya (Pemerintah Provinsi Bali 2010). Nusa Ceningan adalah sebuah pulau kecil yang terletak di daerah selatan Bali di Kabupaten
Klungkung. Mayoritas penduduk Nusa
Ceningan berprofesi sebagai nelayan, petani rumput laut dan pedagang. Masyarakat Desa Nusa Ceningan lebih terbuka terhadap turis karena Nusa Ceningan terletak dekat dengan tempat wisata Bali. Namun, dampak dari turisme terhadap gaya hidup masyarakat Nusa Ceningan belum banyak diketahui.
Gambar 3 Letak empat desa terpilih di Bali Pedawa dan Penglipuran berada di dataran tinggi, sedangkan Legian dan Nusa Ceningan berada di pesisir pantai.
Desa Legian terletak di Kecamatan Kuta, Kabupaten Badung, Bali. Desa Legian merupakan desa yang terletak di pesisir pantai sebelah barat daya Pulau Bali, terletak dekat dengan daerah wisata Pantai Kuta. Penduduk di Desa Legian mayoritas berprofesi sebagai pedagang dan buruh. Desa ini berkembang menjadi salah satu tujuan wisata yang cukup diminati oleh turis. Dampak turisme terhadap gaya hidup masyarakat Legian sangat terlihat, karena masyarakat di sini telah terpapar westernisasi selama setidaknya 20 tahun (Malik et al. 2011).
Berbeda dengan Nusa Ceningan dan Legian, Penglipuran dan Pedawa merupakan
desa yang terletak jauh dari pantai.
Penglipuran merupakan desa yang terletak di bagian tengah Pulau Bali, Kecamatan Kubu, Kabupaten Bangli, Bali. Desa ini terletak di kaki Gunung Batur yaitu sekitar 500-700 meter di atas permukaan laut. Penduduk Penglipuran mayoritas berprofesi sebagai petani, hal ini terlihat dari topografi Desa
Penglipuran yang berupa subak/sawah
berundak-undak. Desa Penglipuran
merupakan salah satu tujuan wisata di Bali karena letaknya yang dekat dengan Denpasar (Murni 2009).
Pedawa terletak di Kecamatan Banjar, Kabupaten Buleleng Bali. Desa tersebut terletak di dataran tinggi dan letaknya jauh dari tempat wisata. Berbeda dengan penduduk di tiga desa lainnya, penduduk di populasi Pedawa cenderung terisolasi dari kehidupan modern dan turisme. Mayoritas penduduk di Desa Pedawa berprofesi sebagai petani dan buruh. Potensi pertanian di desa ini sangat baik. Hal ini terlihat dari organisasi desa yang berkembang pesat berupa subak. Kebudayaan di Desa Pedawa masih terus dijaga dan dilestarikan (Pemerintah Kabupaten Buleleng 2009).
BAHAN DAN METODE
Bahan dan Alat
Bahan yang digunakan dalam penelitian ini terbagi menjadi kelompok bahan untuk tahap isolasi DNA, PCR, RFLP dan elektroforesis. Bahan-bahan untuk tahap isolasi DNA terbagi menjadi dua, yaitu untuk isolasi DNA dari darah dan isolasi DNA dari Guthrie Cards. Bahan-bahan untuk isolasi DNA dari darah adalah: larutan pelisis sel darah merah (NH4Cl [Merck, France]; EDTA
[BDH, Canada]; KHCO3 [Merck, France]);
dan SDS [Sigma, USA]; enzim RNase (5mg/µL) [Qiagen, Germany]; ammonium asetat 5 M; isopropanol [Malinckrodt, USA]; etanol 70% [Malinckrodt, USA]; dan buffer Tris-EDTA yang mengandung Tris-HCL 10 mM pH 7,5 [Invitrogen, USA], EDTA 1 mM [BDH, Canada]. Bahan yang digunakan untuk
isolasi darah dari Guthrie Cards adalah
larutan PBS (Phosphate Buffer Saline) yang terdiri atas NaCl, KCl, Na2HPO4, KH2PO4
[Merck, France] yang dilarutkan ke dalam ddH2O, saponin 5% dan larutan Chelex 20%
dibuat dari Chelex 100% [Bio-Rad, USA]. Bahan-bahan untuk teknik PCR terdiri
atas: PCR buffer (Tris-HCL 10 mM
[Invitrogen, USA] dan KCl 50 mM [BDH,
Canada]); 1,5 mM MgCl2 [Merck, France];
campuran dNTP 10 mM (dATP, dGTP, dCTP, dan dTTP) [Invitrogen, USA]; masing-masing primer forward dan reverse SNPs rs5219 dan SNPs rs727110 sebanyak 4 pmol/µL, [1st Base Singapore]; 1,25 unit enzim Taq DNA polimerase [Gibco-BRL, USA].
Bahan-bahan tahap RFLP adalah ultrapure
ddH2O, enzim BanII 5u/µL, enzim MwOI
5u/µL, NEB buffer 3 dan NEB buffer 4
(terdiri atas 50 mM NaCl; 10 mM Tris –HCl;
10 mM MgCl2; dan 1 mM dithiothreitol pH
7,9); 100 ug/mL BSA pH 7,4 [New England Biolabs, USA]. Bahan untuk elektroforesis adalah: bubuk agarosa [Seakem LE Agar, USA]; etidium bromida (EtBr) 10 mg/mL
[Sigma, USA]; buffer Tris-borat EDTA (TBE)
1x (terdiri atas Tris [Invitrogen, USA]; asam borat [Merck, France]; dan EDTA [BDH,
Canada]); loading buffer 6x (terdiri atas
bromofenol biru 0,25% [Merck, France]; dan sukrosa 40% (b/v) [Merck, France]).
Alat-alat yang digunakan pada penelitian ini adalah vorteks [Thermolyne 37600, USA]; mesin sentrifugasi [Eppendorf 5415C dan Sorvall RT 6000D, Germany]; neraca digital [Sartorius AC 121S, Germany]; inkubator waterbath [Forma Scientific, USA];
spektrofotometer [Nanodrop ND-1000
V3.5.2., UK]; mesin thermal cycler
[GeneAmpR PCR System 9700, USA];
thermomixer Eppendorf 1.5 mL; aparatus elektroforesis wide-mini sub cellR GT Rad, USA]; sumber arus listrik [Bio-Rad,USA]; lemari es [National dan Forma Scientific, USA]; tabung mikrosentrifus(0,2; 0,5, dan 1,5 mL) [Molecular Bio Product, CA]; gelas ukur (25 mL dan 250 mL) [Duran, Germany]; labu Erlenmeyer 250 mL [Duran, Germany]; tabung sentrifugasi (15 mL dan 10 mL) [Falcon, USA]; mikropipet [Eppendorf dan Finpippett, Germany].
Metode Pengambilan Sampel Darah
Sampel darah manusia diambil dari empat desa di Bali, yaitu Desa Nusa Ceningan,
Legian, Penglipuran dan Pedawa
menggunakan metode potong lintang ( cross-sectional), yaitu hanya satu kali pengambilan sampel. Sampel yang diambil dari Desa Nusa Ceningan, Pedawa dan Penglipuran berupa sampel darah utuh, sedangkan sampel dari Desa Legian berupa darah yang diteteskan
pada permukaan Guthrie Cards dan
didiamkan hingga mengering. Pengambilan sampel dilakukan oleh dokter dan peneliti dari RS Sanglah/Fakultas Kedokteran Udayana dan Lembaga Eijkman dengan persetujuan relawan di populasi Bali. Pengambilan sampel telah mendapatkan izin dari Komisi Etik Fakultas Kedokteran Universitas Udayana dan Komisi Etik Riset Lembaga Eijkman (Malik et al. 2011).
Pengambilan sampel dilakukan untuk mengetahui karakteristik dan sebaran sampel secara umum sehingga asosiasi antara karakteristik sampel dengan kriteria DMT2 dapat dianalisis. Beberapa karakteristik klinis diambil dari tiap relawan, yaitu: jenis kelamin, usia dan konsentrasi glukosa puasa relawan sebagai parameter DMT2. Sebanyak 46 sampel dipilih dari tiap desa dengan jenis
kelamin dan usia yang dicocokkan (matched).
Sampel-sampel tersebut dikelompokkan
menjadi kelompok usia <35 tahun, 36-50 tahun, 51-65 tahun dan >65 tahun.
Sampel yang digunakan pada penelitian ini adalah sampel DNA yang diisolasi dari darah manusia berusia di atas 25 tahun. Sebanyak 2 mL darah diisolasi dari pembuluh darah perifer relawan kemudian disimpan pada suhu 4°C hingga tahap isolasi DNA dilakukan, sedangkan untuk sampel Desa Legian, darah diteteskan di permukaan Guthrie Cards dan
dibiarkan mengering. Sampel tersebut
dimasukkan ke dalam amplop pada suhu ruang hingga proses isolasi DNA dilakukan.
Perancangan Primer dan Pemilihan Enzim Restriksi
Primer yang digunakan pada penelitian ini terdiri atas dua pasang primer, satu pasang untuk mendeteksi SNP E23K pada gen KCNJ11 dan satu pasang lainnya untuk
mendeteksi SNP A1369S pada gen ABCC8.
Tahap-tahap perancangan primer dilakukan dengan bantuan software Primer3, REHelper, NetPrimer, Blastn, PCRproducts dan Restriction Digest. Pemilihan enzim
restriksi dilakukan bersamaan dengan
perancangan primer. Tahap perancangan primer diuraikan dalam Lampiran 2. Primer yang digunakan adalah sebagai berikut:
- SNP E23K pada gen KCNJ11
Forward Primer :
5-GACTCTGCAGTGAGGCCCTA-3 Reverse Primer :
5-ACGTTGCAGTTGCCTTTCTT-3
- SNP A1369S pada gen ABCC8
Forward Primer :
5-CGCTACGACAGCTCCCTGAAG-3 Reverse Primer :
5-GTCTCCTTGGTGGATGAGTGAG-3 Enzim restriksi yang digunakan untuk
memotong produk PCR gen KCNJ11 adalah
BanII, dengan situs restriksi : 5’....GRGCYˇC....γ’ γ’....CˆYCGRG....5’
Enzim restriksi yang digunakan untuk
memotong produk PCR gen ABCC8 adalah
MwoI, dengan situs restriksi : 5’....GCNNNNNˇNNGC....γ’ γ’....CGNNˆNNNNNCG....5’ Keterangan :
R=A/G, Y=C/T, N= A/T/G/C; = Letak polimorfisme
Isolasi DNA
a) Metode PuregeneR yang Telah Dimodifikasi (2003)
Sebanyak 6 mL larutan pelisis sel darah merah dicampur dengan 2 mL darah di dalam tabung Falcon™ bervolume 10 mL. Tabung tersebut dibolak-balik sebanyak 2-3 kali kemudian diinkubasi pada suhu ruang (15-25°C) selama 10 menit. Setelah itu, dilakukan sentrifugasi selama 10 menit pada 1500 rpm (Sorvall RT 6000D) untuk mengendapkan sel darah putih. Supernatan dalam tabung dibuang dan dua tahap terakhir diulang tanpa melalui proses inkubasi. Pelet sel darah putih yang
diperoleh kemudian divorteks hingga
homogen dan ditambahkan 500 µL larutan pelisis sel darah putih. Campuran kemudian dihomogenasi kembali dengan menggunakan pipet transfer.
Sebanyak 2 µL enzim RNase A 5 mg/mL
dicampurkan ke dalam larutan kemudian dihomogenasi dan diinkubasi di suhu 37°C
dalam shaker waterbath selama 45 menit.
Sebanyak 334 µL amonium asetat 5 M dimasukkan ke dalam larutan agar terjadi
presipitasi protein. Tabung divorteks hingga homogen kemudian disentrifugasi pada 3.000 rpm (Sorvall RT 6000D) dengan suhu 4°C selama 15 menit.
Supernatan dengan volume ±800 µL yang mengandung DNA dituang ke dalam tabung Falcon™ baru berukuran 15 mL yang berisi 1.540 µL isopropanol. Isopropanol akan
mengikat air sehingga menyebabkan
koagulasi DNA yang dapat diamati berupa gumpalan-gumpalan berwarna putih. Larutan dibolak-balik sebanyak 25-30 kali hingga pelet DNA terlihat. Sentrifugasi dilakukan kembali dengan kecepatan 3000 rpm (Sorvall RT 6000D) pada suhu 4°C selama 15 menit. Supernatan yang berisi pengotor dibuang, kemudian dilakukan pencucian dengan 166 µL etanol 70%. Tabung disentrifugasi kembali, kemudian supernatan berupa etanol
70% dibuang. Pelet DNA kemudian
dikeringkan pada suhu ruang selama semalam. Pelet tersebut selanjutnya direhidrasi dengan menambahkan 100 µL buffer TE dan
diinkubasi pada suhu 37°C dalam shaker
waterbath selama 2 jam. Pelet DNA yang telah dilarutkan disimpan pada suhu -20°C.
b) Isolasi DNA dari GuthrieCards
Daerah pada permukaan Guthrie Cards
yang mengandung darah keringdipotong kecil
dengan ukuran kira-kira 0,7x1cm. Potongan-potongan tersebut dimasukkan ke dalam tabung Eppendorf 1,5 mL. Saponin 0,5% ditambahkan ke dalam tabung Eppendorf tersebut sebanyak 1 mL. Tabung dibolak-balik beberapa kali lalu diinkubasi selama semalam pada suhu 4°C. Setelah melalui proses inkubasi, tabung disentrifugasi pada kecepatan 12000 rpm (Sorvall RT 6000D) selama 5 detik kemudian supernatan dibuang. Selanjutnya, ke
dalam tabung ditambahkan Phosphate Buffer
Pengukuran Konsentrasi DNA
Pengukuran konsentrasi DNA dilakukan dengan prinsip spektrofotometri berdasarkan
Sambrook et al. (2001). Pengukuran
dilakukan dengan spektrofotometer Nanodrop ND-1000 V3.5.2 pada panjang gelombang 260 nm untuk DNA dan 280 nm untuk protein pengotor. Nilai perbandingan antara DNA dengan protein pengotor (A260/A280) berada diantara 1,8-2,0 (Sambrook et al. 2001). Setelah konsentrasi DNA diketahui, sampel DNA diencerkan menjadi 50 ng/µ L dengan menambahkan Ultrapure ddH2O.
Amplifikasi Gen dengan Polymerase Chain Reaction (PCR)
Bahan campuran reaksi PCR memiliki total volume 25 µL dalam tiap tabung, bahan tersebut terdiri atas ddH2O; PCR buffer 1x;
MgCl2 1,5 mM; dNTP 0,2 mM; forward
primer 4 pmol; reverse primer 4 pmol; 0,5
unit Taq polimerase; dan 50 ng DNA sampel.
DNA tersebut diamplifikasi di mesin PCR yang sudah diatur waktu dan suhunya seperti
ditampilkan pada Gambar 3. Fase
predenaturasi dilakukan pada suhu 95°C selama 5 menit, dilanjutkan dengan tahap denaturasi pada suhu yang sama selama 1 menit. Tahap annealing terjadi pada suhu 60°C selama 30 detik, kemudian diikuti tahap ekstensi pada suhu 72°C selama 1 menit.
Tahap denaturasi, annealing dan ekstensi
diulang sebanyak 30x, diakhiri dengan tahap pos ekstensi pada suhu 72°C selama 5 menit.
Gambar 4 Pengaturan suhu dan waktu yang digunakan pada reaksi PCR gen KCNJ11 dan ABCC8.
Elektroforesis Produk PCR
Produk PCR dielektroforesis
menggunakan gel agarose yang terlarut dalam buffer (TBE) 1X hingga mencapai konsentrasi
1% menggunakan microwave oven. Setelah
agarose terlarut dengan sempurna, selanjutnya
dilakukan penambahan EtBr sebanyak 5 µL
ke dalam larutan tersebut. Proses
elektroforesis dilakukan pada tegangan 100 V selama 30 menit. Jumlah DNA yang diisikan ke dalam sumur elektroforesis adalah 4 µL dengan loading buffer sebanyak 2 µL. Hasil PCR yang telah dikonfirmasi kemudian dipotong dengan enzim restriksi.
Restriction Fragment Length Polymorphisms
(RFLP)
Produk PCR dipotong dengan enzim restriksi. Perbedaan satu basa nukleotida atau lebih menyebabkan enzim tidak dapat mengenali situs pemotongannya. Karakteristik ini merupakan prinsip dasar deteksi SNPs dengan metode RFLP. Enzim restriksi akan memotong SNPs dengan jenis alel tertentu,
namun tidak memotong alel lainnya.
Akibatnya, produk PCR dengan polimorfisme yang berbeda akan memiliki ukuran produk restriksi yang berbeda.
Produk PCR gen KCNJ11 didigesti dengan
3 unit enzim BanII untuk mendeteksi SNP
E23K yang dicampur dalam larutan buffer
NEB 4. Produk PCR gen ABCC8 didigesti
dengan 1 unit enzim MwoI untuk mendeteksi SNP A1369S yang dicampur di dalam buffer NEB 3. Produk PCR yang akan didigesti diinkubasi di dalam thermomixer selama 7 jam pada suhu inkubasi 37°C untuk gen KCNJ11 dan 16 jam (overnight) pada suhu
60°C untuk gen ABCC8. Hasil restriksi
kemudian dielektroforesis pada gel agarose 2,5% dengan tegangan 80 V selama 1 jam
untuk gen KCNJ11, sedangkan untuk gen
ABCC8 digunakan gel 3% dengan tegangan 80 V selama 1,5 jam. Pada saat elektroforesis, seluruh DNA hasil restriksi diisikan ke dalam well agarose yang dicampur dengan loading buffer sebanyak 2 µL. Hasil elektroforesis menunjukkan beberapa pita yang dapat
diidentifikasi ukurannya dengan
menggunakan marker DNA ϕx174RTDNA
yang dipotong dengan enzim HaeIII.
Analisis Statistik
Analisis statistik dilakukan menggunakan perangkat lunak R-Statistics 2.13.0
(R-project.org©2011). Hasil analisis yang
signifikan diperoleh pada p-value <0.05.
Kesesuaian frekuensi genotipe dengan
kesetimbangan Hardy-Weinberg (HWE) diuji menggunakan Fisher’s Exact Test. Asosiasi
genotipe dengan DMT2 diuji dengan
Chi-Square test. Analisis haplotipe dilakukan
dengan Expectation Maximization Algorithms
HASIL DAN PEMBAHASAN
Karakteristik Klinis dan Sebaran Sampel Secara Umum
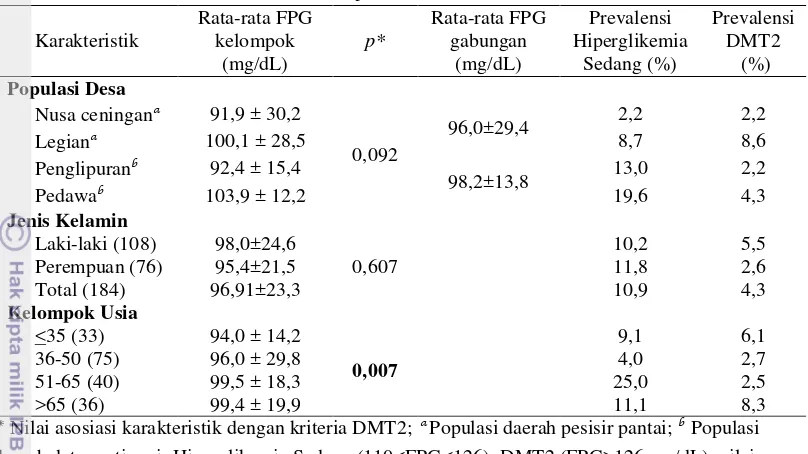
Penelitian ini menggunakan 184 sampel darah yang diperoleh dari 108 orang relawan laki-laki dan 76 orang relawan perempuan pada rentang usia 26-81 tahun pada populasi Bali. Sebaran konsentrasi FPG berkisar antara 60 mg/dL hingga 283 mg/dL. Nilai rata-rata konsentrasi FPG populasi total adalah 96,91 mg/dL dengan rata-rata konsentrasi tertinggi dimiliki oleh Desa Pedawa (103,9 mg/dL), sedangkan nilai rata-rata konsentrasi FPG terendah dimiliki oleh Desa Nusa Ceningan (91,9 mg/dL) (Tabel 2).
Hasil analisis statistik pada tiap desa menunjukkan tidak ada perbedaan yang signifikan antara nilai rata-rata konsentrasi
FPG satu desa dengan desa lainnya (p=0,092)
(Tabel 2). Nilai rata-rata konsentrasi FPG di populasi pesisir (Nusa Ceningan dan Legian) sebesar 96,0 mg/dL, lebih rendah daripada rata-rata konsentrasi FPG populasi dataran tinggi (Penglipuran dan Pedawa) sebesar 98,2 mg/dL, namun tidak berbeda secara signifikan (p=0,452). Nilai standar deviasi gabungan daerah pesisir (Nusa Ceningan dan Legian) dua kali lebih tinggi daripada populasi dataran tinggi (Penglipuran dan Pedawa). Artinya, konsentrasi FPG populasi di daerah pesisir pantai lebih bervariasi dibandingkan dataran tinggi. Hal ini mungkin disebabkan oleh pengaruh turisme yang lebih aktif di Legian dan Nusa Ceningan, sehingga mempengaruhi gaya hidup masyarakat di daerah tersebut. Kemungkinan lain penyebab tingginya variasi FPG adalah faktor makanan yang relatif lebih bervariasi di daerah pesisir bila dibandingkan dengan daerah dataran tinggi. Analisis asosiasi letak geografis (pesisir pantai dan dataran tinggi) dengan kriteria DMT2 tidak signifikan (p=0,052).
Hasil analisis konsentrasi FPG
menunjukkan nilai prevalensi DMT2 total di empat populasi Bali adalah 4,3% (Tabel 2) lebih rendah dibandingkan dengan prevalensi DMT2 di Indonesia yaitu 5,7%. Prevalensi hiperglikemia sedang pada populasi total adalah 10,9%, mendekati prevalensi nasional 10,7% (Depkes 2010). Tingginya prevalensi hiperglikemia sedang perlu diwaspadai karena satu dari tiga kasus hiperglikemia sedang akan berkembang menjadi DMT2 dalam kurun
waktu 5 tahun (Mihardja et al. 2009).
Berdasarkan prevalensi hiperglikemia sedang, dapat diperkirakan sekitar 10-15 tahun mendatang jumlah penderita diabetes akan
meningkat sebanyak dua kali lipat di keempat populasi Bali. Prediksi tersebut diperoleh berdasarkan perhitungan berikut ini :
Prevalensi DMT2 2x lipat = 2x 4,3% = 8,6%
Kurun Waktu Prevalensi IH DMT2
5 tahun 1/3 x 10,9% = 3,6%
10 tahun 2(1/3 x 10,9%) = 7,2%
15 tahun 3(1/3 x 10,9%) = 10,8%
7,2%< prev. DMT2 2x lipat (8,6%)< 10,8% Prevalensi DMT2 di masing-masing populasi Nusa Ceningan (2,2%), Legian (8,6%) Penglipuran (2,2%) dan Pedawa (4,3%). Prevalensi hiperglikemia sedang di Desa Nusa Ceningan (2,2%), Legian (8,7%), Penglipuran (13%) dan Pedawa (19,6%) (Tabel 2).
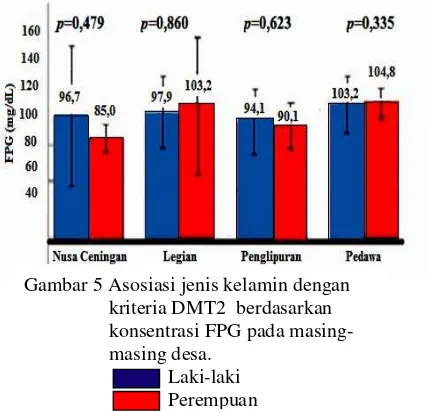
Asosiasi Jenis Kelamin dengan Kriteria DMT2
Analisis asosiasi jenis kelamin dengan kriteria DMT2 pada populasi total tidak menunjukkan perbedaan yang signifikan (p=0,607) (Tabel 2). Perbandingan rata-rata konsentrasi FPG tiap jenis kelamin pada
masing-masing desa ditampilkan pada
Gambar 5. Asosiasi jenis kelamin dengan kriteria DMT2 tidak berbeda secara signifikan baik pada populasi total (p=0,607) maupun di
tiap-tiap desa (Nusa Ceningan p=0,479);
Legian (p=0,860); Penglipuran (p=0,623); dan
Pedawa (p=0,335) (Gambar 5). Berbeda
dengan penelitian lain yang menyatakan
bahwa di Indonesia, kriteria DMT2
berasosiasi dengan jenis kelamin perempuan sebanyak 1,2-1,4 kali lebih tinggi daripada laki-laki (Mihardja et al. 2009). Perbedaan ini mungkin disebabkan oleh jumlah sampel yang sedikit (n=184), sehingga tidak mewakili seluruh populasi.
Gambar 5 Asosiasi jenis kelamin dengan kriteria DMT2 berdasarkan konsentrasi FPG pada masing-masing desa.
Tabel 2 Karakteristik klinis dan sebaran sampel secara umum
* Nilai asosiasi karakteristik dengan kriteria DMT2;a
Populasi daerah pesisir pantai; b
Populasi daerah dataran tinggi; Hiperglikemia Sedang (110<FPG<126); DMT2 (FPG>126 mg/dL); nilai p yang bercetak tebal menunjukkan adanya perbedaan yang signifikan (p<0,05); Semua nilai p
dihitung dengan menggunakan Chi-square test.
Asosiasi Kelompok Usia Tertentu dengan Kriteria DMT2
Sampel yang diperoleh dari keempat desa dibagi menjadi 4 kelompok usia, yaitu <35 tahun (17,9%), 36-50 tahun (40,7%), 51-65 tahun (21,7%) dan >65 tahun (19,6%). Karakteristik sampel berdasarkan kelompok usia dan jenis kelamin tiap desa ditampilkan pada Tabel 3. Sampel yang dianalisis pada penelitian ini banyak tersebar pada rentang usia 36-50 tahun, berjumlah 75 sampel (40,7%). Asosiasi kelompok usia dengan kriteria DMT2 menunjukkan bahwa jumlah penderita hiperglikemia sedang di kelompok usia 51-65 tahun berbeda secara signifikan dari kelompok usia lainnya (p=0,007) (Tabel 2). Perbedaan yang signifikan juga terlihat di populasi Legian (p=0,024), dan populasi laki-laki di Desa Penglipuran (p=0,027). Namun, pada tiga desa lainnya distribusi konsentrasi FPG relatif seragam pada semua kelompok usia (Nusa Ceningan (p=0,452); Penglipuran (p=0,088); dan Pedawa (p=0,071). Hasil penelitian ini berbeda dengan penelitian
sebelumnya (Mihardja et al. 2009) yang
menyatakan bahwa prevalensi DMT2 di Indonesia meningkat pesat pada kelompok usia 35-44 tahun. Tingginya prevalensi hiperglikemia sedang pada kelompok usia 51-65 tahun disebabkan oleh menurunnya aktivitas sel pankreas (Biggs et al. 2010, Mihardja et al. 2009).
Prevalensi DMT2 relatif tinggi pada kelompok usia <35 tahun (6,1%) dan >65 tahun (8,3%). Tingginya prevalensi DMT2 di usia >65 tahun merupakan hal yang wajar sebagai akibat menurunnya efektivitas sel
pankreas dalam memproduksi insulin
sehingga banyak orang lanjut usia mengalami DMT2. Namun, tingginya prevalensi DMT2 di usia <35 tahun mungkin disebabkan perubahan gaya hidup penduduk usia muda sebagai dampak westernisasi. Nilai prevalensi DMT2 di usia <35 tahun merupakan kontribusi dari Desa Penglipuran dan Legian. Desa Legian memang telah dipengaruhi westernisasi setidaknya selama 20 tahun. Hal ini berdampak pada perubahan gaya hidup dan tingkat sosial-ekonomi yang relatif lebih tinggi daripada tiga desa lainnya. Pekerjaan penduduk Legian yang tadinya nelayan dan petani berubah menjadi pekerja hotel dan pedagang sehingga aktivitas fisik mereka berkurang. Makanan yang mereka konsumsi banyak mengandung karbohidrat tinggi, kaya lemak jenuh dan kolesterol (Malik et al. 2011). Berbeda dengan penduduk di Desa Pedawa, Penglipuran dan Nusa Ceningan yang belum terpengaruh oleh westernisasi. Kehidupan mereka relatif lebih sederhana dengan gaya hidup yang masih tradisional. Perubahan gaya hidup dan tingkat sosial-ekonomi yang lebih tinggi berkorelasi dengan
meningkatnya prevalensi DMT2 (Mihardja et
al. 2009). Karakteristik Rata-rata FPG kelompok (mg/dL) p* Rata-rata FPG gabungan (mg/dL) Prevalensi Hiperglikemia Sedang (%) Prevalensi DMT2 (%) Populasi Desa
Nusa ceningana
91,9 ± 30,2
0,092
96,0±29,4 2,2 2,2
Legiana
100,1 ± 28,5 8,7 8,6
Penglipuranb
92,4 ± 15,4
98,2±13,8 13,0 2,2
Pedawab
103,9 ± 12,2 19,6 4,3
Jenis Kelamin
Laki-laki (108) 98,0±24,6
0,607
10,2 5,5
Perempuan (76) 95,4±21,5 11,8 2,6
Total (184) 96,91±23,3 10,9 4,3
Kelompok Usia
<35 (33) 94,0 ± 14,2
0,007
9,1 6,1
36-50 (75) 96,0 ± 29,8 4,0 2,7
51-65 (40) 99,5 ± 18,3 25,0 2,5
Tabel 3 Asosiasi kelompok usia dan jenis kelamin dengan kriteria DMT2 pada masing-masing desa
nilai p yang bercetak tebal menunjukkan adanya perbedaan yang signifikan (p<0,05); Semua nilai p
dihitung dengan menggunakan Chi-square test.
Desa Usia Frekuensi (%)
Rata-rata FPG (mg/dL) P Rata-rata FPG gabungan (mg/dL) Prevalensi Hiperglike mia Sedang (%) Prevalensi DMT2 (%) Nusa Ceningan
Total <35 17,4 84,5+4,7
0,452 91,9 + 30,2
0 0
(n=46) 36-50 39,1 96,9+46,9 0 2,2
51-65 23,9 87,9+9,6 0 0
>65 19,6 93,0+13,7 2,2 0
Laki-laki <35 8,7 87,0+5,3
0,535 96,7+38,6
0 0
(n=27) 36-50 19,6 106,0+59,1 0 3,7
51-65 13,0 89,7+11,4 0 0
>65 13,0 93,0+16,7 3,7 0
Perempuan <35 8,7 82,0+2,4
- 85,0+6,31
0 0
(n=19) 36-50 15,2 82,7+4,6 0 0
51-65 10,9 85,8+7,5 0 0
>65 6,5 93,0+6,0 0 0
Legian
Total <35 17,4 93,9+17,9
0,024 100,1+ 28,5
0 2,2
(n=46) 36-50 34,8 100,7+35,8 2,2 2,2
51-65 21,7 107,3+24,4 6,5 2,2
>65 19,6 96,1+24,7 0 2,2
Laki-laki <35 8,6 90,8+10,1
0,096 97,9+20,7
0 0
(n=27) 36-50 23,9 91,8+6,0 0 0
51-65 13,0 110,7+29,7 7,4 4,8
>65 13,0 101,9+29,8 0 4,8
Perempuan <35 8,6 97,0+24,9
0,541 103,2+37,3
0 5,3
(n=19) 36-50 10,9 113,0+54,3 5,3 5,3
51-65 8,6 102,2+15,9 5,3 0
>65 6,5 86,3+2,1 0 0
Penglipuran
Total <35 19,6 96,6+16,9
0,088 92,4 + 15,4
2,2 2,2
(n=46) 36-50 41,3 86,3+9,8 0 0
51-65 19,6 96,7+19,0 6,5 0
>65 19,6 97,0+17,8 4,3 0
Laki-laki <35 8,6 102,3+20,4
0,027 94,1+17,3
0 3,7
(n=27) 36-50 23,9 85,5+11,9 0 0
51-65 13,0 98,2+23,9 11,1 0
>65 13,0 100,5+12,8 3,7 0
Perempuan <35 10,9 92,0+14,2
0,330 90,1+12,3
5,3 0
(n=19) 36-50 17,4 87,5+6,3 0 0
51-65 6,5 93,7+1,5 0 0
>65 6,5 90,0+27,4 5,3 0
Pedawa
Total <35 17,4 100,6+9,6
0,071 103,9 + 12,2
4,3 0
(n=46) 36-50 41,3 100,2+6,8 4,3 0
51-65 21,7 106,8+11,5 8,1 0
>65 19,6 111,3+19,7 2,2 4,3
Laki-laki <35 8,6 93,3+7.8
0,197 103,2+14,9
0 0
(n=27) 36-50 23,9 99,5+8,3 7,4 0
51-65 13,0 102,7+11,6 3,7 0
>65 13,0 117,2+22,4 3,7 7,4
Perempuan <35 8,6 108,0+2,9
0,054 104,8+7,2
10,5 0
(n=19) 36-50 17,4 101,1+4,5 0 0
51-65 8,6 113,0+9,5 15,7 0
EE EK KK SS SA AA
Tabel 4 Nilai Frekuensi Alel Minor (MAF) gen KCNJ11 pada masing-masing populasi
Angka bercetak tebal menunjukkan nilai yang signifikan p<0,05 (Tidak memenuhi HWE)
Deteksi SNP E23K Gen KCNJ11 dan Gen
ABCC8
Deteksi SNP E23K dan SNP A1369S telah berhasil dilakukan dengan mengamplifikasi gen menggunakan reaksi PCR kemudian mendigesti produk PCR tersebut dengan enzim restriksi yang spesifik. Representasi
deteksi polimorfisme SNP E23K gen KCNJ11
ditampilkan pada Gambar 6 (kiri). Genotipe EE menghasilkan tiga pita, masing-masing berukuran 150bp, 32bp dan 28bp. Genotipe EK menghasilkan empat pita, masing-masing berukuran 178bp, 150bp, 32bp dan 28bp. Genotipe KK menghasilkan dua pita,
masing-masing berukuran 178bp dan 32bp.
Representasi deteksi polimorfisme SNP A1369S ditampilkan pada Gambar 6 (kanan). Genotipe SS menghasilkan 3 pita masing-masing berukuran 203bp, 92bp dan 8bp. Genotipe SA menghasilkan lima pita, masing- masing berukuran 203bp, 92bp, 51bp, 41bp dan 8 bp. Genotipe AA menghasilkan empat pita, masing-masing berukuran 203bp, 51bp, 41bp dan 8bp. Pita-pita berukuran 32bp, 28bp dan 8bp tidak terlihat karena ukurannya kecil. Meskipun begitu, keberadaan masing-masing SNPs dapat dibedakan dengan jelas melalui hasil representasi deteksi SNPs. Pemisahan fragmen berukuran kecil (<50bp) dapat terlihat dengan jelas jika menggunakan gel poliakrilamida (Sambrook et al. 2001).
Frekuensi Alel Minor (Minor Allele
Frequency, MAF) merupakan alel dengan frekuensi paling rendah pada populasi (Foulkes 2009). Hasil analisis populasi total menunjukkan FAM SNP E23K (alel K) di populasi Bali adalah 27% (Tabel 4) sedangkan MAF SNP A1369S (alel A) adalah 29% (Tabel 5). MAF kedua SNPs pada penelitian ini lebih rendah daripada populasi Kaukasia (37% dan 39%), Jepang (37% dan 39%) dan Cina (38% dan 43%), namun tidak serendah populasi Afro-Amerika (8% dan 8%) (Palmer et al 2008; Florez et al 2007; Sakamoto et al
2007;Hani
et al 1998). Perubahan frekuensi alel dalam suatu populasi akibat mutasi menyebabkan munculnya frekuensi alel-alel yang khas dan tetap dipertahankan dari generasi ke generasi. Hal ini yang menyebabkan perbedaan frekuensi alel-alel minor dari SNPs yang sama bervariasi di populasi yang berbeda (Li dan Graur 2001).
MAF SNP E23K gen KCNJ11 dan SNP
A1369S gen ABCC8 di tiap desa mirip satu sama lain, indikasi bahwa keempat desa memiliki latar belakang genetik yang sama (Tabel 4 dan Tabel 5). Jumlah MAF Legian relatif lebih tinggi daripada ketiga desa
lainnya. Setelah dibandingkan dengan
frekuensi gen lain dari penelitian sebelumnya, dapat disimpulkan tingginya MAF di populasi Legian disebabkan oleh pemilihan sampel dan jumlah sampel yang sedikit.
Berdasarkan letak geografis, MAF gen KCNJ11 (p=0,032) dan ABCC8 (p=0,148) di daerah pesisir cenderung lebih tinggi daripada dataran tinggi. Hal tersebut berlaku pada gen KCNJ11 (34% vs 21%; p=0.032) dan ABCC8 (32% vs 24%; p=0,148). Tingginya MAF di daerah pesisir mungkin disebabkan paparan pengaruh genetik dari luar di daerah pesisir lebih besar dibandingkan dataran tinggi.
Gambar 6 Representasi deteksi polimorfisme
SNP E23K gen KCNJ11 dan SNP
A1369S gen ABCC8 yang
dilakukan dengan teknik
PCR-RFLP (Kiri:SNP E23K; Kanan
:SNP A1369S)
Desa N
Frekuensi Genotipe (%) Frekuensi Alel (%) Hardy-Weinberg Equilibrium MAF (%)
EE EK KK E K
Pesisir 92
Nusa Ceningan 46 52 39 9 72 28 0,730
34
Legian 46 44 35 22 61 39 0,069
Dataran Tinggi 92
Penglipuran 46 70 22 9 80 20 0,047
21
Pedawa 46 63 31 6 78 22 0,413
Tabel 5 Nilai Frekuensi Alel Minor (MAF) gen ABCC8 pada masing-masing populasi
Angka bercetak tebal menunjukkan nilai yang signifikan p<0,05 (Tidak memenuhi HWE) Kesetimbangan genotipe pada populasi
perlu dibandingkan dengan HWE untuk
memastikan bahwa frekuensi genotipe
tersebut stabil pada suatu populasi (Kang dan Shin 2004). Prinsip HWE menjelaskan bahwa jika suatu populasi dalam keadaan setimbang
maka terdapat lima kondisi yang
dipertahankan pada populasi tersebut, yaitu tidak terjadi mutasi dan migrasi pada populasi tersebut, ukuran populasi besar, terjadi perkawinan secara acak dan populasi tidak dalam tekanan seleksi alam (Kang dan Shin 2004). Pada populasi total, distribusi frekuensi
genotipe gen KCNJ11 tidak sesuai dengan
Hukum Kesetimbangan Hardy-Weinberg
(HWE) dengan nilai (p=0,008). Distribusi
frekuensi genotipe gen KCNJ11 populasi
Penglipuran juga tidak mengikuti HWE (0,047). Sebaliknya, frekuensi genotipe gen KCNJ11 di tiga desa lainnya menunjukkan kesesuaian dengan HWE (Tabel 4). Analisis distribusi frekuensi genotipe gen ABCC8 pada populasi total (p=0,007) dan Penglipuran
(p=0,012) tidak sesuai dengan HWE. Namun,
frekuensi genotipe gen ABCC8 ketiga populasi lainnya sesuai dengan HWE (Tabel 5). Tidak setimbangnya frekuensi genotipe kedua gen dengan HWE dapat disebabkan oleh pemilihan sampel dan jumlah sampel yang relatif sedikit sehingga tidak dapat mewakili seluruh populasi.
Asosiasi SNP E23K Gen KCNJ11 dan SNP A1369S Gen ABCC8 dengan Kriteria
DMT2
Analisis asosiasi SNP E23K dengan kriteria DMT2 keempat populasi di Bali tidak menunjukkan perbedaan yang signifikan, baik yang dianalisis pada populasi total (p=0,158), pada jenis kelamin laki-laki saja (p=0,247), maupun pada jenis kelamin perempuan saja
(p=0,463). Analisis SNP E23K pada masing-
masing desa juga tidak menunjukkan
perbedaan yang signifikan, baik pada jenis kelamin laki-laki maupun perempuan (Tabel 6). Pada populasi Nusa Ceningan dengan jenis kelamin perempuan, semua sampel termasuk
dalam kriteria FPG normal, sehingga
asosiasinya dengan kriteria DMT2 tidak dapat dianalisis.
Analisis asosiasi SNP A1369S dengan kriteria DMT2 pada populasi total tidak menunjukkan perbedaan yang signifikan (p=0,542), baik yang dianalisis pada populasi
laki-laki saja (p=0,471), maupun pada
populasi perempuan saja (p=0,510). Analisis asosiasi SNP A1369S dengan kriteria DMT2 pada masing-masing desa menunjukkan adanya perbedaan yang signifikan pada populasi laki-laki di Desa Pedawa (p=0,014).
Pada populasi tersebut genotipe AA
berasosiasi dengan kriteria hiperglikemia sedang dengan frekuensi 100%. Penduduk berjenis kelamin laki-laki di Desa Pedawa yang memiliki genotipe AA SNP A1369S perlu berhati-hati karena memiliki resiko DMT2 lebih tinggi daripada populasi lainnya. Hal ini, masih perlu dibuktikan melalui analisis terhadap sampel yang lebih besar. Analisis asosiasi SNP A1369S dengan kriteria DMT2 pada desa lainnya tidak menunjukkan perbedaan yang signifikan, baik pada laki-laki maupun perempuan (Tabel 6).
Penelitian lain menyatakan genotipe mutan
pada kedua gen (genotipe KK pada KCNJ11
dan genotipe AA pada gen ABCC8) berkaitan dengan DMT2 (Lefer et al. 2009; Florez JC et al. 2007; Giurgea et al 2006; Bryan J 2004; Riedel et al. 2004; Schwantecher et al. 2002). Sebaliknya, pada penelitian ini tidak terlihat asosiasi yang signifikan antara haplotipe mutan kedua gen dengan DMT2 di populasi total. Hal ini dapat disebabkan karena jumlah sampel yang sedikit.
Desa N
Frekuensi Genotipe (%) Frekuensi Alel (%) Hardy Weinberg Equilibrium MAF (%)
SS SA AA S A
Pesisir 92
Nusa Ceningan 46 54 37 9 73 27 0,726 32
Legian 46 45 35 20 63 37 0,111
Dataran Tinggi 92
Penglipuran 46 70 20 11 79 21 0,012
24
Pedawa 46 54 37 9 73 27 0,710
Tabel 6 Asosiasi SNP E23K dan SNP A1369S dengan DMT2di tiap populasi
Angka bercetak tebal menunjukkan nilai yang signifikan p<0,05 ; Semua nilai p dianalisis dengan menggunakan Chi –square test.
Variabel Kriteria (%) EE EK KK p SS SA AA P
Total (n=184)
Hiperglikemia sedang 14 3 11
0,158 13 5 13 0,542
DMT2 6 2 5 4 3 5
Laki-laki (n=27)
Hiperglikemia sedang 14 3 14
0,247 12 5 22 0,471
DMT2 6 3 14 5 5 11
Perempuan (n=19)
Hiperglikemia sedang 15 5 7
0,463 14 5 7 0,510
DMT2 5 0 0 5 0 0
Nusa Ceningan (n=46)
Hiperglikemia sedang 4 0 0
0,266 4 0 0 0,179
DMT2 4 0 0 4 0 0
Laki-laki (n=27)
Hiperglikemia sedang 7 0 0
0,735 8 0 0 0,676
DMT2 7 0 0 8 0 0
Perempuan (n=19)
Hiperglikemia sedang 0 0 0
- 0 0 0 -
DMT2 0 0 0 0 0 0
Legian (n=46)
Hiperglikemia sedang 15 0 0
0,345 14 0 0 0,391
DMT2 10 6 10 10 6 11
Laki-laki (n=27)
Hiperglikemia sedang 17 0 0
0,268 14 0 0 0,210
DMT2 0 9 25 0 10 33
Perempuan (n=19)
Hiperglikemia sedang 13 0 0
0,298 14 0 0 0,191
DMT2 25 0 0 29 0 0
Penglipuran (n=46)
Hiperglikemia sedang 19 0 0
0,461 16 11 0 0,830
DMT2 3 0 0 3 0 0
Laki-laki (n=27)
Hiperglikemia sedang 20 0 0
0,342 15 20 0 0,935
DMT2 5 0 0 5 0 0
Perempuan (n=19)
Hiperglikemia sedang 17 0 0
0,521 17 0 0 0,521
DMT2 0 0 0 0 0 0
Pedawa (n=46)
Hiperglikemia sedang 17 14 67
0,219 16 12 75 0,067
DMT2 7 0 0 4 6 0
Laki-laki (n=27)
Hiperglikemia sedang 11 13 100
0,138 8 8 100 0,014
DMT2 11 0 0 8 8 0
Perempuan (n=19)
Hiperglikemia sedang 27 17 50
0,647 23 25 50 0,722
Tabel 7 Asosiasi haplotipe gen KCNJ11 dan ABCC8 dengan kriteria DMT2 .
Haplotipe Gen KCNJ11 dan ABCC8
Gen KCNJ11 dan gen ABCC8 dilaporkan terkait satu sama lain, baik letaknya pada
kromosom, maupun fungsinya secara
fisiologis (Florez et al. 2007; Sakamoto et al 2007; Inagaki et al 1995). Kedua gen terletak sangat berdekatan, hanya dipisahkan 4,5 kbp dengan fungsi fisiologis yang komplementer,
yaitu membentuk subunit protein KATP
channel. Penelitian sebelumnya menyatakan
bahwa kedua gen memiliki Linkage
Disequilibrium (LD)>0,90 (Palmer et al. 2008; Florez et al. 2007; Florez et al. 2004; Aguilar-Bryan et al. 1998; Inagaki et al. 1995). Kedua gen memiliki kecenderungan yang kuat untuk diwariskan bersama-sama sehingga perlu dilakukan analisis haplotipe. Studi haplotipedilakukan ketika studi asosiasi genetik terhadap suatu marka tidak berhasil ditemukan. Kombinasi alel-alel pada dua SNPs atau lebih dapat memberikan
informasi tambahan mengenai hubungan SNPs dengan penyakit (Foulkes 2009).SNP
E23K pada gen KCNJ11 dan SNP A1369S
pada gen ABCC8 membentuk empat
kombinasi haplotipe, yaitu ES, EA, KS dan KA.
Asosiasi Haplotipe Gen KCNJ11 dan
ABCC8 dengan Kriteria DMT2
Analisis asosiasi haplotipe gen KCNJ11
dan ABCC8 dengan DMT2 dilakukan
menggunakan Expectation Maximization
Algorithms dengan model aditif. Hasil yang diperoleh ditampilkan pada Tabel 7. Hasil analisis asosiasi haplotipe dengan kriteria DMT2 menunjukkan hanya terdapat satu haplotipe yang signifikan, yaitu haplotipe KS
pada populasi Nusa Ceningan (p=0,039)
meskipun frekuensinya relatif lebih kecil jika dibandingkan dengan frekuensi haplotipe lain (6%). Pada populasi total dan ketiga populasi lainnya (Legian, Penglipuran dan Pedawa) tidak terlihat adanya asosiasi yang signifikan
Haplotipe Frekuensi Haplotipe(%) P
Total (184)
ES 66 0,138
EA 7 0,642
KS 6 0,662
KA 21 0,098
Nusa Ceningan (46)
ES 64 0,834
EA 8 0,739
KS 6 0,039
KA 22 0,182
Legian (46)
ES 55 0,338
EA 6 0,319
KS 8 0,498
KA 31 0,356
Penglipuran (46)
ES 74 0,187
EA 7 0,806
KS 6 0,489
KA 14 0,328
Pedawa (46)
ES 69 0,406
EA 9 0,539
KS 4 0,746
KA 18 0,513
p merupakan nilai asosiasihaplotipe kedua gen dengan kriteria DMT2 berdasarkan konsentrasi FPG; Angka bercetak tebal menunjukkan nilai yang signifikan p<0,05; Semua nilai p dihitung
antara haplotipe tertentu dengan kriteria DMT2. Penelitian lain yang dilakukan terhadap 3400 sampel Kaukasia menjelaskan bahwa haplotipe KA yang terbentuk dari SNP E23K dan A1369S berasosiasi dengan DMT2 (p=0,003)(Florez et al. 2004). Perbedaan ini disebabkan karena jumlah sampel yang kecil dan perbedaan populasi yang dianalisis.
SIMPULAN DAN SARAN
Simpulan
Nilai prevalensi DMT2 pada populasi Bali yang diamati tidak jauh berbeda dengan prevalensi nasional, yaitu 4,3% untuk DMT2 dan 10,9% untuk hiperglikemia sedang. Prevalensi DMT2 di populasi Nusa Ceningan (2,2%), Legian (8,6%) Penglipuran (2,2%) dan Pedawa (4,3%), sedangkan prevalensi hiperglikemia sedang di populasi Nusa Ceningan (2,2%), Legian (8,7%), Penglipuran (13,0%) dan Pedawa (19,6%). Berdasarkan tingginya prevalensi hiperglikemia sedang, prevalensi DMT2 pada keempat populasi diprediksi akan meningkat sebanyak 2 kali lipat dalam kurun waktu 10-15 tahun mendatang. Pada populasi total, hiperglikemia sedang berasosiasi dengan rentang usia 51-65 tahun. Namun, pada penelitian ini, tidak ditemukan asosiasi antara jenis kelamin
dengan kriteria DMT2. MAF gen KCNJ11
dan ABCC8 di populasi daerah pesisir lebih tinggi daripada populasi di dataran tinggi. Daerah pesisir juga memiliki keragaman rata-rata FPG lebih tinggi daripada dataran tinggi. Pada penelitian ini, tidak ditemukan asosiasi
SNP E23K pada gen KCNJ11 dan SNP
A1369S pada gen ABCC8 dengan DMT2
yang dianalisis di total populasi. Namun, ditemukan asosiasi genotipe AA pada SNP
A1369S gen ABCC8 dengan hiperglikemia
sedang di populasi laki-laki Desa Pedawa. Secara umum, analisis haplotipe yang dibentuk oleh kedua polimorfisme juga tidak
menunjukkan asosiasi terhadap kriteria
DMT2.
Saran
Kecilnya jumlah sampel merupakan
kelemahan dari penelitian ini. Oleh karena itu,
analisis cohort dan penggunaan jumlah
sampel yang lebih besar diperlukan agar dapat memberikan informasi yang lebih akurat dan representatif.
DAFTAR PUSTAKA
Aguilar Bryan L, Bryan J. 1999. Molecular biology of adenosine
triphophat-sensitive potassium channels.
Endocrine rev 20(2):101-135.
Aguilar-Bryan L, Clement JP, Gonzalez G, Kunjilwar K, Babenko A, Bryan J.
1998. Toward understanding the
assembly and structure of KATP channel. Physiol Rev 78:227-245.
Ashcroft FM. 1998. Adenosine
5-triphosphate-sensitive potassium
channels. Annual Review of
Neuroscience. 11:97–118.
Biggs ML, Mukamal KJ, Luchsinger JA, Ix
JH. 2010. Association between
adiposity in midlife and older age and risk of diabetes in older adults. JAMA Vol 303:No.24.
Bryan J. 2004. Toward Linking Structure
With Function in ATP-Sensitive K_ Channels. Diabetes 3: S104–S112 Campbell JD, Sansom MSP, Ascroft FM.
2003. Potassium channel regulation, structural insights into the function of the nucleotide-binding domains of the
human sulphonylurea receptor. EMBO
reports 4, 1038–1042.
[Depkes] Departemen Kesehatan RI. 2010.
Tahun 2030 prevalensi diabetes
mellitus di Indonesia mencapi 21,3 juta
orang. http://www.depkes.go.id.
[Februari 2011]
Feng Y, Mao G, Ren X, Xing H, Tang G, Li
Q. 2008. Ser1369Ala variant in
sulfonylurea receptor gene ABCC8 is associated with antidiabetic efficacy of gliclazide in Chinese type 2 diabetic patients. Diabetes Care 31:1939–1944. Fischer A et al. 2008. KCNJ11 E23K affects
diabetes risk and is associated with the disposition index. Diabetes Care 31: Number 1.
Florez JC, Burtt N, Bakker PIW, Almgren P, Toumi T, Johan T. 2004. Haplotype
structure and genotype-phenotype
correlations of the sulfonylurea
receptor and the islet ATP-sensitive
potassium channel gene region.
Florez JC, Jablonski KA, Khan SE, Frank PW, Dabelea D, Hamman RF,
Knowler WC . 2007. Type 2 Diabetes–
Associated Missense Polymorphisms KCNJ11 E23K and ABCC8 A1369S Influence Progression to Diabetes and Response to Interventions in the Diabetes Prevention Program. Diabetes 56(2): 531-536.
Foulkes A. 2009. Applied statistical genetics with R: for population-based study. Springer, New York: xxiii+252 hlm. Giurgea I, Bellanie-Chantellot C, Ribeiro M,
Hubert L, Sempoux C. 2006.
Molecular mechanisms of neonatal
hyperinsulinism. Horm Res 66:289–
296.
Gloyn AL,Phil D, Pearson ER, Antcliff JF, Proks P.2004. Activating mutations in the gene encoding the ATP-sensitive potassium-channel subunit Kir6.2 and permanent neonatal diabetes. N Engl J
Med 350: 1838-1849.
Gloyn AL, McCarthy. 2001. The genetics of type 2 diabetes. Best Pract Res Clin Endocrinol Metab 15:293-308.
Gong L. 2001. Anti-diabetic drug pathway (potassium channel inhibitor PD) http://www.pharmgkb.org/do/serve?obj Id=PA153627758&objCls=Pathway [18 Februari 2011].
Hani EH, Boutin P, Durand E, Inoue H, Permutt MA, Velho G, Froguel P . 1998. Missense mutations in the pancreatic islet beta cell inwardly
rectifying K+ channel gene
(KIR6.2/BIR):a meta-analysis suggests a role in the polygenic basis of type II
diabetes mellitus in Caucasians.
Diabetologia 41:1511-1515.
Hansen SK, Nielsen EMD, Jakob EK, Andersen G, Glumer C. 2005. Analysis of sparate and combined effects of common variation in KCNJ11 and PPARG on risk of type 2 diabetes. J Clin Endocrinol Metab 90(6) 3629-37. Hansen T, Echwald SM, Hansen L, Moller
AM, Almind K, Clausen JO. 1998.
Decreased tolbutamide-stimulated
insulin secretion in healthy subjects with sequence variants in the high-affinity sulfonylurea receptor gene. Diabetes 47(4):598-605.
Hansen L, Hansen T, Clausen JO, Echwald SM, Urhammer SA, et al. 1997. The Val985Met insulin-receptor variant in the Danish Caucasian population: Lack
of associations with
noninsulin-dependent diabetes mellitus or insulin resistance. Am J Hum Genet 60: 1532– 1535.
Human Genome Project Information. 2011. SNP Fact Sheet. http://www.ornl.gov/ scBi/techresources/Human_Genome/fa q/snps.shtml [20 April 2011]
[IDF] International Diabetes Federation. 2011.
About Diabetes.
http://www.idf.org/about-diabetes. [April 2011]
[IDF] International Diabetes Federation. 2010.
Diabetes Fact Sheet.
http://www.idf.org/about-diabetes. [April 2011]
Inagaki N, Glonoi P, Clement JP, Namba N, Inazawa J, Gonzalez G, Aguilar Bryan L. 1995. Reconstitution of IKATP: an inward rectifier subunit plus the
sulfonylurea receptor. Science
270:1166 –1170.
Jablonski KA, McAteer JB, de Bakker PI, Franks PW, Polin TI, Hanson RL.
2010. Common variants in 40 genes
asssesed fo diabetes incidence and response to metformin and lifestyle interventions in the diabets prevention program. Diabetes 59(10):2672-81. Kang dan Shin. 2004. The size of chi square
test for Hardy-Weinberg Law. Human
Heredity 58:10-11.
Ke X, Taylor MS, Cardon MR . 2008.
Singleton SNPs in the human genome and implications for genome-wide association studies. European Journal of Human genetics 16:506-515.
Kristiansen SB, Lofgren B, Nielsen JM, Stottrup NB, Buhl ES, Nielsen-Kudsk
JE. 2010. Comparison of two
sulfonylureas with