m w a d c r l w - r l C * L r l u
ik,
aewa i s a imci-
ataslor
m a
(I*.j o r l ) r c y S ~ E , ~ i i w i I r ) c l i t
EKSPRESI FENOTIP DAN AKTlVlTAS
BlOLOGl STREPTOKOKUS GRUP
C
ISOLAT
ASAL BABI
DAN
KERA
Oleh
:
IWAN HARJONO UTAMA
PROGRAM PASCASARJANA
INSTITUT
PERTANlAN BOGOR
SUMMARY
IWAN HARJONO UTAMA. Phenotypic Expression and Biological
Activities of Group C Streptococci Isolated from Pigs and Monkeys. (under
supervision of
Hj.
AISJAH GlRlNDRA as chairman, I WAYAN TEGUH WIBAWAN. FACHRIYAN HASMl PASARIBU, H. ENDHIE D. SETIAWAN andH GATUT ASHADI as members).
Thirty four isolates of streptococci from streptococcosis outbreaks in
pigs and monkeys had been -identified and characterized. All isolates
belonged to Lancefield's group C streptococci, and based on biochemical
reactions identified as Streptococcus equi subsp. zooepidemicus.
Twenty eight isolates (82,35%) produced mucoid colonies on blood agar,
turbidity in liquid media, diffuse colonies on soft agar and had hydrophylic
properties. In contrast, six isolates
(17,65%)
formed rough colonies on blood agar, sediment in liquid media with clear supernatant, compact colonies onsoft agar and had hydrophobic character. These phenotypic properties
seemed to be related to the encapsulation of these bacteria. Bacteria with
mucoid colonies expressed capsular structures on their surface. The
capsular structure were confirmed electronmicroscopicaIly. No comparable
result was shown by bacteria with rough colonies.
The acriflavin and decapsulation tests indicated that the main
component of capsule is hyaluronic acid. The hyaluronic acid capsule
phagocytic activity of polymorphonuclear (PMN) cells and for the adhesion of
bacteria on host's epithelial cells.
IWAN HARJONO UTAMA. Ekspresi Fenotip dan Aktivitas Biologi
Streptokokus Grup C Isolat Asal Babi dan Kera (di bawah bimbingan Hj.
AISJAH GlRlNDRA sebagai ketua, I WAYAN TEGUH WIBAWAN,
FACHRIYAN HASMl PASARIBU. H. ENDHIE D. SETIAWAN dan H. GATUT
ASHADI (Alm) sebagai anggota).
Tiga puluh empat isolat Streptokokus yang berasal dari wabah
streptokokosis pada babi dan kera telah diidentifikasi dan dikarakterisasi.
Semua isolat termasuk Streptokokus grup C menurut Lancefield dan
Streptococcus equi subsp. zooepidemicus menurut reaksi
biokimiawinya. Dua puluh delapan isolat (82,35%) tumbuh sebagai koloni
mukoid pada agar darah, menimbulkan kekeruhan dalam media cair, koloni
difus pada' media agar lunak (soft agar) dan memiliki sifat hidrofil.
Sebaliknya, enam isolat (17,65%) tumbuh sebagai koloni kasar pada media
agar darah, membentuk endapan dalam media cair dengan supernatan
jernih, koloni kompak di agar lunak dan rnemiliki sifat hidrofob. Slfat-sifat
fenotip tampaknya berhubungan dengan keberadaan kapsul. Bakteri dengan
koloni mukoid memperlihatkan struktur kapsul di permukaan selnya, ha1 ini
ditunjang dengan pengamatan elektron mlkroskop. Bakteri dengan
tampaknya berperan penting dalarn ketahanan bakteri terhadap aktivitas
fagositosis sel pofimorfonuklear (PMN) dan untuk perlekatan bakteri pada sel
inang.
EKSPRESI FENOTIP DAN AKTlVlTAS BlOLOGl
STREPTOKOKUS GRUP C ISOLAT ASAL BABl
DAN KERA
oleh
:
IWAN HARJONO UTAMA
Disertasi sebagai salah satu syarat untuk memperoleh gelar
Doktor
pada
Program Pascasarjana
lnstitut Pertanian Bogor
PROGRAM PASCASARJANA
INSTITUT PERTANIAN
'BOGOR
Judul disertasi : EKSPRESI FENOTIP DAN AKTlVlTAS
BlOLOGl STREPTOKOKUS GRUP-C ISOLAT ASAL BABl DAN KERA
Nama mahasiswa : IWAN HARJONO UTAMA Nomor pokok : 93550
Program studi : Sains veteriner
Menyetujui
L
Prof. Dr. Hi. Aisiah Girindra Ketua
Dr. Drh. I Wavan T. Wibawan. MS Dr. Drh. Fachrivan
&
Hasmi PasaribuAnggota Anggota
--
ttd.Dr. Drh. H. Endhie D. Setiawan. MS. Prof. Dr. H. Gatut Ashadi (Alrn)
Anggota Anggota
Ketua Program Studi Ssins Veteriner
~roVbr.~-~asduki Partadiredia.
M*!
Tanggal. lulus ujian :
Penulis lahir di Bogor tanggal 6 April 1961, anak tunggal dari Eddy G.
Utama (alrn) dan Mien Ashadi (alrn). Tahun 1980 ia lulus dari SMA
Kesatuan Bogor, masuk lnstitut Pertanian Bogor tahun 1980 (Tingkat
Persiapan Bersama) dan lulus Dokter Hewan tahun 1985. Pendidikan
pascasarjana di tnstitut Pertanian Bogor jurusan sains veteriner dimulai
tahun 1986 dan lulus Magister Sains tahun 1989. Mendapat kesempatan
melanjutkan program doktor di lnstitusi yang sama pada tahun 1993 dan
lulus pada tahun 1998.
Pengalaman bekerja dimulai'dari asisten luar biasa pada tahun 1982
dalam mata ajaran Biokimia ~ a s a r ' l .dan 2, Anatomi ~ e t e r i n e r I dan 2 serta
Mikrobiologi Umurn di Fakultas Kedokteran Hewan IPB. Tahun 1989 hingga
sekarang penulis bekerja sebagai staf pengajar Biokimia Veteriner di
Program Studi Kedokteran Hewan (sekarang Fakultas Kedokteran Hewan)
Universitas Udayana-Bali. Penulis juga ikut rnembantu mengajar Patologi
Klinik Veteriner di institusi yang sama dan Kimia Klinik di Jurusan Kimia
Program Studi Matematika dan llmu Pengetahuan Alam (sekarang telah
menjadi Fakultas Matematika dan llrnu Pengetahuan Alam) Universitas
UCAPAN TERIMA KASlH
Penulis sangat berterima kasih kepada Prof. Dr. Hj. Aisjah Girindra,
pembimbing utamanya atas saran d a n bimbingannya selarna penelitian.
Ucapan terirna kasih juga ditujukan kepada anggota pembimbing lainnya, Dr.
Drh. I Wayan Teguh Wibawan, MS, Dr. Drh. Fachriyan Hasmi Pasaribu, Dr.
Drh. H. Endhie D. Setiawan, MS dan Prof. Dr. H. Gatut Ashadi atas
bimbingannya.
Ucapan terima kasih juga ditujukan kepada Tim Manajemen Program
Doktor (TMPD) Dikti dan Tim Proyek Hibah Bersaing VI I 1 I 1997-1999
Direktorat Pembinaan Penelitian dan Pengabdian pada Masyarakat
(Ditbinlitabrnas) Departemen Pendidikan dan Kebudayaan Republik
Indonesia atas dana penelitian yang diberikan. Kepada Kepala
Laboratorium Biokimia Fakultas Matematika dan llmu Pengetahuan Alam
lnstitut Pertanian Bogor, Kepala Laboratorium Patologi Veteriner dan Kepala
laboratorium Mikrobiologi Fakultas Kedokteran Hewan lnstitut Pertanian
Bogor juga di&apkan terima kasih atas ijin dan fasilitas yang diberikan
selama penelltian ini berlangsung. Tidak lupa juga diucapkan terima kasih
kepada teknisi
laboratorium-laboratorium
tersebut atas bantuan teknis yangdiberikan selarna penelitian. Ucapan terima kasih juga ditujukan kepada
beberapa perpustakaan seperti perpustakaan Balai Penelitian Veteriner-
perpustakaan bagian Mikrobiologi Fakultas Kedokteran Universitas
Indonesia, perpustakaan Fakultas Kedokteran Universitas Indonesia,
perpustakaan pusat lnstitut Pertanian Bogor-Darmaga dan pusat informasi
lainnya yang telah banyak membantu penulis.
~ e n e l i t i a n ini tidak dapat diselesaikan tanpa bantuan dari berbagai
pihak seperti Balai Penyidikan Penyakit Hewan (BPPH) Wilayah VI
Denpasar-Bali, Dinas Peternakan Propinsi Daerah Tingkat I Bali. BPPH
Wilayah I l t Bandarlampung-Lampung, BPPH Wilayah VII Maros-Sulawesi
Selatan dan lnstansi lainnya. Untuk itu diucapkan terima kasih atas
kerjasama yang baik ini.
Kepada Bagian Mikrobiologi Justus Leibig Universitat-Giessen,
Germany diucapkan terima kasih atas bantuannya dalam preparasi sediaan
elektron mikroskop dan beberapa isotat rujukan termasuk isolat streptokokus
grup C yang diberikan.
Akhir kata, disertasi ini tidak mun$kin tersusun tanpa bantuan kedua
orang tua Eddy G. Utama (Alm). dan Mien Ashadi (Alm) yang telah
membesarkan penulis serta istri tercinta Drh. Ninuk Sri Rejeki yang telah
mendampingi penulis dalam suka dan duka. Untuk kolega dan karyawan
dari Fakultas Kedokteran Hewan Universitas Udayana juga diucapkan terima
kasih atas perhatiannya selama penulis mengikuti tugas belajar ini.
Bogor, Juli 1998
KATA PENGANTAR
Puji syukur penulis panjatkan pada Tuhan Yang Maha Esa atas berkat
dan rahmat yang diberikan hingga penelitian ini dapat diselesaikan.
Penelitian ini membahas streptokokus grup
C
yang merupakan bakterikomensal tapi berpeluang menjadi patogen baik pada hewan mau pun
manusia. Permasalahan ini sering terabaikan terutama dari kalangan
praktisi medis
.
dan veteriner. Kemunculan wabah streptokokosis yangmenyerang babi dan kera pada tahun 1994 di Bali yang kemudian menyebar
ke daerah lainnya merupakan salah satu bukti ha1 di atas. Selain merupakan
penyakit yang baru pertama kali dilaporkan di Indonesia, bakteri
penyebabnya juga baru dan telah dipelajari serta diteliti sebagian sifat-
sifatnya dalam disertasi ini.
Akhir kata, tak ada karya manusia yang sempurna meskipun usaha ke
arah itu selalu dilakukan, demikian juga halnya disertasi ini. Kritik dan saran
positif sangat diharapkan untuk perbaikan disertasi ini.
Bogor, Juli 1 998
DAFTAR IS1
Halaman
DAFTAR IS1 ... i
DAFTAR TABEL ... iv
DAFTAR GAMBAR ... vi
DAFTAR LAMPIRAN ... viii
I PENDAHULUAN ... 1
1 1 Latar Belakang ... 1 1 2 Tujuan Penelitian
. .
... 4...
.
1 3 Kegunaan Penelltlan 5
I1 TINJAUAN PUSTAKA ... 6
2.1 Sifat Streptokokus ... : ... 6 2.2 Streptococcus equi subsp
.
zooepidemicus pada Hewan danManusia ... 8 . 2.3 Komponen Kimiawi Permukaan Sel S . equi subsp
.
zooepidemicus ... 9 2.3.1 Kapsul sebagaj Komponen Permukaan Sel ... 9 2.3.2 Dinding Sel ... 14
2.3.2.1 Peptidoglikan, Asam Teikoat dan Lipoteikoat ... 14 2.3.2.2 Karbohidrat Dinding Sel ... 17 2.3.2.3 Antigen Protein ... 19 2.4 Antigen Ekstra Seluler ... 22 2.5 Sifat Permukaan Sel Bakteri ... 23 2.6 Perlekatan Bakteri pada Sel lnang ... 26 2.7 Fagositosis oleh Sel Polimorfonuklear (PMN) ... 28 2.7.1 Biologi PMN ... 28 2.7.2 Proses Fagositosis oleh PMN ... 29 2.8 Kapsul dan lmunopatogenesisnya ... 32
Ill 8AHAN DAN METODA : ... 37
3 1 Bahan ... 37
...
3.1 . 2 Hewan Percobaan 37
... .
3.1 3 Media Umum 37
3.1 . 4 Media untuk Uji Biokimia ... 38 3 1 . 5 Bahan Kimia Padat ... 38 3.1 . 6 Kit untuk Pengujian Serologis Streptokokus ... 39
...
3.7 . 7 Kertas Cakram Antibiotika 39
3.1.8 Bahan Kimia Cair ... 39 3.1 . 9 Bahan-bahan Lain ... 40 3.1.10 Alat-alat ... 40
...
3.1 Metoda Penelitian 41
3.2.1 Isolasi, Pengamatan Makroskopis dan Mikroskopis
Bakteri ...
1
... 41...
3.2.2 Penentuan Grup secara Serologis 42
3.2.2.1 Pembuatan Antigen Utuh untuk Uji
Koaglutinasi ... 42 3.2.2.2 Pembuatan Ekstrak Antigen untuk Uji
.
.
lmunod~fus~ Ganda ... 43 3.2.3 Pengujian Biokimia ... 44 3.2.4 Uji Kepekaan terhadap Antibiotika In
vitro
... 45 3.2.5 Karakterisasi lanjut : Pengamatan Ekspresi Fenotiplsolat ...
.
.
... 46...
3.2.6 Pengujian Kimiawi dan Konfirmasi Struktural Kapsui 47 3.2.6.1 Uji Akriflavin ...
:.
... 47 3.2.6.2 Uji Dekapsulasi ... 48 3.2.6.3 Salt Aggregation Test ( S A T ) ... 48 3.2.6.4 Pengamatan Elektron Mikroskopi ... 49 3.2.7 Elektroforesis SDS-PAGE ... 513.2.8 Uji Aktivitas Biologi Isolat ... 53 3.2.8.1 Uji Fagositosis In
vitro
... 53 3.2.8.2 Uji Fagositosis Invivo
... 56 3.2.9 Uji Perlekatan pada Sel lnang ... 573.2.9.1 Pengaruh Perlakuan terhadap Sel Bakteri
Berkapsul ... 59 3.2.9.2 Pengaruh Perlakuan terhadap Sel Epitel ... 60 3.2.10 Analisis Data ... 61 3.2.1 1 Waktu dan Tempat Penelitian ... 62
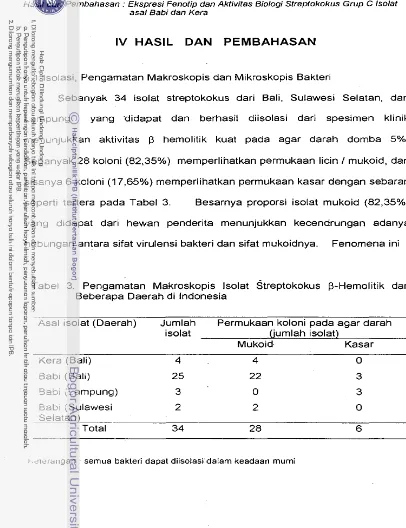
4.1 Isolasi. Pengamatan Makroskopis dan Mikroskopis
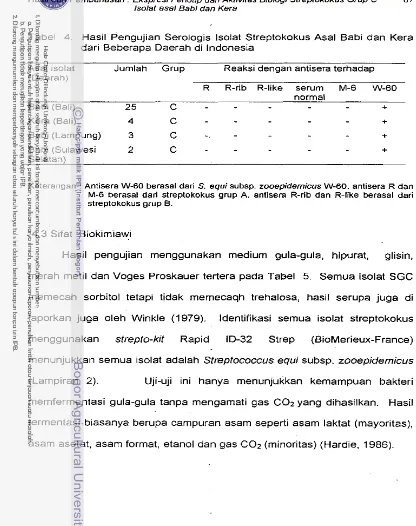
Bakteri ... 63 4.2 Penentuan Grup Secara Serologis . . . ... 65 4.3 Sifat Biok~rn~aw~ ... 67 4.4 Kepekaan lsolat Terhadap Antibiotika ... 68 4.5 Karakterisasi lanjut : Pengamatan Ekspresi Fenotip
lsolat ... 71 4.6 Pengujian Kapsul ... 76 4.6.1 Uji Akriflavin ... 76 4.6.2 Uji Dekepsulasi ... 77 4.6.3 Salt Aggregation Test (SAT) ... 78 4.6.4 Elektron Mikroskopi ... 81 4.7 Elektroforesis SDS-PAGE Ekstrak Antigen lsolat ... 81 4.8 Aktivitas Biologi lsolat ... 83 4.8.1 Fagositosis Bakteri In vitro ... 83 4.8.2 Fagositosis
In
vivo ... 89...
4.8.3 Perlekatan Bakteri pada Sel lnang 94
4.8.3.1 Pengaruh Perlakuan terhadap Sel Bakteri
Berkapsul ... ... 97 4.8.3.2 Pengaruh Perlakuan terhadap Sel
...
Epitel 99
...
V KESIMPULAN DAN SARAN 103
DAFTAR TABEL
Judul Halaman
Kemampuan Memfermentasi Trehalosa dan Sorbitol dari
ke 3 Spesies Streptokokus Grup C ..: ... 7
Sifat Biokimiawi Streptokokus Grup C ... 8
Pengamatan Makroskopis lsolat Streptokokus a-Hemolitik
dari Beberapa Daerah di Indonesia ... 63
Hasil Pengujian Serologis lsolat Streptokokus Asal Babi dan
Kera dari Beberapa Daerah di Indonesia ... 67
Hasil Pengujian Beberapa Sifat Biokimiawi lsolat Streptokokus
Grup C dari Betserapa Daerah di Indonesia ... 68
Sebaran Kepekaan lsolat S. equi subsp. zooepidemicus dari Beberapa Daerah di lndonesia terhadap Bermacam-macam
Antibiotika ...
. . .
... 69Karakterisasi Lanjutan Ekspresi Fenotip lsolat S. equi subsp.
zooepidemicus darj Beberapa Daerah di Indonesia ... 72
Karakterisasi Lanjutan Sifat Permukaan Sel S. equi subsp.
zooepidemicus dari Beberapa Daerah di Indonesia ... 80
Aktivjtas dan Kapasitas Fagositosis Bakteri Berkapsul dan
Tidak Berkapsul oleh PMN Babi
In
vitro ... 85Kapasitas Fagositosis Bakteri berkapsul dan Tidak Berkapsul
oleh Kultur Makrofag Peritoneal Mencit In vitro ... 86
Aktivjtas dan Kapasitas Fagositosis Bakteri Berkapsul dan Tidak Berkapsul olehPMN dan Sel Makrofag Peritoneal
12 Aktivitas dan Kapasitas Fagositosis- Bakteri Berkapsul dan Tidak Berkapsul oleh PMN dan Sel Makrofag Darah Mencit
In vivo. ... 92
13 Perlekatan
S.
equi subsp. zooepidemicus pada Sel EpitelMukosa Buccalis Babi ... 95
14 Pengaruh Perlakuan S. equi subsp. zooepidemicus Berkapsul dengan Hialuronidase terhadap Perlekatan pada Sel Epitel
Buccalis Babi ... 98
15 Pengaruh Berbagai Perlakuan pada Sel Epitel Mukosa Buccalis
...
Babi terhadap Perlekatan S. equi subsp. zooepidemicus 100
16 Preinkubasi Sel Epitel Mukosa Buccalis Babi dengan Asam Hialuronat terhadap Perlekatan S. equi subsp.
DAFTAR GAMBAR
Judul Halaman
Struktur Monomer Asam Hialuronat Disakarida ... ... .. 11
D~agram Skematis Lintasan Biosintesis Asam Hialuronat
Disakarida
...
... ... ... ... ... .... ... ... ... . .. . .. 13Beberapa Struktur Umum Peptidogtikan dan Konfigurasi
.Susunan Kornponennya ... .. 15
Struktur Kimia Asam Teikoat tipe Gliserol dan tipe Ribitol ... 15
Hubungan antara Asam Teikoat (Rantai Sampingnya) dengan
Polimer Polisakarida dari Peptidoglikan ... ... ... ...
...
...
...
.. . .... . .
16Struktur Pembalut Sel Bakteri Kokus Gram Positif ... 18
Pengaturan Tulang Punggung Gliserofosfat dari Asam Lipoteikoat yang Mengandung Sekeiompok Asam Amino
Bermuatan Positif (biasanya Lisin) ... 20
Model Umurn Proses Antigen, Menunjukkan Lintasan
Konvensional MHC-I dan MHC-I1 ... 31
Reaksi Serologis Penentuan Grup Streptokokus asal Babi
Menggunakan Strepfo-Grouping Kit ... 66
Ekspresi Fenotip Isolat S. equi subsp. zooepidemicus pada
Media Cair 74
Kolonr S. equi subsp. zooepidemicus pada Soft Agar ... ... .. 7 5
Reaksi Akriflavin dengan Kapsul S. equi subsp.
zooepidemicus ...
...
.
... ... .. . .. . .. . ... .. ...
. ....
... . ..
.
. . . . ..
. . . 7614 Elektron Mikroskopik Sel S.
equi subsp. zooepidemicus
...
(33.200 x) 82
15 Pola Elektroforetik Ekstrak Antigen S.
equi subsp.
...
zooepiderni~us
8316 Grafik Kapasitas Fagositosis Bakteri Berkapsul dan Tidak
Berkapsul oleh PMN Babi In vitro ... 85
17 Grafik Kapasitas Fagositosis Bakteri Tidak Berkapsul oleh
Kultur Makrofag Peritoneal Mencit In
vitro ... 87
I 8 Fagositosis Bakteri Tidak Berkapsul oleh PMN (Pewarnaan
Giemsa 1 600 x) ...
:
... 8819 Grafik Aktivitas Fagositosis Bakteri Berkapsul dan Tidak Berkapsul oleh PMN dan Sel Makrofag Peritoneal Mencit
In
vivo
...
9020 Grafik Kapasitas Fagositosis Bakteri Berkapsul dan Tidak Berkaps~ii oieh PMN dan Sel Makrofag Peritoneal Mencit
In
vivo ...
9121 Grafik Kapasitas Fagositosis Bakteri Berka-psul dan Tidak Berkapsul oleh PMN dan Sel Makrofag Darah Mencit
In
vivo..
... 9322 Grafik Kemampuan Melekat Sel Bakteri Berkapsul dan
Tidak Berkapsul pada Set HeLa ... 96
23 Grafik Pengaruh Perlakuan Kimia pada Sel Bakteri Berkapsul
terhadap Kemampuan Melekatnya pada Sel HeLa
...
9724 Grafik Pengaruh Preinkubasi Sel HeLa dengan Larutan Asam Hialuronat 4 mg / ml terhadap Kemampuan Melekat
DAFTAR LAMP IRAN
No Judul Halaman
1 Nilai Rujukan Kepekaan Bakteri terhadap Antibiotika
( I n vitro) ... ... ... ... .... 125
2 Hasil Pengujian Sifat Biokimiawi Streptokokus Grup C
Pendahuluan : Ekspresi Fenotip dan Aktivitas Biologi Streptokokus Grup C lsolaf asal
Babi dan Kera
I
'PENDAHULUAN
1 1 Latar Belakang
DI negara berkembang mash ditemukan masyarakat yang gemar
mengkonsurnsr daging babi atau darah yang tidak dimasak atau d~masak
setengah matang Hal ini rnenjadi salah satu faktor pendukung penyebaran
penyakit, terutarna yang bersifat zoonosis. Banyak bakteri yang hidup
komensal dalarn tubuh hewan rnaupun manusia, yang aicibat stress 1
imunodepresi dapat mengakibatkan bakteri tersebut mampu menirnbulkan
penyakit.
Bakteri patogen memiliki banyak faktor yang berperan dalam
rnempertahankan diri terhadap sistem kekebalan inang. Secara garis besar
faktor tersebut dapat dibagi menjadi faktor ekstraseluler (metabolit) dan
faktor yang berasosiasi dengan sel / antigen permukaan (Mims, 1982).
Secara serologis, streptokokus dikelornpokkan oleh Lancefield
rnenjadi beberapa grup di antaranya grup A, B, C. D, G, dan L (Davies.
Dulbecco, Eissen, Ginsberg dan Wood. 1973). Hingga saat ini diketahui
S t r e p t o c o c c u s pyogenes (grup A) banyak dikaitkan dengan infeksi pada
manusia dan S. a g a l a c t i a e banyak dikaitkan dengan kasus mastitis pada
sapi perah dan penyakit neonatal pada manusia (Hardie, 1986).
Streptokokus lainnya bervariasi, baik dalarn serogrup maupun dalam inang
yang dihuninya, rnisalnya S. d y s g a l a c t i a e asal sapi (grup C) dan S. canis
Pendahuluan : Ekspresi Fenotip dan AMivitas Biologi Streptokokus Grup C lsolat asal 2
Babi dan Kera
Devriese, Hommez, Baltz dan Scheiffer (1986) juga menunjukkan
Streptococcus canis tidak hanya berasal dari anjing, tetapi juga dari hewan
la~nnya. Penggolongan streptokokus akhir-akh~r ini bukan saja berdasarkan
atas ekovar dan serologis, tetapi juga dilakukan dengan analisis hibridisasi
DNA-DNA (Devriese, 1991 ).
Streptokokus grup C (SGC) terdiri dari empat spesies, yaitu S.
equisirnills, S. dysgalactiae, S. equi, dan S. zooepidemicus (Davies et
at.,
1973). Berdasarkan analisis hibridisasi DNA-DNA, streptokokus grup C
dikelompokkan menjadi tiga spesies yaitu S. equisimilis, S. dysgalactiae. S.
equi subsp. equi dan S. equi subsp. zooepidemicus (Farrow dan Collins,
1984). Bakteri ini umumnya merupakan komensal pada babi dan manusia
(S. equisimilis), kuda (S. equi subsp. egui), babi, kuda dan hewan lainnya (S. equi subsp. zooepidemicus) (Blood dan Radostits, 1989; Farrow dan Collins.
1984; Jorm, Love dan Bailey. 1994). Meskipun demikian, kasus infeksi
SGC pada babi tidak banyak dilaporkan (Evans, 1944; Roberts, Ramsey,
Switzer dan Layton, 1968; Kahn dan Ross, 7972).
Pada bulan April 1994 dilaporkan terjadi wabah penyakit yang
menyerang babi pada peternakan rakyat di Bali yang dikaitkan dengan
infeksi streptokokus. Pada waktu yang bersamaan terjadi pula wabah pada
kera di beberapa Tarnan Wisata Alam di Bali oleh jenis bakteri yang sarna.
Berjangkitnya wabah streptokokosis pada kera akibat minum air sungai
Pendal~uluan : Ekspresi Fenotip dan AMivitas Biologi Streptokokus Grup C lsolat asal Babi dan Kera
1994). Kasus ini merupakan kejadian pertarna di Indonesia, sebelumnya
tidak pernah dilaporkan kasus infeksi streptokokus yang dikaitkan dengan
kematian babi dan kera. Sebanyak 2200 ekor babi dan ratusan ekor kera di
Taman Wisata Alarn mati akibat penyakit ini (Rudyanto dan Dharrna, 1994;
Suharsono dan Dibia, 1994). Bakteri Streptococcus sp. yang rnenghasilkan
tipe hemolisis beta pada agar darah dapat diisolasi secara konsisten
(Dharma, 1994). Bakteri penyebab wabah ini kernudian dapat digolongkan
ke dalarn grup C (Dartini, Soeharsono, Ekaputra, Dibia, Dharrna dan
Supartika, 1994; Wibawan dan Pasaribu, 1994). Dalarn waktu singkat kasus
ini telah menyebar ke Surnatera Utara dengan kernatian ternak babi
mencapai 8422 ekor dan kerugian mencapai lebih dari Rp. 101.300.000,-
(seratus satu juta tiga ratus ribu rupiah) (Hasibuan, 1994). Wabah
streptokokosis juga terjadi di Lampung sebanyak 2 kasus (Anon. 1995),
Sulawesi Selatan serta Sulawesi Utara sebanyak 14 kasus (Anon. 1996 a,
b).
Kapsul rnerupakan struktur terluar dari sel bakteri dan urnurnnya
terdiri dari karbohidrat. Asarn hialuronat merupakan komponen utarna
pembentuk kapsul streptokokus grup C (Davies et at., 1973); komponen ini
dijumpai pula pada kapsul Pasteurella multocida (Esslinger. Seleirn dan
Blobel, 1989). Mengingat sebagian besar isolat SGC asal babi dan kera
rnemilik~ kapsul (Wibawan dan Pasaribu, 1994) rnaka penelitian ini dilakukan
Pendahuluatl : Ekspresi Fenotip dan Aktivitas Siologi Streptokokus Grup C Isolat asal 4
Babi dan Kera
1. Kasus streptokokosis pada babi dan kera rnerupakan kasus baru di
Indonesia
2 S~fat khas bakteri penyebab. terutama dika~tkan dengan keberadaan
kapsulnya
3 Sifat dan peran kapsul dalam proses infeksi, khususnya sebagai faktor
antrfagositik dan sebagai faktor adhesif bakteri pada perrnukaan sel inang.
? .2 Tujuan Penelitian
Penelitian ini bertujuan untuk :
1. Mengidentifikasi spesies isolat Streptococcus sp. yang didapat dari kasus
streptokokosis pada babi dan kera di Indonesia.
2. Melakukan karakterisasi fenotip bakteri yang dikaitkan dengan
keberadaan kapsul dan sifat perrnukaan sel.
3. Mengetahui peran kapsul dalam proses infeksi, khususnya sebagai faktor
antifagositik dan perannya dalam proses perlekatan bakteri pada sel
inang.
Hipotesis yang akan diuji :
Keberadaan kapsul pada perrnuka'an SGC bertanggung jawab terhadap
ekspresi fenotip, sifat antifagositik dan menjadi perantara proses perlekatan
Pendahuluan : Ekspresi Fenotip dan Aktivitas Biologi Streptokokus Grup C lsolat asal 5 Babi dan Kera
I . 3 Kegunaan Penelitian
Penelitian ini diharapkan mampu menggali informasi dasar khususnya
tentang sifat-sifat agen sehingga dapat digunakan sebagai landasan
pernilihan kandidat vaksin streptokokus. Vaksin dapat dibuat menggunakan
sel bakteri utuh atau komponen permukaan sel sebagai antigen. Komponen
asam hialuronat kapsul merupakan kandidat antigen terpilih untuk
pengembangan vaksin kapsul streptokokus yang dikonyugasikan dengan
protein seperti toksoid tetanus. Percobaan ini pernah dilakukan pada
polisakarida meningokokus grup A, B dan C dan berhasil dengan baik
(Jennings dan Lugowski, 1981 ). Pengembangan vaksin tersebut
dimungkinkan mengingat sifat bakteri ini kurang imunogenik (Hutomo,
1998). Pembuatan bahan diagnostik berupa antigen sel utuh dari S. equi
subsp. zooepidemicus untuk pemantauan kasus di lapangan secara
serologis merupakan alternatif lain dari kepentingan penelitian h i .
Penelaahan lebih lanjut mengenai kemampuan perlekatan bakteri pada sel
inang dan komponen penghambatnya diharapkan menjadi dasar untuk
pengembangan terapi anti perlekatan rnikroba pada sel inang mengingat
telah banyak mikroba resisten terhadap beberapa antibiotika. Terapi anti
perlekatan ini di masa depan diharapkan menjadi terapi alternatif yang
sangat potensial guna pencegahan infeksr oleh bakteri (Ofek, Kahane dan
Tinjatran Pustaka : Ekspresi Fenotip dan Akfivitas Biologi Strepfokokus Grrup C Isolat asal Babi dari Kera
II TINJAUAN PUSTAKA
2 1 S~fat Streptokokus
Streptokokus. merupakan bakteri berbentuk bulat dengan susunan
sepertl rantai dan tersebar luas di alam. Beberapa spesies di antaranya
merupakan flora normal manusia dan newan (Blood dan Radostits, 1989).
Beberapa spesles bers~fat patogen pada manusia misalnya S. pyogenes
(grup A) dikenal sebagai peyebab rheumatoid fever ( Davies et a/., 1973). S.
agalactiae (grup B) sebagai penyebab penyakit neonatal dan mastitis pada
sapi perah (Wibawan dan Laemmler, 1990), Streptococcus suis (grup D)
penyebab meningitis pada babi dan S. canis (grup G) yang patogen pada an- jing (Blood dan Radostits, 1989).
Streptokokus grup C menurut Lancefield merupakan komensal lain
pada manusia dan hewan , terdiri dari 3 spesies yaitu S. equisimilis; S. dys-
galactiae, dan S. equi yang terdirj dari dua subspesies, yaitu S. equi subsp.
equi dan S. equi subsp. zooepidemicus.(Winkle, 1979; Farrow dan Collins,
1984; Jorm er al., 1994). Streptococcus equisimilis merupakan patogen pada
manusiz (Esferatiou, 1989; Fox, Turner dan Fox,
t
993; Lebrun, Gilbert,Wallet. Manneville dan Pilot, 1986). S. dysgalactiae patogen pada sapi
menyebabkan rnasdtis (Baksi dan Singh, 1964; Dodd dan Norcross, 1967).
Streptococcus equi subsp. equi patogen pada kuda penyebab ingus ganas
(strangles) dan S. equi subsp. zooepidemicus penyebab ingus tenarig (stran-
Tinjauan Pustaka : Ekspresi Fenotip dan Aktivitas Biologi Streptokokus Grrup C isolat 7
asal Babi dan Kera
subsp. zooepidemicus juga patogen pada manusia (Low, Young dan Hard-
ing, 1980; Barnham, Thornton dan Lange, 1983; Esferat~ou, Coleman,
Hahn.Timmoney, Boutgrass dan Monget, 1994).
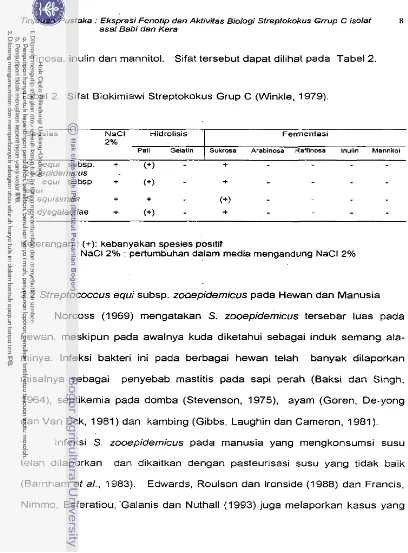
Secara biokimiawi, streptokokbs grup C dapat dibedakan berdasarkan
sifat-sifat sebagai berikut :
1. Kemampuan memfermentasi trehalosa dan sorbitol, ini terutarna untuk
membedakan S. equisimilis, S. equi subsp. zooepidemicus, dan S. equi
[image:173.515.38.458.12.567.2]subsp. equi (Winkle, 1979). Sifat tersebut dapat dilihat pada Tabel 1.
Tabel 1. Kemampuan Memfermentasi Trehalosa dan Sorbitol dari ke 3 Spe- sies Streptokokus Grup C (Winkle, 1979).
Spesies Sorbitol Trehalosa
S. equisimilis -
+
S. equi subsp. equi 4-
-
S. equi subsp. zooepidemicus
-
+
2. Berdasarkan uji aktivitas enzim a-L glutamat amino peptidase bersama
sama dengan fermentasi glikogen dan sorbitol (Esferatiou et a/., 1994).
Cara ini dapat juga di gunakan untuk membedakan S equi subsp. equi dan
S. equi subsp. zooepidemicus. a
3. Winkle (1979) mengklasifikasi ke tiga spesies dan dua subspesies ini den-
gan cara mengamati kemampuan tumbuh dalam medium mengandung
Tinjauan Pustaka : Ekspresi Fenotip dan Aktivitas Biologi Streptokokus Grrup C isokt asal Babi dan Kera
finosa. inulin dan mannitol. Sifat tersebut dapat dilihat pada Tabel 2.
Tabel 2. Sifat Biokimiawi Streptokokus Grup C (Winkle, 1979).
S. equi subsp. (+) +
zooepide~nicus
S. equi subsp + (+) +
equi
S. equisimilis + + (+)
Spesies NaCl H i d r o l i s i s
2%
Pati Gelatin
S. dysgalactiae + (+) +
Fennentasi
Sukrosa Arabinosa Raffinosa lnulin Mannilol
Keterangan : (+): kebanyakan spesies positif
NaCl2% : pertumbuhan dalam media rnengandung NaCl2%
2.2 Streptococcus equi subsp. zoaepidernicus pada Hewan dan Manusia
Norcoss (1969) mengatakan S. zooepidemicus tersebar luas pada
hewan, meskipun pada awalnya kuda diketahui sebagai induk semang ata-
minya. lnfeksi bakteri ini pada berbagai hewan telah banyak dilaporkan
misalnya sebagai penyebab mastitis pada sapi perah (Baksi dan Singh,
1964). septikemia pada domba (Stevenson, 1975). ayam (Goren, De-yong
dan Van Eck, 1981 ) dan kambing (Gibbs, Laughin dan Cameron. 1981 ).
infeksi S. zooepidemicus pada manusia yang mengkonsumsi susu
telah dilaporkan dan dikaitkan dengan pasteurisasi susu yang tidak baik
(Barnham et a/., 1983). Edwards, Roulson dan ironside (1 988) dan Francis.
[image:174.515.42.450.14.567.2]Ti)?jaua~? Pustaka : Ekspresi Fenotip dan Aktivitas Biologi Streptokokus Grrup C isolat 9
asal Sabi dan Kera
Nimmo, Esferatiou, 'Galanis dan Nuthall (1993) juga melaporkan kasus yang
sama akibat konsumsi susu mentah yang berasal dari sapi penderita mastitis
subklinis. Peran kuda dalam penularan penyakit ini ke manusia nampaknya
sangat penting (Barnham. Lyungren dan Mc Intyre, 1987). Barnham, Kerby,
Chandler dan Millar ( I 989) mengatakan infeksi S. equi subsp. zooepidemicus
pada manusia adalah akibat konsumsi susu yang tidak dipasteurisasi (zoo-
nosis) atau kontak dengan kuda. Esferatiou (1989) mengatakan sebelum
tahun 1980-an, infeksi bakteri ini pada manusia masih sangat jarang. lnsiden
penyakit yang dikaitkan dengan bakteri ini belakangan semakin meningkat.
Gejala klinis yang ditimbulkan pada manusia, adalah pharingitis, arthritis dan
septikemia.
2.3 Komponen Kimiawi Permukaan
el
S. equi subsp. zooepidemicus2.3.1 Kapsul sebagai Komponen Peimukaan Sel
Kapsul merupakan struktur terluar permukaan sel bakteri dengan kon-
sistensi seperti lendir (bersifat hidrofil dengan kandungan airnya lebih dari
95%). Komposisi kopsut sangat beragam, umumnya terdiri dari karbohidrat
yang bermuatan negatif (Bayer dan Thurow, 1977). berbagai makromolekul
dan beberapa ion anorganik yang memang dapat diserapnya (Geddie dan
Sutherland, 1993). Ketebalan kapsul umumnya sekitar 10 nm, tetapi pada
Klebsiella pneumoniae bisa mencapai 160 nm (Amako. Meno dan Takade,
1988). P a d a Eschertchia colt ( E . coli) K1, ketebalan kapsul dapat menjngkat
Tinjaua~l P~rstaka : Ekspresi Fenotip dan Aktivitas Biologi Streptokokus Grrup C isolat 10
asal Babi dan Kera
hadapnya Fenomena ini telah diketahui sejak lama pada Sfreptococcus
pneumoniae, tetapi belum jelas penyebabnya (Kroncke, Golecki dan Jann,
1990).
Salah satu senyawa yang umum terdapat sebagai komponen kapsul
~alah asam N-asetil neuraminat (NeuNAc) yang membentuk struktur polirner
dengan ikatan poll a(2-8). Senyawa ini terdapat pada kapsul Nejsseria men-
ingitidis, €. coli dan streptokokus grup 3 (Pelkonen, Hayrinen dan Finne,
1988). Selain pada prokariot, senyawa poli a(2-8) NeuNAc juga terdapat
pada membran sel saraf yang berfungsi untuk melekatkan sesamanya
(Rutishauer, Acheson, Hall, Mann dan Sunshine, 1-980). Senyawa poli a(2-
8) NeuNAc dapat berperan sebagai imunodeterminan kapsul dan dapat
dikenali oleh antibodi spesifik terhadapnya (Finne, Suermann, Goridis dan
Finne, t987).
Kapsul memifiki fungsi biologis bermacam-macam, di antaranya :
1. Mencegah sel dari kekeringan, cadangan energi, dan membantu perleka-
tan bakteri (Bazeley, 1942; Perry dan Staley, 1997).
2. Aktivitas biologis yang berhubungan dengan patogenesis penyakit seperti
melindungi bakteri dari fagositosis, mencegah aktivasi sistem komplemen,
dan membantu perlekatan dan penyebaran bakteri dalam tubuh (Mims,
1982).
3. Melindungi bakteri terhadap bakteriofag, toksin-toksin dan agen antibakte-
Ti~?jajauan Pustaka : ~ k s h s i Fenotip dan Aktivitas Biologi Streptokokus Grrup C isolat I I
asal Babi dan Kera
, Pada S. equi subsp. zooepidemicus dan streptokokus grup C lain-
nya. kapsul terdiri dari asam hialuronat sebagai bahan penyusunnya (Parker,
1983). Senyawa ini berupa polimer dari disakarida asam P-D glukuronat
dan N-asetil glukosamin yang saling berikatan pada posisi P-1,3. Asam hialu-
ronat disakarida merupakan monomernya (Harper, Rodwell dan Mayes,
1980) (Gambar 1 ). Pertama kali asam hialuronat ditemukan oleh Meyer dan
Palmer pada tahun 1934 dari cairan bulbus oculi sapi (Jeanfoz, 1963), na-
mun sekarang telah berhasil dimurnikan dari berbagai jaringan hewan seperti
trachea sapi, umbilicus (tali pusar) manusia, pial ayam jantan, dan dari S.
equi
subsp. zooepidemicus (Anon., 1997).Senyawa ini tergabung dalam kelompok karbohidrat jaringan yang
di-
namakan mukopolisakarida asam 'atau gtikosaminoglikan asam (Jeanloz,
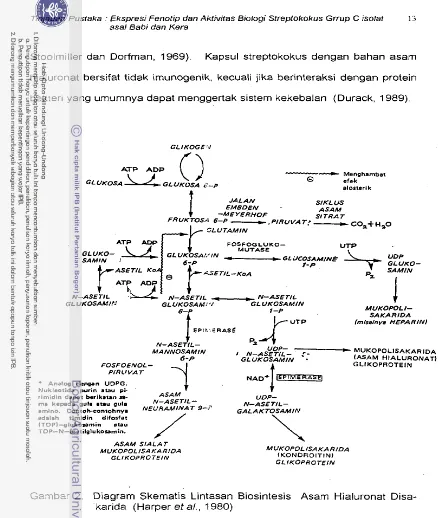
1963). Glukosa adalah prekursor senyawa ini dengan lintasan biosintesis-
nya tertera pada Gambar 2. Pada Gambar 2 nampak lintasan biosintetik
menghasilkan dua senyawa karbohidrat aktif, yaitu uridin di fosfat (UDP), N-
Gambar I .
44r3-Q43~7
NW--co-cHJ-
H OH
ASAM G L ~ u R o ~ T N-ASETIL G L UKOSAMIN
Tinjauan Pustaka : Ekspresi Fenotip dan Aktivifas Bblogi Streptokokus Grrup C isolat 12
asal Babi dan Kera
asetil glukosamin dan UDP- glukuronat. Penggabungan kedua senyawa ini
terjadi akibat bantuan enzirn hialuronat sintase yang rnernerlukan Mg
''
da-lam aktivitasnya (Markovitz dan Dorfrnan,. 1962). Secara biokimiawi
senyawa ini rnerupakan struktur dasar jaringan ikat yang terdiri dari banyak.
molekul protein (proteoglikan) dan anggota kelompok mukopolisakarida lain-
nya seperti kondroitin sulfat dan keratin sulfat. Molekul kelornpok asarn
glikosa~ninoglikan ini banyak dijumpai pada bahan dasar yang bersifat adhe-
sif dan juga dalarn cairan sinovial sendi untuk pelurnas (Lehninger, 1982;
Stryer, 1988).
Asam hialuronat kapsul bakteri bersifat sama dengan asarn hialuronat
jari-ngan. yaitu adhesivitasnya, dan p ~ l i ~ e r i s a s i n y a dirnulai dari bahan dasar
yang sama rnembentuk o-p-D glukopiranosil (7-3) p-D-N asetil glukosamin
piranosida (asam hialuronat disakarida). Dalarn proses ini secara ber-
samaan juga disintesis protein inti (core protein) dengan residu asam amino
serin yang rnenjadi tempat dirnana asarn hialuronat tersebut bertaut (Muir,
1958; Stoolmiller dan Dorfrnan, 1969). Asam hialuronat murni dari strepto-
kokus grup A dan C berbobot rnolekul 580.000 dan mengikat satu residu
serin per rnolekulnya (Markovitz, Cifanolli, dan Dorfrnan, 1959; Stoolmiller
dan Dorfman, 1969). Kapsul streptokokus dengan bahan asarn hialuronat
bersifat tidak irnunogenik, kecuali jika berinteraksi dengan protein bakteri
yang umumnya dapat menggertak sistern kekebalan (Durack, 1989).
Tinjauan P u s t a k a : Ekspresi Fenotip dan A k t i v i t a s Biologi S f r e p f o k o k u s Grrup C i s o l a t 13
asal Babi dan K e r a
Stoolmiller dan Dorfman, 1969). Kapsul streptokokus dengan bahan asam
hialuronat bersifat tidsk imunogenik, kecuali jika berinteraksi dengan protein
bakteri yang urnurnnya dapat menggertak sistern kekebalan (Durack, 1989).
GL IKOGE V
--
MenghambatGL UKOSA GGL UKOSA b-P 0 c f c k
alorteri k 4
JALAN SlKL US EMBDEN - ASAM
-ME YERHOF SI TRA T
FRUKTOSS 6-P
-
, PIRUVAT:--
COa+ H=O-b
GLUTAMINFOSFOGLVKO- P4UTASE
_
,
~ i l C O s A H , N gr-P
N-ASETIL .-- N - A s E T l I GL UKOSAMI.fL GLUKOSAM!'.' GL UKOSAMIN
t
N-ASETIL-
MANNOSAMIN
FOSFOENOL-
R A T
j.
* Anal00 den!+" UDPG. Nukteotida purin etau pi- .
rimidin dapat berikatan sa- ASAM
t
ma kapada gula .tau gula N-ASETIL-
amino. Contch.sontohnya NEURAMINAT
'-'
adalah timidin diforfat (TDPl-glukosmin .tau
TOP-N-autilglu kosmin.
/
ASAM SIALAT MUXOPOLISAKARIDA
GLIKOPROTEIN
t
MUKOPOL I - SAKARIDA (misslnya HEPARIN)
4 N-AS%?L-
-
MUKOPOLISAKARJDAGLUKOSAMIN (ASAM HIALURONAT) 1 GLIKOPROTEIN
NAD+/==='
UDP- N-ASETIL- GALAKTOSAMIh'\
MUKOPOL iSAKAflIDA (KONDROITINJ GLIKOPROTEIN [image:179.518.39.477.13.531.2]Tinja~ian Pustaka : Ekspresi Fenotip dan Aktivitas Biologi Streptokokus Grrup C isolat 14.
asal Babi dan Kera
2.3.2 Dinding Sel
2.3.2.1 Peptidoglikan, Asam Teikoat dan Lipoteikoat
Dinding sel bakteri Gram positif terdjri dari senyawa peptidoglikan
dengan kandungan 40 sampai 96 % materi penyusun dinding sel. Dafam
satu dimensi, peptidoglikan tersusun dari unit disakarida yang terdiri dari N-
asetil glukosamin (G) dan N-asetil muramat (M). Disakar~da tersebut ter-
susun berselang seling secara polimerik yang rnembentuk ikatan glikan.
Dalam dua dimensi, kedua struktur glikan tersebut dihubungkan dengan ika-
tan oligopeptida (interpeptida) yang rnembentuk ikatan seberang silang
(Thorpe, 1984). Struktur kimia peptidogtikan dengan variasi konfigurasinya
tertera pada Gambar 3.
Peptidoglikan berperan sebagai penahan sel dari pengaruh osmotik
lingkungan yang umumnya hipotonik dan secara biofisis struktur ini sangat
efisien (Thorpe, 1984 ). Asam teikoat m@rupakan polimer karbohidrat alkohol
yang dihubungkan melalui ikatan fosfodiester. Panjang senyawa ini rata-rata
6 sampai 10 unit ulangan, tiap unit mengandung residu ester d-alanin dan
N-asetil glukosaminil seperti nampak pada Gambar 3. Ada dua rnacam
asam teikoat yaitu yang mengandung gliserol fosfat dan yang mengandung
gliserol fosfat dan ribitol fosfat. Bentuk gliserol fosfat lebih tersebar luas
dan kedua senyawa ini terdapat baik pada dinding maupun pada membran
sitoplasma sel (Gambar 4) (Davies et
a/.:
1973; Jawetz, Melnick dan Adel-berg. 1982). lnteraksi asam teikoat dengan peptidoglikan tertera pada Gam-
Tinjaoar? Prrsfaka : Ekspresi Fenofip dan Aktivitas Biologi Sfrepfokokus Grrup C isolaf I 5
asal Babi dan Kera
A,-(A..A,.A+i--b-Ala I
o- Ala A, I I A. i
A,
[image:181.511.36.458.14.555.2]I -.G'-.M'-. GI-
Gambar 3. Beberapa Struktur Umum Peptidoglikan dan Konfigurasi Susunan Komponennya (Thorpe, 1984)
Keterangan : G: N-asetil glukosamin, M: N-asetil muramat;
A1
sampai A7 ialah bermacam-macam asam amino pembentuk ikatan peptida( a ) CH.OH
I
CH,OH +I-C-OAla
I RO-C-H
CH,-0 CH,- 0 1
CH,-0
R=Hsturreskhlglubadl CH,- 0 n
Ala = ahliIl R = a- or @N. @ gkkob;rmbr
Ala
-
brbn*lTipe gliserol Tipe ribitol
Garnbar 4. Struktur Kimia Asam Teikoat tipe Gliserol dan tipe Ribitol (Thorpe, 1984)
Ti,ljairat? P~rslalta : Eltsprssi Fe~lotip O ~ I J Alitiv~tas Biologi SStr-eptokok~~s Grrttl~ C isolat I h
asal Babi cia17 Kera
I CH-OR
[image:182.518.57.450.30.562.2]I
Gambar 5 . Hubungan antara Asam Teikoat (Rantai Sampingnya) dengan Polimer Polisakarida dari Peptidoglikan (Thorpe, 1984)
mampu rnengikat M ~ ~ + dan Ca 2' serta memasukannya ke dalam sel.
Setain fraksi ribitol dan gliserol fosfat, asarn teikoat pun mengikat fraksi kar-
bohidrat dan lipid. Pada Streptococcus sanguis, komposisi perbandingan
fosfat, gliserol, karbohidrat dan asam lemak dalam asam lipoteikoat rnernbran
berturut-turut sebesar 1 : 0,95 : 0.71 : 0,046, sedangkan untuk asam lipo-
teikoat dinding sel perbandingan ini berturut-turut sebesar 1 : 0,99 : 0,79 :
0,023 (Chiu, Emdur dan Platt, 1974). Nampak di sini kandungan lemak
asam lipoteikoat membran dua kali kandungan lemak asam iipoteikoat din-
Tir~jauarl Pustaka : Ekspresi Fenofip dan Aktivifas Biologi Strepfokokus Grrup C isolat 17
asal Sabi dan Kera
2.3.2.2 Karbohidrat Dinding Sel
Keberadaan antigen ini mula-mula diketahui oleh Lancefield yang
mengamati suatu komponen polisakarida (substansi C) pada dinding sel
bakteri. Reaksi dengan antiserum spesifik terhadap bakteri tersebut menye-
babkan terbentuknya endapan. Berdasarkan reaksi serologis streptokokus
dibedakan dalam beberapa grup yang dinyatakan dengan A, B, C, dan grup
lainnya.
Streptokokus grup A rnemiliki antigen karbohidrat berupa asam hialu-
ronat yang strukturnya mirip asam hialuronat jaringan mamalia (Davies ef
at.,
1973). Antigen ini beratnya kira-kir.a 10% berat kering sel, dengan N-asetil
glukosamin sebagai determinan antigen untuk grup A dan N-asetil galakto-
samin untuk grup C. Kedua residu ini bertanggung jawab terhadap spesifisi-
tas respon imun yang dihasilkan (Jawetz e t a / . , 1982).
Streptokokus grup B memiliki antigen karbohidrat yang terdiri dari tipe
I, 11, 111, IV, dan V. Tipe I terdiri dari galaktosa dan N asetil glukosamin
(Jelinkova, 1977). Tipe II rnemiliki antigen lebih kompleks, rantai polisakari-
danya rnemiliki 2 rantai sarnping (bercabangf, cabang yang satu mengand-
ung asarn sialat dan cabang lainnya rnemiliki residu galaktosa (Kasper, Gor-
roff dan Baker, 1978; Kasper, Baker, Coldes, Katzenellenbogen dan
Jennings, 1983). Kedua residu ini bertanggung jawab terhadap ekspresi anti-
gen tipe yang bersangkutan. Tipe Ill memiliki antigen karbohidrat yang terdiri
T~njauan Pustaka : Ekspresi Fenotip dan Akfivitas Biologi Streptokokus Grrup C isolat 18
asal Babi dan Kera
Gambar 6. ~truktu; Pembalut Sel Bakteri Kokus Gram Positif (Whithworth, 1990).
Keterangan : P, protein; PL, fosfolipid; h ikatan seberang silang peptida
Asam glukuronat dan asam sialat merupakan imunodeterminan anti-
gen tipe Ill, dimana asam glukuronat lebih dominan mengekspresikan sifat
antigenik tersebut jika dibanding dengan asam sialat (Baker, Kasper dan Da-
vies, 1976; Jelinkova, 1977; Jennings, Lugowski dan Kasper., 1981). Gam-
bar 6 memperlihatkan keberadaan antigen karbohidrat permukaan sel dari
[image:184.520.40.435.26.550.2]Tinjauan Pustaka : Ekspresi Fenotip dan Aktivitas Siologi Streptokokus Grrup C isolat I ! ,
asal Babi dan Kera
2.3.2.3 Antigen Protein
Selain karbohidrat, streptokokus juga memiliki antigen protein. Pada
grup A protein M merupakan antigen utamanya, di samping protein T dan R
yang turut membantu ekspresi sifat antigenik secara keseluruhan (Davies et
a/..
1973; Parker. 1983). Protein M tersebar di permukaan sel, berbentukfimbriae dan memiliki sifat antifagositik. Banyaknya tipe protein M dise-
babkan perbedaan komposisi dan urutan asam amino penyusunnya. Akibat-
nya sifat seperti konformasi dan pelipatan struktur protein menjadi dasar da-
lam kemarnpuan ekspresi sifat antigenik protein M tersebut (Fischetti, 1989).
Cunningham dan Beachey (1 975) mengatakan protein M streptokokus grup A
dapat dikategorikan menjadi tipe spesifik dan non spesifik. Tipe spesifik ber-
peran dalam respon imunogenik ingng yang terinfeksi, sedangkan tipe non
spesifik berperan dalam reaksi siiang dengan komponen endokardium inang.
Protein M pun dapat dikategorikan berdasarkan sensitivitasnya terhadap trip-
sin dan pepsin (Parker, 1983):Atas dasar ini pula, 'beberapa peneliti melaku-
kan klasifikasi antigen protein SGC berdasarkan sifat protein M-nya . (Ti-
money dan Trachman, 1985; Hoffman, Staemfli, Prescott dan Viel, 1991; Ti-
money, Walker, Zhow dan Ding, 1995). Protein M juga menjadi tempat in-
teraksi asam lipoteikoat sehingga diperoleh struktur yang stabil. Sebagian
fraksi asam gliserofosfat I ribitolf~sfat yang bermuatan negatif berinteraksi
dengan residu asam amino protein M yang bermuatan positif (biasanya lisin)
(Ofek, Simpson dan Beachey, 1982). lnteraksi tersebut terjadi dalarn sua-
Tinjaua~l Pustaka : Ekspresi Fenotip dan Aktivitas Biologi Streptokokus Grrup C isolat 2 0
asal Babi dan Kera
hidrofob (Garnbar 7).
Protein T terdiri dari sejumlah protein penting secara irnunokirniawi,
tahan terhadap enzirn proteolitik tetapi sensitif terhadap panas dan asam, ti-
dak larut dalam alkohol dan penyebarannya memberikan sifat antigenik khas
dan dapat dibedakan dengan protein M (Parker, 1983).
Protein R rnerupakan antigen, lain, terdiri dari tipe 3R yang sensitif ter-
hadap tripsin dan pepsin, serta tipe 28R yang hanya sensitif terhadap pepsin.
Protein 3R mirip dengan protein M, tetapi tidak imunogenik (Johnson dan
Beachey, 1979). Timoney (1986) meneliti antigen R pada S . equi subsp. zooepidemicus yang ternyata bersifat tahan asam, panas dan tripsin tetapi
--OOQ90000009090-
1 1 1 , I I 1 1Protein
'
Garnbar 7 . Pengaturan Tulang Punggung Gliserofosfat dari Asarn Lipoteikoat dengan Struktur Alfa Helix Protein M 24 yang Mengandung Se- kelompok Asarn Amino Berrnuatan Positif (biasanya lisin) (Courtney, Hastry dan Ofek,
t
990)peka terhadap pepsin, titik isolistrik 4,8 serta berbobot rnolekul 82 kD.
Protein R ini tidak irnunogenik, dandari beberapa streptokokus grup C lain
Tinjauan Pustaka : Ekspresi Fenotip dan Aktivitas B i o b g ~ Streptokokus Grrup C isolat 2 1 asal Babi dan Kera
Perlrnann, 1952). lsolasi dan karakterisasi protein R pada SGB telah dilaku-
k a n oleh Wibawan dan Laemmler (1991a) dengan bobot molekul 116 kD dan
titik isoelektrik 4,7.
Erickson dan Norcross (1975) mengatakan antigen M dan R dari SGC
mirip dengan antigen M dan R dari Streptococcus grup A, tetapi tidak beraksi
silang. Kemiripan tersebut hanya berdasarkan kepekaan protein-protein ter-
sebut terhadap pepsin dan tripsin, dimana komponen antigen yang peka ter-
hadap tripsin bertanggung jawab terhadap pembentukan antibodi protektif.
Analisis lanjut protein permukaan sel pada
Strepfococcus equi subsp.
equi
memperlihatkan antigen protein 24 dan 29 kD dan 36-37 kD yang bersifat
imunogenik dan diduga mirip protein M dari Strepfococcus grup A, protein
ini dapat bereaksi silang dengan protein 31 dan 55 kD yang berasal dari S.
equi subsp.
zooepidemicus, dan merupakan salah satu reaksi silang dalam
grup C (Tirnoney dah Trachman, 1985).
Keberadaan komponen protein 116,5 kD pada S.
equi
subsp.zooepi-
demicus
menunjukkan perbedaannya dengan S. equi subsp. equi, dan pro-tein ini juga dimiliki oleh S.
equisimilis (Grant, Esferatiou dan Chanter, 1993).
Moore dan Byrans (1969) mengatakanStreptococcus zooepidemicus
me-mitiki paling sedikit 9 tipe antigen yang saling berbeda. Preparasi antigen tipe
I
sampai 8 dapat diekstraksi dengan menggunakan tripsin, sedangkan tipe 9rnenggunakan pepsin. Sifat tahan tripsin ini mirip dengan protein R pada
streptokokus grup A. Antigen tipe 1 sampai 8 tersebar luas pada populasi
Tinjauan Pustaka : Ekspresi Fenotip dan Aktivitas Biologi Streptokokus Grrup C isolat 22 asal Babi dan Kera
Antigen protein lain ialah protein G yang berfungsi sebagai reseptor Fc
dari molekul antibodi (mirip protein A pada Staphylococcus aureus) dengan
bobot molekul 64 kD (Reis, Ayoub dan Boyle, 1984a). Analisa perbandingan
dengan protein A menunjukkan aktivitas saling menghambat antara sesama
prote~n tersebut tetapi secara antigenik kedua antigen protein tersebut ber-
beda. Kedua protein tersebut men-gikat Fc pada lokasi berbeda, selain itu
protein G mengikat IgG sapi dan domba lebih efisien dibandingkan dengan
IgG dari spesies lain (Reis et
a/.,
7984b). Nampaknya protein G terdapat le-bih dari satu macani, selain berbobot 64 kD ada juga yang berbobot 30,dan
45 kD serta dapat membentuk trimer berbobot 130 kD. (Yarnall dan Widders,
1989). Byorck dan Kronvall (1984) juga rnengatakan bahwa secara se-
rologik semua komponen protein G t'ersebut tidak dapat dibedakan.
Hemaglutinin ialah protein permukaan sel streptokokus grup C yang
memiliki aktivitas hemaglutinasi dan sifat ini tidak tergantung pada ke-
beradaan residu asam sialat, hemaglutinasi juga dimiliki oleh Streptococcus
grup C (Kurl, Haataya dan Finne, 1989).
2.4 Antigen Ekstra Seluler
Di samping antigen struktural, streptokokus rnenghasilkan pula antigen
yang dilepaskan ke media pertumbuhannya. Pada umumnya antigen ini
berupa enzim atau protein dengan berbagai aktivitas misalnya :
1 Hemolisin adalah enzim yang bertanggung jawab melisis eritrosit (strepto-
Tifljauan Pustaka : Ekspresi Fenotip dan Aktivitas Biologi Streptokokus Grrup C isolat 23
asal Babi dan Kera
terhadap oksi gen dan streptolisin S (tahan asam dan panas). Strepto-
lisin 0 bertanggung jawab terhadap respon antibodi yang terbentuk, se-
dangkan streptolisin S bertanggung jawab terhadap hemolisis tipe beta
(Parker, 1983)
2 Streptokinase bertanggung jawab terhadap pelarutan fibrin, betfungsi se-
baga~ enzim yang mengaktifkan prekursor plasminogen menjadi plasmin
yang mampu melarutkan fibrin. Streptokinase dihasilkan pula oleh grup A,
C dan G (Jawetz et a/., 1982).
3. Hialuronidase dikenal dengan spreading factor yang berperan untuk
merusak struktur jaringan. Hyaluronidase bersifat antigenik yang khas
untuk grup penghasilnya ( grup A, B, C dan G ) (Davies eta/., 1973).
2.5 Sifat Permukaan Sel Bakteri
Hidrofobik, sifat permukaan sel bakteri yang berkaitan dengan
kemampuannya melekat pada sel inang, telah diamati pada S. mutans dan
S. salivarius (Rosenberg, 1984). Banyaknya protein permukaan ikut me-
nentukan sifat hidrofobisitas sel bakteri seperti protein M pada S. pyogenes
dan protein A pada S. aureus (Tylewska et a/. 1980 yang dikutip dari Mtorner.
Albertson dan Kronvall, 1982). Kaitan antara sifat hidrofobisitas dan per-
lekatan ini pun diamati oleh Laemmler, Pramono, Wibawan, Salasia dan Es-
toepangestie (1993) pada S. suis terhadap eritrosit dan sel Hela. Disimpul-
kan semakin hidrofobik permukaan set bakteri, kemampuan perlekatannya
Tinjauaf? Pustaka : Ekspfesi Fenotip dan Aktivitas Biologi Streptokokus G r u p C isolat 2-1
asal Babi dan Kera
daan galur hidrofobik dan hidrofilik ditentukan oleh jumlah protein yang
diternukan pada permukaan kedua galur bakteri tersebut (Mc Bride, Song.
Krasse dan Olson. 1984). Penentuan jenis dan jumtah protein tersebut dapat
dilakukan dengan mencerna dinding sel bakteri menggunakan mutanolisin
725 unit (U). Ekstraksi protein permukaan sel bakteri menggunakan sodium
dodesil sulfat (SDS) 2 % dan merkaptoetanol 5 %. Setelah itu dilakukan
elektroforesis SDS Poliakrilamide (SDS-PAGE) terhadap ekstrak protein ter-
sebut dan membandingkannya antara bakteri hidrofilik dan hidrofobik (Mc
Bride ef
a/.,
1984). Penentuan derajat hidrofobisitas permukaan sel bakteridapat dilakukan secara kimiawi menggunakan larutan amonium sulfat de-
gan uji Salt-Aggregation Test 1 SAT (Lindahl, Faris, Wadstrom dan Hjerten.
1981 ; Wibawan dan Laemmler, 1990).
Di samping itu, sifat hidrofobik bakteri dapat dideteksi menggunakan
senyawa hidrokarbon ca