ANALISIS KERAGAMAN FENOTIFIK 47 AKSESI SUMBER DAYA GENETIK KELAPA SAWIT (Elaeis guineensis Jacq.) ASAL KAMERUN
THESIS
Oleh:
MAHMUD IRFAN LUBIS NIM: 107001045
PROGRAM MAGISTER AGROEKOTEKNOLOGI FAKULTAS PERTANIAN
ANALISIS KERAGAMAN FENOTIFIK 47 AKSESI SUMBER DAYA GENETIK KELAPA SAWIT (Elaeis guineensis Jacq.) ASAL KAMERUN
THESIS
Oleh:
MAHMUD IRFAN LUBIS NIM: 107001045
Diajukan Sebagai Salah Satu Syarat Untuk Memperoleh Gelar Magister Pertanian Dalam Program Studi Agroekoteknologi pada
Fakultas Pertanian Universitas Sumatera Utara
PROGRAM MAGISTER AGROEKOTEKNOLOGI FAKULTAS PERTANIAN
LEMBAR PENGESAHAN
Judul Penelitian : ANALISIS KERAGAMAN FENOTIFIK 47 AKSESI SUMBER DAYA GENETIK KELAPA SAWIT (Elaeis guineensis Jacq.) ASAL KAMERUN
Mahasiswa : Mahmud Irfan Lubis
N I M : 107001045
Program Studi : Agroekoteknologi
Menyetujui Komisi Pembimbing
Dr. Ir.Lollie Agustina P. Putri, MSi Dr.Ir. A.Razak Purba, MS
Ketua Anggota
Ketua Program Studi Dekan Fakultas Pertanian
Prof. Dr. Ir. Abdul Rauf, MP. Prof. Dr. Ir. Darma Bakti, MS
Telah Diuji Pada :
Tanggal : 4 April 2013
PANITIA PENGUJI TESIS
Ketua : Dr. Ir. Lollie Agustina P. Putri, MSi. Anggota : Dr.Ir.Abdul Razak Purba, MS
Prof. Dr. Ir. B. Sengli J. Damanik, MSc. Prof. Dr. Ir. Rosmayati, MS.
ABSTRAK
MAHMUD IRFAN LUBIS, Analisa Keragaman Fenotifik 47 Aksesi Sumber Daya Genetik Kelapa Sawit (Elaeis guineensis Jacq.) Asal Kamerun dibimbing oleh LOLLIE AGUSTINA P. PUTRI dan ABDUL RAZAK PURBA.
Penelitian ini bertujuan untuk menganalisis keragaman fenotifik data awal vegetatif dari 47 aksesi plasma nutfah introduksi asal Kamerun. Pelaksanaan penelitian dilakukan di Kebun Adolina PT Perkebunan Nusantara IV. Data dianalisa dengan menggunakan metode analisa multivariate yaitu: Komponen Utama (Principal Component Analysis), Analisa Biplot (Biplot Analysis) dan Analisa Neigbor – Joining. Lima karakter morfologi (tinggi tanaman, panjang rachis, jumlah total daun, petiole cross section dan leaf area) dan dua karakter fisiologi (Kandungan klorofil a dan b) menjadi parameter dalam penelitian ini.
Hasil penelitian menunjukkan Analisa Komponen Utama memperoleh dua komponen utama yang masing – masing berperanan sebesar 51,8% dan 24 % terhadap keragaman dari seluruh aksesi Kamerun yang diuji. Analisis biplot menungkap bahwa karakter morfologi vegetatif tidak berhubungan sama sekali dengan karakter fisiologi. Masing – masing aksesi menyebar diantara titik – titik perpotongan dengan garis vektor sesuai dengan besar nilai rerata amatannya. Pohon phylogenetik yang diperoleh dari hasil analisis Neighbor - Joining menunjukkan bahwa seluruh aksesi tidak membentuk suatu kelompok tertentu berdasarkan asal geografisnya.
Analisis komponen utama juga menunjukkan bahwa setiap aksesi menyebar tanpa membentuk suatu kelompok yang mencirikan tempat asal geografisnya, saat diplotkan menurut asalnya. Analisis biplot dan pohon phylogenetik dari Analisis Neigbor Joining menunjukkan bahwa aksesi CMR 32 dan 5 memiliki jarak euclidean yang terbesar diantara aksesi lainnya dan CMR 32 juga memiliki nilai rerata parameter fisiologi yang tinggi namun dengan nilai rerata morfologi vegetatif yang rendah.
Seluruh hasil analisis multivariat menunjukkan bahwa sumberdaya genetik asal kamerun ini memiliki keragaman yang tinggi dan layak digunakan untuk memenambah keragaman populasi kelapa sawit di Indonesia yang telah cukup jenuh.
ABSTRACT
MAHMUD IRFAN LUBIS, Phenotypic Diversity Analysis of 47 Oil Palm (Elaeis guineensis Jacq.) Germplasm of Cameroon Origin, supervised by LOLLIE AGUSTINA P. PUTRI and ABDUL RAZAK PURBA.
The objectives of this reseach was to analyze phenotipe variation of early vegetatif data from 47 germplasm accession introduced from Cameroon. The research was conducted in PT Perkebunan Nusantara IV, Adolina Estate. The data were analyzed by multivariate analysis method which comprised the principal component analysis, biplot analysis, and neigbour – joining analysis. Five morphology characters (the height of plant, the length of rachis, total number of leaves, petiole cross section, and leaf area) and two physiology characters (the content of a and b chlorophyl), became the parameter in this research.
The results of research showed that the Principal Component Analysis indicated two main components which each of them played their role of 51,8% and 24 % on the variation of whole tested Cameroon Accessions. Biplot analysis revealed that morphology characters did not have any correlation with physiology characters. Each accessions scattered amongthe point of intersection lines with vector lines according to the average value of observation. Phylogenetic tree obtained from Neigbor – Joining analysis indicate that whole accessions distributed in a random pattern without composed a spesific group refer to their geographic origin.
Principal Component Analysis (PCA) also showed that each accessions scatters without establishing a cluster which characterized its geographic origin, at the time it was according to its origin. The analysis of biplot and phylogenetic tree from Neighbor – Joining showed that accession CMR 32 and CMR 5 had the bighest Euclidean distance among the others. CMR 32 also had the higher average value of physiology parameters with low mean value of vegetative morphology characters.
All multivariate analysis showed that the genetic resources of Cameroon Origin had high variability which were suitably used in increasing Indonesian’s oil palm population variability which has lately been saturated.
UCAPAN TERIMA KASIH
Alhamdulilah, Puji syukur kehadirat Allah SWT yang telah memberikan rahmat-Nya dan kesempatan kepada penulis dalam menyelesaikan tesis dan program studi Agroekoteknologi, Program Pascasarjana Fakultas Pertanian Universitas Sumatera Utara.
Pada kesempatan ini penulis mengucapkan terima kasih kepada Ibu Dr. Ir. Lollie Agustina P. Putri, MSi sebagai ketua komisi pembimbing dan Bapak Dr.Ir. A. Razak Purba, MS sebagai anggota komisi pembimbing. Dan juga kepada Prof. Dr. Ir. B.Sengli J. Damanik, M.Sc dan Bapak M. Basyuni, S.Hut, Msi, Phd sebagai komisi penguji yang telah banyak memberikan saran, masukan dan bimbingan yang sangat berguna bagi penulis dalam menyelesaikan tesis ini.
Terima kasih penulis sampaikan kepada Rektor Universitas Sumatera Utara Bapak Prof. Dr. Ir. Syahril Pasaribu, DTM&H, MSc (CTM), Sp.A (K)., Direktur Pascasarjana USU Bapak Prof . Dr. Ir. A. Rahim Matondang, MSIE., Dekan Fakultas Pertanian USU Bapak Prof. Dr. Ir. Darma Bakti, MS dan kepada Ketua Program Studi Agroekoteknologi Bapak Prof. Dr. Ir. Abdul Rauf, MP serta segenap dosen Program Magister Agroekoteknologi dan staf tata usaha.
Tidak lupa juga penulis ucapkan terima kasih kepada Direksi PT Perkebunan Nusantara IV dan pimpinan penulis Ibu Ir. Nurmala Dewi Hasibuan MM dan Ir. Sigit Karyadi MSc. atas izin belajar dan bimbingan kepada penulis. Serta kepada kawan-kawan program studi Magister Agroekoteknologi angkatan 2010 dan 2011 Fakultas Pertanian USU.
Medan, April 2013
KATA PENGANTAR
Puji dan syukur penulis ucapkan kepada Allah SWT karena atas kehendakNYA penulis dapat menyelesaikan tesis ini yang berjudul “Analisis
Keragaman Fenotifik 47 Aksesi Sumber Daya Genetik Kelapa Sawit (Elaeis guineensis Jacq.) Asal Kamerun” yang merupakan salah satu syarat untuk dapat
memperoleh gelar magister pada Sekolah Pasca Sarjana Universitas Sumatera Utara, Medan.
Pada kesempatan ini penulis mengucapkan terima kasih kepada dosen pembimbing yang telah banyak membantu penulis untuk menyelesaikan Tesis ini yaitu kepada Ibu. Dr. Ir. Lollie Agustina. P. Putri, MSi. sebagai ketua komisi pembimbing dan kepada Bapak Dr.Ir. A. Razak Purba, MS sebagai anggota komisi pembimbing.
Akhir kata penulis mengucapkan terima kasih, dan semoga tulisan ini dapat bermanfaat.
Medan, April 2013
RIWAYAT HIDUP
Mahmud Irfan Lubis, dilahirkan pada tanggal 25 Desember 1973 di Medan Sumatera Utara Bapak bernama Mohammad Lud Lubis dan Ibu Salmawaty merupakan anak pertama dari tiga bersaudara. Menikah dengan Letty Parinduri, SS dan dikaruniai dua anak putri dan putra yaitu: Annisa Mutiara Irfanty Lubis Kelas V Sekolah Dasar dan Muhammad Rafa Rizqy Lubis Kelas II Sekolah Dasar.
Riwayat pendidikan yang telah dicapai penulis sampai saat ini adalah:
1. Tahun 1980 – 1986, bersekolah di Sekolah Dasar Negeri No.060801 Medan.
2. Tahun 1986 – 1989, bersekolah di Sekolah Menengah Pertama Negeri 10 Medan.
3. Tahun 1989 – 1992, bersekolah di Sekolah Menengah Umum Negeri 1 Medan.
4. Tahun 1992 diterima di Fakultas Pertanian Universitas Sumatera Utara, dan lulus tahun 1997 pada Jurusan Budidaya Pertanian Program Studi Pemuliaan Tanaman.
5. Februari 2011 Penulis diterima menjadi mahasiswa S2 di Sekolah Pasca Sarjana Program Studi Agroekoteknologi, Universitas Sumatera Utara.
DAFTAR ISI
Sumber Daya Genetik, Morfologi, dan Fisiologi Tanaman Kelapa Sawit... 6
Analisis Keragaman Fenotifik Kelapa Sawit... 10
BAHAN DAN METODE PENELITIAN... 15
Tempat Dan Waktu Penelitian... 15
Bahan dan Alat... 15
Desain Percobaan... 15
Analisa Data... 16
Pelaksanaan Penelitian... 19
Perawatan dan Pemupukan Percobaan... 19
Peubah Yang Diamati... 20
Tinggi Tanaman... 20
Panjang Pelepah... 21
Jumlah Lembaran Daun... 21
Penghitungan Lebar dan Tinggi Pelepah... 21
Penghitung Area Luas Daun (Leaf Area)... 21
Pengukuran Kandungan Klorofil a, b dan Total Pada Daun... 22
HASIL DAN PEMBAHASAN... 23
Karakter Morfologi dan Fisiogis Aksesi Kamerun... 23
Analisis Keragaman Genetik... 27
Analisis Faktor... 27
Analisis Komponen Utama (Principal Component Analysis)... 29
Analisis Biplot... 35
Analisis Keragaman dengan Metode Neighbor- Joining... 41
KESIMPULAN DAN SARAN... 45
Kesimpulan... 45
Saran... 46
DAFTAR TABEL
No. Hal
1. Jadwal Pemupukan Tanaman Belum Menghasilkan Tahun Pertama... 20 2. Hasil Statistik Deskriptif Karakter Morfologi dan Fisiologi... 23
3. Hasil Analisis Sidik Ragam Satu Arah Untuk Region Asal Aksesi
Kamerun... 24 4. Nilai Keragaman Proporsi Tiap Faktor... 28
DAFTAR GAMBAR
No. Hal
1. Skema Perkembangan Genus Elaeis... 5 2. Plot Nilai Skor Dari Hasil Analisis Komponen Utama Untuk
Komponen Satu Dan Dua... 32 3. Grafik Analisis Biplot 47 Aksesi Kamerun... 33 4. Pohon Phylogenetik dari 47 Aksesi Sumberdaya Genetik Asal
DAFTAR LAMPIRAN
No. Hal
1. Daftar Plasma Nutfah Dura Aksesi Kamerun Yang ditanam di Adolina... 51 2. Daftar Plasma Nutfah Tenera Aksesi Kamerun Yang ditanam di
Adolina... 52 3. Peta Percobaan AD 03 S dan AD 04 S Kebun Adolina, Afdeling III
Pohon Induk PT.Perkebunan Nusantara IV... 53 4. Peta Zona Agroklimat Asal Aksesi Kamerun... 54
5. Data Curah Hujan Unit Adolina PTPN IV 2009 – 2011... 55 6. Tabel Nilai Rerata Pengamatan 47 Aksesi Sumberdaya Genetik
Asal Kamerun... 56 7. Tabel Hasil Analisa Sidik Ragam Antar Aksesi Plasma Nutfah
Kamerun... 59 8. Tabel Hasil Analisa Kesesuaian Data Metode KMO and Bartlett's
Test dan Tabel Anti-image Matrices... 60 9. Tabel Hasil Analisa Faktor Communalities and Component
Matrix... 61 10. Tabel Hasil Analisa Faktor Rotated Component Matrix dan
Component Transformation Matrix... 62 11. Tabel Hasil Analisa Gerombol Proximities dan Pautan Rata – Rata
(Antar Kelompok)... 63 12. Grafik Hasil Aglomerative Icicle Plotting... 65 13. Tabel Out Put Proximity Matriks Jarak Kuadrat Euclidean Hasil
Analisis Cluster (Gerombol) 47 Aksesi Kamerun... 66 14. Grafik Hubungan Antara Estimasi Berat Kering Daun (Leaf Dry
Weight dengan Kandungan Klorofil A... 69 15. Grafik Hubungan Antara Estimasi Berat Kering Daun (Leaf Dry
Weight dengan Kandungan Klorofil B... 70 16. Grafik Hubungan Antara Estimasi Berat Kering Daun (Leaf Dry
ABSTRAK
MAHMUD IRFAN LUBIS, Analisa Keragaman Fenotifik 47 Aksesi Sumber Daya Genetik Kelapa Sawit (Elaeis guineensis Jacq.) Asal Kamerun dibimbing oleh LOLLIE AGUSTINA P. PUTRI dan ABDUL RAZAK PURBA.
Penelitian ini bertujuan untuk menganalisis keragaman fenotifik data awal vegetatif dari 47 aksesi plasma nutfah introduksi asal Kamerun. Pelaksanaan penelitian dilakukan di Kebun Adolina PT Perkebunan Nusantara IV. Data dianalisa dengan menggunakan metode analisa multivariate yaitu: Komponen Utama (Principal Component Analysis), Analisa Biplot (Biplot Analysis) dan Analisa Neigbor – Joining. Lima karakter morfologi (tinggi tanaman, panjang rachis, jumlah total daun, petiole cross section dan leaf area) dan dua karakter fisiologi (Kandungan klorofil a dan b) menjadi parameter dalam penelitian ini.
Hasil penelitian menunjukkan Analisa Komponen Utama memperoleh dua komponen utama yang masing – masing berperanan sebesar 51,8% dan 24 % terhadap keragaman dari seluruh aksesi Kamerun yang diuji. Analisis biplot menungkap bahwa karakter morfologi vegetatif tidak berhubungan sama sekali dengan karakter fisiologi. Masing – masing aksesi menyebar diantara titik – titik perpotongan dengan garis vektor sesuai dengan besar nilai rerata amatannya. Pohon phylogenetik yang diperoleh dari hasil analisis Neighbor - Joining menunjukkan bahwa seluruh aksesi tidak membentuk suatu kelompok tertentu berdasarkan asal geografisnya.
Analisis komponen utama juga menunjukkan bahwa setiap aksesi menyebar tanpa membentuk suatu kelompok yang mencirikan tempat asal geografisnya, saat diplotkan menurut asalnya. Analisis biplot dan pohon phylogenetik dari Analisis Neigbor Joining menunjukkan bahwa aksesi CMR 32 dan 5 memiliki jarak euclidean yang terbesar diantara aksesi lainnya dan CMR 32 juga memiliki nilai rerata parameter fisiologi yang tinggi namun dengan nilai rerata morfologi vegetatif yang rendah.
Seluruh hasil analisis multivariat menunjukkan bahwa sumberdaya genetik asal kamerun ini memiliki keragaman yang tinggi dan layak digunakan untuk memenambah keragaman populasi kelapa sawit di Indonesia yang telah cukup jenuh.
ABSTRACT
MAHMUD IRFAN LUBIS, Phenotypic Diversity Analysis of 47 Oil Palm (Elaeis guineensis Jacq.) Germplasm of Cameroon Origin, supervised by LOLLIE AGUSTINA P. PUTRI and ABDUL RAZAK PURBA.
The objectives of this reseach was to analyze phenotipe variation of early vegetatif data from 47 germplasm accession introduced from Cameroon. The research was conducted in PT Perkebunan Nusantara IV, Adolina Estate. The data were analyzed by multivariate analysis method which comprised the principal component analysis, biplot analysis, and neigbour – joining analysis. Five morphology characters (the height of plant, the length of rachis, total number of leaves, petiole cross section, and leaf area) and two physiology characters (the content of a and b chlorophyl), became the parameter in this research.
The results of research showed that the Principal Component Analysis indicated two main components which each of them played their role of 51,8% and 24 % on the variation of whole tested Cameroon Accessions. Biplot analysis revealed that morphology characters did not have any correlation with physiology characters. Each accessions scattered amongthe point of intersection lines with vector lines according to the average value of observation. Phylogenetic tree obtained from Neigbor – Joining analysis indicate that whole accessions distributed in a random pattern without composed a spesific group refer to their geographic origin.
Principal Component Analysis (PCA) also showed that each accessions scatters without establishing a cluster which characterized its geographic origin, at the time it was according to its origin. The analysis of biplot and phylogenetic tree from Neighbor – Joining showed that accession CMR 32 and CMR 5 had the bighest Euclidean distance among the others. CMR 32 also had the higher average value of physiology parameters with low mean value of vegetative morphology characters.
All multivariate analysis showed that the genetic resources of Cameroon Origin had high variability which were suitably used in increasing Indonesian’s oil palm population variability which has lately been saturated.
PENDAHULUAN
Latar Belakang Penelitian
Melalui berbagai upaya pengembangan, baik yang dilakukan oleh perkebunan besar, proyek-proyek pembangunan maupun swadaya masyarakat, perkebunan kelapa sawit telah berkembang sangat pesat. Pada tahun 1968, luas areal yang baru 120 ribu ha menjadi 5.160 ribu ha pada tahun 2005 dan pada tahun 2010 diproyeksikan telah mencapai 8.430 ribu ha (Ditjenbun, 2011). Selain pertumbuhan areal yang cukup pesat tersebut, hal lain yang lebih mendasar lagi adalah penyebarannya, yang semula hanya ada pada 3 provinsi saja di Sumatera (dari 27 provinsi), tetapi saat ini telah tersebar di 22 provinsi di Indonesia (dari 33 provinsi). Sumatera masih memiliki areal terluas di Indonesia, yaitu mencapai 65,1% diikuti Kalimantan dan Sulawesi, masing-masing 23,2% dan 2,56%. Untuk skope nasional, produktivitas naik dari 2,93 ton CPO/ha/tahun pada tahun 2005 menjadi 2,98 ton CPO/ha/tahun di tahun 2010. (Kementerian Perindustrian, 2011).
Hal ini sesuai dengan penelitian Carsono (2008) yang menyebutkan Peningkatan produktivitas tanaman umumnya merupakan tujuan yang paling sering dilakukan pemulia dalam merakit suatu kultivar. Hal ini karena peningkatan produktivitas berpotensi menguntungkan secara ekonomi. Bagi petani, peningkatan produktivitas diharapkan dapat mengkompensasi biaya produksi yang telah dikeluarkan. Peningkatan produktivitas (daya hasil per satuan luas) diharapkan akan dapat meningkatkan produksi secara nasional.
Dalam Maizura et al (2006) menyebutkan bahwa kelapa sawit yang ada di Malaysia dan Indonesia berasal dari empat tanaman kelapa sawit yang ditanam di Kebun Raya Buitenzorg di Bogor (sekarang Kebun Raya Bogor) Indonesia. Biji dari pohon ini kemudian ditanam di Deli Sumatra dan kemudian berkembang menjadi populasi “Dura Deli”. Kemudian populasi ini dimamfaatkan sebagai
bagian dari dasar genetik guna perbaikan kelapa sawit di Malaysia dan Indonesia. Akibatnya, dasar genetik di Timur Jauh menjadi sempit.
pertanian, penggantian hutan kelapa sawit alami dengan persilangan DxP berproduksi tinggi yang lebih moderen dan penebangan hutan sawit alami untuk produksi anggur kelapa sawit (palm wine) yang umum di Kamerun wilayah timur. Tujuan yang lain juga untuk memanfaatkan potensi keragaman genetik dari populasi alami kelapa sawit tersebut dan menggabungkannya ke dalam bahan tanaman pemuliaan yang telah lanjut guna mencapai produksi minyak secara kuantitatif dan kualitatif bersamaan dengan karakteristik sekunder lain yang diinginkan. Pada eksplorasi ini tim fokus untuk mengumpulkan contoh yang mewakili dari kelapa sawit alami dari bagian Timur dan Barat Kamerun termasuk sepanjang zona savana. Target permulaan tim adalah untuk mengumpulkan 100 koleksi. Jumlah keseluruhan dapat dikumpulkan 103 koleksi yang dikumpulkan dari 58 lokasi yang mencakup tujuh dari sepuluh provinsi di Kamerun dengan ketinggian tempat berkisar dari permukaan laut sampai dengan 1.326 meter di atas permukaan laut (Purba et al, 2008).
Perumusan Masalah
Keragaman genetik populasi dasar pemuliaan kelapa sawit di Indonesia cukup sempit. Untuk memperluas keragaman tersebut perlu dilakukan introduksi sumber daya genetik baru dari pusat keragaman asal Kelapa Sawit yaitu Afrika dan salah satunya adalah Kamerun. Informasi mengenai karakteristik koleksi plasma nutfah Kamerun yang telah ditanam di Kebun Adolina PT Perkebunan Nusantara IV belum diketahui. Data ini sangat dibutuhkan sebagai informasi awal guna memudahkan dalam penentuan aksesi terbaik untuk diikutsertakan dalam program pemuliaan lanjut guna perakitan varietas baru.
Tujuan Penelitian
Adapun tujuan dari penelitian ini adalah dapat menganalisis keragaman fenotifik data awal vegetatif dari 47 aksesi plasma nutfah introduksi asal Kamerun ditanam di Kebun Adolina PT Perkebunan Nusantara IV
Hipotesis Penelitian
Terdapat perbedaan fenotipik plasma nutfah asal Kamerun berdasarkan aksesi plasma nutfah dan letak geografis asal plasma nutfah yang diamati.
Kegunaan Penelitian
TINJAUAN PUSTAKA
Botani Kelapa Sawit
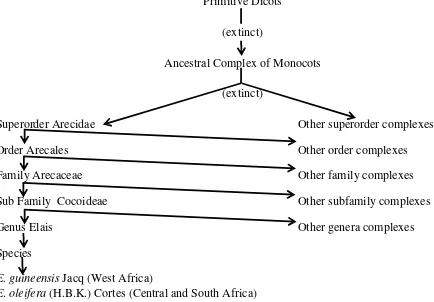
Dalam Latiff (2000) menyatakan bahwa famili Palmae atau Aracaceae genus Elaeis ini dikelompokkan, diperkirakan berusia sama tuanya dengan famili atau tanaman berbunga yang fosilnya ditemukan pada batuan Cretaceous yang berusia kurang lebih 120 juta tahun. Banyak taksonomis tanaman yang mempercayai bahwa monokotiledon yang pertama telah bercabang keluar dari cadangan dikotiledon primitif yang merupakan nenek moyang dari semua monokotiledon. Garis perkembangan keturunan dari genus Elaeis dapat diGambarkan dalam bagian skematik seperti Gambar berikut:
Primitive Dicots
(extinct)
Ancestral Complex of Monocots
(extinct)
Superorder Arecidae Other superorder complexes
Order Arecales Other order complexes
Family Arecaceae Other family complexes
Sub Family Cocoideae Other subfamily complexes
Genus Elais Other genera complexes
Species
E. guineensis Jacq (West Africa)
E. oleifera (H.B.K.) Cortes (Central and South Africa)
Genus Elaeis yang termasuk ke dalam sub famili Cocoidae, yang termasuk juga kelapa sayur, yang dipercaya berasal baik dari Afrika atau Amerika dan merupakan satu dari 240 genera famili Arecaceae yang tidak bisa terlihat pada isolasi dari genera yang lain disebabkan oleh tingginya tingkat homogenitas diantara kromosom dari tanaman palma. Spesies pertama dari genus E. guineensis dinyatakan oleh Jacquin tahun 1763. Spesies kedua merupakan E. oleifera (H.B.K) Cortes, kelapa sawit Amerika Selatan biasanya disebutkan sebagai E.melanococca dan digunakan oleh Gaertner pada tahun 1897 (basionymnya mula
– mula disebutkan sebagai Alfonsia oleifera), oleh H.B.K. di tahun 1816). Spesies
ini dibedakan dari E. guineensis oleh batangnya pertama – tama tegak, segera selanjutnya menjadi terbaring, dan pertumbuhannya lambat. Daunnya berlekatan pada satu bidang dan tidak ada pembengkakan didasarnya, duri pada batang pelepah pendek dan tajam; seludang bunga betina keras, tandan buah mengerucut, dengan mesokarp yang tipis, sewaktu matang berwarna kuning pucat sampai oranye cerah. Selanjutnya ia dikelompokkan pada genus yang lain dan dirujuk sebagai Corozo oleifera oleh (H.B.K.) Bailey.
Sumber Daya Genetik, Morfologi, dan Fisiologi Tanaman Kelapa Sawit Dalam Rajanaidu, et al (2000) bahwa famili Palmae (monocotyledon) dikelompokkan ke dalam enam sub famili. Sub famili Arecordeae dikelompokkan lebih jauh lagi menjadi suku dan sub suku. Genus Elaeis dan Barcella jatuh ke dalam suku Cocoeae dan sub suku Elaeidinae. Genus Elaeis terdiri atas dua spesies, yaitu E. guineenis dan E. Oleifera (2n = 32). Dilaporkan juga bahwa
panjang kromosom kelapa sawit berkisar 1.00 m – 3.89 m. Jumlah DNA dalam
dengan perhitungan satu genom berukuran sekitar 1.8 x 109 pasangan basa. Tidak ada perbedaan dalam penjang kromosom antara E. guineenis dan E. Oleifera. Tanaman kelapa sawit dipercaya berasal dari Afrika Barat meski demikian patut diperhitungkan Amerika Selatan sebagai awalnya. E. oleifera merupakan endemik untuk Amerika Tengah dan Selatan dan telah berhibiridisasi dengan E. guineensis.
lebih setengah panjangnya; berikutnya pelepah akan berpecah ditulang daun, membentuk anak daun yang masih bergabung pada ujung daun; selanjutnya pelepeah memiliki anak daun yang seluruhnya bebas. Jumlah pelepah yang dihasilkan setahun meningkat sebesar 30 – 40 pada umur 3 – 4 tahun dan akhirnya akan menurun menjadi 18 – 25 daun.
Berdasarkan hasil penelitiannya Maizura et al (2006) menemukan bahwa hasil analisa keragaman dari plasma nutfah lainnya yang berasal dari Nigeria, Congo DR, Tanzania, Angola, Senegal, Sierra Leone dan Guinea menunjukkan bahwa populasi liar ini memiliki alel asing yang tidak dimiliki oleh populasi Deli Dura. Ada lima alel yang hilang dalam populasi ini dan diasumsikan bahwa hal ini terjadi akibat pengurangan oleh seleksi (selection drift) dalam proses pemuliaan. Hal ini merupakan penjelasan yang masuk akal untuk fenomena yang terjadi dalam kelapa sawit. Seleksi yang intensif telah dilakukan dalam populasi kelapa sawit selama bertahun – tahun. Bahan tanaman ini hanya membawa sekumpulan alel saja yang hadir di generasi tetuanya. Beberapa alel hilang disebabkan oleh proses seleksi dalam proses program pemuliaan tanaman kelapa sawit. Secara umum untuk semua peubah keragaman genetik yang dihitung, koleksi sumber daya genetik yang diamati menunjukkan tingkat polimorfik yang lebih tinggi dibandingkan dengan populasi Deli Dura. Berdasarkan hasil penelitian ini, polimorfisme kelapa sawit dari Nigeria memiliki nilai tertinggi 67.2%, diikuti dengan Kamerun 65.5%, Kongo DR 62.1%, Tanzania 62.1%, Angola 56.9%, dan Madagaskar 46.6%. Pola yang sama juga muncul pada negara – negara yang berlokasi di bagian barat dari daerah penyebaran kelapa sawit.
Sierra Leone, diikuti oleh Guinea dan Senegal (masing – masing 51.7 dan 55.2%) dan yang terakhir Gambia.
Otitoju dan Onwurah (2010) menyebutkan bahwa produktivitas tanaman merupakan suatu proses yang unik yang sangat tergantung pada jumlah klorofil yang ada di dalam kloroplas. Klorofil merupakan pigmen yang memberikannya karakter warna hijau, yang memainkan peranan yang unik dalam fisiologis, produktivitas, dan ekonomi dari tanaman hijau termasuk Elaeis guineensis. Adapun jumlah klorofil dalam jaringan tanaman dipengaruhi oleh ketersediaan nutrisi dan cekaman lingkungan seperti kekeringan, salinitas, polusi, minyak dan lain sebagainya.
Analisis Keragaman Fenotifik Kelapa Sawit
Dalam Purwantoro et al (2005) kekerabatan secara fenotipe merupakan kekerabatan yang didasarkan pada analisis sejumlah penampilan fenotipe dari suatu organisme. Hubungan kekerabatan antara dua individu atau populasi dapat diukur berdasarkan kesamaan sejumlah karakter dengan asumsi bahwa karakter-karakter berbeda disebabkan oleh adanya perbedaan susunan genetik. Gen merupakan potongan DNA yang hasil aktivitasnya (ekspresinya) dapat diamati melalui perubahan karakter morfologi yang dapat diakibatkan oleh pengaruh lingkungan. Hubungan kekerabatan tersebut dapat dipelajari dengan menggunakan penanda sebagai alat untuk melakukan karakterisasi genetik pada tanaman kelapa sawit karakter dari pelepah kelapa sawit seperti pengukuran karakter daun (panjang pelepah, jumlah pelepah, tebal dan lain – lain), nisbah kelamin dan lainnya.
Hal ini sesuai dengan yang dikemukakan oleh Purba et al, 2000 bahwa induk dalam populasi Deli terkelompok secara erat, yang mencerminkan dasar genetik yang sempit. Keragaman fenotifik antar induk dalam populasi ini mungkin disebabkan oleh tingginya heterozigositas dari empat tetuanya. Adanya kemiripan antara Kamerun dan populasi Deli cukup mengejutkan, disebabkan Populasi Deli dipercaya berasal dari bagian lain Afrika. Bahan tanaman “Kamerun”
diintroduksi ke Indonesia oleh perusahaan perkebunan Jerman pada tahun tiga puluhan. Diduga bahan tanaman itu berasal dari Kamerun disebabkan negara ini dijajah oleh Jerman pada waktu tersebut.
dasar alelik dari populasi ini yang diturunkan hanya dari empat tanaman kelapa sawit yang diintroduksi pada tahun 1848 di Bogor, Indonesia. Rendahnya nilai ini juga menggambarkan seleksi yang berlangsung bertahun – tahun yang mengarah pada tersingkirnya alel – alel tertentu.
Menurut Rajanaidu et al (2000) diketahui secara umum bahwa sempitnya cadangan gen merupakan tantangan utama untuk kemajuan seleksi pada tanaman kelapa sawit. Keadaan ini memberikan dorongan untuk mencari bahan genetik kelapa sawit dari lingkungan alaminya. Setelah perang dunia kedua bahan genetik kelapa sawit di Kongo diambil dari beberapa lokasi. Antara 1961 – 1965, pemulia dari NIFOR (Nigerian Institute for Oil Palm Research) mengumpulkan koleksi bahan genetik kelapa sawit dari pasar lokal dan melalui kepala desa. Bahan tanaman ini kemudian berada di NIFOR. Tujuh puluh dua turunan persilangan terbuka diamati dan tanaman yang terunggul diintroduksi ke dalam program pemuliaan. Blaak (1967) mengambil bahan tanaman di Dataran Tinggi Bamenda di Kamerun dan beberapa diantaranya ditanam di Lobe, Kamerun dan sebagian lainnya disebarkan ke berbagai tempat. Di Pantai Gading, pekerja kelapa sawit Perancis secara sistematis mengamati kelapa sawit liar dan tanaman terpilih dilakukan pengujian keturunan untuk nilai pemuliaannya. Yang paling akhir Institute de Reseherches pour les Huiles et Oleagineux (IRHO) memilih 38
Koleksi Plasma Nutfah Karakterisasi
Evaluasi Pemanfaatan Konservasi
Menurut Soedomo (2006) salah satu cara untuk memperluas keragaman genetik adalah dengan melakukan introduksi tanaman. Namun tidak semua sumber genetik hasil introduksi dapat berkembang dengan baik di Indonesia, sehingga diperlukan adanya persilangan – persilangan dengan jenis lokal. Potensi genetik dari varietas – varietas ini belum teridentifikasi, oleh sebab itu sebelum kita menggunakan sumber genetik tersebut sebagai tertua persilangan perlu dievaluasi terlebih dahulu.
Maizura (2006) menyatakan bahwa di Malaysia sumber daya genetik kelapa sawit tetap dipertahankan diluar lingkungan aslinya pada kebun koleksi telah dikarakterisasi dengan dengan menggunakan beberapa tipe penanda. Usaha pertama yang dilakukan dipusatkan pada sifat – sifat morfologis. Beberapa tanaman dari plasma nutfah ini menunjukkan produksi yang lebih tinggi, pertumbuhan tinggi yang lambat, nilai bilangan iodine yang tinggi, kandungan kernel yang tinggi dan vitamin E yang tinggi.
masukan bagi pengamatan plasma nutfah dan berguna untuk menduga potensi kemajuan genetik (Zulhermana et al, 2010).
Peubah “asal-usul geografis” telah digunakan secara luas oleh pemulia
tanaman kelapa sawit, menjadi kriteria yang penting untuk seleksi populasi saat pembedaan berdasarkan agro morfologi. Dalam skema Seleksi Berulang Timbal Balik (SBB), pemisahan populasi berdasarkan asal geografisnya mampu mengeksploitasi sumber keragaman genetik yang lebih besar dari koleksi plasma nutfah. Penggunaan “asal geografis” sebagai sumber keragaman genetik dari
bahan tanaman kelapa sawit sering digunakan sebagai salah satu cara terbaik untuk menilai plasma nutfah kelapa sawit baru sebelum diintroduksikan ke dalam skema SBB. Penanda molekuler merupakan alat terkini yang digunakan dalam pembedaan populasi jika teknologi ini tersedia. Namun demikian, pada tahap awal penerapan pembedaan berdasarkan “asal geografis” sebagai kriteria untuk
BAHAN DAN METODE PENELITIAN Tempat Dan Waktu Penelitian
Penelitian ini dilakukan di Percobaan AD 03 S dan AD 04 S Kebun Unit Adolina Afd. III Kebun Induk Kelapa Sawit PT. Perkebunan Nusantara IV, yaitu mulai Januari 2012 sampai dengan Juni 2012.
Bahan dan Alat
Bahan yang digunakan dalam penelitian ini meliputi 47 Aksesi plasma nutfah asal kamerun yang ditanam di percobaan AD 03 S dan AD 04 S KebunUnit Adolina Afd. III Kebun Induk Kelapa Sawit PT Perkebunan Nusantara IV.
Alat –alat yang digunakan dalam penelitian ini meliputi: Jangka sorong, Meteran pita 5 m, plat identifikasi individu tanaman, cat, kuas, lembar data isian pengamatan, alat tulis menulis dan lain – lain.
Desain Percobaan
Aksesi Kamerun ini ditanam di Afd 3 benih kebun unit Adolina PT Perkebunan Nusantara IV di Kecamatan Perbaungan Kabupaten Sedang Bedagai Sumatera Utara. Setiap aksesi ditanam dalam 1 plot yang terdiri dari 10 tanaman menurut region asalnya yaitu region 3 (dataran tinggi), region 4 (hutan hujan Monomodal) dan region 5 (hutan hujan bimodal) dengan 2 ulangan.
Analisa Data
1. Statistik Deskriptif, Uji F dan Uji Jarak Berganda Duncan (DMRT)
Data yang dianalisis merupakan nilai rerata dari 20 pohon setiap aksesi di dua ulangan. Analisis statistik desktriptif dilaksanakan untuk setiap variabel pengamatan dari seluruh aksesi. Analisis sidik ragam satu arah dilaksanakan untuk setiap region dari seluruh aksesi dan diuji pada taraf kepercayaan 95% dan dilanjutkan dengan uji jarak berganda Duncan. Menurut Montgomery (2001) model linier statistik tersebut dituliskan sebagai berikut:
Yij = + i + ij
dimana:
Yij = Nilai pengamatan pada region ke-i dan aksesi ke-j
= Nilai Rerata pengamatan
i = Nilai pengamatan pada region ke-i
ij = Nilai galat pada region ke-i dan aksesi ke-j
2. Analisis Keragaman Fenotifik
Minitab 16. Analisis Biplot dilakukan dengan bantuan software MULTBIPLOT (MULTivariate Analysis using BIPLOT) dengan metode yang dikembangkan oleh Villardón (2010).
Pada tahap awal dilaksanakan dua jenis analisa statistik untuk melihat kesesuaian data jika difaktorkan yaitu dengan Uji Keutuhan Barlett dan Ukuran Kesesuaian Contoh Kaiser-Meyer-Olkin (KMO). Hasil Uji Keutuhan Barlett nyata pada nilai p<0.05 memadai untuk dilakukan analisis faktor. Indeks nilai KMO akan berkisar antara 0 – 1 dengan nilai > 0.5 dipertimbangkan cukup untuk dilaksanakan analisis faktor. Analisis faktor dilaksanakan dengan metode principal component (komponen utama) dan dilakukan rotasi dengan metode varimax.
Disamping itu untuk karakter kuantitatif yang memiliki karakter fenotifik yang luas dilakukan penghitungan matrik ketidaksamaan untuk konstruksi pohon phylogenetik. Pohon phylogenetik adalah diagram yang menggambarkan alur dari penurun secara evolusi dari berbagai spesies, organisma, atau gen dari tetua yang sama. Phylogeni berguna untuk menyusun pengetahuan mengenai keragaman biologi, untuk menyusun klasifikasi, dan untuk memberikan pandangan mengenai peristiwa yang terjadi selama proses evolusi (Baum, 2008).
Dasar dari konstruksi matrik ketidaksamaan yang dipergunakan dalam penelitian ini adalah dengan pengukuran jarak atau ke-h misalnya, disimbolkan dengan dih. Nilai dih diperoleh melalui perhitungan jarak kuadrat Euclidean sebagai berikut (Everitt, 1993):
p
dengan nilai dih = jarak kuadrat Euclidean antara objek ke-i dengan objek ke-j atau ke-h,
p = jumlah variabel
Xij = nilai atau data dari objek ke-i pada variable ke-j
Xhj = nilai atau data dari objek ke-h pada variabel ke-j
Data yang dianalisis bervariasi dalam satuannya. Ada data dalam ratusan (panjang rachis) dan ada juga dalam satuan (jumlah klorofil). Perbedaan yang mencolok ini dapat menyebabkan bias dalam Analisis Kelompok, sehingga data asli harus distandarisasi sebelum proses pembentukan matriks ketidaksamaan. Standarisasi dilakukan dengan cara membagi setiap data (xik) dengan simpangan
baku standar dari variabel k yang dihitung pada unit yang terpilih (Perrier et al, 2006).
Pohon phylogenetik dikonstruksi dengan metode Neighbor-Joining yang diperkenalkan oleh Saitou and Nei (1987) yang sering digunakan dalam analisa keragaman genetik. Metode ini menggunakan kedekatan relatif. Pada konstruksi pohon phylogenetik dalam penelitian ini digunakan metode Neighbor-Joining tanpa penyesuaian data (unweighted neighbor – joining), yang menggunakan metode kriteria keseimbangan rata – rata. Oleh karena kriteria kekerabatan dihitung untuk setiap pasangan hasil dari suatu formula kompleks yang mencakup seluruh ketidaksamaan, sehingga kemungkinan dari semua kriteria yang sama untuk beberapa pasangan sangat rendah. Dengan demikian kelompok algoritma yang terjadi hanya satu pasangan pada tiap ulangan dan pohon yang terbentuk selalu pohon biner.
metode unweighted neighbor – joining. Pada metode ini pembentukan pohon menggunakan pohon – pohon yang diperkirakan dari perbedaan – perbedaan bootstrapping yang digunakan untuk menilai ketidakpastian dari struktur pohon
yang dibentuk. Secara nyata, nilai bootstrap yang diberikan pada setiap cabang menunjukkan frekuensi munculnya cabang ini pada pohon yang telah dibootstrap. Sehingga penyebaran aksesi akan lebih jelas pada pohon yang dibootstrap.
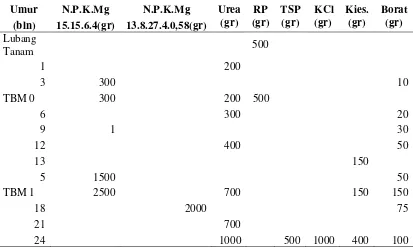
Pelaksanaan Penelitian Perawatan dan Pemupukan Percobaan
Perawatan tanaman dilakukan sesuai dengan Standar Prosedur Operasi untuk PTPN IV diantaranya membuang semua gulma berkayu dengan mendongkelnya, menggarpu lalang atau wiping lalang, membabat gulma lunak yang meninggi pertumbuhannya atau membersihkannya dengan cara kimia, menggaruk pasar pikul, pasar hektar dan piringan dengan ukuran TBM I dan II : 150 cm dan TBM III : 200 cm. Rotasi dongkel kayuan 1x2 bulan, garpu atau wiping 1x2 bulan, piringan dan pasar pikul 1x1 bulan.
Tabel 1. Jadwal Pemupukan Tanaman Belum Menghasilkan Tahun Pertama
Setiap pohon dari tiap aksesi ditandai dengan menggunakan plat dari potongan plat seng yang dicat dengan menambahkan nomor baris dan pohon serta nomor jenis aksesi.
Peubah Yang Diamati
Tinggi Tanaman
Panjang Pelepah
Panjang pelepah diukur merupakan pelepah ke – 17 pada saat pengukuran dengan memulai pengukuran dari pangkal mulai daun rudimenter sampai dengan batas pemisahan daun terminal.
Jumlah Lembaran Daun
Seluruh pada satu sisi pelepah dihitung mulai dari duri rudimenter di pangkal sampai dengan batas pecahnya daun terminal
Penghitungan Lebar dan Tinggi Pelepah
Penghitungan lebar dan tinggi pelepah sering disebutkan dengan istilah pcs (petiole cross section). Lebar dan tinggi pelepah ini diukur pada tempat dimana mulai terbentuknya daun rudimenter .
Penghitung Area Luas Daun (Leaf Area)
Luas daun (leaf area) dapat dihitung dengan menggunakan rumusan yaitu: LA = 2 x n x b x 0,55
Dimana n= jumlah anak daun satu sisi, b = nilai rerata contoh tiga anak daun.
Estimasi Berat Kering Daun (Leaf Dry Weight) Estimasi Berat Kering Daun dihitung dengan rumus:
Pengukuran Kandungan klorofil a, b dan Total Pada Daun
Pengukuran kadar klorofil secara spektrofotometrik didasarkan pada hukum Lamber – Beer. Beberapa metode untuk menghitung kadar klorofil total, klorofil a dan klorofil b telah dirumuskan sesuai dengan metode Arnon (1949) adalah menggunakan pelarut aceton 85 %, dan mengukur nilai absorbansi larutan klorofil pada panjang gelombang (λ) = 663 dan 645 nm.
Setelah diperoleh pembacaan absorbansi lalu dapat dihitung dengan rumus:
klo. a = 12,7 D λ -663 - 2,69 D λ -645 (mg/ l)
klo. b = 22,9 D λ -645 - 4,68 D λ -663 (mg/ l)
HASIL DAN PEMBAHASAN
1. Karakter Morfologi dan Fisiogis Aksesi Kamerun
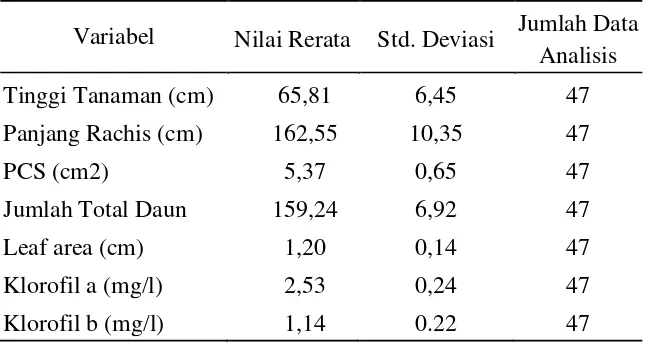
Untuk melihat gambaran secara umum dari seluruh aksesi yang diamati dari masing – masing region, maka dilakukan analisa statistik pendahuluan dengan metode analisis deskriptif. Dari hasil analisis statistik secara deskriptif diperoleh nilai rerata untuk masing – masing variabel karakter morfologi dan fisiologi sebagai berikut:
Tabel 2. Hasil Statistik Deskriptif Karakter Morfologi dan Fisiologi
Variabel Nilai Rerata Std. Deviasi Jumlah Data Analisis
Tinggi Tanaman (cm) 65,81 6,45 47
Panjang Rachis (cm) 162,55 10,35 47
PCS (cm2) 5,37 0,65 47
Jumlah Total Daun 159,24 6,92 47
Leaf area (cm) 1,20 0,14 47
Klorofil a (mg/l) 2,53 0,24 47
Klorofil b (mg/l) 1,14 0.22 47
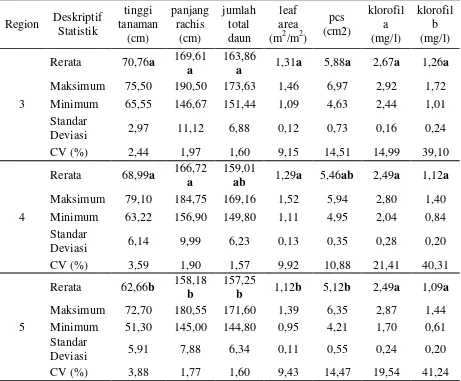
Untuk melihat perbedaan letak geografis dari masing – masing aksesi secara statistik digunakan metode analisis sidik ragam satu arah pada taraf 5 persen, yang dilihat dalam tabel berikut:
Tabel 3. Hasil Analisis Sidik Ragam Satu Arah Untuk Region Asal Aksesi Kamerun Keterangan: angka yang diikuti huruf yang sama pada tiap kolom Rerata peubah menunjukkan tidak berbeda nyata pada uji Duncan pada taraf 5%.
Hasil analisa lanjutan uji jarak berganda Duncan dengan software SPSS menunjukkan adanya perbedaan yang nyata untuk region 3 dan 4 dengan 5 untuk karakter morfologi tinggi tanaman, panjang rachis (pelepah), leaf area namun tidak ditemui adanya perbedaan nyata untuk seluruh karakter di atas antar region 3 dan 4. Untuk variabel jumlah total daun dan petiole cross section tidak ditemui adanya perbedaan yang nyata antar region 3 dan 4 serta 4 dan 5 namun ada perbedaan yang cukup nyata antar region 3 dan 5.
Adanya perbedaan ini menunjukkan lebih jagurnya pertumbuhan aksesi yang berasal dari region 3 yang berlokasi di dataran tinggi (plateau) dibandingkan dengan yang berasal dari hutan hujan bi modal yang berlokasi di dataran yang lebih rendah. Jagurnya aksesi ini dimungkinkan oleh karena saat aksesi ini ditanam di dataran rendah (Adolina) memperoleh intensitas sinar matahari yang lebih tinggi dibandingkan sewaktu ditanam di dataran tinggi.
Untuk karakter fisiologi yaitu kadar klorofil a dan b tidak berbeda nyata untuk semua region. Hal ini berkebalikan dengan seluruh karakter morfologi yang menunjukkan adanya perbedaan yang nyata baik antara region 3 dan 4, 3 dan 5, maupun 4 dan 5.
Meskipun tidak ditemui perbedaan yang nyata pada aksesi antar seluruh region, namun nilai rerata tertinggi untuk jumlah Klorofil a dan b ditemui pada aksesi di region 3 dengan nilai rerata 2,67 mg/l dan 1,26 mg/l. Nilai rerata terendah untuk klorofil a seragam pada region 4 dan 5 yaitu 2,49 mg/l, namun untuk kadar klorofil b nilai rerata terendah diperoleh pada region 5 yaitu 1,09 mg/l.
Nilai koefisien keragaman (CV) terendah terdapat pada variabel jumlah total daun pada seluruh aksesi di region 4 dengan nilai 1,57% dan nilai koefisien keragaman tertinggi terdapat pada karakter fisiologis kadar klorofil a dan b dengan nilai masing – masing 21,41mg/l pada region 4 dan 41,24 mg/l pada region 5.
Karakter morfologis dari masing – masing aksesi yang berasal dari tiga region menunjukkan perbedaan yang nyata secara statistik hal ini sesuai dengan Rafii et al (2002) menemukan bahwa ada perbedaan yang sangat nyata antar lokasi terhadap parameter vegetatif seperti petiole cross section, leaf area, jumlah total daun, panjang rachis dan tinggi tanaman dari beberapa percobaan pengujian keturunan yang terletak di berbagai lokasi di Malaysia.
terbukti memiliki peranan yang penting untuk kadar klorofil sebagai salah satu parameter fisiologis.
2. Analisis Keragaman Genetik
Analisis keragaman genetik dilaksanakan dengan analisis faktor, analisis komponen utama, analisis cluster (kelompok) dan analisis biplot. Untuk rincian lebih jelasnya dapat kita lihat pada bagian berikut:
2.1. Analisis Faktor
Guna mengetahui karakter apa saja yang menimbulkan keragaman secara fenotifik dari seluruh aksesi yang diteliti dari berbagai letak geografis maka dilaksanakan analisi faktor. Pada tahap pertama dilaksanakan uji kelayakan data untuk dianalisis dengan menggunakan metode statistik melalui uji keutuhan Barlett (Bartlett’s test of sphericity) dan uji kesesuaian contoh Kaiser-Meyer-Olkin (KMO) (Kaiser-Meyer-Kaiser-Meyer-Olkin (KMO) measure of sampling adequacy (MSA)). Dalam analisis uji keutuhan Barlett maka hasil uji harus nyata dengan nilai p< 0,05 untuk dilakukan analisis faktor sudah mencukupi. Nilai indeks KMO berkisar antara 0 sampai satu dengan nilai 0,5 sudah mencukupi untuk dilanjutkan dengan analisis faktor (Pallant, 2001).
daun 0.91, leaf area 0.81, kadar klorofil a 0.54, dan kadar klorofil b 0.56. Dengan demikian analisis dapat dilanjutkan dengan analisis multivariate lainnya.
Hasil analisis faktor dengan metode komponen utama dapat dilihat pada tabel berikut:
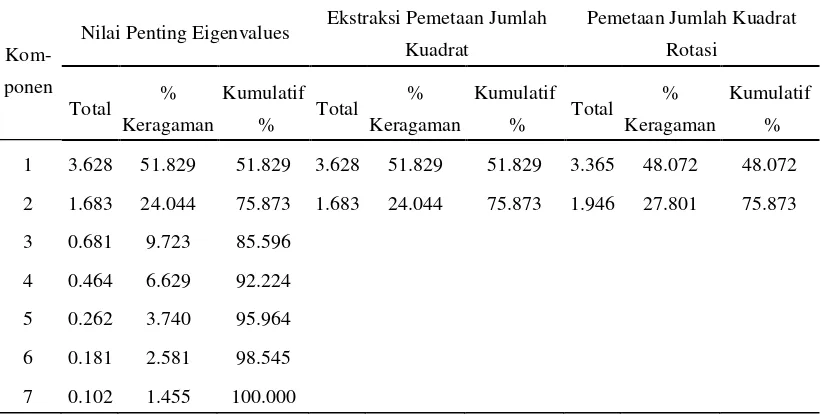
Tabel 4. Nilai Keragaman Proporsi Tiap Faktor
Kom-ponen
Nilai Penting Eigenvalues Ekstraksi Pemetaan Jumlah
Kuadrat besarnya proporsi keragaman yang dapat dijelaskan oleh komponen utama untuk karakter tinggi tanaman (58.4%), panjang rachis (83.6%), petiole cross section (65.8%), jumlah total daun (50.9%), leaf area (84.7%), klorofil a (94.4%) dan klorofil b (93.2%).
variabel yang menyusun sebuah komponen utama lebih jelas dan nyata. Dengan demikian nilai korelasi karakter tinggi tanaman dalam tabel ini adalah 0.75, panjang rachis adalah 0.90, petiole cross section adalah 0.81, jumlah daun adalah 0.69, leaf area adalah 0.92, klorofil a 0.11 dan klorofil b 0.13 pada komponen pertama. untuk komponen kedua tinggi tanaman dalam tabel ini adalah 0.16, panjang rachis adalah 0.19, petiole cross section adalah -0.04, jumlah daun adalah 0.18, leaf area adalah 0.05, klorofil a 0.97 dan klorofil b 0.96.
Pada tabel Component Transformation Matrix pada Lampiran 9 terlihat bahwa nilai koefisien korelasi antara komponen 1 dengan komponen 1 serta komponen 2 dengan komponen 2 memiliki nilai 0.93 yang menunjukkan adanya korelasi yang cukup erat pada saat sebelum maupun sesudah rotasi.
Pengelompokan karakter morfologis maupun fisiologis yang membentuk masing – masing komponen ini sesuai dengan hasil penelitian Oboh dan Fakorede (1992) yang menunjukkan bahwa karakter vegetatif (morfologis) seperti tinggi batang, diameter mahkota, laju penambahan tinggi batang membentuk satu komponen sendiri dan komponen lain yang berasosiasi dengan produksi seperti kadar minyak/mesokarp, persentase mesokarp/buah, persentase cangkang/buah, persentase kernel/buah, dan nilai rerata tandan buah segar, serta produksi tandan buah segar masing – masing membentuk satu komponen tersendiri.
2.2 Analisis Komponen Utama (Principal Component Analysis)
analisis komponen utama. Hubungan antara variabel kuantitatif dan masing – masing komponen utama dapat dilihat dalam Tabel 5.
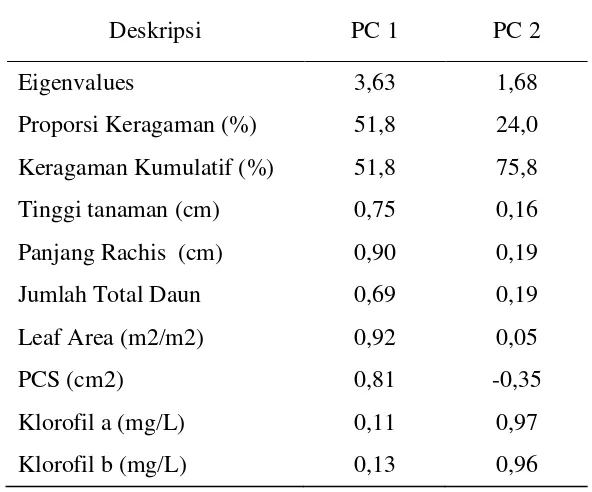
Hasil analisis menyimpulkan ada dua komponen utama yaitu komponen utama 1 (PC 1) dan komponen utama 2 (PC 2) yang membentuk keanekaragaman fenotifik pada 47 aksesi yang diamati. Masing – masing komponen yaitu PC 1 menggambarkan 51.8% dan PC 2 menggambarkan 28.0% keragaman fenotifik pada penelitian ini. Pada PC 1 terdapat lima variabel karakter morfologi yang menyusun komponen tersebut dengan nilai korelasi lebih besar dari 0.5 yaitu tinggi tanaman (0.75), panjang rachis (0.90), jumlah total daun (0.69), leaf area (0.92) dan pcs (0.81). Untuk PC 2 terdapat 2 variabel karakter fisiologi yang menyusunnya yaitu klorofil a (0.97) dan klorofil b (0.96).
Tabel 5. Nilai Eigenvalues Dan Persentase Keragaman Yang Dijelaskan Oleh Kedua Komponen Utama Untuk Semua Aksesi
Deskripsi PC 1 PC 2
Eigenvalues 3,63 1,68
Proporsi Keragaman (%) 51,8 24,0 Keragaman Kumulatif (%) 51,8 75,8
Tinggi tanaman (cm) 0,75 0,16
Panjang Rachis (cm) 0,90 0,19
Jumlah Total Daun 0,69 0,19
Leaf Area (m2/m2) 0,92 0,05
PCS (cm2) 0,81 -0,35
Klorofil a (mg/L) 0,11 0,97
Klorofil b (mg/L) 0,13 0,96
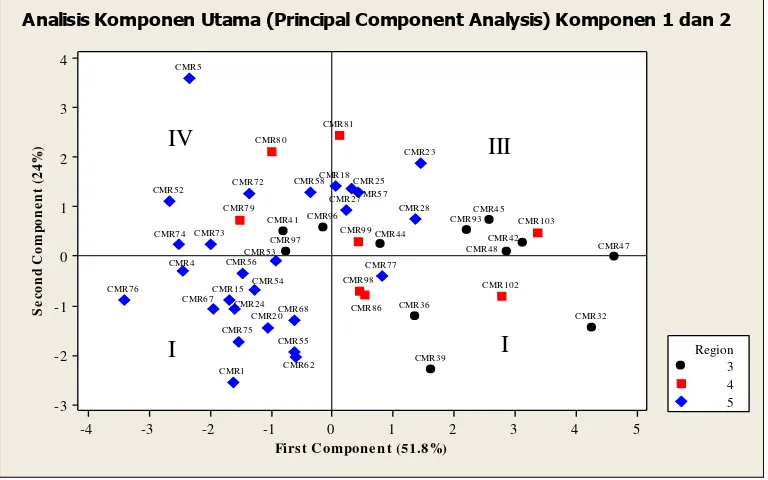
diplotkan pada grafik sesuai dengan nilai skor dari kombinasi linier komponen utama yang dibentuk, dalam penelitian ini diperoleh dua komponen utama yaitu komponen pertama dan komponen kedua. Dari grafik dapat dilihat sebaran dari masing – masing aksesi yang membentuk kelompok yang berdekatan maupun berpisah satu sama lain.
Aksesi CMR 5 terletak paling jauh dari kelompok yang dibentuk oleh nilai skor pada kombinasi linier dari dua komponen utama. Demikian juga halnya dengan CMR 76, CMR 1, CMR 39, CMR 32 dan CMR 47. Dapat dilihat bahwa seluruh aksesi kamerun menyebar secara acak secara fenotifik tanpa mengelompok menurut region asalnya. Namun hampir keseluruhan dari aksesi yang berasal dari region 5 membentuk satu kelompok dan rata – rata terletak pada kwadran pertama dari grafik yang terbentuk yang merupakan kombinasi nilai skor negatif.
Diasumsikan bahwa aksesi yang berasal dari region 5 memiliki pertumbuhan yang kurang jagur jika dibandingkan dengan yang berasal dari region 3, hal ini disebabkan asal lokasi aksesi ini merupakan savana yang memiliki curah hujan lebih rendah jika dibandingkan dengan region 3 yang merupakan dataran tinggi.
meskipun jarak genetik dan geografis berkaitan secara umum, beberapa pengecualian bisa diintroduksikan sebagai introduksi terkini.
Hal yang sama juga ditemui oleh Efombaghn et al (2009) sewaktu nilai rerata dari keseluruhan 16 karakter morfologis dan kualitatif dari penelitian ini diamati, tingkat keberagaman asal aksesi (antara perkebunan dan plasma nutfah) tidak begitu beragam. Meskipun keragaman morfologis menggunakan karakter kuantitatif (PCA) menunjukkan jarak yang berbeda antara aksesi yang berasal dari Perkebunan Barat dan Utara. Namun demikian tidak ditemui adanya perbedaan secara genetik yang dianalisis dengan metode analisis microsatelitte diantara dua sub kelompok ini. Keragaman yang ditemui disebabkan oleh faktor – faktor non
Gambar 2. Plot Nilai Skor dari Hasil Analisis Komponen Utama Untuk
Analisis Komponen Utama (Principal Component Analysis) Komponen 1 dan 2
I I
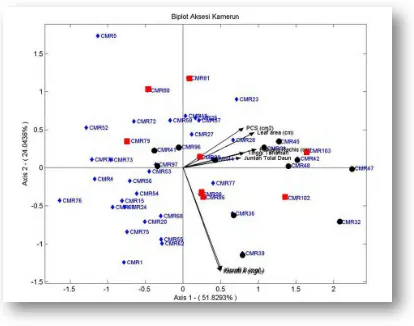
Gambar 3. Grafik Analisis Biplot 47 Aksesi Kamerun
genetik seperti keadaan pertumbuhan kakao secara umum diantara kedua region pertumbuhan ini.
Seluruh aksesi membentuk satu kelompok dengan jarak yang cukup berdekatan secara visual, namun ada beberapa aksesi yang terletak di luar (jauh) dari kelompok yang dibentuk aksesi tersebut yaitu CMR 39, CMR 32 dan CMR 47 yang berasal dari region 3. Sementara dari region 5 terdapat CMR 1, CMR 76 dan CMR 5 yang terletak diluar kelompok.
Seluruh aksesi asal Kamerun tidak membentuk kelompok khas tertentu menurut tempat asal aksesi tersebut dikumpulkan. Dapat disimpulkan bahwa plasma nutfah ini memiliki keragaman yang cukup tinggi diantara aksesi tersebut. Hasil serupa juga diperoleh oleh Ajambang et al (2012) bahwa keragaman genetik populasi liar Kamerun dari tanaman kelapa sawit dan keragaman ini tersebar disemua region (bagian) geografi dari daerah yang disurvei. Pada grafik pohon kekerabatan, 180 individu terkelompok dalam lima kelompok secara jelas menunjukkan hubungan diantara individu. Pengelompokan ini tidak mencerminkan asal dari setiap individu. Kebanyakan dari setiap individu dari tujuh tempat asal ditemui seluruhnya pada keseluruhan lima kelompok yang terbentuk. Keadaan ini merupakan gambaran umum dari Kamerun sebab region – region tersebut berada dalam pengaruh ekologis yang heterogen. Di semua region, kita dapat menemukan dataran tinggi demikian juga dengan dataran rendah, daerah basah dan kering, hutan dan savana dan daerah panas dan dingin. Demikian juga, di setiap provinsi terdapat campuran dari semua varietas Dura, Tenera dan Pisifera atau berdasarkan pada warna buah mentah (Nigrescens atau Virescens). Region atau lokasi asal yang berlainan tidak mengelompokkan aksesi – aksesinya ke dalam kelompok spesifik tertentu namun saling menyebar
Keragaman genetik yang tinggi diperlukan untuk memperoleh heterosis pada turunanya sebagaimana dalam penelitian Maharaj et al (2011) bahwa tetua yang digunakan dalam program pemuliaan tanaman kakao TSH berbeda dari turunannya secara genetik melalui analisis PCA dan gerombol. Kemajuan genetik dalam karakter tertentu ditemukan hal ini menunjukkan bahwa peranan kombinasi heterosis tercapai. Indentifikasi keragaman TSH dapat dilakukan dengan lebih sederhana menggunakan penanda fenotifik.
2.3 Analisis Biplot
Untuk melihat sebaran keragaman aksesi yang cenderung lebih dipengaruhi satu atau beberapa karakter yang diamati diperlukan suatu analisa yang dikenal sebagai analisis Biplot, merupakan visualisasi dari cerminan seperangkat data multivariat yang menunjukkan sejumlah matriks data antara lain:
Struktur varian / kovarian dari struktur variabel
Nilai pengamatan pada variabel
Jarak Euclidean diantara pengamatan dalam ruang multidimensi
Titik perpotongan dari garis tegak lurus dari titik tertentu pada suatu garis variabel, merupakan nilai rerata pengamatan pada variabel yang diwakili garis tersebut. Jika titik perpotongan jatuh pada titik awal garis, diperkirakan nilai pengamatan merupakan rerata dari masing – masing variabel. Jika titik perpotongan jatuh jauh tapi masih searah garis variabel maka ini menunjukkan nilai pengamatan yang tinggi. Sementara jika titik perpotongan jauh di luar garis variabel, yang telah diperpanjang melalui titik asalnya, mewakili nilai yang rendah. Akhirnya jarak antara dua titik kira – kira mewakili jarak Euclidean diantara dua pengamatan dalam ruang multivariat. Pengamatan yang terletak berjauhan satu sama lain memiliki nilai jarak Euclidean yang tinggi dan demikian sebaliknya (Kohler et al,2005).
Dari Gambar 3 dapat dilihat bahwa dari setiap variabel yang diplotkan pada grafik memiliki varians yang beragam dengan keragaman yang tertinggi dimiliki oleh variabel klorofil a dan b. Keragaman yang sedang dimiliki oleh variabel leaf area, pcs, dan panjang rachis. Sementara keragaman yang paling rendah dimiliki oleh jumlah total daun dan tinggi tanaman.
cukup rendah jika dibandingkan dengan panjang rachis yang memiliki keragaman yang lebih tinggi.
Dari seluruh parameter morfologis dapat dilihat bahwa jumlah total daun memiliki hubungan yang cukup erat dengan tinggi tanaman dan panjang rachis dibandingkan dengan leaf area dan pcs. Sudut yang dibentuk oleh jumlah total daun dengan tinggi tanaman dan panjang rachis lebih sempit dibandingkan dengan bentukan sudut bersama variabel morfologis lainnya. Hubungan antara petiole cross section (pcs) dengan leaf area cukup erat dibandingkan dengan variabel – variabel morfologi lainnya. Hal ini ditunjukkan dengan besaran sudut yang di bentuk antara kedua variabel ini lebih kecil dibandingkan dengan variabel morfologis lainnya. Keragaman variabel pcs sedikit lebih rendah jika dibandingkan dengan variabel leaf area. Korelasi antara pcs dan leaf area cukup jauh dengan jumlah total daun yang ditunjukkan dengan bentukan sudut antara garis variabel ini cukup besar dibandingkan bentukan sudut oleh variabel lainnya.
Dari gambaran grafik juga dapat dilihat bahwa bahwa hubungan antara variabel morfologi dan variabel fisiologis baik klorofil a maupun klorofil b sangat kecil atau tidak berhubungan. Hal ini ditunjukkan oleh sudut yang dibentuk oleh garis variabel morfologis dengan variabel fisiologis membentuk sudut sebesar kurang lebih 90o.
untuk aksesi ini lebih mendekati nilai rerata dibandingkan dengan aksesi CMR 52. Dengan demikian peluang pembentukan populasi pemuliaan yang baru cukup besar sehubungan kedua aksesi ini memiliki jarak ketidaksamaan yang cukup jauh dan saling melengkapi diantara keduanya (komplementer), dimana CMR 27 yang sedikit lebih jagur dibandingkan CMR 52 namun aksesi ini memiliki kadar klorofil a dan b yang lebih tinggi . Untuk kombinasi persilangan antara CMR 32 dengan CMR 62 jika merujuk pada Gambar 4 dapat kita lihat bahwa kedua kultivar ini berjarak cukup jauh dan terletak pada kuadran yang berbeda. Posisi CMR 62 yang lebih dekat dengan vektor karakter klorofil a dan b yang diteliti menunjukkan bahwa nilai aksesi ini lebih mendekati nilai rerata jika dibandingkan dengan aksesi CMR 32. Peluang memperoleh populasi baru yang memiliki keragaman yang tinggi dengan karakter seleksi yang penting cukup besar pada kombinasi persilangan ini, CMR 32 juga berasal dari daerah dengan ketinggian tempat yang cukup tinggi sehingga populasi baru yang terbentuk akan memiliki peluang untuk toleran terhadap tempat tumbuh yang cukup tinggi dari permukaan laut.
maupun morfologis yang dapat diasumsikan bahwa aksesi ini memiliki nilai yang rendah dibandingkan dengan nilai rerata dari kedua parameter ini. Sementara meskipun letak dari aksesi CMR 32 menjauhi vektor parameter morfologis namun terletak pada kuadran dimana vektor parameter fisiologis berada. Dengan demikian kedua kombinasi ini bersifat saling melengkapi (komplementer) untuk membentuk suatu populasi pemuliaan baru yang cukup menarik untuk dieksplorasi lebih lanjut.
Strategi pemuliaan tanaman kelapa sawit saat ini beralih pada tujuan untuk memperoleh model tanaman ideal sesuai dengan Chin (2004) bahwa saat ini seleksi untuk tanaman yang ditanam pada populasi tanaman yang bersegregasi secara genetik. Situasi ini akan cenderung mengalami penyimpangan yang mengarahkan seleksi pada tanaman yang lebih vigor dibandingkan dengan tanaman yang lebih lemah. Pengalaman umum dibanyak tanaman lain bahwa produksi dari individu tanaman pada pertanaman campuran tidak berkorelasi dengan baik dengan keturunannya hasil perbanyakan secara seksual maupun klonal yang ditumbuhkan pada tegakan murni. Genotip yang agresif atau vigor tidak menghasilkan kultivar yang sukses. Kultivar modern yang berproduksi tinggi cenderung tumbuh lebih kecil dengan sifat kanopi yang tegak yang memungkinkannya untuk ditanam pada kerapatan yang tinggi dan/atau sanggup menghadapi stress.
saat mengalami defisit air, tergantung pada tingkatan defisit air. Tingkat kerusakan membran berhubungan secara positif dengan kerusakan klorofil. klorofil a, klorofil b, total karotenoid, produksi maksimum kuantum (fotosistem II), dan produksi foton (fotosistem II) pada bibit tanaman yang mengalami defisit air turun secara nyata dibandingkan dengan kelompok kontrol.
Sejarah tanaman kelapa sawit mengarah pada dasar ragam genetik yang sempit pada kultivar – kultivar yang ditanam. Saat ini para pemulia menginginkan karakter – karakter agronomis tertentu seperti: ketahanan penyakit, tingkat kejenuhan minyak, tingkat pertumbuhan yang lambat, dan adaptasi ekologi yang luas, yang keragaman genetiknya terbatas dijumpai pada plasma nutfah E. guineensis. Untuk memperluas dasar genetiknya, pengumpulan plasama nutfah E.
guineensis (Afrika Barat) maupun E. oleifera (Amerika Tropikal) harus
dilaksanakan (Barcelos et al,2002).
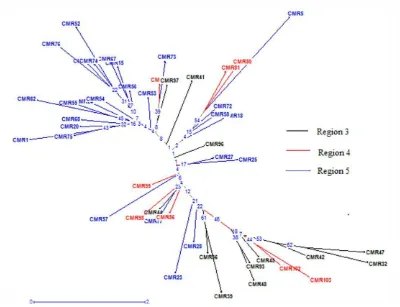
Gambar 4. Pohon Phylogenetik dari 47 Aksesi Sumberdaya Genetik Asal Kamerun dengan Metode Neighbor - Joining
2.4 Analisis Keragaman dengan Metode Neighbor- Joining
membentuk kelompok pada awal grafik phylogenetik ini namun masih ditemui beberapa aksesi asal region ini yang membentuk kelompok dengan aksesi – aksesi yang berasal dari kedua region lainnya. Dengan demikian ekspresi fenotifik yang diukur menunjukkan bahwa keragaman yang muncul dominan dipengaruhi oleh genetik dari aksesi tersebut, sehingga dapat dikatakan peranan lingkungan lebih kecil. Merujuk diagram phylogenetik ini dapat diperkirakan beberapa aksesi yang cenderung digunakan untuk pengembangan populasi ini menjadi suatu program pemuliaan maupun diintroduksikan ke dalam populasi pemuliaan yang sudah ada. Pengembangan suatu populasi pemuliaan baru kelapa sawit seharusnya mempertimbangan 3 prinsip utama yaitu:
1. Daya Gabung Umum (DGU) aksesi tersebut 2. Letaknya berjauhan secara genetik
3. Kombinasi keduanya bersifat komplementer.
Pada prinsip yang pertama untuk nilai daya gabung umum dari aksesi ini masih terlalu dini untuk ditetapkan, dimana nilai DGU tersebut harus diperoleh melalui percobaan pengujian keturunan. Namun demikian dengan mempertimbangkan kedua prinsip lainnya maka dapat diduga kombinasi aksesi mana saja yang berpotensi untuk membentuk satu kelompok populasi pemuliaan baru dan sesuai untuk diintroduksikan ke dalam populasi pemuliaan yang telah ada.
CMR 62 merupakan jenis tanaman dura sedangkan CMR 32 dan 47 merupakan jenis tenera. Dalam program pemuliaan kelapa sawit dengan metode Seleksi Berulang Timbal Balik (SBB), untuk mengetahui daya gabung umum (DGU) dari suatu kombinasi persilangan maka pengujian keturunan dilakukan dengan menyilangkan Dura dan Tenera dan mengevaluasi turunan Teneranya.Untuk tahap awal data kombinasi persilangan antara CMR 62 dengan CMR 32 dan 47 memungkinkan diperolehnya data ini walaupun harus mendapat data kombinasi dari persilangan lainnya.
Salah satu pasangan persilangan lainnya yang memiliki peluang cukup besar adalah CMR 5 dan CMR 32. Kombinasi ini berpeluang untuk membentuk populasi pemuliaan baru yang memiliki keragaman yang tinggi, sehingga akan memungkinkan besarnya intensitas seleksi yang akan dilakukan. Dari Gambar 4 dapat dilihat jarak ketidaksamaan diantara kedua aksesi ini cukup besar dibandingkan dengan aksesi lainnya. Dari analisis biplot juga dapat dilihat bahwa aksesi CMR 32 memiliki karakter morfologis vegetatif yang rendah namun nilai karakter fisiologisnya (klorofil a dan b) cukup tinggi, sedangkan CMR 5 memiliki karakter morfologis yang sama namun nilai karakter fisiologisnya rendah. Kedua aksesi ini cukup memiliki karakter yang komplementer. Karakter fisiologis yang tinggi pada aksesi CMR 32 memungkinkan adanya turunan dari kombinasi persilangan dengan CMR 5 yang memiliki toleransi cekaman kekeringan yang cukup baik.
(Bangoua) dengan ketinggian 1,190 meter dari permukaan laut, maka peluang untuk memperoleh populasi pemuliaan baru yang memiliki daya adaptasi terhadap ketinggian tempat yang cukup tinggi dari permukaan laut lebih besar.
KESIMPULAN DAN SARAN Kesimpulan
1. Berdasarkan hasil analisa komponen utama ditemukan bahwa setiap aksesi menyebar secara merata pada ketiga region asalnya tanpa membentuk pola tertentu.
2. Hasil analisis biplot menunjukkan bahwa aksesi CMR 32 memiliki karakter morfologis vegetatif yang rendah namun nilai karakter fisiologisnya (klorofil a dan b) cukup tinggi, sedangkan CMR 5 memiliki karakter morfologis yang sama namun nilai karakter fisiologisnya rendah.
3. Hasil analisis keragaman berdasarkan metode neighbor – joining juga menunjukkan bahwa tidak ada kelompok khas yang terbentuk dari setiap aksesi yang diamati berdasarkan region tempat asalnya.
4. Dari seluruh hasil analisa multivariat pada penelitian ini baik analisa komponen utama, analisa komponen utama biplot dan analisis keragaman metode neighbor – joining dapat disimpulkan bahwa 47 aksesi kamerun memiliki keragaman fenotifik yang cukup tinggi.
Saran
Disarankan agar dapat dilakukan penelitan lanjutan untuk meneliti keragaman 47 aksesi kamerun dengan penanda molekuler dan morfologis termasuk produksi dan komponen produksi lainnya (kadar minyak, kualitas minyak) untuk mendapat informasi yang lebih lengkap bagi seluruh aksesi Kamerun.
Disarankan agar aksesi CMR 32 dan CMR 5 segera dieksploitasi maupun dintroduksikan ke dalam program pemuliaan PPKS untuk dapat diamati lebih lanjut.
DAFTAR PUSTAKA
Ajambang W., Sudarsono, D. Asmono. dan N. Toruan., 2012.Microsatellite Markers Reveal Cameroon’s Wild Oil Palm Population As A Possible Solution To Broaden The Genetic Base In The Indonesia-Malaysia Oil Palm Breeding Programs. African Journal of Biotechnology Vol. 11(69), pp. 13244-13249, 28 August, 2012.
Allou D., B. Adon., E. Konan, T. D. Gasselin, S.P.N. Assanvo, C. Bakoume, A. Sangare., 2009. Value of Geographical Origin and Parental Genealogy in the Selection of Oil Palm (Elaeis guineensis Jacq.) in Côte d’Ivoire. International Society of Oil Palm Breeding Seminar 2009.
Bakoumé C., R. Wickneswari, N. Rajanaidu, A. Kushairi, P. Amblard, dan N.Billotte, 2011. Allelic diversity of natural oil palm (Elaeis guineensis Jacq.) populations detected by microsatellite markers. Implication in conservation. International Society of Oil Palm Breeder Meeting in Kuala Lumpur 2011.
Barcelos E., A. Philippe, B. Julien dan S. Marc, 2002. Genetic diversity and relationship in American and African oil palm as revealed by RFLP and AFLP molecular markers. Pesq. agropec. bras., Brasília, v. 37, n. 8, p. 1105-1114, ago. 2002.
Baum, D., 2008. Reading a phylogenetic tree: The meaning of monophyletic groups. Nature Education. © 2008 Nature Education.
Carsono N., 2008. Peran Pemuliaan Tanaman dalam Meningkatkan Produksi Pertanian di Indonesia. Dalam Seminar on Agricultural Sciences Mencermati Perjalanan Revitalisasi Pertanian, Perikanan dan Kehutanan dalam kajian terbatas bidang Produksi Tanaman, Pangan, pada tanggal Januari 2008, di Tokyo.
Cha-um S., N. Yamada, T. Takabe, dan C. Kirdmanee, 2011. Manitol-Induce Water Deficit Stress In Oil Palm (Elaeis guineensis Jacq.) Seedlings. Journal of Oil Palm Research Vol.23 December 2011 p.1193 – 1201. Chin S.A., 2004. Selecting The Ideal Oil Palm: What You See Is Not Necessarily
What You Get!.Journal of Oil Palm Research Vol. 16 No. 2, December 2004, p. 121-128.
Dirjenbun, 2011. Buku Statisitik Perkebunan 2009 - 2011. Badan Penelitian dan Pengembangan Pertanian Departemen Pertanian 2011.
Efombagn M. I. B., O. Sounigo, S. Nyassé, M. Manzanares-Dauleux, A. B. Eskes, 2009. Phenotypic variation of cacao (Theobroma cacao L.) on farms and in the gene bank in Cameroon. Journal of Plant Breeding and Crop Science Vol. 1(6). pp. 258-264, August, 2009.
Everitt, B.S. 1993. Cluster Analysis. Third Edition. Halsted Press an Imprint of John Wiley and Sons Inc. New York.