KERAGAMAN MORFOMETRIK
KERANG GELATIK (
Anadara pilula
Reeve 1843)
DI PESISIR PULAU JAWA
YUYUN QONITA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Keragaman Morfometrik Kerang Gelatik (Anadara pilula Reeve 1843) di Pesisir Pulau Jawa adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
RINGKASAN
YUYUN QONITA. Keragaman Morfometrik Kerang Gelatik (Anadara pilula Reeve 1843) di Pesisir Pulau Jawa. Dibimbing oleh YUSLI WARDIATNO dan NURLISA ALIAS BUTET.
A. pilula atau kerang gelatik merupakan salah satu kerang dari genus Anadara yang memiliki distribusi cukup luas di Indonesia. Di dalam daur hidupnya, kerang akan mengalami stadia sebagai meroplankton. Seperti larva kerang meroplankton yang lain, mobilitasnya bergantung kepada arus laut. Setelah melewati stadia meroplankton, kerang akan memasuki fase settlement dan meliang pada substrat. Di habitatnya, kerang akan beradaptasi dengan lingkungan tempat hidupnya, salah satunya melalui adaptasi morfologi cangkang. Tujuan dari penelitian ini adalah untuk mengevaluasi keragaman morfometrik dari kerang gelatik (A. pilula) yang berasal dari pesisir Labuan, Cirebon, Rembang, dan Gresik.
Bahan yang digunakan dalam penelitian ini adalah contoh Anadara pilula. yang diambil dari empat wilayah yang berbeda, yaitu Pesisir Labuan (100 individu), Cirebon (106 individu), Rembang (104 individu), dan Gresik (46 individu). Karakteristik morfometrik kerang A. pilula yang diukur adalah adalah panjang cangkang (P), tinggi cangkang kanan (TA), tinggi cangkang kiri (TI), tebal cangkang (L), panjang ligamen (PL), tinggi umbo (TU), simetri kanan (SA), simetri kiri (SI). Morfometrik kerang diukur menggunakan kaliper digital dengan ketelitian 0.01 mm. Selain itu berat total (BT) dan berat daging (BD) juga diukur menggunakan timbangan digital dengan ketelitian 0.0001 gram. Analisis data yang dilakukan meliputi uji MANOVA satu arah, studi alometri (panjang cangkang-bobot, panjang cangkang-tebal cangkang, panjang cangkang-tinggi cangkang), uji ANCOVA, analisis diskriminan, serta analisis klaster.
Uji MANOVA satu arah dan uji lanjut Games-Howell menunjukkan bahwa terdapat enam karakter yang berbeda secara signifikan antar populasi, yaitu rasio panjang ligamen (PL/P), tinggi cangkang kiri (TI/P), tinggi cangkang kanan (TA/P), tinggi umbo kanan (TUA/P), simetri cangkang kiri (SI/P), dan simetri cangkang kanan (SA/P) (p<0.05). Pola pertumbuhan A. pilula di Pesisir Labuan, Cirebon, dan Rembang berdasarkan hubungan panjang cangkang-bobot total adalah isometrik, sedangkan untuk populasi A. pilula Gresik adalah alometrik negatif. Hubungan panjang cangkang dengan karakter morfometrik lainnya, yaitu tebal dan tinggi cangkang kiri memperlihatkan bahwa pola pertumbuhan A. pilula adalah alometrik negatif. Analisis diskriminan menunjukkan populasi A. pilula dari Pesisir Labuan, Cirebon, Rembang, dan Gresik masing-masing terklasifikasikan secara tepat sebesar 63.0%, 55.7%, 78.3%, dan 64.4%. Analisis klaster menunjukkan bahwa terbentuk dua kelompok, yaitu kelompok pertama yang terdiri dari populasi Labuan dan Rembang serta kelompok kedua yang terdiri dari populasi Cirebon dan Gresik. Analisis morfometrik yang dikaitkan dengan plot arus mengindikasikan bahwa populasi A. pilula di Pesisir Labuan, Cirebon, Rembang, dan Gresik adalah unit stok yang berbeda, sehingga pengelolaannya harus dipisahkan.
SUMMARY
YUYUN QONITA. Morphometric Variation of Pill Ark Cockle (Anadara pilula Reeve 1843) in the Coast of Java Island. Supervised by YUSLI WARDIATNO and NURLISA ALIAS BUTET.
Anadara pilula or pill ark cockle is one of the genus Anadara cockle which distributed widely in Indonesia. It experiences meroplankton stage in its life cycle. All meroplankton likes other larval stage depend on ocean currents for its mobility. Following meroplankton stage, the cockle will experience a settlement phase and stay on the substrate. In its habitat, the cokcle will adapt to the environment, one of them through morphological adaptations. The aim of this study was to evaluate the morphometric variation of A. pilula from the Labuan, Cirebon, Rembang, and Gresik Coast.
Materials used in this study were Anadara pilula samples taken from four different waters, i.e., Labuan (100 pcs), Cirebon (106 pcs), Rembang (104 pcs) and Gresik Coast (46). Morphometric characters which were measured including shell length (P), height of right valve (TA), height of left valve (TI), shell depth (L), ligament length (PL), right valve umbonal height (TUA), left valve umbonal height (TUI), right symmetry (SA), and left symmetry (SI). Morphometric shells were measured using digital caliper with an accuracy of 0.01 mm. Additionally, total weight (BT) and flesh meat (BD) were also measured using digital scales with a precision of 0.0001 grams. The data were analysed using one-way MANOVA test, allometric study (shell length-weight, shell length-shell depth, shell length- shell height), ANCOVA test, discriminant analysis, and cluster analysis.
One way MANOVA and Games-Howell post hoc test showed that there were six characters which were significantly different among the populations, i.e., the ratio of ligament length/shell length (PL/P), heigth of left valve/shell length (TI/P), heigth of right valve/shell length (TA/P), right valve umbonal height/shell length (TUA/P), left symmetry/shell length (SI/P), and right symmetry/shell length (SA/P) (p<0.05). A. pilula growth pattern of shell length-weight relationship in Labuan, Cirebon, and Rembang Coast were isometric, while A. pilula population from Gresik was negative allometric. Shell length relationship with shell depth and shell height of left valve showed that the growth pattern were allometric negative. Based on discriminant analysis, A. pilula from Labuan, Cirebon, Rembang, and Gresik Coast, properly classified as 63.0%, 55.7%, 78.3% and 64.4%, respectively. Cluster analysis showed there were two groups, the first group consisting of Labuan and Rembang population and the second group consisting of Cirebon and Gresik population. Morphometric analysis associated with current plot indicated that A. pilula population in the Labuan Coast, Cirebon Coast, Rembang Coast, and Gresik Coast were different unit stock, so their management should be separated.
© Hak Cipta Milik IPB, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Pengelolaan Sumberdaya Perairan
KERAGAMAN MORFOMETRIK
KERANG GELATIK (
Anadara pilula
Reeve 1843)
DI PESISIR PULAU JAWA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2015
Judul Tesis : Keragaman Morfometrik Kerang Gelatik (Anadara pilula Reeve 1843) di Pesisir Pulau Jawa
Nama : Yuyun Qonita NIM : C251140216
Disetujui oleh Komisi Pembimbing
Dr Ir Yusli Wardiatno, M Sc Ketua
Dr Ir Nurlisa A. Butet, MSc Anggota
Diketahui oleh
Ketua Program Studi
Pengelolaan Sumberdaya Perairan
Dr Ir Sigid Hariyadi, MSc
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
PRAKATA
Puji dan syukur Penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Oktober 2014 ini ialah Keragaman morfometrik kerang gelatik (Anadara pilula Reeve 1843) di Pesisir Pulau Jawa.
Penulis menyampaikan terima kasih kepada:
1. Institut Pertanian Bogor yang telah memberikan kesempatan untuk studi. 2. Pusat Penelitian Lingkungan Hidup (PPLH) IPB yang telah mendanai
sebagian dari penelitian ini.
3. Dr Ir Yusli Wardiatno, MSc selaku ketua komisi pembimbing dan Dr Ir Nurlisa A. Butet, MSc selaku anggota komisi pembimbing yang telah memberi arahan dan masukan dalam penulisan karya ilmiah ini.
4. Prof Dr Ir Mennofatria Boer, DEA selaku penguji tamu dan Dr Ir Etty Riani, MS selaku perwakilan komisi pendidikan Program Studi Pengelolaan Sumberdaya Perairan atas saran dan masukan dalam penulisan karya ilmiah ini.
5. Ayah, ibu, dan keluarga yang telah memberikan dukungan dan doa.
6. Tim Laboratorium Biologi Makro (Bang Aries dan teman-teman) dan tim Laboratorium Biologi Molekuler Departemen Manajemen Sumberdaya Perairan (Kak Wahyu, Kak Panji, Kak Findra, Mba Lela, Mba Fajrin, Mba Yustin, Mba Lita) yang telah memberikan semangat.
7. Teman-teman seperjuangan (Nina, Lufi, Febi, Ayu, Dewi, Nisa, Lusita, Akrom, Agus, Wida, Siska, Mega) atas dukungan yang diberikan.
Semoga karya ilmiah ini bermanfaat.
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
DAFTAR LAMPIRAN vii
1 PENDAHULUAN
Latar Belakang 1
Perumusan Masalah 2
Tujuan Penelitian 3
Manfaat Penelitian 3
2 METODE
Waktu dan Tempat 4
Bahan 4
Analisis Laboratorium 4
Analisis Data 5
3 HASIL DAN PEMBAHASAN
Hasil 10
Pembahasan 19
4 SIMPULAN DAN SARAN
Simpulan 26
Saran 26
DAFTAR PUSTAKA 27
LAMPIRAN 31
DAFTAR TABEL
1 Ukuran tubuh Anadara pilula dari Pesisir Labuan, Cirebon, Rembang,
dan Gresik 10
2 Nilai koefisien keragaman karakter morfometrik Anadara pilula di wilayah Pesisir Labuan, Cirebon, Rembang, dan Gresik 11 3 Hubungan panjang dan bobot Anadara pilula pada Pesisir Labuan,
Cirebon, Rembang, dan Gresik 12
4 Perbandingan hasil regresi nilai logaritmik panjang cangkang (P) terhadap logaritmik bobot total (BT) dan nilai logaritmik panjang cangkang (P) terhadap logaritmik bobot daging (BD) 14 5 Perbandingan karakter morfometrik Anadara pilula dari keempat
wilayah (rata-rata±standar deviasi) 12
6 Perbandingan hasil regresi panjang cangkang (P) terhadap tebal cangkang (L) dan panjang cangkang (P) terhadap tinggi cangkang kiri
(TI) 15
7 Hasil klasifikasi populasi Anadara pilula di masing-masing populasi
berdasarkan analisis diskriminan 17
8 Matriks jarak berdasarkan karakter morfometrik Anadara pilula antara keempat wilayah (Labuan, Cirebon, Rembang, Gresik) 17
DAFTAR GAMBAR
1 Diagram perumusan masalah 3
2 Kerang gelatik (Anadara pilula) yang diambil dari Pesisir Labuan,
Cirebon, Rembang, dan Gresik 4
3 Peta lokasi pengambilan contoh kerang Anadara pilula di Pesisir
Labuan, Cirebon, Rembang, dan Gresik 5
4 Karakter morfometrik kerang gelatik (Anadara pilula) yang diukur pada penelitian ini. Diadaptasi dari Poutiers (1998) 5 5 Distribusi panjang cangkang Anadara pilula di Pesisir Labuan ( ),
Cirebon ( ), Rembang ( ), dan Gresik ( ) 10
6 Hubungan panjang dan bobot Anadara pilula (a) Labuan, (b)
Cirebon (c) Rembang, dan (d) Gresik 13
7 Hubungan panjang dan bobot Anadara pilula (a) Labuan, (b) Cirebon,
(c) Rembang, dan (d) Gresik 14
8 Analisis regresi karakter morfometrik Anadara pilula (a) regresi panjang tebal cangkang (b) regresi panjang cangkang-tinggi cangkang . Labuan, Cirebon, Rembang,
Gresik 16
9 Dendogram populasi Anadara pilula dari empat wilayah (Labuan, Cirebon, Rembang, Gresik) berdasarkan karakter morfometrik 17 10 Plot sebaran morfometrik Anadara pilula dari Pesisir Labuan,
Cirebon, Rembang, dan Gresik berdasarkan analisis diskriminan 18
DAFTAR LAMPIRAN
1 Hasil uji MANOVA satu arah pada karakter morfometrik Anadara
pilula antar lokasi 31
2 Hasil uji ANOVA dan uji t dari regresi antara log panjang dan log bobot total dari Anadara pilula di Pesisir Labuan 32 3 Ringkasan hasil uji ANOVA dan uji t regresi antara log panjang dan
log bobot total dari Anadara pilula di Pesisir Cirebon, Rembang, dan
Gresik 33
4 Hasil uji ANOVA dan uji t dari regresi antara panjang cangkang dengan tebal cangkang total dari Anadara pilula di Pesisir Cirebon 34 5 Ringkasan hasil uji ANOVA dan uji t dari regresi antara panjang
cangkang (P) dengan tebal cangkang (L) dari Anadara pilula di
Pesisir Labuan, Rembang, dan Gresik 35
6 Hasil uji ANCOVA antara populasi Anadara pilula Cirebon dan Gresik pada hubungan linier panjang cangkang-tebal cangkang 35 7 Hasil uji ANCOVA antara populasi Anadara pilula Cirebon dan
Labuan pada hubungan linier panjang cangkang-tebal cangkang 36 8 Ringkasan hasil uji ANCOVA antara populasi Anadara pilula
Labuan, Cirebon, Rembang, dan Gresik pada hubungan linier panjang
cangkang-tebal cangkang 37
9 Ringkasan hasil uji ANCOVA antara populasi Anadara pilula Labuan, Cirebon, Rembang, dan Gresik pada hubungan linier panjang
cangkang-tinggi cangkang 38
1
PENDAHULUAN
Latar Belakang
Wilayah Indo-pasifik merupakan wilayah dengan keragaman bivalvia tertinggi di dunia. Asia Tenggara, termasuk Indonesia merupakan salah satu bagian utama dari wilayah Indo-pasifik (Hutomo dan Moosa 2005). Berdasarkan data terbaru, diketahui terdapat 38 spesies dari subfamili Anadarinae di wilayah ini (Lutaenko 2011), salah satunya adalah Anadara pilula. Distrbusi spesies ini yang telah tercatat terletak di wilayah Indo-Pasifik seperti Filipina (Lozouet dan Plaziat 2008; Dolorosa dan Dangan-Galon 2014), Thailand (Sanpanich 2011), Pulau New Hebrids (Solem 1959), Australia (Li dan Morrison 2011), Hongkong (Cheung dan Wang 2008), Vietnam (Evseev dan Lutaenko 1998), Cina (Feng et al. 2011), dan Indonesia (Mudjiono dan Kastoro 1997; Satrioajie et al. 2013; Akhrianti et al. 2014).
A. pilula atau kerang gelatik merupakan salah satu kerang genus Anadara yang memiliki distribusi cukup luas di Indonesia. Kerang gelatik (A. pilula) banyak ditemukan di pesisir Kota Tegal (Satrioajie et al. 2013), di Panimbang (Mudjiono dan Kastoro 1997), serta di pesisir Belitung Timur (Akhrianti et al. 2014). Di habitatnya, kerang gelatik (A. pilula) sering ditemukan berasosiasi dengan kerang lainnya, seperti kerang darah (A. granosa), A. indica, dan A. inequivalis (Mudjiono dan Kastoro 1997). A. pilula memiliki peran sebagai deposit dan filter feeder (Mudjiono dan Kastoro 1997) yang mengontrol pertumbuhan fitoplankton. Selain itu, kerang Anadara juga berperan sebagai mangsa alami dari gastropoda Natica maculosa, dan Thais carinifera (Broom 1982), T. forbesi, burung pantai, dan ikan pari (Okera 1976).
Kerang gelatik merupakan salah satu spesies famili Arcidae yang memiliki nilai ekonomis penting di Indonesia dan di Asia Tenggara (Poutiers 1998). Kerang ini banyak dimanfaatkan sebagai sumber bahan pangan oleh masyarakat. Kerang gelatik dapat digunakan sebagai salah satu sumber protein alternatif, untuk mengurangi tekanan eksploitasi terhadap kerang darah (A. granosa) yang sangat populer di masyarakat untuk mencegah penurunan stok di alam akibat eksploitasi berlebih.
2
Indonesia, mengingat A. pilula memiliki peran ekonomis dan ekologis yang penting.
Di dalam daur hidupnya, stadia larva kerang merupakan meroplankton yang mobilitasnya bergantung kepada arus laut. Hal ini dapat menyebabkan terjadinya pertukaran genotipe A. pilula antara satu wilayah dengan wilayah lainnya. Pertukaran genotip ini menyebabkan adanya konektivitas antarpopulasi A. pilula. A. pilula memiliki sebaran yang luas di pesisir Pulau Jawa. Kerang ini dapat ditemukan di peisir Labuan, Cirebon, Rembang, serta Gresik. Distribusi yang cukup luas memungkinkan terdapat konektivitas antara A. pilula antarwilayah tersebut.
Setelah melewati masa meroplankton, kerang akan mengalami fase settlement. Setelah meliang pada substrat, kerang akan beradaptasi dengan lingkungan tempat hidupnya. Perbedaan karakter habitat yang terdapat di pesisir Jawa menyebabkan kerang A. pilula harus beradaptasi melalui mekanisme adaptasi fisiologi, morfologi, dan tingkah laku. Adaptasi morfologi yang dilakukan kerang A. pilula dapat menyebabkan kerang tersebut memiliki karakteristik morfologi yang berbeda. Perbedaan karakter morfometrik ini juga dapat disebabkan adanya perbedaan genotip dari masing-masing populasi. Menurut Irie (2006), kajian keragaman morfometrik perlu dilakukan untuk mengetahui perubahan morfometrik biota dalam siklus hidupnya dan dapat memperjelas mekanisme penyebab variasi morfometrik akibat perbedaan geografis. Kajian keragaman morfometrik juga mampu mengevaluasi adanya penciri fenotip pada suatu unit stok, sehingga menjadi penting dalam pengelolaan sumber daya perikanan (Cadrin 2000; Cadrin dan Friedland 2005). Oleh karena itu, dibutuhkan data keragaman morfometrik A. pilula sebagai data base awal serta informasi dalam penentuan strategi pengelolaan dan konservasi A. pilula di pesisir Pulau Jawa.
Perumusan Masalah
Kerang gelatik (A. pilula) memiliki peran ekologis penting dalam rantai makanan sebagai filter feeder dan deposite feeder di wilayah pesisir bersubstrat lumpur atau pasir berlumpur. Selain itu, A. pilula juga memiliki potensi ekonomis yang besar sebagai bahan makanan oleh masyarakat Indonesia karena kebutuhan pangan yang terus meningkat. Kerang ini memiliki sebaran yang cukup luas di pesisir Jawa dan dapat ditemukan di pesisir Labuan, Cirebon, Rembang, serta Gresik dan menjadi salah satu komoditas pangan ekonomis di wilayah-wilayah tersebut. Meskipun demikian, kajian mengenai kerang ini masih sangat terbatas di Indonesia dan di dunia, sehingga upaya pengelolaan terhadap sumber daya menjadi sulit dilakukan.
3
\
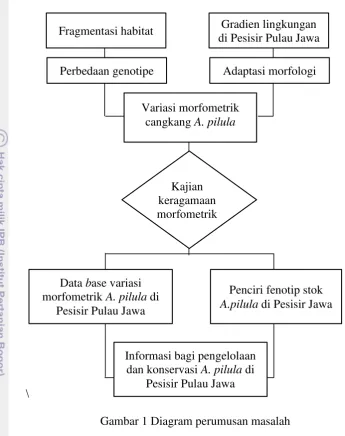
Gambar 1 Diagram perumusan masalah
Tujuan Penelitian
Tujuan dari penelitian ini adalah untuk mengevaluasi keragaman morfometrik (panjang cangkang, panjang ligamen, tebal cangkang, tinggi cangkang kanan dan kiri, tinggi umbo cangkang kanan dan kiri, simetri cangkang kanan dan kiri) dari kerang gelatik (A. pilula) yang berasal dari Pesisir Labuan, Cirebon, Rembang, dan Gresik.
Manfaat Penelitian
Manfaat dari penelitian ini adalah untuk mendapatkan data base keragaman morfometrik kerang gelatik (A. pilula) di Pesisir Labuan, Cirebon, Rembang, dan Gresik, serta mengevaluasi penciri fenotip pada masing-masing unit stok A. pilula yang akan digunakan sebagai salah satu dasar dalam melakukan upaya pengelolaan dan konservasi sumber daya A. pilula di Pesisir Pulau Jawa.
Informasi bagi pengelolaan dan konservasi A. pilula di
Pesisir Pulau Jawa Kajian keragamaan morfometrik Fragmentasi habitat
Variasi morfometrik cangkang A. pilula
Gradien lingkungan di Pesisir Pulau Jawa Perbedaan genotipe Adaptasi morfologi
Penciri fenotip stok A.pilula di Pesisir Jawa Data base variasi
4
2
METODE
Waktu dan Tempat
Penelitian ini dilaksanakan pada bulan Oktober 2014 hingga Maret 2015. Penelitian dilakukan di Laboratorium Biologi Makro, Departemen Manajemen Sumberdaya Perairan, Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor.
Bahan
Bahan yang digunakan dalam penelitian ini adalah contoh Anadara pilula. Contoh A. pilula (Gambar 2) diambil dari empat wilayah yang berbeda, yaitu pesisir Labuan, Cirebon, Rembang, dan Gresik (Gambar 3). Jumlah contoh yang digunakan dalam penelitian ini 100, 106, 104, dan 46 individu masing-masing untuk wilayah pesisir Labuan, Cirebon, Rembang dan Gresik.
Gambar 2 Kerang gelatik (Anadara pilula) yang diambil dari Pesisir Labuan, Cirebon, Rembang, dan Gresik
Analisis Laboratorium
Identifikasi spesies
Identifikasi dilakukan untuk memastikan spesies contoh kerang yang didapatkan berdasarkan karakteristik morfologi. Identifikasi morfologi dilakukan berdasarkan buku identifikasi FAO kelompok bivalvia (Poutiers 1998).
Analisis morfometrik
5 kerang dan bobot kerang masing-masing diukur menggunakan kaliper digital (0.01 mm) dan timbangan digital (0.0001 gram).
Gambar 3 Peta lokasi pengambilan contoh kerang Anadara pilula di Pesisir Labuan, Cirebon, Rembang, dan Gresik
Gambar 4 Karakter morfometrik kerang gelatik (Anadara pilula) yang diukur pada penelitian ini. Diadaptasi dari Poutiers (1998)
Analisis Data
Koefisien keragaman
Nilai koefisien keragaman dihitung dari data karakter morfometrik (PL, L, TA, TI, TUA, TUI, SA, SI) yang telah dirasiokan dengan panjang cangkang (P). Koefisien keragaman digunakan untuk mengetahui keragaman dari masing-masing karakter morfometrik di masing-masing-masing-masing populasi. Rumus koefisien keragaman adalah sebagai berikut.
̅ Keterangan:
6
SD = standar deviasi (simpangan baku) ̅ = nilai rataan
Perbandingan karakter morfometrik antar wilayah
Data yang digunakan dalam analisis perbandingan karakter morfometrik antar wilayah adalah data rasio terhadap panjang cangkang. Hal ini dilakukan untuk menghindari perbedaan karakter morfometrik yang diakibatkan oleh struktur umur yang berbeda. Rasio karakter morfometri yang digunakan adalah ukuran morfometrik (PL, L, TA, TI, TUA, TUI, SA, SI) terhadap panjang cangkang (P). Perbandingan data rasio morfometrik antar wilayah dilakukan melalui uji Multivariate Analysis of Variance (MANOVA) satu arah yang dilanjutkan dengan uji post-hoc Games-Howell pada taraf α=0.05. Model MANOVA yang digunakan dirumuskan sebagai berikut.
ij k ik ijk Keterangan:
yijk = nilai rasio karakter morfometrik terhadap panjang cangkang ke-k
ulangan ke-j pada lokasi ke-i
k = rataan umum dari rasio karakter morfometrik terhadap panjang cangkang ke-k
ik = pengaruh aditif lokasi ke-i terhadap rasio karakter morfometrik terhadap panjang cangkang ke-k
ijk = komponen galat yang timbul pada rasio karakter morfometrik terhadap panjang cangkang ke-k dari ulangan ke-j dan lokasi ke-i
Hubungan panjang dan bobot
Hubungan panjang dan bobot kerang dianalisis untuk masing-masing popualasi kerang. Analisis ini dibutuhkan untuk mengetahui pola pertumbuhan kerang. Hubungan panjang dan bobot dirumuskan sebagai berikut (Ricker 1973).
Keterangan :
W = bobot total (gram) a,b = konstanta
L = panjang (mm)
Hubungan linear dari panjang dan bobot dapat diketahui dengan cara meregresikan nilai 10log dari data panjang dan bobot, sehingga didapatkan persamaan logW=loga+blogL. Pola pertumbuhan kemudian ditentukan dengan menguji nilai b terhadap nilai 3 melalui uji t dengan taraf α=0.05. Jika nilai b=3, maka pola pertumbuhan yang didapatkan adalah isometrik, sedangkan jika nilai b≠3, maka pola pertumbuhannya adalah alometrik Statistik uji t untuk menentukan pola pertumbuhan tersebut dirumuskan sebagai berikut.
7 pertumbuhan alometrik positif (pertumbuhan bobot lebih dominan dibandingkan pertumbuhan panjang), dan jika nilai b<3, maka diketahui pola pertumbuhannya adalah alometrik negatif (pertumbuhan panjang lebih dominan dibandingkan pertumbuhan bobot). Jika nilai thitung<ttabel, maka didapatkan keputusan Gagal Tolak H0 (pola pertumbuhan isometrik). Pola pertumbuhan isometrik mengindikasikan pertumbuhan panjang sama atau sebanding dengan pertumbuhan bobot.
Perbedaan slope antar persamaan linear panjang-bobot logW=loga+blogL antar lokasi diketahui menggunakan Analysis of Covariance (ANCOVA) pada taraf α=0.05, seperti yang telah dilakukan oleh Filguiera et al. (2008) dan Thomas (2013). Nilai slope dibandingkan dengan mempelajari interaksi antara variabel konkomitan (logaritmik panjang cangkang) dengan faktor perlakuan (lokasi). Jika interaksi berbeda signifikan terhadap nol, maka disimpulkan terdapat perbedaan nilai slope antar lokasi. Perbedaan slope ini mengindikasikan adanya perbedaan laju pertumbuhan relatif bobot antar lokasi. Ketika interaksi antara panjang cangkang dan lokasi tidak berbeda signifikan terhadap nol, uji ANCOVA dilanjutkan untuk mengetahui perbedaan nilai intercept antar lokasi. Hal ini dilakukan dengan melihat pengaruh lokasi terhadap variabel respon (logaritmik bobot). Jika pengaruh lokasi berbeda signifikan terhadap nol, maka diindikasikan terdapat perbedaan bobot antar lokasi pada panjang cangkang yang sama. Model ANCOVA yang digunakan untuk menguji pengaruh lokasi terhadap nilai logaritmik bobot adalah sebagai berikut.
ij i ( ij ̅) ij
Keterangan:
ij = nilai logaritmik bobot pada lokasi ke-i dan ulangan ke-j = rataan umum
i = pengaruh aditif dari lokasi ke-i
= koefisien regresi yang menunjukkan ketergantungan yij pada xij
xij = nilai logaritmik ukuran panjang cangkang pada lokasi ke-i dan ulangan ke-j
̅ = rata-rata nilai logaritmik panjang cangkang
8
Hubungan panjang cangkang dengan karakter morfometrik lainnya
Analisis alometri dilakukan dengan meregresikan data panjang cangkang terhadap tebal cangkang serta panjang cangkang terhadap tinggi cangkang kiri sehingga didapatkan persamaan y=a+bx. Nilai a dan b pada persamaan regresi dirumuskan sebagai berikut.
n∑i nn i i ∑ni i (∑ni i)
∑ni i ∑ni i
̅ ̅
Nilai slope yang didapatkan kemudian diuji terhadap nilai 1 menggunakan
uji t p d t r f α 0.05. Uji t dilakukan untuk mengetahui pola pertumbuhan kerang. Jika b=1, maka pola pertumbuhan yang didapatkan adalah isometrik,
sed ngk n jik ≠ , m k pol pertum uh nn dalah alometrik.
thit
∑ ∑
Keterangan:
thit = nilai t hitung b = slope
Sb = standar eror nilai b
Nilai thitung kemudian dibandingkan dengan nilai ttabel ( ; n-2) pada tabel t-student dengan taraf α=0.05. Jika nilai thitung>ttabel, maka didapatkan keputusan Tolak H0 (pola pertumbuhan alometrik). Jika nilai b>1, maka didapatkan pola pertumbuhan alometrik positif (pertumbuhan tebal atau tinggi cangkang lebih dominan dibandingkan pertumbuhan panjang cangkang), dan jika nilai b<1, maka diketahui pola pertumbuhannya adalah alometrik negatif (pertumbuhan panjang cangkang lebih dominan dibandingkan pertumbuhan tebal atau tinggi cangkang). Jika nilai thitung<ttabel, maka didapatkan keputusan Gagal Tolak H0 (pola pertumbuhan isometrik). Pola pertumbuhan isometrik mengindikasikan pertumbuhan panjang cangkang sama atau sebanding dengan pertumbuhan tebal atau tinggi cangkang.
Perbedaan slope dan intercept persamaan regresi antar lokasi kemudian diuji menggunakan uji ANCOVA untuk mengetahui perbedaan laju pertumbuhan dan ukuran antara dua populasi kerang seperti yang telah dilakukan oleh Thomas (2013). Prosedur dan model ANCOVA yang digunakan dapat dilihat pada subbab sebelumnya (hubungan panjang cangkang dan bobot).
Analisis klaster
9 Minitab 16. Dendrogram kemudian dikonstruksi berdasarkan matriks jarak tersebut menggunakan metode UPGMA (Unweighted Pair Group Method with Arithmetic Mean) pada software MEGA 5.0 (Tamura et al. 2011) sesuai petunjuk Kumar et al. (1993). Analisis klaster digunakan untuk melihat kekerabatan dan tingkat kesamaan masing-masing populasi berdasarkan karakter morfometrik yang diukur. Berikut merupakan rumus jarak Mahalanobis yang digunakan sebagai dasar konstruksi dendrogram.
̅i ̅j ̅i ̅j
Keterangan :
D2 = nilai jarak Mahalanobis antara kerang dari wilayah ke-i dan wilayah ke-j i
̅ = vektor nilai rataan pengamatan dari wilayah ke-i pada masing-masing peubah kuantitatif
j
̅ = vektor nilai rataan pengamatan dari wilayah ke-j pada masing-masing peubah kuantitatif
- = kebalikan matriks gabungan ragam peragam antar peubah
Analisis diskriminan
Analisis diskriminan dilakukan untuk mengelompokkan data berdasarkan variabel-variabel kuantitatif. Analisis ini dilakukan untuk mengetahui pengelompokkan kerang A. pilula berdasarkan karakter morfometrik yang telah diukur. Melalui analisis ini juga akan didapatkan persentase nilai sharing component yang terjadi akibat kemiripan morfometrik antar wilayah yang menyebabkan terjadinya kesalahan pengelompokkan. Analisis diskriminan dilakukan berdasarkan metode stepwise menggunakan software SPSS 17. Model analisis diskriminan yang digunakan adalah sebagai berikut.
k k Keterangan:
D = skor diskriminan b = koefisien diskriminasi
x = variabel independen (rasio karakter morfometrik)
Analisis pola arus
10
3
HASIL DAN PEMBAHASAN
Hasil
Ukuran tubuh A. pilula
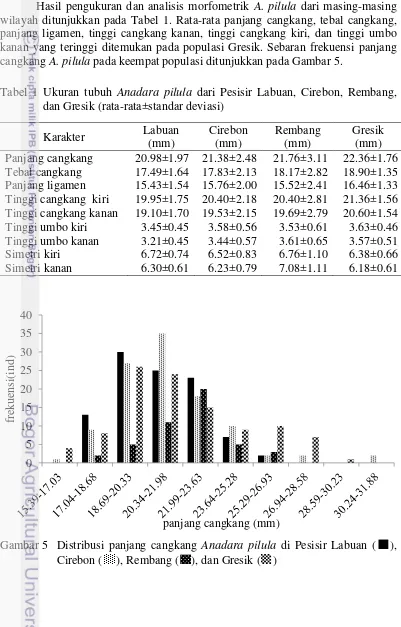
Hasil pengukuran dan analisis morfometrik A. pilula dari masing-masing wilayah ditunjukkan pada Tabel 1. Rata-rata panjang cangkang, tebal cangkang, panjang ligamen, tinggi cangkang kanan, tinggi cangkang kiri, dan tinggi umbo kanan yang teringgi ditemukan pada populasi Gresik. Sebaran frekuensi panjang cangkang A. pilula pada keempat populasi ditunjukkan pada Gambar 5.
Tabel 1 Ukuran tubuh Anadara pilula dari Pesisir Labuan, Cirebon, Rembang, Panjang cangkang 20.98±1.97 21.38±2.48 21.76±3.11 22.36±1.76 Tebal cangkang 17.49±1.64 17.83±2.13 18.17±2.82 18.90±1.35 Panjang ligamen 15.43±1.54 15.76±2.00 15.52±2.41 16.46±1.33 Tinggi cangkang kiri 19.95±1.75 20.40±2.18 20.40±2.81 21.36±1.56 Tinggi cangkang kanan 19.10±1.70 19.53±2.15 19.69±2.79 20.60±1.54 Tinggi umbo kiri 3.45±0.45 3.58±0.56 3.53±0.61 3.63±0.46 Tinggi umbo kanan 3.21±0.45 3.44±0.57 3.61±0.65 3.57±0.51 Simetri kiri 6.72±0.74 6.52±0.83 6.76±1.10 6.38±0.66 Simetri kanan 6.30±0.61 6.23±0.79 7.08±1.11 6.18±0.61
11 Berdasarkan sebaran panjang cangkang individu dari keempat populasi terlihat bahwa populasi A. pilula yang berasal dari Pesisir Cirebon memiliki sebaran terluas dengan panjang minimum 16.6 mm dan panjang maksimum sebesar 31.85 mm. Populasi A. pilula dari Pesisir Labuan memiliki wilayah (range) data yang tersempit, yaitu dengan panjang cangkang minimum 17.4 mm dan panjnag cangkang maksimum 26.18 mm. Menurut Poutiers (1998), ukuran maksimum panjang cangkang A. pilula adalah 40 mm.
Koefisien keragaman karakter morfometrik
Hasil perhitungan koefisien keragaman dari masing-masing karakter morfometrik di masing-masing populasi menunjukkan bahwa rasio karakter tinggi umbo terhadap panjang merupakan karakter yang memiliki nilai koefisien keragaman terbesar di populasi Labuan, Cirebon, dan Gresik. Berbeda halnya dengan populasi A. pilula di Pesisir Rembang, karakter tinggi umbo kiri merupakan karakter dengan nilai koefisien keragaman terbesar dibandingkan karakter lainnya. Nilai koefisien keragaman secara rinci dari masing-masing karakter morfometrik di masing-masing populasi disajikan pada Tabel 2.
Tabel 2 Nilai koefisien keragaman karakter morfometrik Anadara pilula di wilayah Pesisir Labuan, Cirebon, Rembang, dan Gresik
Rasio karakter Koefisien keragaman (%)
Labuan Cirebon Gresik Rembang Rataan
L/P 3.72 4.51 4.27 4.07 4.14
PL/P 4.44 3.57 3.57 4.49 4.02
TI/P 3.01 3.01 2.86 2.85 2.93
TA/P 2.85 3.20 2.94 2.81 2.95
TUI/P 8.93 9.24 9.27 9.73 9.29
TUA/P 10.19 10.78 11.22 8.34 10.13
SI/P 6.69 7.15 7.21 7.26 7.08
SA/P 6.51 7.49 7.45 6.65 7.03
Perbandingan karakter morfometrik antarlokasi
12
Hasil analisis hubungan panjang dan bobot total dari masing-masing wilayah menunjukkan bahwa pola pertumbuhan A. pilula di Pesisir Labuan, Cirebon, dan Rembang adalah isometrik, sedangkan pola pertumbuhan populasi A. pilula dari Pesisir Gresik adalah alometrik negatif (Tabel 3). Grafik hubungan panjang dan bobot ditunjukkan pada Gambar 6 dan Gambar 7.
Tabel 4 Hubungan panjang dan bobot Anadara pilula pada Pesisir Labuan,
13 hubungan panjang cangkang-bobot total pada umumnya menunjukkan nilai koefisien determinasi (R2) yang lebih besar dibandingkan persamaan panjang cangkang-bobot daging. Nilai logaritmik panjang cangkang dan bobot diregresikan dari masing-masing wilayah lalu dibandingkan dengan uji ANCOVA untuk melihat adanya perbedaan laju pertumbuhan atau kemungkinan adanya perbedaan bobot daging antar populasi. Contoh perhitungan tabel sidik ragam regresi log panjang cangkang-log bobot dan uji t nilai b ditunjukkan pada Lampiran 2. Hasil uji ANCOVA bobot total dan bobot daging A. pilula dari keempat lokasi disajikan pada Tabel 4.
(a) (b)
(c) (d)
Gambar 6 Hubungan panjang dan bobot Anadara pilula (a) Labuan, (b) Cirebon, (c) Rembang, dan (d) Gresik
14
(c) (d)
Gambar 7 Hubungan panjang dan bobot Anadara pilula (a) Labuan, (b) Cirebon, (c) Rembang, dan (d) Gresik
Tabel 5 Perbandingan hasil regresi nilai logaritmik panjang cangkang (P) terhadap logaritmik bobot total (BT) dan nilai logaritmik panjang cangkang (P) terhadap logaritmik bobot daging (BD)
Lokasi y=a+bx R2 N ANCOVA
Slope Intercept Panjang cangkang-bobot total
Labuan logW =-3.05+2.71logL 0.82 100 Paralel
(p>0.05) a1 ≠ 2 Cirebon logW =-3.21+2.85logL 0.91 106
Labuan logW =-3.05+2.71logL 0.82 100 Paralel
(p>0.05) a1 ≠ 2 Gresik logW =-2.76+2.58logL 0.87 37
Cirebon logW =-3.21+2.85logL 0.91 106 Paralel
(p>0.05) a1 ≠ 2 Gresik logW =-2.76+2.58logL 0.87 37
Labuan logW =-3.05+2.71logL 0.82 100 Paralel
(p>0.05) a1 ≠ 2 Rembang logW =-3.13+2.88logL 0.95 104
Cirebon logW =-3.21+2.85logL 0.91 106 Paralel
(p>0.05) a1 ≠ 2 Rembang logW =-3.13+2.88logL 0.95 104
Gresik logW =-2.76+2.58logL 0.87 37 Paralel
(p>0.05) a1 ≠ 2 Rembang logW =-3.13+2.88logL 0.95 104
Panjang cangkang-bobot daging
Labuan logW =-3.65+2.68logL 0.68 100 Paralel
(p>0.05) a1 ≠ 2 Cirebon logW =-3.89+2.92logL 0.86 106
Labuan logW =-3.65+2.68logL 0.68 100 Paralel
(p>0.05) a1 ≠ 2 Gresik logW =-4.14+3.10logL 0.62 37
Cirebon logW =-3.89+2.92logL 0.86 106 Paralel
(p>0.05) a1 =a2 Gresik logW =-4.14+3.10logL 0.62 37
Labuan logW =-3.65+2.68logL 0.68 100 Non-Paralel
(p<0.05) - Rembang logW =-4.02+3.03logL 0.95 104
Cirebon logW =-3.89+2.92logL 0.86 106 Paralel
(p>0.05) a1 ≠ 2 Rembang logW =-4.02+3.03logL 0.95 104
Gresik logW =-4.14+3.10logL 0.62 37 Paralel
(p>0.05) a1 ≠ 2 Rembang logW =-4.02+3.03logL 0.95 104
15
Hubungan panjang cangkang dengan karakter morfometrik lainnya
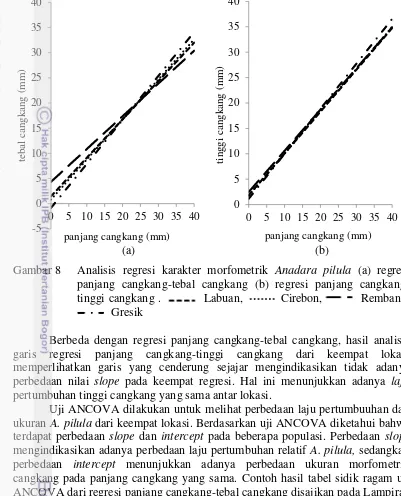
Analisis regresi antara panjang cangkang-tebal cangkang dan panjang cangkang-tinggi cangkang memperlihatkan bahwa pola pertumbuhan A. pilula adalah alometrik negatif (Tabel 6). Hal ini menunjukkan pertumbuhann panjang cangkang lebih dominan dibandingkan tebal cangkang dan tinggi cangkang. Contoh perhitungan tabel sidik ragam regresi panjang cangkang-tebal cangkang ditunjukkan pada Lampiran 4.
Hasil analisis regresi antara panjang cangkang dengan tebal cangkang dan tinggi cangkang A. pilula dari keempat wilayah ditunjukkan pada Gambar 8 Berdasarkan grafik hubungan panjang cangkang-tebal cangkang, terlihat bahwa terdapat titik potong yang mengindikasikan adanya perbedaan slope antar regresi. Hal ini mengindikasikan adanya laju pertumbuhan tebal cangkang yang tidak sama antar populasi yang dikaji. Populasi dengan kemiringan (slope) yang tercuram menunjukkan pertumbuhan relatif tebal cangkang yang paling cepat dibandingkan dengan populasi lainnya.
Tabel 6 Perbandingan hasil regresi panjang cangkang (P) terhadap tebal cangkang (L) dan panjang cangkang (P) terhadap tinggi cangkang kiri (TI)
Lokasi y=a+bx R2 N Alometri ANCOVA
16
(a) (b)
Gambar 8 Analisis regresi karakter morfometrik Anadara pilula (a) regresi panjang tebal cangkang (b) regresi panjang cangkang-tinggi cangkang . Labuan, Cirebon, Rembang,
Gresik
Berbeda dengan regresi panjang cangkang-tebal cangkang, hasil analisis garis regresi panjang cangkang-tinggi cangkang dari keempat lokasi memperlihatkan garis yang cenderung sejajar mengindikasikan tidak adanya perbedaan nilai slope pada keempat regresi. Hal ini menunjukkan adanya laju pertumbuhan tinggi cangkang yang sama antar lokasi.
Uji ANCOVA dilakukan untuk melihat perbedaan laju pertumbuuhan dan ukuran A. pilula dari keempat lokasi. Berdasarkan uji ANCOVA diketahui bahwa terdapat perbedaan slope dan intercept pada beberapa populasi. Perbedaan slope mengindikasikan adanya perbedaan laju pertumbuhan relatif A. pilula, sedangkan perbedaan intercept menunjukkan adanya perbedaan ukuran morfometrik cangkang pada panjang cangkang yang sama. Contoh hasil tabel sidik ragam uji ANCOVA dari regresi panjang cangkang-tebal cangkang disajikan pada Lampiran 6 dan 7.
Analisis klaster
Analisis klaster dibangun berdasarkan matriks jarak dari karakter morfometrik (Tabel 8). Berdasarkan matriks jarak tersebut diketahui bahwa jarak terjauh dimiliki oleh populasi A. pilula yang berasal dari Rembang dan Gresik, sedangkan jarak terdekat dimiliki oleh populasi Labuan dan Rembang.
Konstruksi dendrogram memperlihatkan bahwa populasi A. pilula yang berasal dari Pesisir Labuan membentuk kelompok dengan populasi Rembang, sedangkan populasi A. pilula dari Cirebon membentuk kelompok dengan populasi A. pilula Gresik (Gambar 10).
17 Tabel 7 Matriks jarak berdasarkan karakter morfometrik Anadara pilula antara
keempat wilayah (Labuan, Cirebon, Rembang, Gresik)
Lokasi Labuan Cirebon Rembang Gresik
Labuan 0 1.574 1.205 2.780
Cirebon 1.574 0 1.989 1.561
Rembang 1.205 1.989 0 3.204
Gresik 2.780 1.561 3.204 0
Gambar 9 Dendogram populasi Anadara pilula dari empat wilayah (Labuan, Cirebon, Rembang, Gresik) berdasarkan karakter morfometrik
Analisis diskriminan
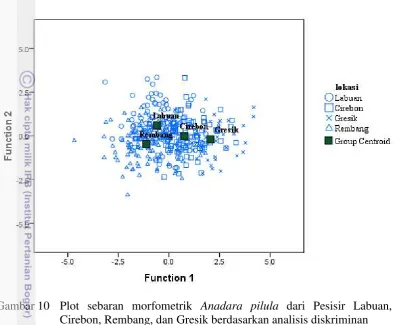
Analisis diskriminan yang dilakukan menunjukkan bahwa keempat populasi tidak terpisah secara sempurna (Gambar 9). Populasi A. pilula dari Pesisir Labuan, Cirebon, Rembang, dan Gresik masing-masing terklasifikasikan secara tepat sebesar 63.0%, 55.7%, 78.3%, dan 64.4% (Tabel 7). Total persentase klasifikasi pada keseluruhan wilayah adalah sebesar 63.2%. Berdasarkan analisis diskriminan diketahui terdapat tujuh variabel yang berpengaruh terhadap pengelompokkan populasi A. pilula, yaitu rasio tebal cangkang (L/P), panjang ligamen (PL/P), tinggi cangkang kanan (TA/P), tinggi cangkang kiri (TI/P), tinggi umbo cangkang kanan (TUA/P), tinggi umbo cangkang kiri (TUI/P), simetri cangkang kanan (SA/P), dan simetri cangkang kiri (SI/P) (Lampiran 10).
Tabel 8 Hasil klasifikasi populasi Anadara pilula di masing-masing populasi berdasarkan analisis diskriminan
Lokasi Kelompok prediksi (%)
Labuan Cirebon Rembang Gresik Total Kelompok
Berdasarkan analisis diskriminan juga diketahui bahwa adanya sharing component dari masing-masing populasi dengan populasi lainnya akibat kesalahan klasifikasi. Adanya sharing component ini diakibatkan oleh kesamaan karakter
18
morfometrik pada individu-individu A. pilula di antara keempat populasi. Hasil analisis menunjukkan adanya sharing component antar populasi, kecuali pada populasi Rembang dan Gresik.
Gambar 10 Plot sebaran morfometrik Anadara pilula dari Pesisir Labuan, Cirebon, Rembang, dan Gresik berdasarkan analisis diskriminan
Analisis peta arus di Laut Jawa
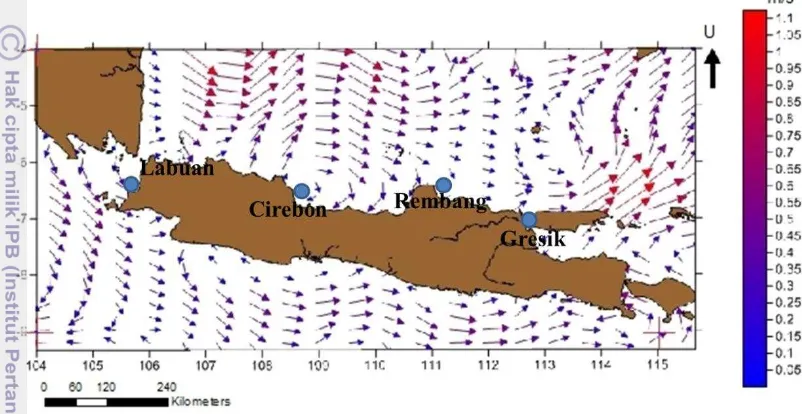
Analisis plot arus di Laut Jawa berdasarkan data NOAA memperlihatkan kecepatan dan arah arus di antara keempat lokasi yang dapat mengindikasikan ada atau tidak adanya konektivitas antar lokasi akibat sebaran larva A. pilula yang terbawa oleh arus seperti yang ditunjukkan pada Gambar 11. Analisis plot arus dibuat berdasarkan data arus pada tahun 2014.
Masa dispersal larva bivalvia jenis cockle pada umumnya memiliki periode sekitar 30 hari (Dare et al. 2004). Pada periode ini, larva kerang yang berada dalam fase meroplankton akan terbawa arus dan tersebar ke wilayah lainnya, sehingga memungkinkan adanya konektivitas antara satu lokasi dengan lokasi yang lain. Setelah melewati stadia meroplankton, bivalvia akan memasuki masa settlement dan pada umumnya menjadi sesil di substrat, sehingga sangat sulit terjadi penyebaran individu antar lokasi pada fase ini.
19 Jarak keempat wilayah (Labuan, Cirebon, Rembang, Gresik) yang cukup jauh (lebih dari 200 km) dengan kecepatan rata-rata arus di Laut Jawa yang relatif lemah (hanya sekitar 0.4 m/s) dan banyaknya barrier lingkungan menyebabkan konektivitas larva A. pilula antar lokasi menjadi sangat sulit. Oleh karena itu, diduga empat populasi A. pilula di masing-masing wilayah (Labuan, Cirebon, Rembang, Gresik) merupakan unit stok yang berbeda.
Gambar 11 Peta sebaran arus tahun 2014 di Laut Jawa
Pembahasan
Keragaman morfometrik A.pilula intra dan interpopulasi
Secara deskriptif keragaman morfometrik intra dan interpopulasi digambarkan melalui nilai koefisien keragaman (KK). Berdasarkan analisis yang telah dilakukan, nilai koefisien keragaman terendah dan tertinggi pada populasi Labuan masing-masing diperoleh dari karakter tinggi cangkang kanan dan tinggi umbo cangkang kanan dengan nilai sebesar 2.85% dan 10.19%. Koefisien keragaman terendah dan tertinggi dari populasi A. pilula di Pesisir Cirebon dan Gresik ditemukan pada karakter tinggi cangkang kiri dan tinggi umbo kanan. Nilai koefisien keragaman untuk tinggi cangkang kiri untuk populasi A. pilula di Labuan dan Gresik berturut-turut adalah 3.01% dan 2.86%, sedangkan nilai koefisien keragaman untuk karakter tinggi umbo kanan di wilayah Labuan dan Gresik, masing-masing adalah 10.78% dan 11.22%. Hal yang sedikit berbeda ditemukan pada populasi A. pilula di Rembang dengan nilai koefisien keragaman terendah ditemukan pada karakter tinggi cangkang kanan dan nilai koefisien keragaman tertinggi ditemukan pada karakter tinggi umbo kiri, dengan nilai masing-masing 2.81% dan 9.73%.
20
memiliki nilai koefisien keragaman rendah menggambarkan konsistensi ukurannya, sehingga diindikasikan mampu menjadi penciri fenotip populasi, meskipun harus dipastikan lebih lanjut untuk melihat ada atau tidak adanya perbedaan interpopulasi yang signifikan terhadap karakter tersebut.
Uji MANOVA satu arah dan uji lanjut Games-Howell yang dilakukan menunjukkan bahwa terdapat enam karakter yang berbeda secara signifikan antar populasi, yaitu rasio panjang ligamen (PL/P), tinggi cangkang kiri (TI/P), tinggi cangkang kanan (TA/P), tinggi umbo kanan (TUA/P), simetri cangkang kiri (SI/P), dan simetri cangkang kanan (SA/P) (p<0.05). Karakter rasio panjang ligamen dan rasio tinggi cangkang kiri A. pilula dari populasi Rembang berbeda secara signifikan dengan tiga populasi lainnya, yaitu rasio panjang ligamen dan rasio tinggi cangkang kiri terhadap panjang cangkang pada populasi A. pilula Rembang lebih kecil dibandingkan populasi lainnya. Hal yang sama juga terlihat pada rasio tinggi cangkang kanan terhadap panjang cangkang (TA/P) yang menunjukkan A. pilula di wilayah Rembang memiliki karakter TA/P yang secara signifikan lebih kecil dibandingkan dengan populasi Gresik (p<0.05), namun tidak berbeda nyata dengan dua populasi lainnya (p>0.05). Selain itu, diketahui pula bahwa rasio karakter simetri cangkang kanan terhadap panjang cangkang memiliki perbedaan secara signifikan antar keempat populasi (p<0.05).
Jika perbedaan karakter interpopulasi yang diuji dengan MANOVA dikaitkan dengan nilai koefisien keragaman intrapopulasi, maka karakter-karakter dengan nilai koefisien keragaman rendah lebih ideal untuk dijadikan penciri fenotip populasi. Karakter tersebut secara berturut-turut untuk populasi Labuan, Cirebon, Rembang, dan Gresik adalah rasio tinggi cangkang kanan (TA/P), rasio tinggi cangkang kiri (TI/P), rasio tinggi cangkang kanan (TA/P), dan rasio tinggi cangkang kiri (TI/P). Meskipun demikian, setelah karakter-karakter tersebut dikaitkan dengan keragaman interpopulasi melalui uji MANOVA dan Games-Howell, diketahui bahwa karakter-karakter tidak dapat menjadi penciri fenotip bagi masing-masing populasi. Hal ini disebabkan karakter tersebut tidak berbeda nyata terhadap tiga populasi lainnya sekaligus, namun hanya berbeda nyata dengan satu atau dua populasi lainnya. Hal ini dapat terlihat salah satunya pada rasio karakter tinggi cangkang kanan (TA/P) pada populasi Rembang yang signifikan lebih kecil dibandingkan populasi Gresik (p<0.05), namun tidak berbeda signifikan terhadap dua populasi lainnya (p>0.05) sehingga penentuan penciri fenotip dari karakter ini menjadi kurang tepat.
Berdasarkan uji lanjut Games-Howell, diketahui pula bahwa rasio karakter simetri cangkang kanan (SA/P) berbeda signifikan pada keempat populasi. Meskipun demikian, karakter ini juga beresiko untuk dijadikan penciri fenotip bagi masing-masing populasi karena kisaran nilai SA/P antar populasi sangat dekat dan terdapat nilai-nilai yang tumpang tindih sehingga akan menjadi kurang tepat pula bila dijadikan sebagai penciri populasi.
21 salinitas (Eisma 1965), periode pasang surut (De Montaudoin 1996), tipe sedimen (Newell dan Hidu 1982), dan lainnya. Laudien et al. (2003) menyatakan bahwa terdapat perbedaan karakter morfometrik pada kerang Donax serra yang hidup di perairan dengan temperatur berbeda. Kerang yang hidup di perairan yang lebih dingin memiliki bentuk cangkang yang lebih pipih dibandingkan dengan kerang yang hidup di perairan yang lebih hangat. Tarnowska et al. (2009) juga menyatakan bahwa salinitas dapat mempengaruhi morfometrik bivalvia yang ditunjukkan oleh populasi Cerastoderma glaucum yang memiliki ukuran lebih kecil ketika hidup di Laut Baltik yang memiliki salinitas di bawah rata-rata.
Hubungan panjang cangkang dan bobot A. pilula
Analisis hubungan panjang dan bobot dilakukan terhadap panjang cangkang dengan bobot total serta panjang cangkang dengan bobot daging. Berdasarkan analisis yang telah dilakukan, diketahui bahwa pola pertumbuhan A. pilula di Pesisir Labuan, Cirebon, dan Rembang berdasarkan hubungan panjang cangkang-bobot total adalah isometrik, sedangkan untuk populasi A. pilula Gresik adalah alometrik negatif. Hal ini menunjukkan bahwa pertumbuhan panjang A. pilula di Pesisir Labuan, Cirebon, Rembang, dan Gresik sebanding dengan pertumbuhan bobotnya, sedangkan pertumbuhan panjang lebih dominan dibandingkan bobot untuk populasi Gresik. Satrioajie et al. (2013), yang melakukan studi di Pesisir Tegal juga menemukan bahwa hubungan panjang dan bobot total A. pilula yang hidup di Pesisir Tegal adalah alometrik negatif. Perbedaan pertumbuhan ini dapat disebakan oleh beberapa faktor eksternal seperti kompetisi intraspesies (Jensen 1993), gangguan parasit (Miura dan Chiba 2007), kepadatan populasi (Ivell 1981), dan periode makan (Jensen 1992). Selain itu, pertumbuhan juga sangat dipengaruhi oleh faktor internal seperti genetik (Meyer dan Manahan 2010).
Pola pertumbuhan alometrik negatif pada populasi A. pilula dapat disebabkan oleh adaptasinya terhadap lingkungan. Menurut Gaspar et al. (2002) pertumbuhan alometrik negatif pada bivalvia dapat menjadi salah satu adaptasinya dalam melakukan strategi penggalian substrat dan menghindari terlepasnya bivalvia dari substrat dasar akibat hidrodinamika lokal.
Analisis hubungan panjang cangkang terhadap bobot daging memperlihatkan bahwa pola pertumbuhan untuk keempat populasi adalah isometrik, yang diartikan sebagai pertumbuhan panjang cangkang sebanding dengan pertumbuhan bobot daging A. pilula. Secara umum, nilai koefisien determinasi (R2) pada hubungan panjang cangkang-bobot total lebih besar dibandingkan panjang cangkang-bobot daging. Hal ini mengindikasikan bahwa pertumbuhan bobot daging lebih bervariasi dibandingkan bobot total, sehingga panjang cangkang lebih mampu menjelaskan variasi bobot total dibandingkan bobot daging.
22
daging dapat disebabkan oleh perbedaan ketersediaan makanan di alam antar kedua lokasi.
Hubungan panjang cangkang dengan karakter morfometrik lainnya pada A. pilula
Hubungan panjang cangkang dengan karakter morfometrik lainnya, yaitu tebal cangkang dan tinggi cangkang kiri memperlihatkan bahwa pola pertumbuhan A. pilula adalah alometrik negatif. Hal ini menunjukkan pertumbuhan panjang cangkang lebih dominan dibandingkan pertumbuhan tebal dan tinggi cangkang. Satrioajie et al. (2013) yang telah melakukan penelitian mengenai morfometrik A. pilula di Pesisir Tegal menemukan bahwa hubungan panjang cangkang dengan tebal cangkang serta hubungan panjang cangkang dengan tinggi cangkang adalah alometrik negatif, dengan nilai slope masing-masing adalah 0.912 dan 0.842. Hasil tersebut tidak berbeda jauh dengan studi ini yang dilakukan pada empat populasi A. pilula yang menunjukkan pertumbuhan panjang cangkang lebih dominan dibandingkan tebal cangkang dan tinggi cangkang.
Berdasarkan uji ANCOVA dari regresi panjang cangkang terhadap tebal cangkang memperlihatkan bahwa populasi A. pilula di Labuan dan Cirebon serta Labuan dan Gresik memiliki pertumbuhan tebal cangkang yang sama (p>0.05). namun populasi A. pilula di Labuan dan Gresik memiliki ukuran tebal cangkang yang berbeda pada panjang cangkang yang sama (p<0.05), sedangkan populasi A. pilula di Labuan dan Cirebon memiliki ukuran tebal cangkang yang sama pada panjang cangkang yang sama.
Hal yang berbeda terjadi salah satunya antara populasi A. pilula di Cirebon dan Gresik yang memiliki pertumbuhan tebal cangkang yang berbeda (p<0.05). Populasi di Gresik cenderung memiliki pertumbuhan tebal cangkang yang lebih lambat dibandingkan populasi Cirebon. Hal ini ditandai oleh nilai slope yang lebih rendah pada populasi A. pilula Gresik. Populasi A. pilula Rembang merupakan populasi dengan laju pertumbuhan tebal cangkang yang paling besar dibandingkan dengan ketiga populasi lainnya (p<0.05).
Uji ANCOVA dari regresi panjang cangkang terhadap tinggi cangkang kiri memperlihatkan bahwa populasi A. pilula di Cirebon dan Labuan memiliki pertumbuhan tinggi cangkang dan ukuran tinggi cangkang yang tidak berbeda (p>0.05). Hal yang berbeda terjadi pada perbandingan populasi lainnya yang memiliki laju pertumbuhan tinggi cangkang yang relatif sama (p>0.05), namun memiliki ukuran tinggi cangkang yang berbeda pada panjang cangkang yang sama (p<0.05). Secara deskriptif, dapat diketahui bahwa pada panjang cangkang yang sama, tinggi cangkang kiri A. pilula dari yang terkecil dan terbesar secara berurutan dimiliki oleh populasi Rembang, Labuan, Cirebon, dan Gresik.
23 populasi A. pilula Cirebon, namun setelah melewati panjang 24.36 mm, populasi A. pilula dari Cirebon akan memiliki tebal cangkang yang lebih besar dibandingkan A. pilula Gresik pada panjang cangkang yang sama. Secara singkat dapat dikatakan bahwa populasi A. pilula Gresik memiliki bentuk lebih membulat daripada populasi A. pilula Cirebon sebelum mencapai panjang cangkang 24.36 mm.
Titik potong antara populasi A. pilula yang berasal dari Pesisir Labuan dan Rembang terjadi pada saaat panjang cangkang mencapai 20.86 mm. Hal ini menunjukkan bahwa tebal cangkang A. pilula di Pesisir Rembang lebih rendah dibandingkan populasi A. pilula di Pesisir Labuan sebelum mencapai panjang cangkang 20.86 mm. Setelah melewati panjang cangkang 20.86 mm, bentuk cangkang A. pilula di Pesisir Rembang akan lebih membulat dibandingkan populasi Labuan akibat pertumbuhan tebal cangkangnya yang cepat. Populasi A. pilula di Pesisir Cirebon dan Rembang juga memiliki laju pertumbuhan tebal cangkang yang berbeda, sehingga titik potong ditemukan pada saat panjang cangkang mencapai 21.21 mm. Sebelum mencapai panjang ini, bentuk cangkang A. pilula di Pesisir Cirebon akan lebih membulat dibandingkan populasi A. pilula yang berasal dari Rembang karena tebal cangkang yang lebih tinggi pada populasi Cirebon. Perbedaan laju pertumbuhan relatif tebal cangkang juga terjadi antara populasi Gresik dan Rembang. Titik potong ini terjadi pada saat panjang cangkang mencapai 23.25 mm. Setelah mencapai panjang tersebut, populasi A. pilula Rembang yang awalnya memiliki tebal cangkang yang lebih rendah dari populasi A. pilula Gresik akan berbalik memiliki ukuran tebal cangkang yang lebih besar dibandingkan populasi Gresik pada panjang cangkang yang sama. Beberapa penelitian sebelumnya membuktikan bahwa perbedaan laju pertumbuhan dan bentuk cangkang pada bivalvia dapat dipengaruhi oleh lamanya periode pasang surut (Griffiths 1981; De Montaudoin 1996), kepadatan populasi yang berhubungan dengan kompetisi intrapopuasi (Jensen 1993), perbedaan salinitas (Tarnowska et al. 2009; Mariani et al. 2002), serta perbedaan tipe sedimen (Newell dan Hidu 1982).
24
Dendrogram dan analisis diskriminan karakter morfometrik
Analisis klaster yang dibangun berdasarkan matriks jarak akar kuadrat jarak Mahalanobis antar populasi menunjukkan jarak terdekat dimiliki oleh populasi Labuan dan Rembang dengan jarak sebesar 1.205 dan jarak terjauh dimiliki oleh populasi Rembang dan Gresik dengan jarak sebesar 3.204. Hasil dendrogram yang terbentuk pun menunjukkan bahwa terbentuk dua kelompok, yaitu kelompok pertama yang terdiri dari populasi Labuan dan Rembang serta kelompok kedua yang terdiri dari populasi Cirebon dan Gresik. Hal ini mengindikasikan bahwa karakter morfometrik populasi A. pilula di Labuan memiliki kemiripan dengan populasi A. pilula Rembang. Hal yang sama juga ditunjukkan oleh populasi A. pilula di Cirebon yang memiliki kemiripan karakter morfometrik dengan populasi A. pilula di Gresik.
Berdasarkan analisis diskriminan yang telah dilakukan diketahui bahwa terdapat empat pusat sebaran (centroid) yang terbentuk untuk masing-masing populasi. Meskipun demikian, berdasarkan karakter morfometriknya individu-individu tidak dapat terpisah dan terklasifikasikan secara sempurna. Hal ini terlihat pada sebaran masing-masing populasi yang masih tumpang tindih. Adanya sebaran yang tumpang tindih menandakan kemiripan karakter morfometrik dari A. pilula antar lokasi. Analisis sharing component memperlihatkan bahwa 63% populasi A. pilula dari Pesisir Labuan terklasifikasi secara tepat, namun sebesar 23% terklasifikasi sebagai populasi Rembang. A. pilula dari populasi Rembang pun hanya terklasifikasi secara tepat sebesar 64.4% dan sebagian populasinya terklasifikasi ke dalam populasi Labuan sebesar 18.3%. Hal yang sama juga ditunjukkan oleh populasi Cirebon dan Gresik yang menunjukkan bahwa 19.8% populasi A. pilula Cirebon salah terklasifikasi sebagai populasi Gresik, dan 17.4% populasi A. pilula Gresik salah terklasifikasi sebagai populasi Cirebon. Analisis diskriminan menunjukkan hasil yang serupa dengan analisis klaster, yaitu terdapat kemiripan karakter morfometrik antara populasi Labuan dan Rembang serta antara populasi Cirebon dan Gresik.
Kemiripan karakter morfometrik A. pilula antar populasi menyebabkan terjadinya sharing component yang cukup besar. Berdasarkan analsis sharing component juga diketahui bahwa tidak terdapat tumpang tindih antara populasi Rembang dan Gresik yang terlihat dari nilai sharing component sebesar 0%. Hal ini mengindikasikan karakter morfometrik A. pilula di Pesisir Rembang berbeda dengan populasi A. pilula di Gresik.
25
Pengelolaan sumber daya A. pilula di pesisir Pulau Jawa
Kajian keragaman morfometrik diperlukan dalam bidang pengelolaan sumber daya perikanan untuk mengetahui adanya penciri fenotip dari suatu stok sumber daya. Penelitian ini belum berhasil menemukan fenotip penciri dari masing-masing populasi A. pilula di Pesisir Labuan, Cirebon, Rembang, dan Gresik. Meskipun demikian, informasi keragaman morfometrik yang dikaitkan dengan plot arus dapat mengindikasikan bahwa keempat populasi merupakan empat unit stok yang berbeda. Berdasarkan plot arus yang dibuat, penyebaran dan konektiitas larva A. pilula antar keempat lokasi sulit terjadi akibat adanya barrier lingkungan dan jarak yang jauh antar lokasi. Menurut De Montaudoin (1996), jika antar populasi tidak dimungkinkan terjadinya pertukaran genotipe, maka keragaman morfometrik cenderung dipengaruhi oleh interaksi faktor genetik dan lingkungan lokal.
26
4
SIMPULAN DAN SARAN
Simpulan
Terdapat enam karakter morfometrik yang memiliki perbedaan signifikan antar keempat lokasi (Pesisir Labuan, Cirebon, Rembang, dan Gresik), yaitu panjang ligamen, tinggi cangkang kanan, tinggi cangkang kiri, tinggi umbo cangkang kanan, simetri cangkang kanan dan simetri cangkang kiri. Belum ditemukan penciri fenotip untuk masing-masing populasi pada penelitian ini. Analisis morfometrik dan plot arus mengindikasikan bahwa populasi A. pilula di Pesisir Labuan, Cirebon, Rembang, dan Gresik adalah unit stok yang berbeda sehingga pengelolaannya harus dipisahkan.
Saran
27
DAFTAR PUSTAKA
Afiati. 2007. Hermaphroditism in Anadara granosa (L.) and Anadara antiquata (L.) (bivalvia: Arcidae) from Central Java. Journal of Coastal Development. 10(3):171-179.
Akhrianti I, Bengen DG, Setyobudiandi I. 2014. Distribusi spasial dan preferensi habitat bivalvia di pesisir perairan Kecamatan Simpang Pesak Kabupaten Belitung Timur. Jurnal Ilmu dan Teknologi Kelautan Tropis. 6(1):171-185. Akmal AT. 2013. Perbedaan jarak antar gigi pada alat tangkap garuk terhadap
hasil tangkapan kerang gelatik (Anadara pilula) di perairan Blanakan, Subang, Jawa Barat [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Benton MJ, Diamond SA, Guttman SI. 1994. A genetic and morphometric comparison of Helisoma trivolvis and Gambusia holbrooki from clean and contaminated habitats. Ecotoxicology and Environmental Safety. 29:20-37. Broom MJ. 1982. Size-selection, consumption rates and growth of the gastropods
Natica maculosa (Lamarck) and Thais carinifera (Lamarck) preying on the bivalve Anadara granosa (L.). Journal of Experimental Marine Biology and Ecology. 56:213-233.
Butet NA. 2013. Plastisitas fenotip kerang darah Anadara granosa L. dalam merespon pencemaran lingkungan: studi kasus di perairan Pesisir Banten. [Disertasi]. Bogor (ID): Institut Pertanian Bogor.
Cadrin SX. 2000. Advances in morphometric identification of fishery stocks. Rev. Fish Biol. Fish. 10:91-112. Cypraeidae). Journal of Molluscan Studies.72:31-38.
Cadrin SX, Friedland KD. 2005. Morphometric Outlines. Di dalam: S. X. Cadrin Friedland KD, Waldmann JR, editor. Stock identification methods: applications in fishery science. London (UK): Elsevier Academic Press. Hlm 173-183.
Cheung MS, Wang WX. 2008. Analyzing biomagnification of metals in different marine food webs using nitrogen isotopes. Marine Pollution Bulletin. 56:2082-2088.
Dalziel B, Boulding EG. 2005. Water-borne cues from a shell-crushing predator induce a more massive shell in experimental populations of an intertidal snail. Journal of Experimental Marine Biology and Ecology. 317:25-35. Dare PJ, Bell MC, Walker P, Bannister RCA. 2004. Historical and current status
of cockle and mussel stocks in The Wash. The Wash (UK): CEFAS Lowesto. 85hlm.
De Montaudouin X. 1996. Factors involved in growth plasticity of cockles Cerastoderma edule (L.), identified by field survey and transplant experiments. Journal of Sea Research. 36:251-265.
Dolorosa RG, Dangan-Galon F. 2014. Species richness of bivalves and gastropods in Iwahig River-Estuary, Palawan, the Philippines. International Journal of Fisheries and Aquatic Studies. 2:207-215.
Eisma D. 1965. Shell characteristics of Cardium edule L. as indicators of salinity. Netherlands Journal of Sea Research 2:493-540.
28
Faulkner P. 2010. Morphometric and taphonomic analysis of granular ark (Anadara granosa) dominated shell deposits of Blue Mud Bay, northern Australia. Journal of Archaeological Science. 37:1942-1952.
Feng Y, Li Q, Kong L, Zheng X. 2011. COI-based DNA barcoding of Arcoida species (Bivalvia: Pteriomorphia) along the coast of China. Molecular Ecology Resources. 11:435-441.
Filgueira R, Labarta U, Fernández-Reiriz M J. 2008. Effect of condition index on allometric relationships of clearance rate in Mytilus galloprovincialis Lamarck, 1819. Revista de Biología Marina y Oceanografía. 43(2): 391-398. Gaspar B, Miguel N, Miguel S, Paulo, Carlos CM. 2002. Shell morphometric
relationship of the most common bivalve species (Mollusca:Bivalvia) of the Algarve coast (southern Portugal). Hydrobiologia. 477:73-80.
Griffiths RJ. 1981. Population dynamics and growth of the bivalve Choromytilus meridionalis (Kr.) at different tidal levels. Estuarine, Coastal and Shelf Science. 12:101-118.
Hossen MF, Hamdan S, Rahman MR. 2014. Cadmium and lead in blood cockle (Anadara granosa) from Asajaya, Sarawak, Malaysia. The Scientific World Journal. 2014: 1-4.
Hutomo M, Moosa MK. 2005. Indonesian marine and coastal biodiversity: present status. Indian Journal of Marine Sciences. 34(1):88-97.
Irie T. 2006. Geographical variation of shell morphology in Cypraea annulus (Gastropoda: Cypraeidae). Journal of Molluscan Studies. 72:31-38.
Ivell R. 1981. A quantitative study of a Cerastoderma-Nephthys community in the Limfjord, Denmark, with special reference to production of Cerastoderma edule. Journal of Molluscan Studies. 47:147-170.
Jahangir S, Siddiqui G, Ayub Z. 2014. Temporal variation in the reproductive pattern of blood cockle Anadara antiquata from Pakistan (northern Arabian Sea).Turkish Journal of Zoology. 38:263-272.
Jensen KT. 1992. Dynamics and growth of the cockle, Cerastoderma edule, on an inter-tidal mudflat in the Danish Wadden Sea: effects of submersion time and density. Netherlands Journal of Sea Research. 28:335-345.
Jensen KT. 1993. Density-dependent growth in cockles (Cerastoderma edule): evidence from interannual comparisons. Journal of the Marine Biological Association of the United Kingdom.73:333-342.
Kawecki TJ, Ebert D. 2004. Conceptual issues in local adaptation. Ecological Letters. 7:1225-1241.
Krakau M. 2008. Biogeographic patterns of the marine bivalve Cerastoderma edule along European Atlantic coasts. [Disertasi]. Kiel (DE): University of Kiel.
Kumar SK, Tamura, Nei M. 1993. MEGA. Molecular Evolutionary Genetics Analysis. Version 3.0 [Internet]. Pennsylvania (US). [diunduh pada Agustus 2015]. Tersedia pada: http://www.megasoftware.net/MEGA-v1.01.pdf Laudien J, Flint NS, Van der Bank FH, Brey T. 2003. Genetic and morphological
variation in four populations of the surf clam Donax serra (Roding) from southern African sandy beaches. Biochemical Systematics and Ecology. 31:751-772.
29 Li B, Morrison RJ. 2011. Analysis of macrobenthic assemblages of Lake Illawarra,
New South Wales, Australia. Transitional Waters Bulletin. 5:85-94.
Lozouet P, Plaziat JC. 2008. Mangrove Environments and Molluscs: Abatan River, Bohol and Panglao Islands, Central Philippines. Hackenheim (DE): ConchBooks.
Lutaenko KA. 2011. Status of The Knowledge of The Indo-Pacific Anadarinae (Mollusca: Bivalvia). Di dalam: Lutaenko, editor. Proceedings of the Workshop Coastal Marine Biodiversity and Bioresources of Vietnam And Adjacent Areas to The South China Sea; 2011 November 24-25; Nha Trang, Vietnam (VN): Asia-Pacific Network for Global Change Research. hlm 95-101.
Mariani S, Piccari F, De Matthaeis E. 2002 Shell morphometry in Cerastoderma spp.(Bivalvia: Cardiidae) and its significance for adaptation to tidal and non-tidal coastal habitats. Journal of the Marine Biological Association of the United Kingdom 82:483-490.
Meyer E, Manahan DT. 2010. Gene expression profiling of genetically determined growth variation in bivalve larvae (Crassostrea gigas). Journal of Experimental Biology. 213:749-758.
Miura O, Chiba S. 2007. Effects of trematode double infection on the shell size and distribution of snail hosts. Parasitology International. 56:19-22.
Mudjiono, Kastoro WW. 1997. Density, biomass and distribution of cockle, Potiarca pilula (Reeve, 1844) in The Bay of Miskam, West Java. Indonesia. Phuket Marine Biological Center Special Publication. 17(1):193-197. Mzighani S. 2005. Fecundity and population structure of cockles, Anadara
antiquata L. 1758 (Bivalvia: Arcidae) from a sandy/muddy beach near Dar es Salaam, Tanzania. Western Indian Ocean Journal Marine Science. 4(1):77–84.
Newell CR, Hidu H. 1982. The effects of sediment type on growth rate and shell allometry in the soft shelled clam Mya arenaria L. Journal of Experimental Marine Biology and Ecology.65:285-295.
[NOAA] National Oceanic and Atmospheric Administration. 2015. Ocean surface current analysis-real time. [diacu 18 Agustus 2015]. Tersedia dari: http://www.oscar.noaa.gov/.
Okera W. 1976. Observations on some population parameters of exploited stocks of Senilia senilis (= Arca senilis) in Sierra Leone. Marine Biologi. 38:217-229.
Pigllucci M. 1996. How organisms respond to environmental changes: from phenotypes to molecules (and vice versa). Trends in Ecology and Evolution 11:168-173.
Poutiers JM. 1998. Bivalves (Acephala, Lamellibranchia, Pelecypoda). Di dalam: Carpenter KE dan Niem V H. FAO Species Identification Guide for Fishery Purposes. The Living Marine Resources of The Western Central Pacific. Volume 1. Seaweeds, Corals, Bivalves, and Gastropods. Rome, Italia. Rome (IT): FAO. hlm 123-362.