KERAGAMAN FENOTIPIK DAN GENOTIPIK
KERANG BULU (Genus:
Anadara
)
DI PESISIR UTARA PULAU JAWA
DEWI FITRIAWATI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul Keragaman Fenotipik dan Genotipik Kerang Bulu (Genus: Anadara) di Pesisir Utara Pulau Jawa adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
RINGKASAN
DEWI FITRIAWATI. Keragaman Fenotipik dan Genotipik Kerang Bulu (Genus: Anadara) di Pesisir Utara Pulau Jawa. Dibimbing oleh YUSLI WARDIATNO dan NURLISA A. BUTET.
Kerang bulu termasuk dalam golongan dari famili Arcidae dan merupakan cryptic species karena morfologinya yang sulit dibedakan. Kerang bulu memiliki karakteristik unik dengan cangkang yang diselimuti bulu. Distribusi dari kerang bulu sangat luas meliputi wilayah Indo-Pasifik, termasuk didalamnya perairan Indonesia dan tersebar di kawasan pesisir, intertidal, daerah berbatu, berpasir, berlumpur, bahkan sering dijumpai bersimbiosis dengan biota lain. Penelitian ini bertujuan untuk mengkaji keanekaragaman kerang bulu berdasarkan karakter fenotip (morfologi) dan molekuler (genetik) yang berasal dari enam lokasi di pesisir utara Pulau Jawa.
Sampel kerang bulu yang diukur sebanyak 316 individu. Kerang bulu diambil dari enam wilayah yang berbeda di beberapa pesisir di utara Pulau Jawa, yaitu Pesisir Banten (92 individu), Subang (49 individu), Cirebon (13 individu), Rembang (25 individu), Gresik (22 individu), dan Probolinggo (115 individu). Karakteristik morfometrik kerang bulu yang diukur adalah panjang cangkang (PC), lebar cangkang (LC), tebal cangkang (TC), panjang ligamen (PL), tinggi umbo (TU), simetri kanan (Ska), dan simetri kiri (Ski). Karakter meristik yang dihitung dari kerang adalah jumlah alur (JA). Karakter morfometrik kerang diukur dengan menggunakan kaliper digital dengan ketelitian 0,01 mm. Analisis data yang dilakukan meliputi analisis pohon fenetik, uji Mann-Whitney, analisis diskriminan dan analisis kluster dilakukan untuk membandingkan dan melihat keanekaragaman karakter morfologi antarlokasi. Variasi genetik dilakukan dengan menggunakan analisis molekuler, meliputi isolasi dan ekstraksi DNA, uji kualitas DNA, amplifikasi DNA, dan sekuensing DNA.
Hasil identifikasi secara morfologi dan molekuler menunjukkan bahwa terdapat 3 spesies kerang bulu yang teridentifikasi, yaitu Anadara inaequivalvis (Banten, Subang, Cirebon, Rembang, dan Gresik), A. cornea (Probolinggo), dan A.gubernaculum (Subang). Tiga spesies kerang bulu tersebut memiliki ciri fenotip yang berbeda, berupa bentuk dan ukuran tubuh kerang, posisi umbo, warna cangkang, serta jumlah alur. Pohon fenetik yang dibentuk membuktikan bahwa terdapat tiga spesies kerang bulu dengan ciri fenotip yang berbeda dari enam wilayah di pesisir utara Pulau Jawa. Hasil analisis diskriminan, analisis kluster, dan pohon fenetik menunjukkan hasil yang sama, yaitu populasi dari Banten, Subang (Ai), Cirebon, Rembang, dan Gresik membentuk kelompok pertama, sedangkan populasi yang berasal dari Subang (Ag) dan populasi yang berasal dari Probolinggo membentuk kelompok masing-masing. A. cornea dan A. gubernaculum berhasil diidentifikasi dengan menggunakan marka gen COI. A. inaequivalvis belum berhasil diidentifikasi dengan menggunakan marka gen yang sama. Hasil analisis dari karakter fenotip dan genotip yang diperoleh beberapa karakter penciri dari tiap spesies dapat digunakan sebagai informasi awal dalam menentukan pengelolaan sumberdaya kekerangan.
SUMMARY
DEWI FITRIAWATI. Phenotipic and Genotipic Variation of Hairy Ark Cockle (Genus: Anadara) in Northcoast of Java Island. Supervised by YUSLI WARDIATNO and NURLISA ALIAS BUTET.
Hairy ark cockle is one of the bivalves of family Arcidae. Most of them are determined as cryptic species, due to high similarity on morphology among those species of hairy ark cockles leading to difficulty on identification. Hairy ark cockles have unique characteristic, their shells are covered by black fine hairs. Distribution of cockles is wide covering Indo-Pacific region, including Indonesian waters. They inhabit coastal areas of intertidal, rocky areas, sandy, muddy, and often encounter in symbiosis with other organisms. This study aims at assessing the diversity of phenotype (morphology) and genotype (molecular) characters of the hairy ark cockle from six locations in northcoast of Java Island.
Hairy ark cockle samples were measured as much as 316 individuals. Hairy ark cockles taken from six different regions in some coastal areas in the north of Java, i.e., Banten (92 ind), Subang (49 ind), Cirebon (13 ind), Rembang (25 ind), Gresik (22 ind), and Probolinggo (115 ind). Morphometric characters were measured including shell length (PC), shell width (LC), shell height (TC), ligament length (PL), umbonal height (TU), right symmetry of shell (Ska), and left symmetry of shell (Ski). The number of ribs (JA) as meristic character was counted. Morphometric characters were measured using digital caliper with accuracy 0,01 mm. The data were analyzed using phenetic-tree method, Mann-Whitney test, discriminant and cluster analyses to compare the diversity of morphological characters among locations. Genotypic variation was performed by molecular analysis covering molecular analysis DNA isolation, DNA quality test, amplification of DNA fragments, and DNA sequencing.
Morphology and molecular identification showed that three species of hairy ark cockles, i.e., Anadara inaequivalvis (Bantam, Subang, Cirebon, Rembang, and Gresik), A. cornea (Probolinggo), and A. gubernaculum (Subang). Those species had a different phenotype characteristic, such as shell shape and size, umbonal position, shell color, as well as ribs number. Phenetic tree proved that there are three species of hairy ark cockles with different phenotype characteristic of the six regions in the northcoast of Java Island. Discriminant analysis and cluster analysis showed similar results, i.e., the population of Banten, Subang (Ai), Cirebon, Rembang, and Gresik occupied the first group. While the population from Subang (Ag) and population from Probolinggo occupied different group each. A. cornea and A. gubernaculum were identified successfully using COI gene marker. On the other hand, A. inaequivalvis has not been successfully identified using similar genetic marker. This result can be generally used as initial information to support management of bivalve resource.
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Pengelolaan Sumber Daya Perairan
KERAGAMAN FENOTIPIK DAN GENOTIPIK
KERANG BULU (Genus:
Anadara
)
DI PESISIR UTARA PULAU JAWA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2016
PRAKATA
Puji syukur Penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala limpahan rahmat, hidayah, serta karunia-Nya, sehingga karya ilmiah berjudul Keragaman Fenotipik dan Genotipik Kerang Bulu (Genus: Anadara) di Pesisir Utara Pulau Jawa dapat diselesaikan.
Pada kesempatan ini Penulis menyampaikan terima kasih kepada:
1. Institut Pertanian Bogor yang telah memberikan kesempatan untuk menempuh studi di sekolah pascasarjana pada program studi Pengelolaan Sumber Daya Perairan.
2. Pusat Penelitian Lingkungan Hidup (PPLH) IPB yang telah bersedia mendanai sebagian dari penelitian ini.
3. Dr Ir Yusli Wardiatno, MSc dan Dr Ir Nurlisa A. Butet, MSc selaku komisi pembimbing yang telah memberikan arahan dan masukan dalam penulisan karya ilmiah ini.
4. Dr Ir Isdradjad Setyobudiandi, MSc selaku penguji tamu dan Dr Ir Sigid Hariyadi, MSc selaku perwakilan komisi pendidikan Program Studi Pengelolaan Sumberdaya Perairan atas saran dan masukan yang sangat berharga.
5. Keluarga tercinta, Ayah Sukarman, Ibu Sutitah, Mas Ari, Mba Femi, Ashila serta Miftah, Ayu, dan Lia yang selalu memberikan doa dan motivasi. 6. Tim Laboratorium Biologi Molekuler (Panji, Wahyu, Findra, Lela, Fajrin,
Yustin, Lita) dan Tim Laboratorium Biologi Makro (Pak Ruslan, Bang Aries, Bang Zahid, Teh Tina, Teh Dewi, Reiza, Widi, Septyan, Ano) atas semangat dan dukungannya.
7. Teman-teman seperjuangan (Lufi, Yuyun, Agus, Mega, Ayu, Lusita, Febi, Nina, Siska, Akrom, Nisa, Wida) atas dukungan yang diberikan.
Semoga karya ilmiah ini bermanfaat.
DAFTAR ISI
DAFTAR ISI v
DAFTAR TABEL vi
DAFTAR GAMBAR vi
1 PENDAHULUAN 1
Latar Belakang 1
Perumusan Masalah 2
Tujuan Penelitian 3
Manfaat Penelitian 3
2 METODE 4
Lokasi dan Waktu Penelitian 4
Pengambilan Contoh 4
Bahan 5
Alat dan Bahan Analisis Molekuler 6
Analisis Contoh 6
Analisis Data 8
3 HASIL DAN PEMBAHASAN 9
Hasil 9
Pembahasan 21
4 KESIMPULAN DAN SARAN 29
Kesimpulan 29
Saran 29
DAFTAR PUSTAKA 30
LAMPIRAN 35
DAFTAR TABEL
1 Koordinat dan lokasi sampling 5
2 Karakter morfometrik yang diukur 7
3 Perbandingan karakter antara A. inaequivalvis, A. gubernaculum, A.
cornea di beberapa pesisir utara Jawa berdasarkan pengamatan 11 4 Ukuran tubuh genus Anadara dari beberapa pesisir utara Jawa (mm) 11 5 Perbandingan karakter morfometrik genus Anadara dari beberapa
perairan di utara Pulau Jawa (mm) 13
6 Perbandingan hasil regresi panjang cangkang terhadap tebal cangkang
spesies dominan A. inaequivalvis 14
7 Perbandingan hasil regresi panjang cangkang terhadap tebal cangkang
spesies A. Inaequivalvis dan A. gubernaculum 14
8 Perbandingan hasil regresi panjang cangkang terhadap tebal cangkang
spesies A. inaequivalvis dan A. cornea 15
9 Hasil klasifikasi dari tujuh populasi kerang bulu berdasarkan analisis
diskriminan 17
10 Hasil klasifikasi dari populasi dominan A. inaequivalvis dari lima
populasi berdasarkan analisis diskriminan 17
11 Hasil BLASTn gen COI pada AQP dan AQS.1 19
12 Matriks jarak genetik spesies A. inaequivalvis, A. cornea dan famili
Arcidae berdasarkan metode pairwise distance 20
13 Kriteria pencemaran berdasarkan ukuran morfologi 28
DAFTAR GAMBAR
1 Diagram perumusan masalah 3
2 Peta lokasi pengambilan contoh kerang bulu di pesisir utara Pulau Jawa 4 3 Beberapa jenis kerang bulu yang diambil dari Pesisir Banten, Subang,
Cirebon, Rembang, Gresik, dan Probolinggo 5
4 Karakter morfometrik kerang bulu yang diukur dalam penelitian 6 5 Kerang bulu yang ditemukan di enam pesisir utara Pulau Jawa 10 6 Konstruksi pohon fenetik dari tujuh populasi berbeda berdasarkan
karakter fenotip 10
7 Distribusi panjang cangkang kerang bulu yang ditemukan di pesisir utara
Pulau Jawa 12
8 Distribusi panjang cangkang kerang bulu spesies A. inaequivalvis yang
ditemukan di lima lokasi penelitian 12
9 Plot sebaran morfometrik kerang bulu dari tujuh populasi berdasarkan
analisis diskriminan 16
10 Plot sebaran morfometrik A. inaequivalvis dari lima populasi berdasarkan
analisis diskriminan 16
11 Analisis kluster kerang bulu dari tujuh populasi berdasarkan karakter
morfometrik 17
12 Analisis kluster spesies dominan A. inaequivalvis dari lima populasi
13 Pengujian hasil isolasi DNA total pada gel agarosa 1,2% 18 14 Elektroforesis DNA hasil pre-test produk PCR pada gel agarosa 1,2% 19 15 Pohon filogeni berdasarkan gen COI dari famili Arcidae 21 16 Penampakan luar, dalam, dorsal, dan ventral dari kerang A. inaequivalvis,
A. gubernaculum dan A. cornea 22
17 Sebaran kerang bulu di pesisir Banten, Subang, Cirebon, Rembang,
Gresik, dan Probolinggo 24
DAFTAR LAMPIRAN
1 Kaliper digital dengan ketelitian 0,01 mm 36 2 Skoring karakter fenotip dari tiga spesies kerang bulu di enam lokasi
penelitian 36
3 Klasifikasi dari tiga spesies kerang bulu A. inaequivalvis, A. gubernaculum, dan A. cornea berdasarkan literatur 37
4 Output hasil diskriminan 37
5 Contoh hasil uji Mann-Whitney antara kerang bulu dari pesisir Banten
1
PENDAHULUAN
Latar Belakang
Kelas Bivalvia memiliki penyebaran sekitar 15.000 spesies yang hidup di berbagai substrat dan kedalaman perairan laut serta sisanya di air tawar (Pechenik 2005). Salah satu famili dari kelas Bivalvia yang terdistribusi dengan luas yaitu famili Arcidae dan subfamili Anadarinae sebanyak 38 spesies (Lutaenko 2011). Berdasarkan database moluska di Indopasific (OBIS 2005), biodiversitas famili Arcidae mencapai 300 spesies di lautan. Sekitar 180 spesies dari 30 genera berada di perairan Indopasifik, termasuk di dalamnya adalah perairan Indonesia. Hal tersebut menyebabkan tingginya biodiversitas bivalvia di Indonesia. Biodiversitas yang dimaksud bukan hanya terkait pada jumlah atau banyaknya spesies, tetapi juga termasuk keragaman bentuk maupun ukuran dari spesies, keragaman habitat, jaring-jaring makanan hingga tingkat trofik dari suatu spesies (Hendrickx et al. 2007).
Menurut FAO (1998), family Arcidae memiliki distribusi yang luas meliputi wilayah New Caledonia, Australia, Indopasifik, Laut Merah, Jepang, China, Laut Cina Selatan, Hong Kong, Vietnam, Thailand, Filipina, hingga bagian pesisir Indonesia. Famili Arcidae banyak dijumpai di perairan pesisir, Karena merupakan daerah yang potensial sebagai penghasil sumber daya kekerangan. Kerang bulu memiliki distribusi luas di kawasan pesisir di daerah sublitoral, sedimen berpasir (Afiati 2007), daerah estuari bersubstrat lumpur (Pattikawa 2007). Perairan pesisir baik untuk perkembangan organisme dengan substrat yang berlumpur. Bahkan kawasan Banten terkenal akan sumber daya kekerangan atau bivalvia yang melimpah di Indonesia (Komala 2011). Kerang ini sering dijumpai di wilayah pesisir di beberapa daerah di Indonesia, contohnya Banten, Subang, Cirebon, Rembang, Gresik, dan Probolinggo.
Salah satu famili Arcidae yang memiliki karakteristik yang unik adalah kerang bulu. Kerang bulu tersebut memiliki cangkang yang diselimuti oleh bulu-bulu halus. Menurut Dixon et al. (1995), bulu tersebut berasal dari periostrakum yang merupakan lapisan terluar dari cangkang. Bulu tersebut berfungsi sebagai alat perlindungan diri dari ancaman predator dan lingkungan dikarenakan struktur cangkang yang relatif tipis. Selain itu, bulu berfungsi sebagai alat penempel pada substrat. Umumnya famili Arcidae memiliki peran yang sama yaitu sebagai deposit dan filter feeder (Mudjiono dan Kastoro 1997). Syafril et al. (2004) melaporkan bahwa kandungan proksimat dari kerang bulu (Anadara inaequivalvis) tergolong tinggi dengan kandungan protein berkisar 13,22%, kandungan lemak berkisar 2,90%, dan kandungan karbohidrat berkisar 3,77%. Berdasarkan pernyataan nelayan dan masyarakat pesisir, diketahui bahwa kerang bulu dapat berfungsi sebagai makanan penambah darah. Hal tersebut menyebabkan kerang bulu memiliki peran ekonomis yang penting. Sebagai komoditas penunjang ekonomi, kerang ini diperdagangkan baik lokal hingga kegiatan ekspor ke mancanegara.
2
mengenai kerang bulu dilakukan oleh Ambarwati dan Trijoko (2011) mengenai kekayaan jenis Anadara (Bivalvia: Arcidae) di perairan Pantai Sidoarjo. Penelitian tersebut hanya menemukan dua jenis kerang bulu, yaitu A. rufuscens dan A. gubernaculum. Penelitian mengenai keragaman A. pilula juga pernah dilakukan oleh Qonita (2015) di beberapa pesisir di Pulau Jawa, distribusi spasial dan preferensi habitat bivalvia di Belitung Timur oleh Akhrianti (2014). Beberapa penelitian tersebut membuktikan bahwa masih kurangnya data dan informasi bagi kerang bulu di Indonesia.
Kajian molekuler sangat dibutuhkan dalam mengidentifikasi keakuratan suatu spesies dan mendukung hasil identifikasi secara morfologi. Kajian molekuler diketahui memiliki sedikit kesalahan dalam mengidentifikasi. DNA mitokondria yang memiliki sifat conserve secara evolusi, sehingga keragaman genetik populasi kerang bulu dapat dideteksi menggunakan marka mitokondria. Gen Cythocrome Oxsidase subunit I (COI) pada DNA mitokondria dapat dijadikan sebagai marka molekuler untuk penentuan spesies kerang bulu, karena gen COI pada mitokondria merupakan gen yang berevolusi sangat lambat (Solihin 1994). Teknik ini bertujuan untuk mengetahui sumber informasi genetik kerang bulu serta mengetahui data base mengenai situs spesifik yang menggambarkan perbedaan dan distribusi keragaman genetik intra- dan inter populasi kerang bulu, sehingga dapat mempermudah upaya pengelolaan dan konservasi sumber daya kerang bulu di perairan pesisir Indonesia berdasarkan karakter fenotip dan genotipnya.
Perumusan Masalah
Kerang bulu banyak dimanfaatkan karena harganya yang murah sehingga dapat dijadikan sebagai bahan substitusi ketika harga kerang ekonomis lainnya tinggi. Kerang bulu tersebar luas pada substrat berlumpur dan berpasir di pesisir Indonesia. Beberapa diantaranya ditemukan di pesisir utara Pulau Jawa, yaitu pesisir Banten, Subang, Cirebon, Rembang, Gresik, dan Probolinggo. Kerang bulu memiliki cangkang yang ditutupi oleh bulu dan memiliki bentuk yang yang mirip sehingga sulit untuk dibedakan secara morfologi, oleh karena itu kerang bulu tergolong dalam cryptic species. Kondisi tersebut dapat menyebabkan terjadinya kesalahan dalam identifikasi yang dapat memberikan dampak kesalahan terhadap pengelolaan sumber daya.
3 Saat ini kajian mengenai aspek biologi terutama keragaman fenotip dan genotip dari kerang bulu masih sangat minim dilakukan di Indonesia. Kajian ini perlu karena akan menghasilkan informasi awal sumber daya kerang kerang bulu di beberapa pesisir Indonesia. Data keragaman genetik dan keragaman fenotip kerang bulu merupakan salah satu informasi penting dalam penentuan upaya pengelolaan dan konservasi kerang bulu di Indonesia.
Gambar 1 Diagram perumusan masalah
Tujuan Penelitian
Penelitian ini bertujuan untuk 1) mengkaji karakter fenotip yaitu karakter morfometrik (panjang cangkang, lebar cangkang, tebal cangkang, panjang ligamen, tinggi umbo, simetri kanan kerang dan simetri kiri kerang serta karakter meristik (jumlah alur) dari kerang bulu yang berasal dari Pesisir Banten, Subang, Cirebon, Rembang, Gresik, dan Probolinggo, dan 2) mengidentifikasi kerang bulu berdasarkan marka gen COI sebagai informasi dalam menentukan strategi pengelolaan sumber daya.
Manfaat Penelitian
Manfaat dari penelitian ini adalah memberikan pengembangan ilmu pengetahuan berupa informasi keragaman fenotipik dan genotipik dari kerang bulu (Genus: Anadara) di Pesisir Banten, Subang, Cirebon, Rembang, Gresik,
Perbedaan habitat: Banten, Subang, Cirebon, Rembang, Gresik, dan
Probolinggo
Karakter cangkang unik (Cryptic species)
Poor taxonomic knowledge
Populasi kerang bulu
Kajian fenotipik kerang bulu (morfometrik dan meristik)
Kajian genotipik kerang bulu (gen COI)
4
dan Probolinggo, sebagai salah satu aspek biologi yang menunjang strategi pengelolaan sumber daya kerang bulu di Indonesia, khususnya Pulau Jawa.
2
METODE
Waktu dan Lokasi Penelitian
Pengambilan contoh dan penelitian dilaksanakan pada bulan Maret 2015 hingga bulan Juli 2016. Kegiatan analisis morfologi dilakukan di Laboratorium Biologi Makro dan kegiatan analisis molekuler dilakukan di Laboratorium Biologi Molekuler Akuatik, Departemen Manajemen Sumberdaya Perairan, Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor.
Pengambilan Contoh
Contoh kerang bulu diambil dari enam pesisir di Pulau Jawa mewakili bagian daerah di Pulau Jawa, yaitu pesisir Banten dan Subang mewakili Jawa Barat, Cirebon dan Rembang mewakili Jawa Tengah, serta Gresik dan Probolinggo mewakili Jawa Timur (Gambar 3). Pengambilan contoh kerang bulu dilakukan dengan cara purposive sampling. Seluruh contoh kerang bulu yang diambil langsung dari nelayan yang berada di lokasi penelitian, langsung dipreservasi dengan alkohol 96%. Contoh kerang yang diambil disesuaikan dengan kondisi yang ada di lapangan, dan hanya dilakukan dalam satu kali pengambilan contoh tanpa ulangan. Kerang yang diambil dari nelayan dibawa ke laboratorium untuk diidentifikasi dan dianalisis karakter morfometriknya.
5 Tabel 1 Koordinat dan lokasi sampling
Lokasi pengambilan contoh Koordinat N
Karangantu, Banten Mayangan, Subang Mundu, Cirebon Kaliori, Rembang Sidayu, Gresik
6º01’22,08”S-106º09’37,44”E 6º13’01,92”S-107º46’49,44”E 6º45’00,00”S-108º35’51,36”E 6º41’06,72”S-111º16’07,68”E 6º58’49,44”S-112º36’41,76”E
92 49 13 25 22 Mayangan, Probolinggo 7º44’06,00”S-113º14’06,00”E 115
Bahan
Bahan yang digunakan dalam penelitian ini adalah kerang bulu (Gambar 3) yang diambil dari enam wilayah berbeda di sepanjang pesisir utara Pulau Jawa (Gambar 2, Tabel 1). Jumlah contoh kerang bulu digunakan dalam penelitian ini adalah sebanyak 316 individu yang berasal dari enam pesisir, yaitu Pesisir Banten, Pesisir Subang, Pesisir Cirebon, Pesisir Rembang, Pesisir Gresik, dan Pesisir Probolinggo.
6
Alat dan Bahan Analisis Molekuler
Alat yang digunakan dalam penelitian ini diantaranya sarung tangan, masker, rak, baki, gunting, timbangan, tube, kertas saring, pipet mikro, vortex, heating, pinset, stirer, spatula, cetakan agarose, mesin elektroforesis, UV transluminator, mesin PCR Thermal cycler, mesin sentrifuge. Bahan yang digunakan adalah jaringan otot kaki dari kerang bulu, kit isolasi DNA Gene aid, kita PCR Kappa 2G Extra Hotstart, agarose, aquades, etanol absolut, dan buffer TEA.
Analisis Contoh
Analisis Morfologi
Identifikasi spesies secara morfologi dan analisis fenotip
Identifikasi spesies dilakukan untuk menentukan spesies contoh kerang berdasarkan karakteristik fenotip (morfometrik dan meristik). Identifikasi dilakukan berdasarkan buku identifikasi FAO untuk kelompok bivalvia (Poutiers 1998) dan hasil penelitian Vongpanich di Thailand (1996).
Analisis morfometrik
Karakter morfometrik kerang bulu yang diukur adalah panjang cangkang (PC), lebar cangkang (LC), tebal cangkang (TC), tinggi umbo (TU), simetri kanan (Ska), dan simetri kiri (Ski) (Butet 2013), panjang ligamen (PL) (Faulkner 2010) (Gambar 4), serta karakter meristik yang dihitung adalah jumlah alur. Karakter morfometrik diukur dengan menggunakan kaliper digital (Lampiran 1) dengan ketelitian 0,01 mm.
Gambar 4 Karakter morfometrik kerang bulu yang diukur dalam penelitian TU
TC
PC SK
A
SKI
PL
7 Tabel 2 Karakter morfometrik yang diukur
No Keterangan Karakter morfometrik
1
Tinggi cangkang dikurangi dengan tinggi dari ventral sampai ligamen
Jarak antara bagian tengah ligamen sampai dengan bagian terjauh cangkang sebelah kanan Jarak antara bagian tengah ligamen sampai dengan bagian terjauh cangkang sebelah kiri
Analisis Molekuler
Isolasi dan ekstraksi DNA
Otot kaki diambil dari tiga contoh kerang bulu yang masih segar dengan bobot 30 mg. Kemudian dicuci untuk menghilangkan darah dan kotoran serta mencegah kontaminasi. Contoh otot tersebut kemudian dikeringkan dan dimasukkan ke dalam tube untuk dilakukan proses isolasi dan ekstraksi DNA. Isolasi dan ekstraksi DNA dilakukan menggunakan kit komersil (Gene Aid) berdasarkan manual kit tersebut dengan beberapa modifikasi.
Uji kualitas DNA
Uji kualitas DNA dilakukan untuk mengetahui kualitas DNA yang baik. Metode yang digunakan adalah metode elektroforesis. Hasil isolasi dan ekstrasi DNA yang telah diuji dan memiliki kualitas yang baik diambil sebanyak 2,5 μl. Kualitas DNA diuji pada gel agarosa 1,2% yang diwarnai dengan ethidium bromide (EtBr) dan direndam dalam larutan buffer TAE 1x. Visualisasi DNA total dilakukan dengan menggunakan mesin ultraviolet (UV) selama 30 menit.
Amplifikasi dan visualisasi fragmen DNA Gen COI
DNA yang memiliki kualitas yang baik layak dijadikan sebagai cetakan (template) untuk amplifikasi fragmen DNA gen COI dengan teknik PCR (Polymerase Chain Reaction) menggunakan kit komersial Kappa 2G Extra Hotstart dan disesuaikan prosedur manual kit tersebut. Primer yang digunakan adalah primer universal COI, LCO1490 dan HC02198 (Folmer et al. 1994) serta primer Fish F1 dan Fish F2 yang didesain oleh Ward et al. (2005).
LCO1490 5’GGTCAACAAATCATAAAGATATTGG3’
HC02198 5’TAAACTTCAGGGTGACCAAAAAATCA3’
Fish F1 5’TCAACCAACCACAAAGACATTGGCAC3’
Fish F2 5’TCGACTAATCATAAAGATATCGGCAC3’
8
72ºC selama 1 menit, pascaelongasi pada suhu 72ºC selama 5 menit, dan penyimpanan pada suhu 15ºC selama 10 menit. Produk PCR kemudian divisualisasi selama 60 menit dengan direndam dalam gel agarosa 1,2% pada mesin ultraviolet.
Pengurutan produk PCR (Sekuensing) DNA Genus Anadara
Sekuensing adalah metode yang digunakan untuk mengetahui variasi genetik gen COI. Pembacaan sekuen DNA dilakukan untuk mengetahui komposisi basa nukleotida dan asam amino suatu gen serta menganalisis kekerabatan dan jalur evolusinya. Produk PCR yang memiliki kualitas baik layak dilanjutkan ke tahap sekuensing untuk ditentukan sekuen basa nukleotidanya. Sekuensing dilakukan menggunakan metode Sanger (1977) dengan mengirimkan produk PCR tersebut ke perusahaan jasa pelayanan sekuensing.
Analisis Data
Analisis Data Morfologi Analisis pohon fenetik
Analisis pohon fenetik dibentuk berdasarkan 6 karakter fenotip dengan software Phylogenetic Analysis Using Parsimony (PAUP). Karakter yang digunakan dalam analisis ini adalah bentuk cangkang, posisi umbo pada kerang, keseimbangan antar cangkang, jumlah alur, jarak antar alur, dan warna cangkang. Pada karakter fenotip tersebut dilakukan skoring (Lampiran 2), kemudian dianalisis dengan software PAUP.
Perbandingan karakter morfometrik antarwilayah
Analisis perbandingan morfometrik antarwilayah dilakukan untuk menghindari perbedaan umur pada populasi contoh. Perbandingan morfometrik dianalisis dengan membandingkan ukuran dari parameter yang diukur. Panjang total cangkang dibandingkan dengan parameter lain seperti lebar cangkang, tebal cangkang, tinggi umbo, simetri kanan, dan simetri kiri, kemudian dianalisis dengan uji Mann-Whitney dengan selang kepercayaan 95%.
Hubungan panjang cangkang dengan karakter morfometrik lainnya
Hubungan alometrik dilakukan dengan membandingkan variabel tebal cangkang (dependent) terhadap panjang cangkang (independent). Kedua variabel tersebut diregresikan untuk mendapatkan persamaan. Untuk mengetahui perbedaan pola pertumbuhan relatif dan perbedaan ukuran dilanjutkan dengan pengujian nilai slope dan intercept dengan analisis covariance (ANCOVA) menggunakan software SPSS.
9 terdapat perbedaan ukuran pada tebal cangkang pada ukuran panjang cangkang yang sama.
Analisis diskriminan
Analisis diskriminan dilakukan untuk mengelompokkan data berdasarkan variabel-variabel kuantitatif. Analisis ini digunakan untuk menentukan pengelompokkan kerang bulu berdasarkan karakter morfometrik yang telah diukur. Analisis ini juga diperkuat dengan nilai sharing component untuk melihat terjadinya kesalahan pengelompokkan. Analisis diskriminan dilakukan dengan menggunakan software SPSS 15.
Analisis kluster
Analisis kluster dilakukan dengan menggunakan data morfometrik kerang yang diukur dari tiap wilayah. Analisis ini digunakan untuk melihat tingkat kesamaan masing-masing populasi berdasarkan karakter morfometrik tiap individu. Dendrogram dikonstruksi dengan menggunakan software SPSS 15.
Analisis Data Molekuler
Pensejajaran sekuen nukleotida Gen COI Genus Anadara
Sekuen basa nukleotida hasil sekuensing disejajarkan dengan menggunakan metoda Clustal W pada software MEGA 5.0 (Tamura et al. 2011). Sekuen nukleotida gen COI dengan primer forward dan reverse diedit dan dianalisis untuk mendapatkan sekuen DNA dari gen COI. Sebagai ingroup, sekuen gen COI genus Anadara disejajarkan dengan genus lain dari famili yang sama, Anadarinae dan Arcinae.
Jarak genetik
Jarak genetik sekuen gen COI dihitung menggunakan metode pairwise distance yang terdapat pada program MEGA 5.0 (Tamura et al. 2011). Hasil perhitungan jarak genetik disajikan dalam bentuk matriks data yang digunakan untuk menganalisis hubungan kekerabatan antarspesies berdasarkan pohon filogeni.
Analisis filogeni
10
3
HASIL DAN PEMBAHASAN
Hasil
Keanekaragaman kerang bulu yang ditemukan di pesisir utara Pulau Jawa
Hasil identifikasi dengan menggunakan beberapa literatur, menunjukkan bahwa dari enam lokasi penelitian diperoleh tiga spesies kerang bulu (Gambar 5, Lampiran 3). Ketiganya terdapat dalam kelas Bivalvia, ordo Arcida, famili Arcidae, dan genus Anadara. Spesies yang berasal dari Banten, Subang, Cirebon, Rembang, dan Gresik teidentifikasi sebagai Anadara inaequivalvis, spesies yang berasal dari Probolinggo teridentifikasi A. cornea, dan spesies kedua yang berasal dari Subang teridentifikasi sebagai A. gubernaculum.
Gambar 5 Kerang bulu yang ditemukan di enam pesisir utara Pulau Jawa
(1.Banten, 2. Subang, 3. Cirebon, 4. Rembang, 5. Gresik, 6. Probolinggo)
Karakter fenotip kerang bulu
Analisis fenotip disajikan dengan menganalisis karakter fenotip yang diduga dari tiga spesies yang ditemukan (Gambar 6). Persamaan dari ketiga spesies tersebut adalah memiliki rambut-rambut halus yang menutupi cangkang, sehingga untuk menganalisis lebih lanjut, rambut halus tersebut dihilangkan terlebih dahulu. Hasil pengamatan menunjukkan bahwa terdapat karakter yang menjadi kunci identifikasi, yaitu bentuk cangkang, posisi umbo, warna cangkang, serta jumlah alur. Penjelasan lebih lanjut mengenai perbandingan karakter dari ketiga spesies tersebut terdapat pada Tabel 3.
Gambar 6 Konstruksi pohon fenetik dari tujuh populasi berbeda berdasarkan karakter fenotip
Spesies Banten
Spesies Subang (Ai)
Spesies Cirebon
Spesies Rembang
Spesies Gresik
Spesies Probolinggo
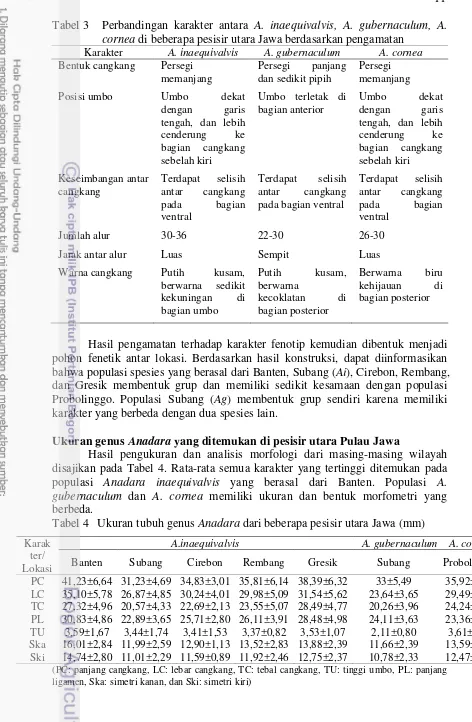
11 Tabel 3 Perbandingan karakter antara A. inaequivalvis, A. gubernaculum, A.
cornea di beberapa pesisir utara Jawa berdasarkan pengamatan
Karakter A. inaequivalvis A. gubernaculum A. cornea
Bentuk cangkang Persegi
Hasil pengamatan terhadap karakter fenotip kemudian dibentuk menjadi pohon fenetik antar lokasi. Berdasarkan hasil konstruksi, dapat diinformasikan bahwa populasi spesies yang berasal dari Banten, Subang (Ai), Cirebon, Rembang, dan Gresik membentuk grup dan memiliki sedikit kesamaan dengan populasi Probolinggo. Populasi Subang (Ag) membentuk grup sendiri karena memiliki karakter yang berbeda dengan dua spesies lain.
Ukuran genus Anadara yang ditemukan di pesisir utara Pulau Jawa
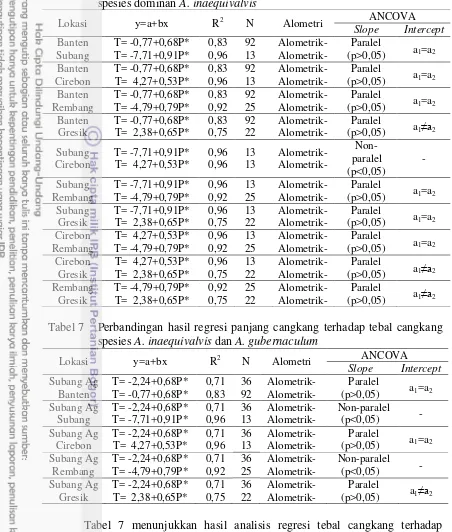
Hasil pengukuran dan analisis morfologi dari masing-masing wilayah disajikan pada Tabel 4. Rata-rata semua karakter yang tertinggi ditemukan pada populasi Anadara inaequivalvis yang berasal dari Banten. Populasi A. gubernaculum dan A. cornea memiliki ukuran dan bentuk morfometri yang berbeda.
Tabel 4 Ukuran tubuh genus Anadara dari beberapa pesisir utara Jawa (mm) Karak
ter/ Lokasi
A.inaequivalvis A. gubernaculum A. cornea
Banten Subang Cirebon Rembang Gresik Subang Probolinggo
PC
TC 27,32±4,96 20,57±4,33 22,69±2,13 23,55±5,07 28,49±4,77 20,26±3,96 24,24±2,69
PL 30,83±4,86 22,89±3,65 25,71±2,80 26,11±3,91 28,48±4,98 24,11±3,63 23,36±2,19
TU 3,59±1,67 3,44±1,74 3,41±1,53 3,37±0,82 3,53±1,07 2,11±0,80 3,61±1,46 (PC: panjang cangkang, LC: lebar cangkang, TC: tebal cangkang, TU: tinggi umbo, PL: panjang
12
Gambar 7 Distribusi panjang cangkang kerang bulu yang ditemukan di pesisir utara Pulau Jawa
Gambar 8 Distribusi panjang cangkang kerang bulu spesies A. inaequivalvis yang ditemukan di lima lokasi penelitian
Berdasarkan sebaran panjang cangkang dari tujuh populasi kerang bulu, populasi Banten memiliki sebaran selang kelas terluas, dengan panjang cangkang maksimum sebesar 54,72 mm. Populasi Subang (Ag) memiliki sebaran selang kelas panjang cangkang yang paling sempit. Populasi dominan kerang A. inaequivalvis yang berasal dari Banten memiliki selang kelas terluas, sedangkan populasi A. inaequivalvis yang berasal dari Cirebon memiliki selang kelas
13 panjang yang sempit. Perbedaan ukuran cangkang dapat terjadi dikarenakan perbedaan habitat antar populasi.
Perbandingan karakter morfometrik tiap spesies dan lokasi
Hasil perbandingan rasio karakter morfometrik terhadap panjang cangkang dari ke enam lokasi menunjukkan adanya perbedaan yang signifikan untuk spesies A. inaequivalvis.
Tabel 5 Perbandingan karakter morfometrik genus Anadara dari beberapa perairan di utara Pulau Jawa
Karakter Banten Subang Cirebon Rembang Gresik Subang
(Ag)
Probo linggo
LC/PC 0,85±0,04 0,86±0,03 0,86±0,06 0,84±0,04b 0,82±0,05ab 0,72±0,04 0,82±0,07
TC/PC 0,66±0,05 0,65±0,04 0,65±0,04 0,65±0,05 0,72±0,10abcd 0,61±0,04 0,68±0,07
TU/PC 0,09±0,04 0,11±0,04 0,10±0,03 0,09±0,02 0,10±0,04 0,07±0,03 0,10±0,04
PL/PC 0,75±0,03 0,73±0,02a 0,74±0,03a 0,73±0,05 0,74±0,05bc 0,73±0,03 0,65±0,06
Ska/PC 0,39±0,02 0,38±0,03 0,37±0,01ab 0,38±0,03 0,36±0,03ab 0,35±0,02 0,38±0,04
Ski/PC 0,36±0,04 0,35±0,03 0,33±0,02ab 0,33±0,03ab 0,33±0,02ab 0,33±0,03 0,35±0,04
Ket: Berdasarkan hasil uji Mann-Whitney, huruf pada superskrip yang berbeda menunjukkan sifat berbeda nyata (p<0,05), sedangkan superskrip yang sama menunjukkan adanya sifat sama (p>0,05)
(PC: panjang cangkang, LC: lebar cangkang, TC: tebal cangkang, TU: tinggi umbo, PL: panjang ligamen, Ska: simetri kanan, dan Ski: simetri kiri)
Hasil uji Mann-Whitney berdasarkan 6 karakter morfometrik kerang bulu, kerang bulu yang berasal dari Subang (Ag) dan Probolinggo memiliki karakter yang berbeda (p<0,05) dengan kerang bulu yang berasal dari Banten, Subang, Cirebon, Rembang, dan Gresik. Hal tersebut dikarenakan spesies yang berbeda, sehingga ukuran karakter morfometrik juga berbeda. Populasi dominan A. inaequivalvis yang berasal dari Gresik memiliki karakter rasio tebal cangkang dengan panjang cangkang yang berbeda dengan populasi lainnya. Karakter rasio tinggi umbo dengan panjang cangkang dari populasi A. inaequivalvis tidak berbeda secara signifikan (p>0,05) dari semua lokasi.
Hubungan panjang cangkang dengan karakter morfometrik lainnya
Karakter morfometrik yang digunakan sebagai variabel dependent dalam analisis ini adalah tebal cangkang. Analisis regresi panjang cangkang-tebal cangkang menunjukkan bahwa pola pertumbuhan kerang bulu alometrik negatif (Tabel 6). Hal ini mengindikasikan bahwa pertumbuhan panjang cangkang lebih dominan dibandingkan dengan tebal cangkang.
14
Tabel 6 Perbandingan hasil regresi panjang cangkang terhadap tebal cangkang spesies dominan A. inaequivalvis
Tabel 7 Perbandingan hasil regresi panjang cangkang terhadap tebal cangkang spesies A. inaequivalvis dan A. gubernaculum
Lokasi y=a+bx R2 N Alometri ANCOVA
15 Subang dan Cirebon memiliki persamaan pertumbuhan tebal cangkang terhadap panjang cangkang yang sama.
Tabel 8 Perbandingan hasil regresi panjang cangkang terhadap tebal cangkang spesies A. inaequivalvis dan A. cornea
Lokasi y=a+bx R2 N Alometri ANCOVA dengan populasi A. inaequivalvis. Berdasarkan nilai slope, A. cornea memiliki laju pertumbuhan yang sama (p>0,05) dengan populasi A. inaequivalvis dari Cirebon dan A. gubernaculum dari Subang. Nilai intercept menunjukkan bahwa terdapat perbedaan pertumbuhan tebal cangkang pada panjang cangkang yang sama antara populasi A, gubernaculum dari Subang dengan A. cornea, sedangkan memiliki persamaan pertumbuhan tebal cangkang pada panjang cangkang yang sama antara populasi A. cornea dengan A. inaequivalvis dari Cirebon.
Analisis diskriminan kerang bulu
Analisis diskriminan yang dilakukan menunjukkan bahwa 7 populasi kerang bulu terpisah secara jelas (Gambar 9). Populasi kerang bulu yang berasal dari Banten, Subang, Cirebon, dan Rembang terlihat mengelompok, dan dekat dengan populasi dari Gresik. Populasi kerang bulu yang berasal dari Subang (Ag) dan Probolinggo terpisah jauh dari populasi lainnya.
16
Gambar 9 Plot sebaran morfometrik kerang bulu dari tujuh populasi berdasarkan analisis diskriminan
17 Berdasarkan hasil analisis diskriminan, diperoleh nilai sharing component dari masing-masing populasi (Tabel 9 dan Tabel 10). Adanya nilai sharing component diakibatkan oleh kesamaan karakter morfometrik pada kerang bulu antar lokasi. Berdasarkan hasil analisis diketahui bahwa populasi Subang dan Cirebon memiliki karakter morfometrik yang tersebar hampir di seluruh lokasi pengambilan contoh. Populasi Subang (Ag) dan populasi Probolinggo memiliki karakter yang sedikit ditemui di lokasi lainnya.
Tabel 9 Hasil klasifikasi dari tujuh populasi kerang bulu berdasarkan analisis
Tabel 10 Hasil klasifikasi dari populasi dominan A. inaequivalvis berdasarkan analisis diskriminan
Kelompok observasi
Lokasi Kelompok prediksi (ind)
Banten Subang(Ai) Cirebon Rembang Gresik Total
Banten 47 6 0 33 5 92
Hasil analisis kluster dibangun berdasarkan semua karakter morfometrik (Gambar 11). Analisis kluster menunjukkan bahwa populasi kerang bulu yang berasal dari Banten, Gresik, dan Rembang membentuk satu kelompok serta populasi dari Subang (Ai) dan Cirebon membentuk satu kelompok, keduanya membentuk satu kelompok besar. Populasi kerang bulu yang berasal dari Subang (Ag) membentuk kelompok sendiri.
18
Gambar 12 Analisis kluster spesies dominan A. inaequivalvis dari lima populasi berdasarkan karakter morfometrik
DNA Total
Isolasi dan ekstraksi DNA dari enam lokasi menghasilkan kualitas DNA yang baik. Penentuan kualitas dilakukan melalui pengujian terhadap DNA total yang dimigrasikan pada agarosa 1,2% (Gambar 13). DNA yang baik ditunjukkan dengan pita DNA yang terang dan DNA tersebut layak untuk dijadikan cetakan proses amplifikasi gen COI dengan teknik PCR. DNA dari semua lokasi akan dilanjutkan ke proses amplifikasi.
Gambar 13 Uji kualitas DNA total yang dimigrasikan pada gel agarosa 1,2% Ket : 1. AQK (Banten), 2. AQS (Subang), 3. AQS.1 (Subang), 4. AQC (Cirebon), 5. AQR (Rembang), 6. AQG (Gresik), 7. AQP (Probolinggo)
Amplifkasi Fragmen DNA Gen COI pada AQP dan AQS.1
Amplifikasi fragmen DNA gen COI dilakukan dengan metode PCR pada suhu optimum annealing sebesar 53ºC untuk primer LCO1490 dan HC02198
19 (Folmer et al. 2004) serta 50ºC untuk primer Fish F1 dan Fish F2 (Ward et al. 2005). Amplifikasi gen COI target berukuran 500-750 pb (Gambar 14). Dua contoh DNA hasil PCR yang muncul (ditunjukkan dengan pita terang), yaitu DNA spesies yang berasal dari Subang dan Probolinggo, sedangkan untuk DNA dari lokasi yang lainnya tidak muncul.
Gambar 14 Elektroforesis DNA hasil pre-test produk PCR pada gel agarosa 1,2%
Urutan nukleotida Gen COI pada AQP dan AQS.1
Sekuen nukleotida gen COI pada genus Anadara yang berasal dari Subang (AQS.1) dan Probolinggo (AQP) yang diperoleh dari hasil sekuensing, dicocokkan pada BLASTn yang terdapat pada GenBank. Hasil BLASTn pada situs NCBI disajikan pada Tabel 11.
Tabel 11 Hasil BLASTn gen COI pada AQP dan AQS.1
Kode Sampel Deskripsi Kemiripan Query cover Kode Akses AQP
(Probolinggo)
Scapharca cornea* isolate H401 cytochrome oxidase subunit I (COI) gene, partial cds; mitochondrial
86% 64% KU341864.1
S. cornea
(China)
AQS.1 (Subang)
Scapharca gubernaculum*
isolate H364 cytochrome oxidase subunit I (COI) gene, partial cds; mitochondrial
98% 63% KU341857.1
S. gubernaculum
(China)
Ket: * Validasi berdasarkan WoRMS (World Register of Marine Species) nama ilmiah Scapharca berstatus unaccepted, sehingga nama imiah yang diakui adalah Anadara.
AQP
750 pb 500 pb
AQS.1
20
Berdasarkan hasil BLASTn, urutan nukleotida pada kode sampel AQP yang berasal dari Probolinggo memiliki kemiripan sebesar 86% dengan spesies Scapharca cornea yang berasal dari China. Kode sampel AQS.1 yang berasal dari Subang memiliki kemiripan sebesar 98% dengan spesies S. gubernaculum yang berasal dari China. Sementara itu, belum ada sekuen kedua spesies tersebut yang berasal dari Indonesia di GenBank. Berdasarkan hasil pencarian, diketahui bahwa sejauh ini belum terdapat individu yang memiliki kemiripan di atas 90% untuk spesies S. cornea di GenBank.
Jarak Genetik dan Analisis Filogeni Gen COI pada Famili Arcidae
Jarak genetik fragmen gen COI spesies yang diteliti AQP dan AQS.1 yaitu berkisar antara 0,005-0,125. Jarak genetik menggambarkan hubungan kekerabatan antarspesies. Nilai jarak genetik dapat dilihat pada matriks di bawah (Tabel 12).
Tabel 12 Matriks jarak genetik spesies A. inaequivalvis, A. cornea dan famili Arcidae berdasarkan metode pairwise distance
1 2 3 4 5 6 7 8 9
Data dari matriks tersebut digunakan untuk analisis hubungan kekerabatan berdasarkan pohon filogeni. Perbedaan jarak genetik terendah dari spesies yang diteliti dimiliki oleh spesies AQS.1 yang berasal dari Subang dengan spesies S. gubernaculum yang berasal dari China sebesar 0,005. Hal tersebut menunjukkan bahwa kedua spesies tersebut memiliki hubungan kekerabatan yang dekat karena merupakan spesies yang sama. Jarak genetik terbesar dimiliki oleh spesies AQP yang berasal dari Probolinggo dengan S. cornea yang berasal dari China sebesar 0,125 yang menandakan bahwa kedua spesies tersebut memiliki hubungan kekerabatan yang jauh. Semakin besar jarak genetik, semakin jauh hubungan kekerabatan, karena perbedaan basa nukleotida yang tinggi.
21 menunjukkan hubungan kekerabatan yang sangat erat. Hal ini disebabkan spesies yang berada pada subfamili Anadarinae merupakan satu keturunan (satu nenek moyang) dan digolongkan dalam spesies monofiletik.
Hasil analisis filogeni menunjukkan bahwa spesies AQS. 1 yang berasal dari Subang berada pada grup yang sama dengan spesies S. gubernaculum yang berasal dari China. Spesies AQP yang berasal dari Probolinggo membentuk grup yang sama dengan S. cornea yang berasal dari China. Gambar 15 menunjukkan pohon filogeni berdasarkan kekerabatan antarspesies dalam famili Arcidae.
Gambar 15 Pohon filogeni berdasarkan gen COI dari famili Arcidae
Pembahasan
Kajian morfologi kerang bulu
Kerang bulu yang ditemukan pada penelitian ini hanya berjumlah tiga spesies dari enam lokasi penelitian, yaitu Anadara inaequivalvis, A. cornea, dan A. gubernaculum. A. inaequivalvis dan A. gubernaculum ditemukan pada kedalaman 0-10 meter pada substrat yang berlumpur. A. cornea dapat ditemukan pada perairan dengan kedalaman 0-25 meter, mulai dari substrat berlumpur di ekosistem mangrove hingga ekosistem terumbu karang seperti yang ditemukan oleh Prabhakaran et al. (2012) di ekosistem mangrove pada Laguna Minicoy, Lakshadweep India. Menurut nelayan dari enam lokasi penelitian, ketiga spesies tersebut dapat ditemukan pada hasil tangkapan nelayan, sebagai spesies bycatch, namun beberapa daerah mengumpulkan kerang bulu tersebut dengan menggunakan alat tangkap garok atau dengan mengambil langsung menggunakan tangan.
Ketiga genus Anadara tersebut memiliki karakteristik unik, yaitu bagian cangkang luar yang ditutupi oleh rambut-rambut halus berwarna cokelat kehitaman. Rambut-rambut halus tersebut menurut Dixon et al. (1995) struktur yang dibentuk dari lapisan periostrakum yang berfungsi penting untuk melindungi organisme dari predator mau pun hewan-hewan yang bersimbiosis. Contoh lainnya dapat ditemukan pada famili Mytiloida dan Veneroida (Watabe 1988). Beberapa kemungkinan fungsi lain dari rambut-rambut halus tersebut adalah melindungi dari serangan predator (Harper dan Skelton 1993; Bottjer dan Carter
Subfamili Anadarinae
22
1980), sebagai proteksi kerang dari ancaman perubahan lingkungan, menstabilisasi kerang di substrat, kamuflase, melindungi bagian mantel dan organ-organ dalam dari kerang, membantu mengapungkan spat (kerang juvenil) ketika melakukan proses settlement, membantu menyaring makanan yang masuk ke dalam tubuh kerang, dan sebagai mekanisme kerang untuk menjaga tubuhnya agar tetap bersih terutama pada bagian bysus.
a. Anadara inaequivalvis b. Anadara gubernaculum
c. Anadara cornea
Gambar 16 Penampakan luar, dalam, dorsal, dan ventral dari kerang A. inaequivalvis, A. gubernaculum, dan A. cornea
23 bentuk cangkang, jumlah alur dan warna cangkang yang berbeda. Bahkan menurut Albano et al. (2009), chevron yang berada pada ligamen memerupakan karakter penting sebagai pembeda antar spesies.
A. cornea memiliki karakter yang unik dengan warna cangkang bagian posterior biru kehijauan, karakter tersebut juga ditemukan dalam penelitian mengenai A. cornea di Laguna Minicoy Lakshadweep India (Prabhakaran et al. 2012). Brake et al. (2004) melaporkan bahwa adanya variasi pigmentasi pada kerang dapat disebabkan oleh faktor genetik dan faktor lingkungan, seperti pengaruh makanan terhadap warna cangkang (Bauchau 2001). Cangkang kerang mengandung protein (Souji et al. 2014) dan karbohidrat yang sangat berpengaruh terhadap struktur morfologi cangkang dan dapat berpengaruh terhadap susunan lapisan luar cangkang (Hedegaard et al. 2005).
Variasi bentuk cangkang kerang dapat dipengaruhi dari perbedaan variasi genetik dari masing-masing kerang (Krakau 2008). Banyak spesies yang mirip secara fenotip, baik dari bentuk tubuh hingga anatomi tubuh. Bahkan identifikasi dengan karakteristik morfometrik dan meristik masih banyak kesalahan, hal tersebut dapat menjadi hambatan dalam menentukan taksonomi dari spesies. Beberapa dekade terakhir, muncul metode yang lebih baik dalam menentukan taksonomi pada cryptic atau sibling species, yaitu dengan marka genetik melalui DNA (Kruse et al. 2003).
Spesies A. inaequivalvis tidak berbeda secara signifikan (p>0,05), kecuali pada spesies yang berasal dari Gresik. Hal tersebut dapat disebabkan karena perbedaan kondisi perairan di daerah Gresik sedikit berbeda dengan daerah yang lain, seperti salinitas, suhu, tipe sedimen, dsb, sehingga adaptasi kerang terhadap lingkungan juga berbeda. Pada dasarnya, proses adaptasi terhadap kondisi lingkungan seperti salinitas dan karakter sedimen dialami oleh beberapa spesies, seperti Cerastoderma edule dan C. glaucum di perairan Atlantik Utara (Mariani et al. 2002). Fluktuasi lingkungan mempunyai pengaruh yang sangat besar terhadap morfologi organisme, karena plastisitas fenotip tinggi pada organisme yang menetap seperti kerang (Imre et al. 2001).
Pertumbuhan cangkang kerang bulu pada penelitian ini adalah alometrik negatif, yaitu panjang cangkang memiliki pertumbuhan panjang cangkang yang lebih cepat dibandingkan dengan karakter lainnya. Hasil uji ANCOVA dari regresi panjang cangkang terhadap tebal cangkang memperlihatkan bahwa populasi kerang bulu di Subang, Cirebon, dan Probolinggo memiliki ukuran tebal cangkang yang berbeda pada panjang cangkang yang sama. Perbedaan laju pertumbuhan dan bentuk cangkang dapat dipengaruhi oleh pasang surut (De Montaudoin 1996), kepadatan populasi dan kompetisi antar populasi, perbedaan salinitas dan temperatur (Mariani et al. 2002) serta perbedaan tipe sedimen.
24
hasil analisis keragaman fenotipik dengan plot diskriminan dan analisis kluster yang dibentuk dari hasil pengukuran morfometrik tidak menunjukkan perbedaan.
Pembentukan kelompok ini diduga adanya perbedaan populasi dari wilayah tersebut dapat disebabkan karena adanya perbedaan wilayah berdasarkan topografi (Krakau 2008), tipe habitat dan gradien lingkungan lainnya meliputi substrat, salinitas, suhu (Mariani et al. 2002; Peyer et al. 2010), kedalaman, pasang-surut, arus, serta paparan ombak (Kandratavicius dan Brazeiro 2014; Versteegh 2012; Conde-Padin et al. 2007) dan fluktuasi lingkungan memiliki dampak yang besar terhadap karakter morfologi dari kerang, seperti bentuk dan ukuran kerang yang berasal dari perbedaan ekosistem (Gaylord dan Gaines 2000; Funk dan Reckendorfer 2008). Perbedaan kondisi habitat tersebut dapat menyebabkan faktor stres secara alami terhadap organisme. Keadaan tersebut memaksa spesies yang tinggal di perairan untuk beradaptasi. Strategi penyesuaian organisme dengan lingkungannya dilakukan dengan cara perubahan biokimia, fisiologis, dan adaptasi dengan perubahan genetik (Pigliucci 2005) dalam jangka waktu yang panjang (Peyer et al. 2010; Evans dan Hofmann 2012).
Berdasarkan hasil penelitian ini diperoleh sebaran kerang bulu dari beberapa pesisir utara Pulau Jawa (Gambar 17). Sebaran kerang bulu diperlukan sebagai informasi awal dalam membantu menentukan pengelolaan sumber daya agar tidak terjadi kesalahan dalam penentuan spesies. Perbedaan spesies dan habitat dapat memberikan dampak yang berbeda dalam pengelolaannya.
Gambar 17 Sebaran kerang bulu di pesisir Banten, Subang, Cirebon, Rembang, Gresik, dan Probolinggo
25 alien species, karena keberadaannya mengganggu spesies endemik dan spesies lain disekitarnya. Penelitian Anistratenko et al. (2014) di Laut Azov Rusia, Sahin et al. (2009), Kolyuchina dan Miljutin (2013), dan Aydin et al. (2014) di Laut Hitam Turki membahas mengenai A. inaequivalvis yang menjadi spesies introduksi yang diduga berasal dari air ballast yang dibawa oleh kapal yang berasal dari bagian Asia Selatan ke bagian Asia Barat hingga ke Eropa. Awalnya, Sahin et al. (1999) menduga bahwa spesies A. inaequivalvis teridentifikasi sebagai A. cornea karena memiliki bentuk tubuh yang sama. Beberapa tahun setelahnya diinformasikan bahwa spesies tersebut merupakan A. inaequivalvis. Tetapi pada tahun 2006, Lutaenko memperhatikan bahwa terjadi perbedaan morfologi antara spesimen yang berasal dari Eropa dengan yang berasal dari Asia, dan dibuktikan oleh Huber (2010) bahwa spesies yang menyerang kawasan Eropa merupakan spesies A. kagosimensis yang berasal dari Jepang. Krapal et al. (2014) telah melakukan konfirmasi bahwa secara molekuler spesies yang berada di Eropa merupakan A. kagosimensis.
Diketahui bahwa spesies A. inaequivalvis merupakan spesies native yang berasal dari indo-pasifik, tetapi di Laut Hitam, Turki dan di Laut Azou Rusia teridentifikasi sebagai invansive alien species. Alien species merupakan spesies introduksi dari luar distribusi aslinya (Lutaenko 2007). Spesies invasive dapat mengancam keberadaan spesies endemik dan spesies lainnya, sehingga dapat memberikan dampak negatif terhadap keseimbangan ekosistem di alam hingga dapat menurunkan keanekaragaman dan kelimpahan spesies. Kondisi tersebut dapat memperparah dan mengancam kesehatan serta kesejahteraan masyarakat. Spesies ini merupakan spesies ekonomis penting yang dapat dikonsumsi, sehingga perlu adanya pemantauan secara kontinu sebagai salah satu langkah awal pengelolaan sumberdaya kerang bulu. Pada penelitian ini hanya menemukan penciri dari suatu spesies, tetapi pada spesies dominan A. inaequivalvis belum diketahui perbedaan penciri antar lokasi.
Kajian molekuler kerang bulu
Berdasarkan asumsi dari Combosh dan Giribet (2016), famili Arcidae ordo Arcida (ark shells) merupakan salah satu famili tertua di kelas Bivalvia, yang diketahui sudah ada pada zaman Ordovician (~450 Mya). Berdasarkan fosil yang ditemukan, Arcida banyak ditemukan pada masa Mesozoic dan pertama kali muncul pada zaman Jurassic. Perkembangan famili Arcidae, khususnya Anadarinae meningkat pada zaman Cretaceous, karena pada saat itu sudah masuk dalam iklim yang hangat dan tinggi permukaan laut mulai meningkat, sehingga dapat menyediakan habitat baru untuk Anadarinae (Oliver dan Holmes 2006; Jackson et al. 2015). Beberapa spesies dari famili Arcidae terdistribusi luas dan melimpah di perairan tropis. Famili ini memiliki nilai ekonomis penting sebagai bahan konsumsi, contohnya kerang darah (Anadara granosa) yang dibudidayakan di China, Taiwan, Korea, Malaysia, dan Thailand; Scapharca dibudidayakan di Jepang dan China (Oliver dan Holmes 2006). Penyebutan nama “kerang darah” di beberapa daerah membuat adanya kesalahpahaman dalam membedakan spesies yang berada pada famili Arcidae.
26
dengan penelitian Alfaya et al. (2015) mengenai DNA barcoding cacing Malacobdella. Urutan basa nukleotida fragmen gen COI pada cacing Malacobdella hanya memiliki kemiripan sebesar 85% dengan spesies cacing Malacobdella yang terdapat pada GenBank berdasarkan analisis BLASTn. Beberapa alasan hal tersebut dapat terjadi karena spesies tersebut sangat sulit diidentifikasi bahkan dengan menggunakan karakter morfologi, dan adanya variasi intraspesifik atau perubahan karakter morfologi karena pengaruh lingkungan juga dapat mempengaruhi perbedaan hingga perubahan susunan basa nukleotida dari suatu spesies.
Menurut Marko (2002), urutan basa nukleotida suatu organisme dipengaruhi oleh faktor lingkungan eksternal seperti kondisi habitat dan letak geografis habitat dari organisme tersebut. Organisme mengembangkan daya adaptasi untuk merespon lingkungan yang berbeda. Fragmentasi habitat menyebabkan terjadinya aliran gen (gene flow) pada organisme yang memiliki fase hidup sebagai larva planktonik. Seringkali fragmentasi habitat bukan hanya menjadi faktor penyebab diferensiasi populasi, tetapi juga diferensiasi spesies bahkan subspesies. Konektivitas organisme atau spesies dapat terjadi pada kondisi lingkungan yang minim atau tidak ada penghalang fisik, kimiawi maupun biologi (isolasi geografis) (Luttikhuizen et al. 2003; Chiu et al. 2013). Pengaruh lingkungan dan posisi geografis yang berjauhan walaupun dengan tingkat konektivitas yang tinggi juga dapat menyebabkan suatu organisme membutuhkan adaptasi lokal, sehingga organisme tersebut perlu mengembangkan plastisitas fenotip (Sokolowski et al. 2004; Tarnowska et al. 2012; Butet 2013). Plastisitas fenotip merupakan kemampuan suatu organisme memiliki karakter genotip yang sama pada habitat yang berbeda untuk mengembangkan keragaman fenotip (Pigliucci et al. 2006). Hal yang berlainan terjadi pada spesies contoh AQP yang memiliki urutan nukleotida yang jauh berbeda dengan spesies A. cornea dari China. Kedua organisme ini diduga berasal dari nenek moyang yang sama, karena letak geografis yang berjauhan maka kedua organisme tersebut mengembangkan daya adaptasi yang berbeda pada tingkat molekuler walaupun secara morfologi memiliki kemiripan yang sama. Fenomena ini sering dikaitkan dengan cryptic species (Marko 2002).
Spesies A. gubernaculum yang ditemukan di Subang memiliki kemiripan sebesar 98% dengan spesies A. gubernaculum yang berasal dari China. Menurut Hebert et al. (2003), persentase minimal untuk mengidentifikasi kemiripan suatu spesies adalah sebesar 97,7%, di bawah persentase tersebut dapat disimpulkan bahwa spesies yang diteliti kemungkinan bukan spesies yang ditargetkan. Isolasi geografis yang terjadi pada A. gubernaculum tidak menjadi faktor penghambat aliran gen. Baik karakter morfologi maupun susunan basa nukleotida gen COI, keduanya masih menunjukkan kemiripan yang tinggi. Susunan basa nukleotida gen COI A. gubernaculum mampu beradaptasi dengan lingkungan lokal dengan cara tetap mempertahankan karakter nukleotida dari nenek moyang.
27 bahwa karena adanya kesamaan keturunan (nenek moyang), maka memungkinkan terjadinya fenomena cryptic species (Geller et al. 2010).
Spesies AQP dan AQS.1 memiliki kekerabatan dengan spesies A. inaequivalvis, dan A. antiquata yang diketahui merupakan kelompok kerang bulu. Mereka termasuk ke dalam sister’s relationship dan bersifat monofiletik karena berasal dari nenek moyang yang sama dan genus yang sama. Adanya hubungan tersebut menyebabkan morfologi dari kelompok kerang bulu tersebut terlihat sama dan sulit dibedakan. Berdasarkan kajian morfologi dari bab sebelumnya, disebutkan bahwa perbedaan morfologi dari spesies yang dikaji A. cornea, A. gubernaculum, dan A. inaequivalvis adalah terdapat pada bentuk dan ukuran tubuh kerang, warna cangkang serta jumlah alur. Oleh sebab itu, pendekatan morfologi saja tidak cukup untuk membuktikan kepastian taksonomi dari suatu spesies. Berdasarkan hal tersebut, dapat disimpulkan bahwa kerang bulu merupakan salah satu kelompok cryptic species. Sehingga analisis molekuler merupakan solusi yang tepat untuk mengkaji bidoversitas cryptic (Miura dan Chiba 2007).
Pengelolaan sumber daya kerang bulu
Telah terjadi penurunan moluska pada tahun 1937 - 1938 di Teluk Jakarta dikarenakan pengaruh lingkungan. Beberapa koleksi moluska yang terdiri dari kelas bivalvia dan gastropoda yang dikumpulkan dari Teluk Jakarta hingga ke wilayah Kepulauan Seribu mengalami perbedaan kekayaan jenis dari tahun 1937/1938 hingga tahun 2005. Menurut Meij et al. (2009), 16 genera dari 70 genera yang ditemukan telah hilang, pada tahun 1937/1938 ditemukan 171 spesies sedangkan pada tahun 2005 hanya ditemukan 58 spesies. Penyebab terjadinya penurunan moluska tersebut dikarenakan faktor lingkungan dan pengaruh antropogenik. Peningkatan jumlah masukan limbah ke perairan, jumlah sedimen yang masuk ke badan perairan dari hulu hingga ke hilir dapat mengganggu ekosistem pesisir dari mangrove hingga terumbu karang, sehingga moluska yang berasosiasi dengan ekosistem tersebut banyak yang hilang.
Filum moluska tersebar luas di perairan, tetapi hal tersebut tidak membatasi ancaman terhadap filum ini. Beberapa diantaranya adalah hilangnya ekosistem mangrove dan ekosistem terumbu karang; perubahan habitat seperti sedimen, salinitas, dsb; meningkatnya tinggi muka air laut pengaruh dari global warming; adanya bencana seperti badai dan tsunami; adanya perubahan pola curah hujan; terjadinya overfishing hingga ke arah overeksploitasi; adanya pencemaran perairan; masuknya spesies introduksi dan adanya gangguan fisik dari pesisir akibat kegiatan ekowisata. Alasan tersebut dapat menurunkan kelimpahan dan keanekaragaman moluska yang ada di perairan, termasuk kerang bulu yang diteliti.
28
cangkang, bobot tubuh, dan bobot total dari cangkang kerang dengan kriteria sebagai berikut. Ukuran tersebut dapat dijadikan sebagai alat mengetahui pencemaran suatu perairan, contohnya logam berat. Dengan demikian, dari karakter morfologi dapat diketahui batas aman dalam mengkonsumsi kerang. Tabel 13 Kriteria pencemaran berdasarkan ukuran morfologi
Karakter morfologi Kriteria pencemaran
Tinggi Sedang Rendah
Tinggi (tebal) (mm) >2 1,5-2 <1,5
Panjang (mm) >3 2,5-3 <2,5
Lebar (mm) >2 1,9-2 <1,9
Bobot tubuh (gr) >2,2 1,7-2,2 <1,7
Bobot total (gr) >10 5,3-10 <5,3
Pengelolaan kerang bulu sangat dipengaruhi oleh faktor lingkungan, mengingat perubahan morfologi pada kerang sangat dipengaruhi oleh fluktuasi lingkungan. Bentuk yang abnormal dapat menyulitkan proses identifikasi. Beberapa kasus yang ditemukan adanya kesalahan identifikasi karena kerang bulu merupakan cryptic species, maka proses identifikasi harus dilakukan dengan benar, salah satunya dibutuhkan kajian molekuler sebagai informasi awal mengenai keberadaan suatu spesies kerang bulu. Cryptic species memiliki pengelolaan tersendiri. Cryptic species tidak boleh diambil dan dikoleksi secara terus menerus, karena akan mengurangi populasinya di alam. Bisa terjadi kemungkinan spesies yang dikoleksi merupakan spesies endemik dari suatu wilayah. A. cornea dan A. gubernculum yang ditemukan pada penelitian ini sangat langka dan jarang ditemukan di Indonesia, sehingga membutuhkan pengelolaan yang baik.
Beberapa daerah di dunia, seperti Jepang dan China (FAO 1998; Combosch dan Giribet 2016) melakukan budidaya kerang. Hal ini dikarenakan nilai ekonomis dari kerang bulu tersebut sangat tinggi, sehingga untuk mencegah berkurangnya stok di alam. Kajian molekuler sangat dibutuhkan dalam penentuan spesies yang dibudidaya, karena dikhawatirkan apabila salah dalam menentukan spesies yang tepat, terdapat spesies “opportunis” yang dapat menjadi predator bagi spesies yang lainnya.
29 Secara morfologi, ketiga spesies yang ditemukan dapat digolongkan sebagai cryptic species. Hasil identifikasi morfologi menunjukkan bahwa A. cornea yang berasal dari Probolinggo memiliki karakter warna cangkang yang unik dibandingkan dengan spesies lainnya, yaitu berwarna biru kehijauan di bagian posterior sehingga karakter unik tersebut dianggap eksotis. Dengan demikian A. cornea memiliki pengelolaan yang berbeda, yaitu dengan dikonservasi agar keberadaannya tidak punah. Sementara itu, pengelolaan untuk spesies A. gubernaculum yang sulit ditemukan sebaiknya juga dikelola dengan baik dengan cara melakukan konservasi agar tetap mempertahankan dan melestarikan stok A. gubernaculum di alam.
4
SIMPULAN DAN SARAN
Simpulan
Secara visual, morfologi contoh kerang bulu dari enam lokasi penelitian memiliki kemiripan yang tinggi sehingga sulit dibedakan. Perbedaan karakter morfometrik yang mencolok dari tiga spesies kerang bulu terdapat pada bentuk dan ukuran tubuh kerang, warna cangkang serta jumlah alur. Analisis morfologi berdasarkan 7 karakter, berhasil mengidentifikasi tiga spesies yang ditemukan sebagai Anadara inaequivalvis, A. cornea, dan A. gubernaculum. Analisis molekuler gen COI pada A. inaequivalvis belum berhasil ditemukan, sedangkan spesies AQS.1 yang berasal dari Subang teridentifikasi sebagai A. gubenaculum. Spesies AQP (A. cornea) yang berasal dari Probolinggo memiliki tingkat kekerabatan yang rendah dengan A. cornea yang berasal dari China, sehingga tidak menutup kemungkinan bahwa spesies tersebut merupakan spesies baru atau spesies yang belum terdaftar.
Saran
30
DAFTAR PUSTAKA
Afiati. 2007. Hermaphroditism in Anadara granosa (L.) and Anadara antiquata (L.) (Bivalvia: Arcidae) from central java. J of Coast Develop. 10 (3): 171-179.
Akhrianti I, Bengen DG, Setyobudiandi I. 2014. Distribusi spasial dan preferensi habitat bivalvia di pesisir perairan Kecamatan Simpang Pesak Kabupaten Belitung Timur. J Ilm Tek Kel Trop. 6(1): 171-185.
Albano PG, Rinaldi E, Evangelisti F, Kuan M, Sabelli, B. 2009. On the identity and origin of Anadara demiri (Bivalvia: Arcidae). JMBA. 89(6): 1289-1298. Alfaya JEF, Bigatti G, Kajihara H, Sundberg P, Machordom A. 2015. DNA barcoding supports identification of Malacobdella species (Nemertea: Hoplonemertea). Zool Studies. 54(10): 1-9. doi: 10.1186/s40555-014-0086-3.
Ambarwati R, Trijoko (2011) Kekayaan jenis Anadara (Bivalvia:Arcidae) di perairan Pantai Sidoarjo. Hayati 4B. 43: 1-8.
Anistratenko VV, Anistratenko OY, Khaliman IA. 2014. Conchological variability of Anadara inaequivalvis (Bivalvia, Arcidae) in the Black-Azov Sea Basin. Vestnik zool. 48(5): 457-466. doi: 10.2478/vzoo-2014-0054. Aydin M, Karadurmus U, Tunca E. 2014. Moprhometric aspects and growth
modeling of exotic bivalve blood cockle Scapharca inaequivalvis from the Black Sea, Turkey. Biologia. 69(12): 1707-1715. doi: 10.2478/s11756-014-0476-3.
Bauchau V. 2001. Developmental stability as the primary function of the pigmentation patterns in bivalve shells. Belgium J of Zool. 131(2):23-28. Bottjer DJ, Carter JG. 1980. Functional and phylogenetic significance of
projecting periostracal structure in the Bivalvia (Mollusca). J of Palaent. 54: 200-216.
Brake J, Evans F, Langdon C. 2004. Evidence for genetic controlt\ of pigmentation of shell and mantle edge in selected families of Pasific oysters, Crassostrea gigas. Aquaculture. 229: 89-98. doi: do.1016/S0044-8486(03)00325-9.
Butet NA. 2013. Plastisitas fenotip kerang darah Anadara granosa L. dalam merespon pencemaran lingkungan: studi kasus di perairan Pesisir Banten. [Disertasi]. Bogor (ID): Institut Pertanian Bogor.
Chiu YW, Bor H, Tan MS, Lin HD, Jean CT. 2013. Phylogeography and genetic differentiation among populations of the moon turban snail Lunella granulate Gmelin, 1791 (Gastropoda: Turbinidae). Int J Mol Sci. 14: 9062-9079. doi: 10.3390/ijms14059062.
Combosh DJ, Giribet G. 2016. Clarifying phylogenetic relationships and the evolutionary history of the bivalve order Arcida (Mollusca: Bivalvia: Pterimorphia). Mol Phylogenet Evol. 94: 298-312. doi: 10.1016/j.mpev.2015.09.016.