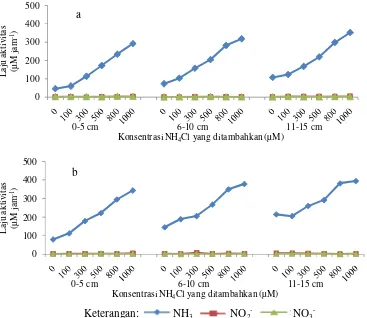
Fluks Bentik dan Potensi Laju Aktivitas Bakteri Terkait Siklus Nitrogen pada Sedimen Perairan Mangrove Karangantu dan Pulau Dua, Banten
Teks penuh
Gambar
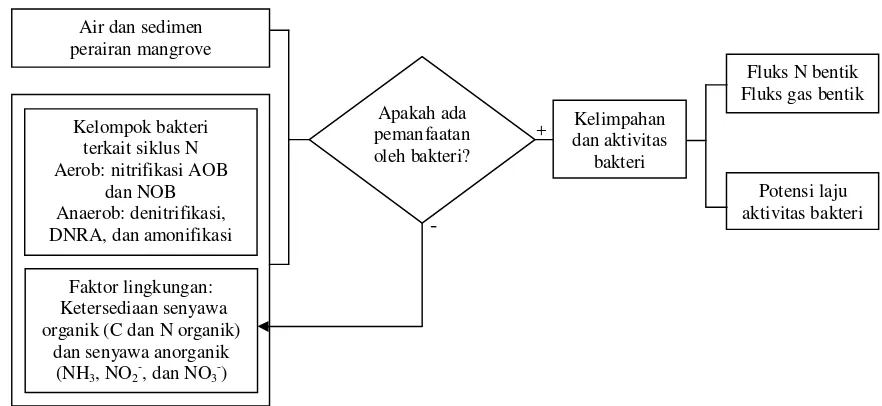
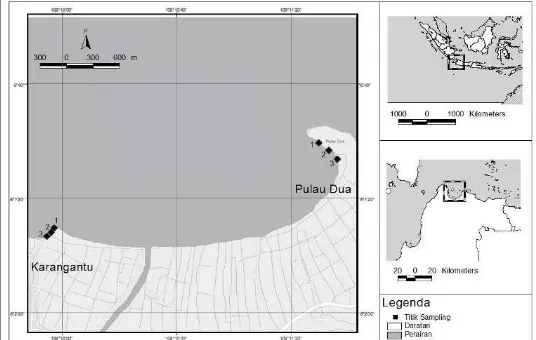

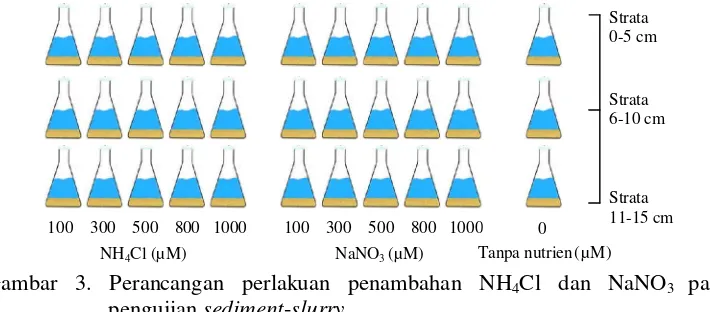
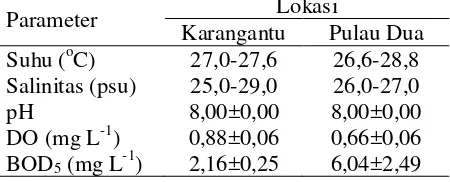
Dokumen terkait
Karbon yang tersimpan pada sedimen dan vegetasi mangrove di sana dapat membantu menurunkan laju emisi gas rumah kaca (GRK) dalam rangka mengurangi pemanasan
Tujuan penelitian ini adalah (1) Menelaah karakteristik habitat mangrove Pulau Panjang berdasarkan parameter kualitas perairan dan substratnya, (2) Mengkaji produktifitas serasah
Tujuan penelitian ini adalah (1) Menelaah karakteristik habitat mangrove Pulau Panjang berdasarkan parameter kualitas perairan dan substratnya, (2) Mengkaji produktifitas serasah
Penurunan kerapatan vegetasi mangrove dipesisir pulau sembilan tentunya akan berpengaruh terhadap penurunan kemampuan fisik akar mangrove tersebut sebagai pemerangkap sedimen
Studi Kandungan Nitrogen (N) dan Fosfor (P) Pada Sedimen Mangrove Di Wilayah Ekowisata Wonorejo Surabaya dan Pesisir Jenu Kabupaten Tuban.. Dosen Pembimbing
