IDENTIFIKASI MORFOLOGI DAN MARKA GENETIK
TERPAUT SIFAT TIDAK BERBUNGA JANTAN PADA MUTAN
PISANG KEPOK
NETTYANI NAIPOSPOS
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Identifikasi Morfologi dan Marka Genetik Terpaut Sifat Tidak Berbunga Jantan pada Mutan Pisang Kepok adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, September 2013 Nettyani Naipospos NIM G353110161
RINGKASAN
NETTYANI NAIPOSPOS. Identifikasi Morfologi dan Marka Genetik Terpaut Karakter Tidak Berbunga Jantan pada Mutan Pisang Kepok. Dibimbing oleh MIFTAHUDIN dan SOBIR.
Pisang merupakan komoditi hortikultura yang penting di Indonesia. Akan tetapi, kapasitas produksi terkendala akibat adanya serangan penyakit darah Blood Disease Bacterium (BDB) pada pisang olahan. Pisang tipe cooking banana seperti pisang kepok merupakan komoditi yang penting di beberapa daerah Indonesia Timur sebagai makanan yang dikonsumsi sehari-hari. Pada akhir tahun 1990, produksi pisang kepok asal Kalimantan secara drastis menurun akibat adanya penyakit darah yang disebabkan oleh BDB. Akibatnya, banyak petani yang kehilangan sumber pendapatannya. Penyebaran penyakit ini sangat cepat, hampir 100 km per tahun yang ditularkan melalui perantaraan serangga. Infeksi penyakit ini dapat dikurangi dengan menghindari terjadinya kontak antara serangga dengan kumpulan bunga jantan (male bud). Cara yang dapat dilakukan adalah dengan memotong male bud atau dengan menghasilkan pisang kepok yang tidak memiliki kumpulan bunga jantan (male budless).
Pusat Kajian Hortikultura Tropika (PKHT) Institut Pertanian Bogor (IPB) telah mengoleksi beberapa pisang kepok mutan male budless yang enak dimakan, dan diberi nama Unti Sayang (US-1). Stabilitas genetik dari mutan ini masih belum diketahui. Oleh karena itu, diperlukan sebuah penanda DNA yang terpaut dengan sifat male budless. Penanda ini dapat digunakan untuk menetapkan sistem perbanyakan pisang mutan male budless yang stabil dan juga untuk mempelajari mekanisme tidak terbentuknya male bud.
Penelitian ini bertujuan untuk mengidentifikasi karakter morfologi dan marka ISSR dan RAPD serta mengembangkan marka molekuler yang dikembangkan dari gen Pistillata dan Agamous terpaut karakter male budless pada mutan pisang kepok Unti Sayang (US-1).
Identifikasi karakter morfologi berdasarkan panduan deskriptor pisang International Plant Genetic Research Institute (IPGRI). Pengamatan morfologi menunjukkan tidak ada variasi morfologi antara tipe liar, mutan dan mutan yang kembali memiliki kumpulan bunga jantan (revertrant mutant). Revertrant mutant ditemukan dari perbanyakan planlet sub kultur ke-6 dan dari perbanyakan anakan generasi kedua.
Sifat male budless diduga berhubungan dengan fungsi gen Pistillata yang mengontrol pembentukan petal dan stamen dan gen Agamous yang mengontrol pembentukan karpel. Oleh karena itu, dikembangkan primer dari gen Pistillata dan Agamous berdasarkan database dari tanaman Musa yang terdapat di GeneBank. Hasil amplifikasi PCR dengan menggunakan kombinasi primer tersebut juga tidak menghasilkan pita polimorfik. Analisis sekuen nukleotida antara fragmen DNA menunjukkan adanya tiga nukleotida yang berbeda antara tanaman tipe liar dan revertrant mutant pada posisi nukleotida ke-445, 461 dan 507. Variasi nukleotida tersebut terletak pada daerah 3’UTR (untranslated region), dimana perbedaan nukleotida tersebut tidak menyebabkan perubahan pada sekuen protein dari gen tersebut. Dari hasil tersebut dapat disimpulkan bahwa kemungkinan terdapat gen lain yang mengontrol karakter male budless atau kemungkinan adanya variasi epigenetik. Oleh karena itu, diperlukan studi lebih lanjut untuk mengkaji latar belakang genetik terkait dengan terjadinya fenomena male budless pada mutan pisang kepok.
SUMMMARY
NETTYANI NAIPOSPOS. Identification of Morphological and Genetic Markers Related to Male Budless Characters on Kepok Banana Mutant. Supervised by MIFTAHUDIN and SOBIR.
Bananas is an important horticultural commodity in Indonesia. However, the improvement of production capacity of cooking banana is hampered by Blood Disease Bacterium (BDB). Cooking banana such as kepok banana is very important commodity in several areas in eastern part of Indonesia for cash crop as well as for daily consumption. Since late 90s, production of kepok banana from Kalimantan drastically declined as a result of devastating outbreak of BDB. As a result many farmers are losing their main source of income. The disease is quickly spread at about 100 km distance per year as the bacterium is insect transmitted. Disease infection could be reduced by avoiding direct contact between insect vector and male bud by breaking peduncula supporting male bud or by cultivating male budless kepok banana mutant.
Recently Centre for Tropical Horticultural Studies (PKHT) at Bogor Agricultural University (IPB) already collected several male budless kepok banana mutants with good eating quality named Unti Sayang (US-1). However, the genetic stability of these mutants is still unknown. Therefore, discovering DNA markers related to male budless character in kepok banana is very important to establish a stable mass propagation system for male budless character on kepok banana, and valuable for science to elucidate male budless mechanism in banana.
The objectives of this research were to identify morphological characters and RAPD and ISSR markers and develop Pistillata and Agamous gene based moleculer marker related to male budless characters in kepok banana mutant Unti Sayang (US-1).
Identification of morphological characters were carried out based on the International Plant Genetic Research Institute (IPGRI) descriptor for banana. The results showed that there is no morphological variation among wild type, mutants and revertrant other than a male bud character. The revertrant mutants were found among planlets derived from the sixth sub culture and second generation of suckers.
Polymerase Chain Reaction (PCR) was used to amplify DNA of 24 plants derived from the first and sixth sub cultures using 20 RAPD and 12 ISSR primer combinations. Those RAPD and ISSR primers produced monomorphic DNA bands with the amount ranged from 2 to 6 and 6 to 8 per plant, respectively. No polymorphism was found between male budless mutants and revertrant mutants. The absence of genetic variation among wild type, mutants and revertrants has been reported in several cases of molecular marker analysis in in vitro cultured banana, although there was a variation in their morphology. Based on the PCR analysis, it was suggested that there was no genetic change between wild type and mutants. However, it is possible that some changes might occurred as point mutation located outside of the priming sites.
Musa GeneBank database. PCR analysis using those primer combination have also produced no polymorphic bands among the analyzed banana plants. Carefull nucleotide sequence analysis of both DNA fragments showed three single nucleotide polymorphism between wild type and revertrant mutants at the position of 445, 461 and 507. Unfortunately, the nucleotide variation was located at
3‘UTR (untranslated region), which no any interference into protein sequence expressed by the genes. This result suggested that there might be other genes controlling the male budless variation or possibly an epigenetic variation occurred behind the variation. Further study needs to be conducted to clarify the genetic background underlaying male budless phenomenon occurred in kepok banana mutants.
© Hak Cipta Milik IPB, Tahun 2013
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
IDENTIFIKASI MORFOLOGI DAN MARKA GENETIK
TERPAUT SIFAT TIDAK BERBUNGA JANTAN PADA MUTAN
PISANG KEPOK
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2013
NETTYANI NAIPOSPOS
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Judul Tesis : Identifikasi Morfologi dan Marka Genetik Terpaut Sifat Tidak Berbunga Jantan pada Mutan Pisang Kepok
Nama : Nettyani Naipospos NIM : G353110161
Disetujui oleh Komisi Pembimbing
Dr Ir Miftahudin, MSi Ketua
Prof Dr Ir Sobir, MSi Anggota
Diketahui oleh
Ketua Program Studi Biologi Tumbuhan
Dr Ir Miftahudin, MSi
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
PRAKATA
Puji dan syukur penulis ucapkan kepada Tuhan Yang Maha Esa Yesus Kristus atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Agustus 2012 ini ialah identifikasi morfologi dan molekular, dengan judul Identifikasi Morfologi dan Marka Genetik Terpaut Sifat Tidak Berbunga Jantan pada Mutan Pisang Kepok
Terima kasih penulis ucapkan kepada Bapak Dr Ir Miftahudin MSi dan Bapak Prof Dr Ir Sobir MSi selaku pembimbing. Ucapan terima kasih penulis kepada Pusat Kajian Hortikultura Tropika (PKHT) IPB yang telah mendanai penelitian ini atas nama Bapak Prof Dr Ir Sobir MSi. Ucapan terima kasih juga penulis sampaikan kepada Direktorat Jendral Pendidikan Tinggi (DIKTI) melalui program Beasiswa Unggulan (BU) yang diberikan. Selain itu, penghargaan penulis sampaikan kepada staff pengajar Biologi Tumbuhan (BOT) dan kepada teman-teman 2011 BOT serta kepada kak Sulasih dan Pipit beserta semua keluarga besar PKHT. Terimakasih kepada seluruh keluarga, orang tua Ayahanda Birma Naipospos dan Ibunda Kezia Juliana, Adik Hevyana Naipospos dan Richy Rinad Naipospos, serta sahabat Ennie Chahyadi dan seluruh keluarga Wisma Rosa atas segala doa, dukungan moral dan materil serta kasih sayangnya.
Semoga karya ilmiah ini dapat bermanfaat bagi pengembangan ilmu pengetahuan.
DAFTAR ISI
DAFTAR TABEL vii
DAFTAR GAMBAR vii
PENDAHULUAN 1
Latar Belakang 1
Perumusan Masalah 3
Tujuan Penelitian 3
Manfaat Penelitian 3
TINJAUAN PUSTAKA 4
Botani Tanaman Pisang (Musa spp.) 4
Penyakit Darah (Blood Disease) pada Tanaman Pisang 4
Variasi Somaklonal Kultur Jaringan 5
Pisang Kepok Unti Sayang (US-1) 6
Bulk Segregation Analysis (BSA) 8
Penanda Molekuler Random Amplified polymorphic DNA (RAPD) dan
Inter Simple Sequence Repeat (ISSR) 9
Gen Pembungaan 10
METODE PENELITIAN 11
Waktu dan Tempat Penelitian 11
Bahan Penelitian 11
Identifikasi Morfologi 12
Isolasi DNA 12
Teknik PCR 13
Analisis Sekuen DNA 13
HASIL DAN PEMBAHASAN 14
Karakter Morfologi Mutan Pisang Kepok Unti Sayang 14 Marka Molekular RAPD dan ISSR Terpaut Sifat Male Budless 21 Analisis Marka Molekular dan Sekuen DNA dari Primer Gen
Pistillata (PI) dan Agamous (AG) 24
SIMPULAN 29
UCAPAN TERIMA KASIH 29
DAFTAR PUSTAKA 29
LAMPIRAN 33
DAFTAR TABEL
1 Deskripsi pisang kepok varietas Unti Sayang (US-1) 7 2 Primer Random Amplified Polymorphism DNA (RAPD) yang digunakan
untuk amplifikasi PCR 11
3 Primer Inter Simple Sequence Repeat (ISSR) yang digunakan untuk
amplifikasi PCR 12
4 Primer yang digunakan untuk amplifikasi gen PI dan AG 12 5 Karakter morfologi male bud pada mutan pisang kepok Unti Sayang
(US-1) 16
6 Karakter morfologi pada mutan pisang kepok Unti Sayang (US-1) male budless hasil perbanyakan kultur jaringan sub kultur ke-1, ke-4
dan revertrant dari sub kultur ke-6 dan anakan 16
7 Total pita yang dihasilkan dari 20 primer RAPD 21 8 Total pita yang dihasilkan dari 12 primer ISSR 22 9 Homologi antara sekuen fragmen DNA gen Pistillata tanaman pisang
male budless mutant dengan aksesi-aksesi yang telah dideposit pada
pangkalan data dari bank gen NCBI 24
DAFTAR GAMBAR
1 Bercak coklat pada beberapa bagian. (A) bagian pembungaan, (B) daging buah, (C) jaringan pembuluh kulit buah, (D) pseudostem, (E) jaringan pembuluh tengah peduncula, (F) jaringan pembuluh tepian tengah peduncula, (G) jaringan pembuluh tepian dasar peduncula, (H) pelepah, (I) permukaan tangkai daun, (J) jaringan pembuluh petiole,
(K) tepian helaian daun menguning, (L) akar 5
2 Pisang kepok Unti Sayang (US-1) male budless. (A) munculnya bunga, (B) munculnya buah pisang pada setiap tandan, (C) lepasnya braktea seiring dengan munculnya buah pada tiap tandan, (D) buah pisang terbentuk sampai pada bagian ujung tanpa menyisakan male
bud 6
3 ABC model dan identitas organ pembungaan pada mutan Arabidopsis
(Robles & Pelaz 2005) 10
4 Pisang kepok Unti Sayang (US-1). 1=Male budless mutant; 2=Revertrant mutant. (A) munculnya jantung, (B) terbukanya braktea, (C) jatuhnya braktea, (D1) tidak ada male bud yang tersisa, (D2) male
bud tersisa pada bagian ujung 14
5 Morfologi batang semu. (A) ketegaran batang, (B) bercak pada batang semu, (C) warna bagian dalam batang semu, (D) warna bercak
DAFTAR GAMBAR (lanjutan)
6 Morfologi tangkai dan helai daun. (A) bentuk pangkal helai daun, (B) bentuk ujung daun, (C) warna permukaan atas daun, (D) warna permukaan bawah daun, (E) warna tepian daun, (F) warna permukaan bawah tulang daun, (G) bentuk kanal tangkai daun, (H) bentuk
tangkai daun dekat pelepah 18
7 Morfologi struktur bunga betina dan buah. (A) bentuk bunga, (B) warna tepal lekat, (C) bentuk tangkai putik, (D) warna tepal lepas, (E) bentuk anter, (F) sisa tangkai bunga pada buah, (G) bentuk buah, (H)
susunan bakal biji 19
8 Pola pita ISSR yang dihasilkan oleh primer PKBT-2 pada 24 tanaman pisang hasil kultur jaringan sub kultur 1-6. M=marker (1 kb DNA ladder, Promega), 1-3=sub kultur ke-1, 4-5=sub kultur ke-2, 6-10=sub kultur ke-3, 11-14=sub kultur ke-4, 15-21=sub kultur ke-5, 22-24=sub
kultur ke-6 22
9 Pola pita DNA hasil amplifikasi dengan primer RAPD. (A) OPA 2, (B) OPA 12 dan OPA 13, (C) OPA 16, (D) OPJ 9 dan OPJ 16. M=marker (1 Kb DNA ladder, Promega), WT=tipe liar, S1=sub kultur ke-1 male budless mutant, S6=sub kultur ke- 6 revertrant mutant,
A=anakan revertrant mutant 23
10 Pola pita DNA hasil amplifikasi dengan primer ISSR. (A) PKBT 2, (B) PKBT 11, (C) PKBT 9, (D) PKBT 8, (E) PKBT 6. M=marker (1 Kb DNA ladder, Promega), WT=tipe liar, S1=sub kultur ke-1 male budless mutant, S6=sub kultur ke- 6 revertrant mutant, A=anakan
revertrant mutant 23
11 Amplifikasi primer spesifik. (A) Amplifikasi primer dari gen PI, (B) amplifikasi primer dari gen AG. M=marker (1 Kb DNA ladder, Promega), WT=tipe liar, S1=sub kultur ke-1 male budless, S6=sub
kultur ke-6 revertrant, A=anakan revertrant 25
12 Contoh bagian kromatogram hasil sekuensing DNA male budless
mutant sub kultur ke-1 dengan primer gen PI reverse 26 13 Contoh bagian kromatogram hasil sekuensing DNA male budless
mutant sub kultur ke-1 dengan primer gen AG reverse 26 14 Hasil penyejajaran sekuen DNA dengan primer gen PI pada tanaman
pisang kepok kuning (wild), male budless mutant dan revertrant mutant. Simbol (.)=basa DNA yang sama, simbol (_)=basa DNA yang
DAFTAR LAMPIRAN
1 Kromatogram sekuen gen Pistillata forward pada sampel tanaman
pisang kepok wild type 33
2 Kromatogram sekuen gen Pistillata reverse pada sampel tanaman
pisang kepok wild type 34
3 Kromatogram sekuen gen Pistillata forward pada sampel tanaman
pisang kepok male budless 35
4 Kromatogram sekuen gen Pistillata reverse pada sampel tanaman
pisang kepok male budless 37
5 Kromatogram sekuen gen Pistillata forward pada sampel tanaman
pisang kepok revertrant 38
6 Kromatogram sekuen gen Pistillata reverse pada sampel tanaman
1
PENDAHULUAN
Latar Belakang
Pisang (Musa spp.) merupakan buah yang banyak dikonsumsi dan mendapat prioritas pemerintah untuk dikembangkan dalam memenuhi kebutuhan konsumsi dalam maupun luar negeri. Salah satu kendala dalam pengembangan tanaman pisang adalah adanya serangan penyakit darah (blood disease) yang disebabkan oleh Blood Disease Bacterium (BDB). Nama latin dari bakteri BDB masih belum ada kesepakatan, sering disebut Pseudomonas celebensis Gauman 1923 (Hadiwiyono 2011). Penyakit darah ditularkan oleh serangga pengunjung bunga yang membawa BDB, menginfeksi kumpulan bunga jantan (male bud) melalui celah ketika bunga rontok dan menyebar kesemua bagian tanaman (Rustam 2005). Penyakit BDB lebih banyak menyerang pisang olahan (cooking banana) seperti pisang kepok yang memiliki genom ABB. Pisang ini mudah terserang BDB karena male bud memiliki kandungan gula yang tinggi sehingga menjadi sumber makanan bagi serangga pengunjung bunga. Gejala penyakit darah yang terlihat adalah apabila buah dipotong maka bagian dalam buah kelihatan berwarna merah kecokelatan atau menjadi busuk berlendir (Hadiwiyono 2011).
Penyakit darah pertama kali dilaporkan sejak 80 tahun yang lalu di Sulawesi. Pada tahun 1980 menyebar dengan cepat di Jawa. Akhir tahun 1990, menyebabkan penurunan produksi pisang Kalimantan secara drastis dan sekarang menjadi masalah di Sumatera. Serangan penyakit ini menurunkan produksi buah pisang sebesar 140.135 ton, yaitu dari 5.177.607 ton di tahun 2005 menjadi 5.037.472 ton di tahun 2008 (Deptan 2010). Penelitian sebelumnya (Rustam 2005) mengupayakan pengendalian penyakit ini dengan menggunakan agens antagonis, khususnya kelompok bakteri yang mengoloni perakaran tanaman (rizobakteria) sehingga memacu pertumbuhan tanaman dan menginduksi ketahanan tanaman terhadap patogen.
2
diketahui setelah tanaman tersebut berbunga yaitu selama 1 sampai 2 tahun. Jaligot et al. (2000) menemukan bahwa abnormalitas terjadi pada rata-rata 5 sampai 10% dari populasi bibit kelapa sawit hasil kultur jaringan dan abnormalitas tersebut bersifat epigenetik. Mutasi balik yang diperkirakan terjadi dapat menimbulkan genotipe yang bervariasi untuk sifat tertentu, tetapi sifat lainnya tidak berubah dan berpotensi sebagai Near Isogenic Lines (NIL).
Untuk mengembangkan teknik kultur jaringan sebagai alat perbanyakan klonal tanaman pisang kepok Unti Sayang (US-1) diperlukan suatu teknik yang mampu mendeteksi adanya abnormalitas, diantaranya pada tingkat morfologi dan DNA. Informasi tersebut dapat digunakan untuk menentukan banyaknya sub kultur yang dapat digunakan untuk perbanyakan pisang varietas kepok US-1, sehingga ada jaminan jika mutan ini diperbanyak maka tidak menghasilkan revertrant mutant yang bermanfaat untuk usaha pemuliaan tanaman. Identifikasi melalui pendekatan morfologi dilakukan untuk melihat apakah ada variasi lain yang timbul akibat munculnya revertrant mutant, akan tetapi pendekatan morfologi mudah berubah akibat pengaruh lingkungan sehingga digunakan juga analisis molekuler menggunakan marka genetik. Identifikasi molekuler berdasarkan pola pita DNA digunakan karena karakter ini didasarkan pada sifat genetik tanaman dan tidak dipengaruhi oleh faktor lingkungan.
Marka molekuler yang digunakan adalah Random Amplified Polimorphic DNA (RAPD) dan Inter Simple Sequence Repeats (ISSR). Teknik RAPD telah banyak digunakan untuk menganalisis hasil improvement pisang dan analisis kelompok genom pisang (Crouch et al. 1998) serta analisis integritas genetik planlet pisang dari kultur in vitro (Mathius & Hutabarat 1997). RAPD adalah teknik amplifikasi fragmen DNA dengan menggunakan primer pendek yang panjangnya 10-20 pb. Dengan menggunakan teknik ini polimorfisme antar genotipe dapat terdeteksi jika ada mutasi titik atau inversi pada tempat melekatnya primer dan adanya penyisipan atau delesi dalam satu fragmen amplifikasi. ISSR melibatkan amplifikasi segmen DNA yang berada pada jarak yang dapat teramplifikasi antara dua daerah mikrosatelit berulang yang identik tetapi dengan orientasi arah yang berbeda (Narayanan et al. 2007). ISSR terdistribusi secara melimpah dan merata dalam genom dengan target multiple-locus genomik, variabilitasnya sangat tinggi dengan panjang primer (16-25 pb). Lokasi ISSR pada genom juga dapat diketahui dengan ketepatan yang sangat tinggi sehingga akurat untuk membedakan genotipe (Azrai 2005). Umumnya metode ini digunakan untuk mengetahui adanya penyimpangan genetik, hubungan kedekatan secara genetik ataupun variasi genetik yang ada (Kumar et al. 2006).
3
Perumusan Masalah
Permasalahan utama yang sering dihadapi dalam pengembangan tanaman pisang kepok adalah adanya serangan dari penyakit darah yang disebabkan oleh Blood Disease Bacterium (BDB). Bakteri ini ditularkan melalui perantaraan serangga pengunjung bunga yang mencari nektar. Salah satu cara yang dapat dilakukan untuk mencegah penularan penyakit ini adalah dengan menghasilkan tanaman pisang mutan yang bersifat male budless. Mutan ini telah diperbanyak dengan kultur jaringan, akan tetapi stabilitas genetik dari mutan ini belum diketahui dengan baik. Tingginya sub kultur hingga sub kultur ke-6 diduga dapat menyebabkan munculnya revertrant mutant sehingga mempengaruhi karakter morfologi dari tanaman tersebut. Oleh karena itu, diperlukan suatu teknik yang dapat mendeteksi adanya abnormalitas dengan cara mengembangkan marka molekular yang terpaut dengan sifat male budless.
Tujuan Penelitian
Penelitian ini bertujuan untuk (1) mengidentifikasi karakter morfologi, (2) mengidentifikasi marka ISSR dan RAPD, dan (3) mengidentifikasi marka molekuler yang terpaut dengan sifat male budless pada mutan pisang kepok Unti Sayang (US-1) yang dikembangkan dari gen Pistillata (PI) dan Agamous (AG).
Manfaat Penelitian
4
TINJAUAN PUSTAKA
Botani Tanaman Pisang (Musa spp.)
Tanaman pisang termasuk kedalam tanaman monokotil, berbentuk pohon yang tersusun atas batang semu (pseudostem). Batang semu merupakan tumpukan pelepah daun yang tersusun secara rapat teratur. Percabaangan tanaman bertipe simpodial dengan meristem ujung memanjang dan membentuk bunga lalu buah. Bagian bawah batang menggembung berupa umbi yang disebut bonggol. Tanaman pisang mempunyai bonggol yang dapat membentuk 2-5 mata tunas. Anakan (suckers) muncul dari mata tunas pada bonggol yang selanjutnya tumbuh menjadi tanaman pisang. Daun yang paling muda terbentuk dibagian tengah tanaman, keluarnya menggulung dan terus tumbuh memanjang, kemudian secara progresif membuka. Helaian daun bentuknya lanset memanjang, mudah koyak, permukaan bawah daun berlilin, tulang tengah penopang daun dapat terlihat dengan jelas, pertulangan daun tersusun sejajar dan berwarna hijau.
Tunas pucuk terletak pada ujung atas tanaman dan tumbuh dari bonggol ditanah. Ketika tunas dewasa, bentuk tunas pucuk berubah menjadi struktur bunga semu dan muncul dipermukaan sebagai sebuah bunga raksasa. Pada saat pembungaan dan produksi buah selesai, tunas mati karena pucuk telah digunakan secara maksimal, tetapi tunas baru akan segera terbentuk pada bonggol ditanah. Bunga pada tanaman pisang merupakan bunga majemuk, tiap kuncup bunga dibungkus oleh seludang berwarna merah kecoklatan (braktea). Seludang akan lepas dan jatuh jika bunga telah membuka. Bunga betina akan berkembang secara normal, sedangkan kumpulan bunga jantan (male bud) yang berada diujung tandan tidak berkembang dan tetap tertutup oleh seludang dan biasa disebut sebagai jantung (Ortiz & Vuylsteke 1995).
Tanaman pisang memiliki sifat partenokarpi, yaitu buah bisa terbentuk tanpa adanya fertilisasi pollen (Simmonds & Shepherd 1995). Sebagian besar kultivar banana dan plantain mempunyai kromosom triploid (2n=3x=33) (Retnoningsih et al. 2010). Ovul yang ada pada ovari akan mendukung pembentukan buah. daging buah (pulp) dihasilkan oleh sebagian besar jaringan ovari, khususnya dari permukaan dalam kulit, sebagian besar lagi dari septa dan aksis bagian tengah (Rohmah 2004).
Penyakit Darah (Blood Disease) pada Tanaman Pisang
5
Gambar 1 Bercak coklat pada beberapa bagian. (A) bagian pembungaan, (B) daging buah, (C) jaringan pembuluh kulit buah, (D) pseudostem, (E) jaringan pembuluh tengah peduncula, (F) jaringan pembuluh tepian tengah peduncula, (G) jaringan pembuluh tepian dasar peduncula, (H) pelepah, (I) permukaan tangkai daun, (J) jaringan pembuluh petiole, (K) tepian helaian daun menguning, (L) akar
Menurut Leiwakabessy (1999) patogen dapat ditularkan melalui bibit yang terinfeksi, alat-alat pemangkasan, tanah yang dihanyutkan air, kontak akar dan paling banyak melalui serangga pengunjung bunga (inflorescense). Serangga tersebut terdiri dari beberapa ordo yaitu Diptera (family Chloropidae, Platypezidae, Drosophilidae, Tephritidae, Dolichopodidae, Cilicidae, Calliphoridae, Anthomylidae dan Muscidae); ordo Lepidoptera (family Coleophoridae); ordo Hymenoptera (family Apidae); ordo Blattaria (family Blattidae).
Variasi Somaklonal Kultur Jaringan
6
Variasi somaklonal dapat terjadi akibat adanya induksi selama proses kultur. Faktor-faktor penginduksi tersebut seperti penggunaan berbagai macam zat pengatur tumbuh, perlakuan tingkat konsentrasi zat pengatur tumbuh, lama fase pertumbuhan kalus dalam media, umur kultur, frekuensi sub kultur, tipe kultur yang digunakan (sel, protoplasma, kalus, jaringan), klon atau kultivar tanaman yang digunakan, sumber eksplan yang digunakan, kondisi fisik selama kultur serta digunakan atau tidaknya media seleksi dalam kultur in vitro. Beberapa hasil penelitian menunjukkan bahwa frekuensi sub kultur yang terlalu tinggi serta penggunaan zat pengatur tumbuh dalam konsentrasi yang tinggi diketahui dapat menyebabkan terjadinya variasi somaklonal. Penggunaan zat pengatur tumbuh kelompok auksin sinetik 2,4-D dan 2,4,5-T biasanya dapat menyebabkan terjadinya variasi somaklonal. Pada tanaman kelapa sawit, perlakuan 2,4-D pada kultur kalus yang mampu beregenerasi membentuk tunas diketahui dapat menyebabkan variasi somaklonal saat aklimatisasi di lapangan (Linacero & Vazquez 1992).
Pisang Kepok Unti Sayang (US-1)
Pisang kepok Unti Sayang (US-1) merupakan pisang mutan yang tidak memiliki kumpulan bunga jantan (male budless). Pisang ini pertama kali ditemukan di pulau Selayar, Sulawesi pada tahun 1992 (Gambar 2). Karakteristik pisang mutan ini adalah bersifat male budless sehingga terhindar dari penyakit darah (blood disease). Selain itu, pisang ini memiliki buah dengan kulit yang halus, warna menarik, ukuran lebih besar, warna daging buah kuning, memiliki rasa manis 20,2 sampai 23,8 briks, produktivitas tinggi (lebih dari 40 kg per tandan), penampilan buah menarik dan buah tidak mudah lepas dari sisirnya serta memiliki perakaran yang kuat sehingga tahan rebah (Sobir et al. 2008).
Gambar 2 Pisang kepok Unti Sayang (US-1) male budless. (A) munculnya bunga, (B) munculnya buah pisang pada setiap tandan, (C) lepasnya braktea seiring dengan munculnya buah pada tiap tandan, (D) buah pisang terbentuk sampai pada bagian ujung tanpa menyisakan male bud
7 Tabel 1 Deskripsi pisang kepok varietas Unti Sayang (US-1)
No Karakter Hasil
1 Asal tanaman Desa Bontobangun, Kecamatan Bontoharu, Kabupaten Kepulauan Selayar, Sulawesi Selatan
2 Umur tanaman 17 bulan
3 Umur berbunga (dari bibit anakan) 12-13 bulan 4 Umur panen (dari bibit anakan) 16-17 bulan
5 Nama Unti Sayang
6 Nama local Loka Nipah
7 Propinsi Sulawesi Selatan
8 pH tanah 6,2
15 Warna batang semu Hijau-kuning
16 Warna batang bagian dalam Hijau terang 17 Pigmentasi batang dalam Hijau kekuningan 18 Bentuk penampang batang Bulat
19 Tekstur kulit batang semu Halus 20 Lilin pada batang semu Berlilin 21 Pigmentasi merah dipangkal batang Ada
22 Warna getah Bening
23 Warna pangkal batang Hijau kemerahan 24 Tipe pertumbuhan daun Tegak
25 Warna permukaan atas daun Hijau tua 26 Warna Permukaan bawah daun Hijau muda
27 Bentuk daun Jorong memanjang dengan ujung tumpul
28 Bentuk ujung daun Tumpul
29 Tepi daun Rata
30 Warna tulang daun Hijau kekuningan sedikit bintik ungu 31 Warna bibir pelepah daun Merah keunguan
32 Tipe daun/letak daun Tegak
33 Jumlah daun <20
34 Panjang helai daun 258-282
35 Lebar helai daun 68-86
36 Rasio panjang lebar 3
37 Panjang tangkai daun 51-70
38 Lebar tepi tangkai daun >1 cm 39 Lebar bibir tangkai daun Sedikit sekali 40 Bercak dipangkal tangkai daun Jarang
41 Banyak bercak Sedikit
42 Warna bercak Hitam-ungu
43 Pigmentasi tepi tangkai daun Ada 44 Pigmentasi tulang daun Ada
45 Tipe lekuk tangkai daun Lurus dengan tepi tegak
46 Tipe sayap Kering
47 Warna tepi tangkai daun Hijau
48 Ujung tepi tangkai daun Tidak berwarna 49 Titik pangkal helai daun Simetris 50 Bentuk pangkal helai daun Hati
51 Perkembangan anakan ¾ sama dengan induk
52 Posisi tandan buah Vertical
8
Tabel 1 (Lanjutan)
No Karakter Hasil
56 Pola pelepasan brakrea Tidak menggulung
57 Tekstur braktea Berombak tumpul
58 Warna luar braktea Ungu kecoklatan 59 Warna dalam braktea Merah keunguan 60 Warna ujung luar braktea Kuning
61 Warna pangkal dalam braktea Warna berubah 62 Lapisan lilin pada braktea Ada
63 Panjang tangkai bunga (tandan) 36 cm
64 Jumlah buku kosong 2
65 Diameter tangkai bunga 5-6 cm 66 Warna tangkai bunga Hijau muda 67 Umur berbunga hingga panen 4-5 bulan
68 Umur panen 120-150 hari
69 Posisi tandan Sedikit menyudut
70 Bentuk tandan Silinder
71 Kenampakan tandan Kompak
72 Panjang tandan 96-110 cm
73 Diameter tandan 34-45 cm
74 Jumlah sisir/tandan 13-15 sisir
75 Jumlah buah/sisir 20-22
76 Jumlah baris/sisir 2 baris
77 Bobot tandan 40-41 kg
78 Arah puntiran daun Memuntir berlawanan putaran jam 79 Panjang buah dari sisir pangkal 15-16 cm
80 Panjang buah dari sisir tengah 14-15 81 Panjang buah dari sisir ujung 10-11
82 Diameter buah 4,1-4,5 cm
83 Bobot/buah 105-158 gr
84 Jumlah segi buah 5
85 Bentuk buah Lurus
86 Bentuk melintang Tonjolan jelas
87 Ujung buah Leher botol
88 Sisa stylus Masih ada
89 Panjang tangkai buah 3-4 cm
90 Diameter tangkai buah 1,5-1,7
91 Fusi tangkai buah Tidak ada
92 Warna kulit buah sebelum masak Hijau 93 Warna kulit bua masak Kuning
94 Tebal kulit 2-2,5 mm
95 Tekstur kulit hijau Halus
96 Tekstur kulit matang Halus
97 Kemudahan mengupas Mudah
98 Warna daging mentah Putih kekuningan
99 Warna daging masak Kuning
100 Brix 20,29o-23,8o brix
101 Kandungan vitamin C 3,5-7,2 mg/100g
Bulk Segregation Analysis (BSA)
9 menemukan penanda yang terpaut dengan gen tertentu. Metode ini akan menghasilkan suatu fenotipe atau genotipe tertentu, dimana akan muncul suatu alel yang spesifik yang tidak muncul pada bulk lainnya.
Penanda Molekuler Random Amplified polymorphic DNA (RAPD)dan Inter Simple Sequence Repeat (ISSR)
Teknik RAPD merupakan metode analisis DNA genom dengan cara melihat pola pita DNA yang dihasilkan setelah DNA genom diamplifikasi menggunakan primer acak. RAPD menggunakan primer tunggal atau sekuen nukleotida pendek (10 sampai 20 pb) yang susunan basanya dibuat secara acak dengan kandungan molekul GC minimal 50%. Primer ini mempunyai banyak kelebihan, diantaranya adalah tidak dipengaruhi oleh lingkungan, membutuhkan sedikit DNA dengan kemurnian yang tidak terlalu tinggi, prosedurnya sederhana tanpa radioaktif serta dapat mendeteksi perubahan basa tunggal dalam DNA genom jika cukup banyak primer yang digunakan. Primer tersebut akan berpasangan dengan utas tunggal DNA genom yang satu dan pada utas DNA pasangannya dengan orientasi yang berlawanan. Selama situs penempelan primer masih berada dalam jarak yang dapat diamplifikasi, maka akan diperoleh produk DNA amplifikasi. Jarak tersebut umumnya tidak lebih dari 5000 pb. Deng (1995), menggunakan teknik RAPD dan hasil yang diperoleh teknik tersebut mampu membedakan mutan dan tanaman normal dalam satu genotipe lemon. Rata-rata jumlah fragmen DNA yang dihasilkan sebuah primer tunggal tergantung pada kekomplekan genom. Makin kompleks suatu genom akan makin kompleks pola fragmen DNA yang dihasilkan dari teknik RAPD. Jumlah dan kualitas fragmen DNA yang dihasilkan bergantung pada panjang dan komposisi nukleotida penyusun primer, konsentrasi dan kemurnian DNA cetakan, dan suhu penempelan pada reaksi PCR.
10
Gen Pembungaan
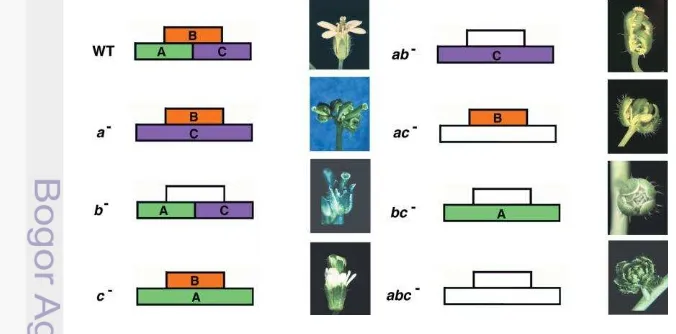
Gen-gen dalam pengaturan proses pembungaan tergolong kedalam kelompok gen MADS-box yang berperan sebagai identitas organ-organ bunga. Gen pembungaan berfungsi dari mulainya fase awal pembentukan identitas pembungaan hingga pembentukan bunga. Bunga terdiri dari empat organ dasar yaitu sepal, petal, stamen dan carpels yang dapat dijelaskan dengan model ABC. Model ini pertama kali ditemukan oleh E. Coen dan Elliot Mayerowitz pada tahun 1991. Ekspresi dari gen A berfungsi terhadap pembentukan sepal dan petals, gen B berfungsi terhadap pembentukan petals dan stamens dan gen C berfungsi terhadap pembentukan stamens dan carpels. Pada tanaman model Arabidopsis aktivitas dari fungsi gen A diatur oleh APETALA1 (AP1) dan (AP2), fungsi gen B diatur oleh APETALA3 (AP3) dan PISTILLATA (PI) bunga dan gen C yang diatur oleh AGAMOUS (AG) selama masa perkembangan (Robles & Pelaz 2005).
Berdasarkan hubungan antara masing-masing gen ABC diperoleh tiga bentuk fenotipik bunga yaitu, (1) apabila gen pembungaan yang bekerja hanya gen A dan B saja maka bunga yang terbentuk hanya memiliki sepal dan petal saja, sehingga bunga tersebut tidak memiliki stamen dan carpel, (2) bila hanya gen B dan C saja yang bekerja maka bunga yang terbentuk hanya memiliki stamen dan carpel saja dan tidak memiliki sepal dan petal, (3) apabila gen yang bekerja hanya gen A dan C maka bunga yang terbentuk hanya memilii sepal dan carpel saja dan tidak memiliki petal dan stamen (Gambar 3). Semakin pesatnya penelitian gen pembungaan pada tanaman, maka model gen ABC berkembang menjadi model ABCDE, yang disebut dengan model MADS-box. Tipe D dibutuhkan untuk perkembangan bakal buah (ovule), dan tipe E dibutuhkan untuk perkembangan seluruh organ bunga dari tanaman. Tipe A terdiri dari APETALLA1 (AP1), tipe B terdiri dari kelas gen APETALLA3 (AP3) dan PISTILLATA (PI), tipe C terdiri dari AGAMOUS (AG), tipe D terdiri dari SEEDSTICK (STK), SHATERPROOF1 (SHP1) dan SHP2, tipe E terdiri dari SEPALLATA1 (SEP1), SEP2, SEP3, SEP4.
11
METODE PENELITIAN
Waktu dan Tempat Penelitian
Penelitian dilakukan dari bulan Agustus 2012 sampai Mei 2013. Identifikasi morfologi dilaksanakan di Kebun Koleksi PKHT Ciomas, Bogor. Analisis molekuler dilaksanakan di Laboratorium Molekuler PKHT.
Bahan Penelitian
Bahan tanaman yang digunakan adalah mutan pisang kepok Unti Sayang yang tidak memiliki kumpulan bunga jantan (Male budless mutant). Jumlah bahan tanaman sebanyak 24 tanaman dari hasil sub kultur ke-1 sampai ke-6. Jumlah tiap sub kultur adalah sub kultur ke-1 sebanyak 3 tanaman, sub kultur ke-2 sebanyak 2 tanaman, sub kultur ke-3 sebanyak 5 tanaman, sub kultur ke-4 sebanyak 4 tanaman, sub kultur ke-5 sebanyak 7 tanaman dan sub kultur ke-6 sebanyak 3 tanaman, serta satu tanaman mutan yang kembali memiliki kumpulan bunga jantan (revertrant mutant) dari perbanyakan anakan dan tipe liarnya yaitu pisang kepok kuning.
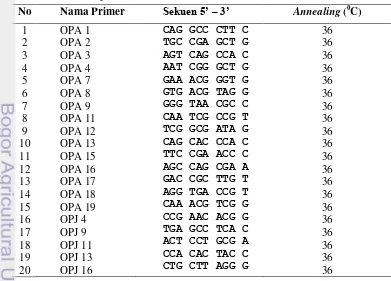
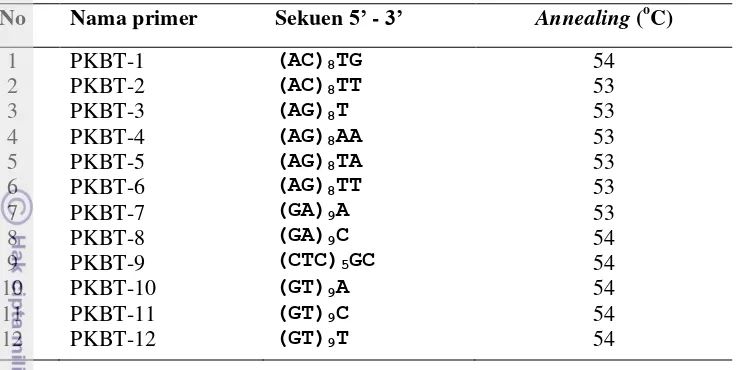
Primer yang digunakan sebanyak 32 primer yang diperoleh dari PKHT (Tabel 2 dan Tabel 3), yang terdiri dari 20 primer RAPD dan 12 primer ISSR. Dua pasang primer yang dikembangkan dari sekuen gen Pistillata (PI) dan Agamous (AG) dari Musa acuminata menggunakan program primer3 (http://frodo.mit.edu) juga digunakan dalam penelitian ini (Tabel 4)
12
Tabel 4 Primer yang digunakan untuk amplifikasi gen PI dan AG
No Primer Sekuen primer
Identifikasi morfologi menggunakan panduan deskriptor pisang dari International Plant Genetik Resources Institute (IPGRI 1996).
Isolasi DNA
Isolasi DNA mengikuti prosedur CTAB (Doyle & Doyle 1987) dengan modifikasi, yaitu menaikkan konsentrasi CTAB dari 2% menjadi 10% dan dengan penambahan polyvinil phenolphthalaein (PVP) dan -mercaptanol pada saat awal penggerusan (Couch & Fritz 1990). Daun yang digunakan adalah daun muda yang masih menggulung yang kemudian digerus didalam buffer CTAB. Purifikasi DNA dilakukan dengan penambahan campuran Chloroform:Isoamil Alkohol (24:1
v/v) sebanyak 500 μl yang kemudian disentrifugasi pada 11000 rpm selama 10
13 air bebas ion steril sebagai stok DNA. Pemurnian DNA dapat dilakukan dengan menambahkan 5μl RNAse (10 mg/ml) dan diinkubasikan pada suhu 370C selama 1 sampai 2 jam. Penentuan konsentrasi DNA dilakukan dengan elektroforesis (Takara USA) yang dimigrasikan bersamaan dengan DNA standar (λ) (Promega USA) 10 dan 50 ng/ml pada gel agarosa 1.2% dalam buffer TBE 1 X dengan tegangan 50 volt selama 15 menit.
Teknik PCR
Amplifikasi PCR mengikuti metode Kurokawa et al. (2003). Seleksi primer dilakukan dengan mengunakan metode Bulk Segregant Analysis (BSA) (Michelmore et al. 1991.) Amplifikasi dilakukan dengan menggunakan mesin PCR (Applied Biosystem USA) sebanyak 35 siklus setelah pra-denaturasi selama 4 menit 940C. Setiap siklus terdiri atas 30 detik 940C denaturasi, 30 detik annealing (36-650C), 1 menit pada 720C elongasi dan elongasi akhir 5 menit pada 720C. Fragmen DNA hasil amplifikasi kemudian dielektroforesis selama 47 menit pada tegangan 50 volt bersama DNA standar 1 kb DNA ladder (Promega USA) pada gel agarosa 1.2%. Gel kemudian divisualisasi dengan cara direndam dalam ethidium bromida 10% selama 15 menit dan dibilas dengan akuades. Pola pita hasil amplifikasi kemudian didokumentasi menggunakan alat dokumentasi gel.
Analisis Sekuen DNA
14
HASIL DAN PEMBAHASAN
Karakter Morfologi Mutan Pisang Kepok Unti Sayang
Identifikasi morfologi dilakukan pada 24 tanaman pisang kepok Unti Sayang (US-1) male budless mutant yang telah diperbanyak dengan kultur jaringan sub kultur ke-1 sampai ke-6. Dari tanaman yang telah diperbanyak hanya beberapa tanaman saja yang telah berbunga dan berbuah yaitu dari sub kultur ke-1 sebanyak dua tanaman, sub kultur ke-4 sebanyak satu tanaman dan sub kultur ke-6 sebanyak satu tanaman. Selain itu, juga digunakan male budless mutant dari perbanyakan anakan serta pisang kepok kuning male bud yang juga telah berbunga dan berbuah sebagai tipe liar. Hasil pengamatan pada beberapa tanaman menunjukkan bahwa terdapat tanaman yang memunculkan fenotipe kembali memiliki kumpulan bunga jantan (revertrant mutant), yaitu dari perbanyakan sub kultur ke-6 dan juga dari perbanyakan anakan (Gambar 4).
Gambar 4 Pisang kepok Unti Sayang (US-1). 1=Male budless mutant; 2=Revertrant mutant. (A) munculnya jantung, (B) terbukanya braktea, (C) jatuhnya braktea, (D1) tidak ada male bud yang tersisa, (D2) male bud tersisa pada bagian ujung
15 sebanyak 6 sampai 7 sisir. Jumlah buah pada male budless mutant juga lebih banyak daripada revertrant mutant yang menghasilkan jumlah sisir per tandan yang lebih sedikit. Ini menunjukkan bahwa male budless mutant menghasilkan lebih banyak buah daripada pisang yang memiliki male bud (Tabel 5).
Tabel 5 Karakter morfologi male bud pada mutan pisang kepok Unti Sayang (US-1)
No Karakter Kepok kuning
Male budless Revertrant
1 Asal perbanyakan Anakan Sub kultur ke-1 dan ke-4 Sub kultur ke-6 Anakan 2 Panjang tangkai
tandan
± 60 cm ± 63 cm 56 cm 60 cm
3 Panjang tandan ± 100 cm 100-113 cm 110 cm 115 cm
4 Jumlah sisir/tandan 6-7 6-8 5 7
5 Jumlah buah/sisir 13-16 14-16 ≥17 ≥20
6 Jarak antar sisir buah
± 12 cm ± 16 cm 8 cm 12 cm
7 Male bud Ada Tidak ada Ada Ada
8 Bekas braktea
pada gagang bunga
Ada bekas Tidak ada bekas Ada bekas Ada bekas
Identifikasi morfologi juga telah dilakukan oleh Pusat Kajian Hortikultura Tropika (PKHT) pada tanaman male budless mutant lainnya, yaitu pada pisang kepok tanjung. Hasil identifikasi menunjukkan bahwa ada karakter morfologi yang berubah akibat munculnya sifat male budless dan sifat ini berbeda dengan tipe liarnya. Pisang kepok tanjung male budless memiliki bentuk helaian daun yang lebih ramping dan panjang jika dibandingkan dengan tipe liarnya kepok tanjung yang memiliki male bud (Sobir et al. 2008).
16
Tabel 6 Karakter morfologi pada mutan pisang kepok Unti Sayang (US-1) male budless hasil perbanyakan kultur jaringan sub kultur ke-1, ke-4 dan revertrant dari sub kultur ke-6 dan anakan
No Karakter
5 Tipe pertumbuhan daun Tegak Tegak
6 Dwarfisme Normal Normal
7 Tinggi batang semu 2.1-2.9 2.1-2.9
8 Bentuk batang semu Normal Normal
9 Warna batang semu Hijau-merah Hijau-merah
10 Lilin pada batang semu Berlilin Berlilin 11 Warna batang semu bagian dalam Hijau Hijau
12 Pigmentasi batang dalam Merah Merah
13 Warna getah Bening Bening
14 Lilin pada pelepah daun Sedang Sedang
15 Jumlah anakan 2-4 2-4
16 Perkembangan anakan Diantara ¼ dan ¾ dari tinggi induk
Diantara ¼ dan ¾ dari tinggi induk
17 Posisi anakan Dekat dengan induk Dekat dengan induk 18 Bercak dipangkal tangkai daun Luas Luas
19 Warna bercak pada tangkai daun Coklat Coklat 20 Tipe lekuk tangkai daun III Tepi melengkung
kedalam
Tepi melengkung kedalam
21 Tepi tangkai daun Bersayap dan lekat batang
Bersayap dan lekat batang
22 Tipe sayap Kering Kering
23 Warna tepi tangkai daun Pink-ungu sampai merah
Pink-ungu sampai merah
24 Ujung tepi tangkai daun Ada garis berwarna yang panjang
30 Warna permukaan atas daun Hijau tua Hijau tua 31 Penampilan permukaan atas daun Bercahaya Bercahaya 32 Warna permukaan bawah daun Hijau sedang Hijau sedang 33 Penampilan permukaan bawah daun Pudar Pudar
34 Lilin pada daun Sedang Sedang
35 Titik pangkal helai daun Asimetrik Asimetrik 36 Bentuk pangkal daun Kedua sisi bulat Kedua sisi bulat 37 Bengkok pada daun Sedikit membengkok Sedikit membengkok 38 Warna belakang tulang daun Hijau terang Hijau terang
39 Warna depan tulang daun Hijau tua Hijau tua 40 Warna bibir pelepah daun Merah-ungu Merah-ungu 41 Bercak daun pada anakan Tanpa bercak Tanpa bercak
42 Lebar tangkai tandan 7-12 cm 7-12 cm
43 Warna tangkai tandan Hijau Hijau
44 Rambut pada tangkai tandan Tidak berambut Tidak berambut 45 Posisi buah terikat Menggantung vertical Menggantung vertical
17
49 Posisi tandan Menggantung vertical Menggantung vertical
50 Penampilan tandan Botak Botak
52 Bentuk pangkal braktea Bahu kecil Bahu kecil 53 Bentuk ujung braktea Agak meruncing Agak meruncing
54 Pola pelepasan braktea Menggulung Menggulung
55 Warna luar braktea Merah-ungu Merah-ungu
56 Warna dalam braktea Merah Merah
57 Warna ujung luar braktea Kuning Kuning
58 Warna goresan pada braktea Tidak berwarna Tidak berwarna 59 Bekas braktea pada gagang bunga Sangat menonjol Sangat menonjol 60 Kepudaran warna pada pangkal braktea Warna relatif sama Warna relatif sama 61 Bentuk braktea x/y < 0.28 (lancet) x/y < 0.28 (lancet) 62 Lapisan lilin pada braktea Sedang Sedang
63 Lekuk pada braktea Sedikit berlekuk Sedikit berlekuk
64 Susunan bakal biji Empat Empat
65 Bentuk buah Lurus membengkok Lurus membengkok
67 Garis potong buah Runcing Runcing
68 Ujung buah Tumpul Tumpul
69 Sisa tangkai bunga pada ujung buah Ada sisa Ada sisa
70 Panjang tangkai buah ≤10 mm ≤10 mm
71 Lebar tangkai buah < 5 mm < 5 mm
72 Permukaan tangkai buah Tidak berambut Tidak berambut 73 Fusi tangkai buah Fusi sebagian Fusi sebagian 74 Warna kulit buah sebelum masak Hijau terang Hijau terang
75 Warna kulit buah masak Kuning Kuning
76 Tebal kulit buah 2.5 mm 2.5 mm
77 Kemudahan mengupas Mudah dikupas Mudah dikupas
78 Retakan pada kulit buah Tidak retak Tidak retak
Pengamatan morfologi terhadap beberapa karakter batang semu (pseudostem) pada male budless mutant dan revertrant mutant menunjukkan bahwa tidak ada perbedaan karakter batang semu. Semua tanaman memiliki tipe pertumbuhan daun tegak dan memiliki ketegaran batang semu yang termasuk kedalam kategori normal atau kekar (Gambar 5). Kultivar-kultivar yang merupakan turunan dari M. balbisiana yang memiliki genom B pada umumnya memiliki batang semu yang tinggi, kekar dan memiliki daun yang lebih tebal misalnya pisang kepok (ABB) (Simmonds & Shepherd 1995).
18
Gambar 5 Morfologi batang semu. (A) ketegaran batang, (B) bercak pada batang semu, (C) warna bagian dalam batang semu, (D) warna bercak pelepah dekat daun
19 Morfologi struktur bunga dan buah yang diamati juga tidak menunjukkan adanya variasi pada warna tepal lekat, pigmentasi tepal, warna tepal lepas, penampakan tepal lepas, bentuk ujung tepal lepas, perkembangan ujung tepal lepas, warna kepala sari, bentuk tangkai putik, warna kepala putik, dan susunan bakal biji (Gambar 7).
Gambar 7 Morfologi struktur bunga betina dan buah. (A) bentuk bunga, (B) warna tepal lekat, (C) bentuk tangkai putik, (D) warna tepal lepas, (E) bentuk anter, (F) sisa tangkai bunga pada buah, (G) bentuk buah, (H) susunan bakal biji
20
akibat epigenetik (Kaeppler et al. 2000). Variasi somaklonal dapat disebabkan oleh jenis dan konsentrasi zat pengatur tumbuh yang ditambahkan kedalam medium, tipe regenerasi, kultivar atau klon yang digunakan, sumber eksplan, umur kultur, frekuensi sub kultur dan lingkungan kultur (Karp 1995).
Pada penelitian ini, zat pengatur tumbuh yang digunakan adalah dari dua kelompok sitokinin yaitu turunan adenine yaitu BAP dan turunan fenilurea yaitu TDZ. Konsentrasi BAP yang digunakan adalah sebesar 0.08 ppm dan konsentrasi TDZ yang digunakan adalah sebesar 2 ppm. BAP dan TDZ mempunyai respon fisiologi yang sama yaitu berperan dalam regulasi pembelahan sel serta diferensiasi dan pertumbuhan jaringan. Penggunaan zat pengatur tumbuh sitokinin dalam konsentrasi yang tinggi dapat berpengaruh negatif karena dapat meningkatkan frekuensi tanaman regeneran tumbuh abnormal akibat tingginya kecepatan multiplikasi.
Beberapa kultivar dalam satu spesies tanaman juga dapat menunjukkan tingkat variasi yang berbeda ketika diperbanyak secara in vitro. Terdapat kultivar yang menunjukkan variasi yang berlebihan, sedangkan yang lainnya stabil. Pada pisang kepok tanjung menunjukkan perubahan bentuk helaian daun akibat adanya sifat male budless, pada pisang cavendish terjadi abnormalitas berupa kekerdilan yang mencapai 70% dan terus terbawa hingga beberapa siklus perbanyakan vegetatif. Pada mutan pisang kepok Unti Sayang male budless menunjukkan adanya revertrant mutant akan tetapi tidak menyebabkan perubahan pada karakter morfologi yang lain. Munculnya revertrant mutant diduga disebabkan oleh adanya perubahan susunan olionukleotida pada untai DNA yang terjadi secara acak dan berbeda untuk masing-masing tanaman.
Tingginya sub kultur hingga sub kultur ke-6 diduga menyebabkan ketidakseimbangan metabolisme dari tanaman sehingga menginduksi munculnya variasi somaklonal yaitu ditemukannya revertrant mutant. Sub kultur dilakukan untuk meningkatkan kecepatan multiplikasi. Akan tetapi, jika sub kultur dilakukan secara berlebihan maka juga dapat menginduksi variasi (Skirvin et al. 1994). Eksistensi keragaman genetik diduga berhubungan dengan kestabilan jaringan dari genotipe pisang tersebut untuk merespon perlakuan didalam kultur. Sub kultur dapat dilakukan terus menerus tetapi dengan bertambahnya umur kultur maka sub kultur menjadi kurang responsif dan muncul ketidakstabilan genetik, sehingga sub kultur harus dibatasi. Cote et al. (1993) menemukan bahwa pada pisang cavendish sub kultur maksimal dilakukan 10 kali. Variasi yang sering muncul pada kultur in vitro pisang berupa kelainan bentuk, ukuran bunga, perubahan bentuk daun, batang semu, dan warna.
Tregear et al. (2002) juga menemukan adanya abnormalitas yang terjadi pada bibit kelapa sawit hasil kultur jaringan dengan beberapa kali sub kultur, dimana bunga kelapa sawit stamen dan staminodes berubah menjadi struktur buah semu (buah mantel) sehingga menyebabkan sterilitas dan abnormalitas tersebut bersifat epigenetik. Phillips et al. (1994) menemukan bahwa tanaman yang beregenerasi dari kultur kalus yang relatif tidak berdiferensiasi menyebabkan kemungkinan terjadinya perubahan genetik yang sangat besar.
21 dan mutasi. Perbedaan morfologi belum bisa untuk memastikan adanya perubahan secara genetik karena faktor adaptasi sementara biasanya tidak permanen. Oleh karena itu, analisis dengan menggunakan penanda molekular juga digunakan untuk menganalisis pola pita DNA dari semua sampel tanaman baik dari kultur jaringan maupun anakan.
Marka Molekular RAPD dan ISSR Terpaut Sifat Male Budless
Marka molekular digunakan untuk mengetahui kestabilan genetik tanaman hasil regenerasi secara in vitro dan anakan. Pengujian stabilitas genetik tanaman melalui pengujian molekuler terhadap hasil mikropropagasi merupakan tahapan penting dalam produksi benih secara massal, karena permasalahan utama dalam mikropropagasi melalui embriogenesis somatik adalah munculnya variasi somaklonal. Marka molekular digunakan untuk memverifikasi secara genetik serta untuk memperoleh pita DNA pembeda antara genotipe male budless mutant dan revertrant mutant serta membandingkan pola pita DNA tersebut dengan tipe liarnya. Deng et al. 1995 menggunakan penanda RAPD dan dapat membedakan antara genotip lemon mutan dengan normalnya.
Amplifikasi DNA dengan menggunakan 20 primer RAPD menghasilkan 101 pita DNA dengan ukuran berkisar antara 250-1250 pb. Jumlah pita yang dihasilkan per primer bervariasi, mulai dari 2 pita (OPA 9, OPA 12 dan OPJ 16) sampai 9 pita (OPA 2 dan OPA 7) (Tabel 7). Amplifikasi DNA dengan menggunakan 12 primer ISSR menghasilkan 52 pita DNA dengan ukuran berkisar antara 250-1750 pb. Jumlah pita yang dihasilkan per primer juga bervariasi, mulai dari 3 pita (PKBT 5 dan PKBT 11) sampai 7 pita (PKBT 2 dan PKBT 3) (Tabel 8). Semua pita DNA yang dihasilkan merupakan pita monomorfik untuk semua sampel hasil sub kultur serta anakan.
Tabel 7 Total pita yang dihasilkan dari 20 primer RAPD
22
Tabel 8 Total pita yang dihasilkan dari 12 primer ISSR
No Nama Primer Jumlah Pita DNA Ukuran Pita DNA (pb)
Amplifikasi PCR dengan menggunakan primer RAPD dan ISSR dilakukan untuk semua sampel tanaman pisang yang berasal dari hasil kultur jaringan sub kultur ke-1 sampai ke-6 sebanyak 24 tanaman. Berdasarkan pengamatan morfologi ditemukan adanya revertrant mutant, akan tetapi hasil amplifikasi DNA pada semua tanaman baik dari kultur jaringan maupun anakan vegetatif tidak menunjukkan adanya pola pita DNA yang berbeda. Amplifikasi terhadap tanaman yang berasal dari anakan serta tipe liarnya dengan menggunakan primer ISSR juga menghasilkan pola pita DNA monomorfik (Gambar 8).
Gambar 8 Pola pita ISSR yang dihasilkan oleh primer PKBT-2 pada 24 tanaman pisang hasil kultur jaringan sub kultur 1-6. M=marker (1 kb DNA ladder, Promega), 1-3=sub kultur ke-1, 4-5=sub kultur ke-2, 6-10=sub kultur ke-3, 11-14=sub kultur ke-4, 15-21=sub kultur ke-5, 22-24=sub kultur ke-6
23
Gambar 9 Pola pita DNA hasil amplifikasi dengan primer RAPD. (A) OPA 2, (B) OPA 12 dan OPA 13, (C) OPA 16, (D) OPJ 9 dan OPJ 16. M=marker (1 Kb DNA ladder, Promega), WT=tipe liar, S1=sub kultur ke-1 male budless mutant, S6=sub kultur ke- 6 revertrant mutant, A=anakan revertrant mutant
Gambar 10 Pola pita DNA hasil amplifikasi dengan primer ISSR. (A) PKBT 2, (B) PKBT 11, (C) PKBT 9, (D) PKBT 8, (E) PKBT 6. M=marker (1 Kb DNA ladder, Promega), WT=tipe liar, S1=sub kultur ke-1 male budless mutant, S6=sub kultur ke- 6 revertrant mutant, A=anakan revertrant mutant
Primer-primer yang digunakan belum dapat menunjukkan adanya variasi genetik atau belum terpaut dengan sifat male budless. Lakshmanan et al. (2007) juga telah menganalisis kestabilan genetik tanaman pisang dengan menggunakan 30 primer RAPD serta 5 primer ISSR. Hasil yang diperoleh adalah pola pita DNA monomorfik, dimana tidak terdapat pola pita yang berbeda antara tanaman induk dengan tanaman yang diperbanyak dengan kultur jaringan meskipun terdapat variasi morfologi.
24
perubahan fenotipik, dimana ekspresi gen dari perubahan tersebut memberikan implikasi yang cukup besar pada perubahan fenotipiknya, khususnya dalam menentukan muncul atau tidaknya male bud.
Tidak adanya variasi pita DNA yang dihasilkan dari primer erat hubungannya dengan perubahan dalam satu atau dua oligonukleotida pada utas DNA yang belum dapat dideteksi dengan teknik ISSR dan RAPD. Hasil amplifikasi primer RAPD dan ISSR tidak terdeteksi adanya perubahan genetic. Akan tetapi, kemungkinan terdapat beberapa perubahan genetik yang tidak dapat terdeteksi seperti adanya mutasi titik diluar situs penempelan primer. Polimorfisme antar genotipe dapat terdeteksi jika ada mutasi titik atau inversi pada tempat melekatnya primer dan adanya penyisipan atau delesi dalam satu fragmen amplifikasi (Debener 2002).
Mathius et al. (2001) menggunakan penanda RAPD untuk menganalisis genotip normal dan abnormal. Hasil yang diperoleh tidak ada satupun pita DNA dari 15 primer yang mencirikan abnormalitas pada semua klon yang diuji. Teknik RAPD tersebut hanya mampu membedakan antar genotip normal, abnormal dan berbunga jantan dalam klon yang sama, namun tidak ditemukan pita DNA pembeda abnormalitas yang dapat digunakan untuk semua klon. Mathius et al. (2005) juga telah menggunakan 20 primer AFLP untuk menganalisis genotip normal dan abnormal pada klon kelapa sawit. Primer yang digunakan menghasilkan 264 pita DNA, akan tetapi tidak ditemukan pita pembeda spesifik yang dapat membedakan antar genotip normal, abnormal atau berbunga jantan, dimana variasi ini erat hubungannya dengan pola metilasi DNA dan metilasi tersebut bersifat epigenetik.
Pada penelitian ini digunakan sebanyak 20 primer RAPD dan 12 primer ISSR. Beberapa hasil penelitian melaporkan bahwa semakin banyak jumlah primer yang digunakan, semakin rendah nilai koefisien keragaman hasil analisis yang diperoleh. Sepuluh sampai duapuluh primer dianggap sudah mencukupi untuk keperluan analisis karena dengan sepuluh primer pengaruh kesalahan percobaan telah dapat diperkecil hingga mendekati nol (Hallden et al. 1994). Analisis dengan menggunakan 200 karakter RAPD menurut Andebrham & Furtek (1994) telah dapat memberikan hasil analisa yang dapat dipercaya. Dengan mengkombinasikan pengamatan karakter morfologi dan molekuler maka kelemahan masing-masing karakter dapat dieliminasi sehingga didapatkan hasil evaluasi variasi yang kuat.
Analisis Marka Molekuler dan Sekuen DNA dari Primer Gen Pistillata (PI) dan Agamous (AG)
25 organ reproduktif tumbuhan, selain itu adanya gangguan dari mikroorganisme juga dapat menyebabkan ekspresi gen AG2 terganggu.
Verifikasi dilakukan pada tanaman yang menunjukkan adanya variasi morfologi yaitu ada atau tidaknya male bud, yaitu menggunakan tanaman pisang male budless dari sub kultur ke-1 (S1), tanaman pisang revertrant dari sub kultur ke-6 (S6), tanaman pisang revertrant dari anakan vegetatif (A), serta tanaman pembanding yaitu pisang kepok kuning male bud (WT). Dua pasang primer PI dan AG didesain berdasarkan informasi sekuen gen Pistillata dan Agamous dari aksesi Musa yang terdapat pada database GeneBank. Dua pasang primer yang diuji dapat mengamplifikasi semua sampel tanaman, ditunjukkan dengan terbentuknya pita tunggal dengan ukuran antara 500-750 pb (Gambar 11). Ini menunjukkan bahwa semua sampel tanaman yang diuji baik tanaman pisang male bud, male budless dan revertrant memiliki gen PI dan AG, sehingga kemungkinan terdapat gen lain yang mengontrol karakter male budless.
Pita DNA hasil amplifikasi tidak dapat memastikan bahwa semua sampel tanaman memiliki sekuen DNA yang sama, karena perbedaan satu atau beberapa basa seperti Single Nucleotide Polymorphism (SNP) tidak dapat terdeteksi melalui elektroforesis agarosa. Diduga terjadi variasi nukleotida pada tanaman yang diuji. Oleh karena itu, pita hasil amplifikasi tersebut diproses lebih lanjut untuk selanjutnya dilakukan analisis sekuen DNA (sekuensing) tiap sampel tanaman.
Gambar 11 Amplifikasi primer spesifik. (A) Amplifikasi primer dari gen PI, (B) amplifikasi primer dari gen AG. M=marker (1 Kb DNA ladder, Promega), WT=tipe liar, S1=sub kultur ke-1 male budless, S6=sub kultur ke-6 revertrant, A=anakan revertrant
26
Gambar 12 Contoh bagian kromatogram hasil sekuensing DNA male budless mutant sub kultur ke-1 dengan primer gen PI reverse
Hasil sekuensing dengan menggunakan primer dari gen AG menghasilkan kromatogram yang kurang baik karena terlalu banyak noise yang dapat menggangu penentuan posisi basa dari DNA sampel sehingga tidak dapat dilakukan contig atau perbaikan susunan sekuen (Gambar 13). Hasil sekuensing dengan menggunakan primer dari gen AG yang kurang baik diduga disebabkan karena adanya multi template dari sekuen AG. Beberapa hasil penelitian menemukan bahwa gen AG berkorelasi dengan beberapa gen lainnya, seperti gen AG2 dan gen EGAD1 yang diduga berada pada satu ukuran yang sama, sehingga jika ingin memperoleh single template maka tahapan yang bisa dilakukan adalah dengan menggunakan teknik kloning. Oleh karena itu, hanya hasil sekuensing fragmen PCR dengan primer dari gen PI yang digunakan untuk analisis tahap selanjutnya.
Gambar 13 Contoh bagian kromatogram hasil sekuensing DNA male budless mutant sub kultur ke-1 dengan primer gen AG reverse
27 fragmen gen PI asal Musa dengan nomor aksesi: EU869311.1, DQ005604.1, AY941798.1, DQ005603.1, EU433562.1 (Tabel 9).
Tabel 9 Homologi antara sekuen fragmen DNA gen Pistillata tanaman pisang male budless mutant dengan aksesi-aksesi yang telah dideposit pada pangkalan data dari bank gen NCBI
Aksesi
EU869311.1 Musa acuminata AAA Group MADS-box protein MADS6 mRNA, complete cds
118 317 31 100
DQ005604.1 Musa ornata Pistillata-like protein (Pla) mRNA, partial cds
118 317 31 100
AY941798.1 Musa acuminata putative MADS box protein (MADS1) mRNA, partial cds
118 317 31 100
DQ005603.1 Musa ornata Pistillata-like protein (Plb) mRNA, partial cds
104 263 31 94
EU433562.1 Musa basjoo Pistillata-like protein (PI-1) mRNA, partial cds
104 254 31 94
Hasil homologi yang diperoleh menunjukkan nilai query coverage sebesar 31%. Data hasil sekuensing yang diperoleh kemudian dianalisis dengan menggunakan program MEGA 5.0 untuk menentukan apakah antara satu sekuen DNA homolog dengan yang lainnya. Analisis homologi dilakukan dengan membandingkan basa DNA antara sekuen dari male budless mutant dengan revertrant mutan serta tipe liarnya sebagai tanaman kontrol. Tahap ini akan menampilkan setiap basa nukleotida yang terdapat pada setiap sampel DNA yang digunakan. Analisis yang dilakukan setelah penyejajaran sekuen adalah penghitungan perbedaan basa antara kedua sampel tanaman.
Hasil penyejajaran sekuen DNA terlihat bahwa antara male budless mutant dan revertrant mutant memiliki sekuen DNA yang hampir keseluruhannya conserve jika disejajarkan dengan sekuen DNA tipe liarnya, yaitu pisang kepok kuning. Alignment sekuen DNA dengan program MEGA diperoleh adanya variasi nukleotida dari fragmen dengan ukuran 532 pb. Terdapat tiga nukleotida yang berbeda (SNP) pada revertrant mutant, yaitu pada posisi nukleotida ke-445, 461 dan 507. Perbedaan nukleotida male budless mutant dan revertrant mutant terjadi
pada daerah 3’UTR (Untranslated region) (Gambar 14).
Berdasarkan hasil sekuen DNA dari gen PI dapat diketahui bahwa tidak terjadi perubahan pada gen PI, baik pada male budless mutant dan revertrant mutant. Perubahan diduga terjadi pada gen AG yang dalam penelitian ini belum berhasil diketahui. Gen AG adalah keluarga gen MADS-box yang diperlukan dalam pembentukan dan perkembangan stamen (kepala dan benang sari) dan karpel (putik). Adam et al. (2007) menemukan bahwa tingkat ekspresi gen AG dapat ditekan oleh gen lain sehingga pengaruh gen AG dalam pembentukan meristem bunga dan inflorensia menjadi terhambat.
28
perbedaan ekspresi gen PI pada male budless mutant dan revertrant mutant juga diduga terjadi akibat adanya variasi epigenetik. Variasi yang terjadi diduga berhubungan dengan perubahan dalam pola metilasi dari komponen genomik. Hal yang sama juga ditemukan dari hasil penelitian Mathius et al (2001) yang menyatakan bahwa status metilasi dapat dideteksi pada sekuen spesifik namun tidak diperoleh polimorfisme tunggal yang konsisten berbeda antara klon normal dan abnormal pada tanaman kelapa sawit.
Wild CTCTTTCTCT CTCTCTCTAT ATTTTTTATC TTTGCCATTT TTCTTGTTGG [ 50] Budless ... ... ... ... ... [ 50] Revertrant... ... ... ... ... [ 50]
Wild ATTTCGGGGT ATTGACGGTG CTGCTGGTGA ATCGGTGGAA GGTTGCCGAA [100] Budless ... ... ... ... ... [100] Revertrant... ... ... ... ... [100]
Wild AATCTTGGAG AGGTACCAGC AGAACTCCGG GAGGAAGTTC TGGGATGCCA [150] Budless ... ... ... ... ... [150] Revertrant... ... ... ... ... [150]
Wild AGCATGAGGT AGCGCTTCGC AAAATCTCGT CTTTCACGGT GCTTTGTCTT [200] Budless ... ... ... ... ... [200] Revertrant... ... ... ... ... [200]
Wild TAAAAACCTA TGTTTCAAAG TGATTACAAG GCGGAGTTCT CCTTTCGGAA [250] Budless ... ... ... ... ... [250] Revertrant... ... ... ... ... [250]
Wild ATCTCGTCTT GCTCCCAGAA AAGCTTCTTC TTCTTTCCTT TTTGATGAAG [300] Budless ... ... ... ... ... [300] Revertrant... ... ... ... ... [300]
Wild ACAAGTAATA AAAAATGGGA ACTTTAGGGA GTTGCTCACC AGTTTACGGG [350] Budless ... ... ... ... ... [350] Revertrant... ... ... ... ... [350]
Wild TGTGTATGGC AGAGTCTGAG TGCTGAGATC GATCGGATTA AGAAAGAGAA [400] Budless ... ... ... ... ... [400] Revertrant... ... ... ... ... [400]
Wild CGACAACATG CAGATCGAGT TGAGGTATGT TGTGCCATCT CATAGTTTCT [450] Budless ... ... ... ... ... [450] Revertrant... ... ... ... ....T... [450]
Wild CTCAACAGAA GAATTTGGTG GTCTTTTTAT TTTGGCTTTT GTTTGTGGAT [500] Budless ... ... ... ... ... [500] Revertrant... T... ... ... ... [500]
Wild GATTAGGCAT CTGAAGGGGG AGGATCTCAA CT [532] Budless ... ... ... .. [532] Revertrant...T... ... ... .. [532]
Gambar 14 Hasil penyejajaran sekuen DNA dengan primer gen PI pada tanaman pisang kepok kuning (wild), male budless mutant dan revertrant mutant. Simbol (.)=basa DNA yang sama, simbol (_)=basa DNA yang berbeda
29 melibatkan perubahan pada sekuen DNA. Terjadinya proses epigenetik merupakan konsekuensi adanya interaksi antara gen dan lingkungannya dan dapat terjadi akibat tidak terekspresinya suatu gen (silenced genes). Pada male budless mutant diduga terjadi hipermetilasi, yaitu terikatnya metil pada basa sitosin dan apabila terjadi pada daerah promotor maka gen tersebut tidak terekspresi. Sedangkan pada revertrant mutant diduga terjadi hipometilasi, yaitu terlepasnya metil dari basa sitosin dan apabila terjadi pada daerah promotor maka gen-gen akan terekspresi kembali (Fraga & Esteller 2002).
SIMPULAN
Identifikasi karakter morfologi menunjukkan adanya revertrant mutant, yaitu dari perbanyakan sub kultur ke-6 dan anakan. Munculnya revertrant mutant tidak menyebabkan adanya perubahan morfologi lain, kecuali ada atau tidaknya male bud.
Sebanyak 20 primer RAPD menghasilkan sebanyak 101 pita DNA dan 12 primer ISSR menghasilkan sebanyak 52 pita DNA. Pita yang dihasilkan adalah pita DNA monomorfik, baik pada tanaman pisang tipe liar male bud, male budless mutant maupun revertrant mutant.
Amplifikasi menggunakan primer yang dikembangkan dari gen PI dan AG tidak menghasilkan pita DNA polimorfik pada male budless mutant dan revertrant mutant. Ini menunjukkan bahwa tanaman tersebut memiliki gen homeotik Pistillata (PI) dan Agamous (AG), diduga terdapat gen lain yang mengontrol karakter male budless. Analisis sekuen nukleotida dari gen PI ditemukan adanya tiga variasi nukleotida (SNP) pada daerah 3’UTR posisi nukleotida ke-445, 461 dan 507.
UCAPAN TERIMA KASIH
Penelitian ini didanai oleh Pusat Kajian Hortikultura Tropika (PKHT) Institut Pertanian Bogor (IPB) Tahun 2012. Atas dukungan tersebut penulis mengucapkan terimakasih.
DAFTAR PUSTAKA
Adam H, Jouannic S, Orieux Y, Morcillo F, Richaud F, Duval Y, Tregear JW. 2007. Functional characterization of Mads Box genes involved in the determination of oil palm flower structure. J Exp Bot 58:1245-1259.
Andebrhan T, Furtek DB. 1994. Random amplified polymorphic DNA (RAPD) analysis of crinipellis perniciosa isolates from different host. Plant Pathol 43:1020-1027.