TEKNOLOGI SUPLEMENTASI DAN PAKAN KOMPLIT PADA
RANSUM BERBASIS JERAMI PADI TERHADAP POPULASI
BAKTERI DAN PROTOZOA RUMEN
SKRIPSI FATMA SARI
DEPARTEMEN ILMU NUTRISI DAN TEKNOLOGI PAKAN FAKULTAS PETERNAKAN
i RINGKASAN
FATMA SARI. D24070117. 2012. Teknologi Suplementasi dan Pakan Komplit pada Ransum Berbasis Jerami Padi terhadap Populasi Bakteri dan Protozoa Rumen. Skripsi. Departemen Ilmu Nutrisi dan Teknologi Pakan. Fakultas Peternakan. Institut Pertanian Bogor.
Pembimbing Utama : Ir. Anita S. Tjakradidjaja, M.Rur.Sc. Pembimbing Anggota : Dr. Ir. Ibnu Katsir Amrullah, MS
Penelitian ini bertujuan untuk mengetahui pengaruh dari aplikasi teknologi suplementasi dan pakan komplit pada ransum berbasis jerami padi terhadap populasi bakteri dan protozoa rumen. Jerami padi adalah limbah pertanian yang dapat dimanfaatkan sebagai makanan ternak. Zat-zat yang terkandung di dalamnya seperti selulosa sebenarnya masih dapat dimanfaatkan oleh sapi, namun terselubung oleh dinding keras, yakni silika dan lignin, sehingga sulit dicerna. Teknologi suplementasi merupakan salah satu cara yang dapat meningkatkan kemampuan mikroba terutama bakteri rumen dalam meningkatkan kapasitas ternak untuk mencerna jerami padi. Teknologi suplementasi yang dicobakan adalah dalam bentuk suplemen yang pemberiannya dipisahkan dari jerami padiatau digabungkan dengan jerami padi dalam bentuk pakan komplit. Suplemen yang dikembangkan adalah suplemen kaya nutrien yang mengandung bahan-bahan berikut: dedak padi, tepung daun (daun singkong, daun lamtoro, dan daun turi), tepung ikan, dan campuran mineral.
Terdapat empat perlakuan, yaitu R1 (ransum mengandung 100% jerami padi), R2 (ransum mengandung 82,78% jerami padi dan 17,22% dedak padi), R3 (ransum mengandung 80,39% jerami padi, 16,72%, dedak padi dan 2,89% SKN), dan R4 (ransum mengandung 100% pakan komplit). Keempat ransum perlakuan ini difermentasi in vitro dengan metoda Tilley dan Terry (1963) yang dimodifikasi oleh Sutardi (1979), dan populasi bakteri dan protozoa dihitung pada waktu inkubasi 1, 3 dan 5 jam. Rancangan percobaan yang digunakan dalam penelitian ini adalah rancangan acak kelompok (RAK) berpola 4 x 3 dengan 4 perlakuan faktor A (ransum perlakuan) dan 3 perlakuan faktor B (waktu inkubasi) serta 5 ulangan. Peubah yang diamati antara lain populasi protozoa total, bakteri total, bakteri proteolitik, bakteri amilolitik, dan bakteri selulolitik.
Hasil penelitian menunjukkan bahwa lama waktu inkubasi memberikan pengaruh yang nyata (P<0,05) terhadap populasi bakteri selulolitik, sedangkan penambahan suplemen kaya nutrien di dalam ransum berbasis jerami padi tidak berpengaruh nyata terhadap populasi protozoa, bakteri total, bakteri proteolitik, bakteri amilolitik, dan bakteri selulolitik. Kesimpulan dari penelitian ini adalah populasi bakteri selulolitik lebih dipengaruhi oleh waktu inkubasi, sementara itu pemakaian suplemen kaya nutrien dan pakan komplit pada ransum berbasis jerami padi belum dapat memberikan pengaruh yang baik terhadap populasi protozoa dan bakteri rumen.
ii ABSTRACT
Supplementation Technology and Complete Feed in Rice Straw Based Diet on Rumen Bacterial and Protozoal Population
Sari, F., A. S. Tjakradidjaja, and I. K. Amrullah
The purpose of this experiment is to investigate the effect of supplementation technology and complete feed in rice straw based diet on rumen bacterial and protozoal populations. This study was conducted using a factorial randomized block design 4 x 3 with five replicates. Factor A was four types of ration whereas factor B was incubation times: B1 = 1 hour, B2 = 3 hours, and B3 = 5 hours. The four types of ration were R1= 100% of rice straw on dry matter (DM) basis; R2 = 82.78% of rice straw + 17.22% of rice bran (DM basis); R3 = 80.39% of rice straw + 16.72% of rice bran + 2.89% of rich nutrient supplement (DM basis); R4 = 100% of complete feed consisting of 40% rice straw, 30.5% rice bran, 9% leaf meal, 8.5% fish meal, 10% molasses, 1% palm oil, and 1% mineral mix (DM basis). The result showed that cellulolytic bacteria were influenced by incubation time (P<0.05). The result of contrast orthogonal test showed that 1 hour incubation was the optimal time for cellulolytic bacterial growth. Protozoa, total bacteria, proteolytic, amilolytic and cellulolytic bacteria were not affected by treatments. It is concluded that supplementation technology and complete feed has not yet produced greater effects on rumen bacterial and protozoal population.
iii
TEKNOLOGI SUPLEMENTASI DAN PAKAN KOMPLIT PADA
RANSUM BERBASIS JERAMI PADI TERHADAP POPULASI
BAKTERI DAN PROTOZOA RUMEN
FATMA SARI D24070117
Skripsi ini merupakan salah satu syarat untuk memperoleh gelar Sarjana Peternakan pada
Fakultas Peternakan Institut Pertanian Bogor
DEPARTEMEN ILMU NUTRISI DAN TEKNOLOGI PAKAN FAKULTAS PETERNAKAN
iv Judul Skripsi : Teknologi Suplementasi dan Pakan Komplit pada Ransum Berbasis
Jerami Padi terhadap Populasi Bakteri dan Protozoa Rumen
Nama : Fatma Sari
NIM : D24070117
Menyetujui,
Pembimbing Utama,
(Ir. Anita S. Tjakradidjaja, M.Rur.Sc.) NIP: 19610930 198603 2 003
Pembimbing Anggota,
(Dr. Ir. Ibnu K. Amrullah, MS.) NIP: 19521110 198003 1 004
Mengetahui : Ketua Departemen
Ilmu Nutrisi dan Teknologi Pakan
(Dr. Ir. Idat Galih Permana, M.Sc.Agr.) NIP: 19670506 199103 1 001
v RIWAYAT HIDUP
Penulis dilahirkan pada tanggal 9 November 1988 di Payakumbuh, Sumatera Barat. Penulis adalah anak kesepuluh dari sepuluh bersaudara dari pasangan bapak Ali Syarkawi dan ibu Darusni.
Pendidikan dasar diselesaikan pada tahun 2001 di SDN 45 Limbanang, pendidikan lanjutan menengah pertama diselesaikan pada tahun 2004 di SMPN 1 Kecamatan Suliki dan pendidikan lanjutan menengah atas diselesaikan pada tahun 2007 di SMAN 1 Limbanang. Penulis diterima sebagai mahasiswa Tingkat Persiapan Bersama (TPB), Institut Pertanian Bogor melalui Undangan Seleksi Masuk IPB (USMI) pada tahun 2007 dan diterima sebagai mahasiswa Departemen Ilmu Nutrisi dan Teknologi Pakan, Fakultas Peternakan, Institut Pertanian Bogor pada tahun 2008.
vi KATA PENGANTAR
Alhamdulillah, segala puji dan syukur penulis panjatkan kehadirat Allah SWT yang telah melimpahkan rahmat dan hidayah-Nya sehingga penulis dapat menyelesaikan penelitian dan penyusunan skripsi ini. Shalawat serta salam semoga tercurah kepada Nabi Besar Muhammad SAW, keluarga, sahabat dan para pengikutnya hingga akhir zaman.
Skripsi ini berjudul “Teknologi Suplementasi dan Pakan Komplit pada
Ransum Berbasis Jerami Padi terhadap Populasi Bakteri dan Protozoa Rumen”.
Penelitian ini dilakukan di Laboratorium Biokimia, Fisiologi dan Mikrobiologi Nutrisi, Departemen Ilmu Nutrisi dan Teknologi Pakan, Fakultas Peternakan, Institut Pertanian Bogor selama 7 bulan dari bulan November 2010 sampai Juni 2011.
Semoga skripsi ini dapat bermanfaat sebagai sumber informasi.Penulis mengucapkan terima kasih kepada seluruh pihak yang telah ikut berperan serta sehingga skripsi ini dapat diselesaikan.
Bogor, Pebruari 2012
vii
Pakan Komplit Berbasis Jerami Padi ... 3
Suplemen Pakan... 5
Urea Molasses Block dan Urea Cassava Block ... 6
Urea Molasses Multinutrient Block ... 7
Feed Block Supplement ... 9
Suplemen Pakan Multinutrien ... 11
Suplemen Kaya Nutrien ... 12
Bahan Baku Suplemen Kaya Nutrien sebagai Sumber Energi ... 13
Dedak Padi ... 13
Molases ... 14
Bahan Baku Suplemen Kaya Nutrien sebagai Sumber Protein ... 14
viii
Cairan Rumen ... 23
Alat ... 23
Bahan ... 23
Prosedur ... 23
Pengambilan Cairan Rumen ... 23
Pembuatan Media Pengenceran... 23
Pembuatan Larutan McDougal ... 24
Pembuatan Media Stok Bakteri ... 24
Percobaan in vitro ... 24
Perhitungan Populasi Protozoa... 25
Perhitungan Bakteri Total, Proteolitik, Amilolitik dan Selulolitik ... 25
Rancangan dan Analisis Data ... 26
Perlakuan ... 26
Model ... 26
Peubah ... 27
HASIL DAN PEMBAHASAN ... 28
Kandungan Zat Makanan dan Ransum Komplit ... 28
Populasi Mikroba Rumen ... 31
Populasi Protozoa ... 31
Bakteri Total ... 32
Bakteri Proteolitik ... 35
Bakteri Amilolitik ... 36
Bakteri Selulolitik ... 38
KESIMPULAN ... 40
SARAN……. ... 40
UCAPAN TERIMAKASIH ... 41
DAFTAR PUSTAKA ... 42
ix DAFTAR TABEL
Nomor Halaman
1. Kandungan Nilai Gizi Jerami Padi ... 3
2. Komposisi Bahan Urea Molasses Block dan Urea Cassava Block ... 7
3. Komposisi Kimia Urea Molasses Block dan Urea Cassava Block ... 7
4. Komposisi Bahan Urea Molasses Multinutrient Block... 8
5. Komposisi Kimia Urea Molasses Multinutrient Block ... 8
6. Komposisi Bahan Feed Block Supplement A dan Feed Block Supplement B ... 10
7. Komposisi Kimia Feed Block Supplement A dan Feed Block Supplement B ... 10
8. Komposisi Kimia Suplemen Pakan Multinutrien ... 11
9. Komposisi Kimia Suplemen Kaya Nutrien ... 12
10. Komposisi Ransum Percobaan ... 22
11. Kandungan Zat Makanan Pakan Penelitian ... 28
12. Komposisi Nutrisi Ransum Perlakuan ... 29
13. Populasi Protozoa Rumen ... 32
14. Populasi Bakteri Total ... 33
15. Populasi Bakteri Proteolitik ... 36
16. Populasi Bakteri Amilolitik ... 37
x DAFTAR GAMBAR
Nomor Halaman
xi DAFTAR LAMPIRAN
Nomor Halaman
1. Komposisi dan Pembuatan Garam Formalin ... 51
2. Komposisi dan Pembuatan Media ... 51
3. Komposisi dan Pembuatan Larutan McDougall ... 52
4. Sidik Ragam untuk Pengaruh Perlakuan terhadap Populasi Protozoa ... 53
5. Sidik Ragam untuk Pengaruh Perlakuan terhadap Populasi Bakteri Total... 53
6. Sidik Ragam untuk Pengaruh Perlakuan terhadap Populasi Bakteri Proteolitik ... 53
7. Sidik Ragam untuk Pengaruh Perlakuan terhadap Populasi Bakteri Amilolitik ... 54
8. Sidik Ragam untuk Pengaruh Perlakuan terhadap Populasi Bakteri Selulolitik ... 54
1 PENDAHULUAN
Latar Belakang
Hijauan merupakan bahan pakan yang penting bagi ternak ruminansia. Namun ketersedian hijauan ini sangat dipengaruhi oleh faktor musim dimana pada musim penghujan tersedia dalam jumlah banyak dan melimpah sedangkan pada musim kemarau ketersediaannya semakin berkurang. Oleh karena itu perlu diupayakan pemanfaatan sumber pakan lain seperti produk samping pertanian, seperti jerami padi.
Jerami padi merupakan salah satu produk samping pertanian yang tersedia cukup melimpah.Jerami padi yang dihasilkan dapat mencapai 5 ton/ha setiap kali panen (Haryanto et al., 2002). Kandungan bahan kering (BK) jerami padi segar sekitar 40–45%, sehingga potensi produksi BK jerami padi adalah 2,0–2,25 ton/ha. Adapun kebutuhan ternak sapi adalah sekitar 6–7 kg BK jerami padi per hari untuk sapi dengan bobot hidup 250 kg. Dengan demikian produksi jerami padi (per satuan BK) dapat digunakan sebagai pakan untuk 357 ekor sapi.
Namun demikian penggunaan jerami padi secara langsung sebagai pakan tunggal tidak dapat memenuhi pasokan nutrisi yang dibutuhkan ternak. Hal ini disebabkan kandungan protein kasarnya rendah sementara kandungan serat kasarnya tinggi. Kandungan serat kasar yang tinggi dapat menghambat mikroba rumen dalam mencerna pakan. Oleh karena itu, penelitian dan pengembangan terus dilakukan untuk meningkatkan kualitas jerami padi agar dapat dimanfaatkan sebagai bahan pakan secara optimal, terutama untuk ternak ruminansia. Teknologi suplementasi merupakan satu alternatif yang diharapkan dapat meningkatkan kemampuan mikroba, terutama bakteri rumen, dalam meningkatkan kapasitas ternak untuk mencerna jerami padi.
2 campuran mineral. Bahan-bahan tersebut merupakan bahan yang tersedia di dekat lokasi peternakan sapi pedaging seperti yang terdapat di Kabupaten Rembang.
Tujuan
3 TINJAUAN PUSTAKA
Pakan Komplit Berbasis Jerami Padi
Jerami padi merupakan bagian dari batang tumbuhan tanpa akar yang tertinggal setelah dipanen butir buahnya (Shiddieqy, 2005). Jerami padi merupakan salah satu limbah pertanian yang sangat potensial sebagai sumber energi untuk ternak ruminansia, karena produksi jerami padi sangat banyak dan tersedia sepanjang tahun. Menurut Departemen Pertanian Republik Indonesia, Deptan RI, (2001), produksi padi tahun 2000 sebanyak 50.866.387 ton, bila diasumsikan jumlah jerami padi adalah 50% dari produksi padi yang dihasilkan pada tahun yang sama, maka produksi jerami padi sebanyak 25.433.194 ton. Dari jumlah tersebut baru sekitar 7,8% yang sudah dimanfaatkan sebagai makanan ternak. Pemanfaatan jerami padi sebagai pakan baru mencapai 31–39%, sedangkan yang dibakar atau dikembalikan ke tanah sebagai pupuk 36–62%, dan sekitar 7–16% digunakan untuk keperluan industri (Syamsu, 2007).
Jerami padi merupakan bahan pakan herbivora yang tergolong bahan pakan yang berkualitas rendah karena dinding selnya tersusun oleh selulosa, hemiselulosa, lignin dan silika (Budiman, 2007). Penggunaan jerami padi sebagai makanan ternak mengalami kendala karena adanya faktor pembatas nilai nutrisi yang rendah yaitu kandungan protein rendah, serat kasar tinggi, dan kecernaan rendah (Tabel 1).
Tabel 1. Kandungan Nilai Gizi Jerami Padi Komposisi Kimia
Bahan Kering (%) 76,93
Protein Kasar (%) 5,06
Serat Kasar (%) 34,98
Lemak Kasar (%) 3,85
Gross Energy (kalori/g) 43,0 Sumber: Prasetiyono et al. (2007)
4 dikonsumsi 10 kg, maka hanya 3 kg saja yang dapat dicerna. Daya cerna yang rendah dari jerami padi akibat struktur jaringan penyangga tanaman yang sudah tua. Jaringan tanaman ini sudah mengalami proses lignifikasi, dimana terjadi ikatan lignoselulosa dan lignohemiselulosa yang sulit dicerna (Shiddieqy, 2005). Menurut Suminar (2005), lignin merupakan faktor yang lebih banyak mempengaruhi rendahnya daya cerna dari jerami tanaman pada umumnya, sedangkan pada jerami padi rendahnya daya cerna disebabkan oleh tingginya kandungan silika. Proses lignifikasi dan silifikasi secara bersama-sama mempengaruhi rendahnya daya cerna jerami padi. Lebih lanjut Suminar (2005) menyatakan bahwa jerami padi dalam keadaan segar relatif lebih hijau, mempunyai kadar air, palatabilitas dan kecernaan lebih tinggi dibandingkan dengan yang sudah kering dan bertumpuk.
Perlakuan pada jerami padi untuk meningkatkan nilai gizinya dapat dilakukan secara fisik, kimia, dan biologis atau kombinasi diantaranya. Perlakuan secara fisik dapat dilakukan dengan pelayuan, penggilingan, pembuatan pellet, penyinaran radiasi dan proses penguapan. Perlakuan secara kimia dapat dilakukan dengan perendaman dalam larutan asam atau basa, penambahan urea, amonia dan molases. Perlakuan secara biologis dapat dilakukan penambahan enzim dan penambahan kapang pada jerami padi (Ibrahim, 1983). Kartadisastra (1967) menyatakan bahwa nilai cerna jerami yang rendah dapat ditingkatkan menjadi lebih dari 50% dengan cara melakukan proses pengolahan jerami tersebut dengan perlakuan kimiawi menggunakan urea, molases, larutan NaOH teknis dan dengan perlakuan fermentasi. Menurut Suryahadi dan Piliang (1994), perlakuan secara fisik mampu meningkatkan kualitas, namun hasil yang diperoleh tidak sebanding dengan energi dan biaya yang dikeluarkan. Perlakuan kimia dapat meningkatkan kecernaan lebih besar, tetapi biaya yang dikeluarkan tetap tinggi.
5 Ransum komplit merupakan pakan yang cukup gizi untuk hewan tertentu dalam tingkat fisiologi, dibentuk atau dicampur untuk diberikan sebagai satu-satunya makanan dan memenuhi kebutuhan pokok atau produksi, atau keduanya tanpa tambahan bahan atau substansi lain kecuali air (Tillman et al., 1997). Menurut Ensminger et al. (1990), beberapa keuntungan yang diperoleh dari penggunaan ransum komplit antara lain : 1) meningkatkan efisiensi pemberian pakan, 2) meningkatkan konsumsi ketika hijauan kurang disukai ternak, 3) jika konsentrat terbatas dapat digunakan hijauan sebagai campuran ransum komplit, dan 4) ransum komplit dapat mempermudah ternak untuk mendapatkan pakan yang lengkap. Ransum komplit untuk ternak ruminansia biasanya disusun dari hijauan dan konsentrat dengan perbandingan tertentu. Saragih (2003) menyatakan bahwa konsentrat adalah pakan yang kaya akan sumber protein atau sumber energi, dan dapat mengandung pelengkap pakan dan imbuhan pakan. Konsentrat banyak digunakan pada usaha penggemukan sapi potong untuk meningkatkan bobot badan. Kandungan protein konsentrat umumnya berkisar antara 12–13% dengan kandungan energi 2400 kkal per kg bahan (Winugroho, 2004).
Suplemen Pakan
6 kekurangan nutrien ketika kualitas dan kuantitas hijauan sangat kurang untuk menghasilkan performa ternak yang diharapkan. BATAN (2005) menambahkan bahwa suplemen yang tersusun dari kombinasi bahan limbah protein dengan tingkatan jumlah tertentu dapat mendukung pertumbuhan, perkembangan dan kegiatan mikroba secara efisien di dalam rumen.
Urea Molasses Block (UMB) dan Urea Cassava Block (UCB)
Urea Molasses Block (UMB) merupakan suplementasi mineral dalam bentuk blok dengan bahan baku lokal sebagai bahan baku utama terutama dari limbah industri pertanian dan pakan non konvensional yang ketersediaannya murah dan mudah diperoleh (Devendra, 1988). Thu dan Uden (2000) menyatakan bahwa UMB merupakan suplemen mineral dalam bentuk blok yang terdiri dari bahan-bahan berupa molases, urea, singkong, minyak biji-bijian dan mineral. Bahan-bahan tersebut berfungsi sebagai sumber energi mudah tercerna, sumber N dan sumber mineral yang dapat meningkatkan pertumbuhan mikroba rumen untuk memperbaiki nitrisi ternak sebagai induk semang. Suplementasi UMB dapat memperbaiki kualitas pakan yang rendah seperti jerami padi yaitu dengan adanya tambahan zat makanan dari UMB.
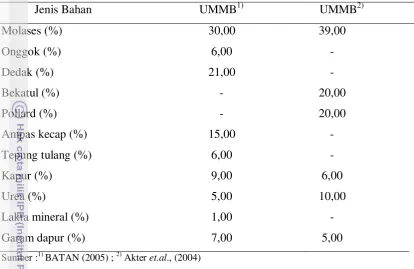
7 Tabel 2. Komposisi Bahan Urea Molasses Block dan Urea Cassava Block*
Jenis Bahan UMB UCB
Molases (%) 47,00 -
Cassava, ubi kayu (%) - 49,00
Dedak Padi (%) 25,00 1,50
CaO (%) 17,00 5,10
Urea (%) 4,00 5,00
Garam (%) 4,00 5,00
TSP (%) 2,00 3,00
Campuran mineral (%) 1,00 2,00
Keterangan :* Komposisi berdasarkan Suryahadi(1999)
Tabel 3. Komposisi Kimia Urea Molasses Block dan Urea Cassava Block
Komposisi Kimia UMB UCB
Protein Kasar (%) 17,60 19,50
Serat Kasar (%) 7,60 5,40
Lemak Kasar (%) 1,40 1,50
Ca (%) 7,10 5,10
P (%) 0,90 1,30
K (%) 2,20 1,20
Sumber :Widasari (2002)
Urea Molasses Multinutrient Block (UMMB)
8 Tabel 4. Komposisi Bahan Urea Molasses Multinutrient Block
Jenis Bahan UMMB1) UMMB2)
Molases (%) 30,00 39,00
Onggok (%) 6,00 -
Dedak (%) 21,00 -
Bekatul (%) - 20,00
Pollard (%) - 20,00
Ampas kecap (%) 15,00 -
Tepung tulang (%) 6,00 -
Kapur (%) 9,00 6,00
Urea (%) 5,00 10,00
Lakta mineral (%) 1,00 -
Garam dapur (%) 7,00 5,00
Sumber :1) BATAN (2005) ; 2) Akter et.al., (2004)
Tabel 5. Komposisi Kimia Urea Molasses Multinutrient Block
Komposisi Kimia UMMB
Bahan Kering (%) 93,30
Bahan Organik (%) 72,30
Protein Kasar (%) 24,50
Serat Kasar (%) 6,50
Lemak Kasar (%) 3,50
Ca (%) 7,30
P (%) 1,10
S (%) 1,50
K (%) 9,40
Fe (%) 2,90
Cu (ppm) 4985
Zn (ppm) 2204
Sumber : BATAN (2005)
9 NPN sebagai sumber proteinnya. Penggunaan urea sebagai sumber NPN harus diimbangi dengan sumber energi yang akan digunakan sebagai kerangka karbonnya. Oleh karena itu penggunaan urea akan lebih efektif digunakan dalam ransum rendah protein, tetapi tinggi kandungan energinya (Suryahadi, 1989; Orskov, 1998).
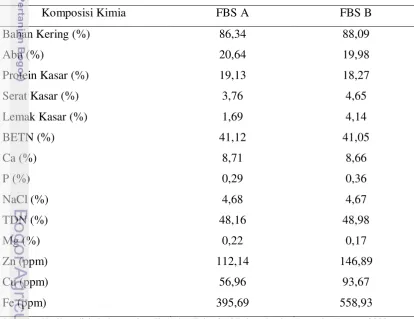
Feed Block Supplement (FBS)
Feed Block Supplement (FBS) merupakan pakan non-konvensional bagi ternak ruminansia yang di dalamnya terdapat berbagai macam bahan pakan yang dikemas dalam bentuk blok. Bahan-bahan penyusun FBS antara lain molases, pollard, urea, kapur, garam, mineral mikro, minyak ikan lemuru, daun kembang sepatu, ragi tape, dan vanila. Formula FBS yang telah diujikan terdiri dari 2 jenis yaitu FBS A dan FBS B. FBS A mempunyai unsur suplemen berupa nitrogen bukan protein (NPN), mineral dari penelitian Widasari (2002) dan Winarni (2002). FBS B merupakan formula dari penelitian Danirih (2004) dan Sahlan (2005) dengan komponen aktif selain yang ada pada FBS A, juga ditambahkan agen defaunasi (defaunating agent) berupa tepung daun kembang sepatu, penghambat metan (methan inhibitor) berupa minyak ikan lemuru, probiotik Saccharomyces cerevisiae berupa ragi tape. Komposisi bahan dan komposisi kimia FBS A dan FBS B dapat dilihat pada Tabel 6 dan 7.
10 Tabel 6. Komposisi Bahan Feed Block Supplement A dan Feed Block Supplement B
Jenis Bahan FBS A FBS B
Molases (%) 44,00 34,80
Pollard (%) 30,00 14,00
Urea (%) 5,00 5,00
Kapur (%) 14,00 14,00
Garam (%) 5,00 5,00
Mikromineral (%) 2,00 2,00
Minyak lemuru (%) - 5,00
Kembang sepatu (%) - 10,00
Ragi tape (%) - 10,00
Vanila (%) - 0,20
Sumber : Danirih (2004)
Tabel 7. Komposisi Kimia Feed Block Supplement A dan Feed Block Supplement B
Komposisi Kimia FBS A FBS B
Bahan Kering (%) 86,34 88,09
Abu (%) 20,64 19,98
Protein Kasar (%) 19,13 18,27
Serat Kasar (%) 3,76 4,65
Lemak Kasar (%) 1,69 4,14
BETN (%) 41,12 41,05
Ca (%) 8,71 8,66
P (%) 0,29 0,36
NaCl (%) 4,68 4,67
TDN (%) 48,16 48,98
Mg (%) 0,22 0,17
Zn (ppm) 112,14 146,89
Cu (ppm) 56,96 93,67
Fe (ppm) 395,69 558,93
11 Suplemen Pakan Multinutrien (SPM)
Suplemen Pakan Multinutrien merupakan suplemen yang dikembangkan oleh BATAN dari suplemen sebelumnya yaitu UMMB. SPM dikembangkan untuk mengatasi beberapa kendala seperti ketersediaan pakan lokal, harga dan bahan penyusun formula suplemen pakan UMMB. Bahan-bahan yang sulit didapat yaitu molases, tepung tulang, dan bungkil kedelai (BATAN, 2005). Tabel 8 memperlihatkan komposisi kimia SPM.
Kandungan molases dan bungkil kedelai SPM lebih rendah dibandingkan dengan UMMB yaitu sebesar 10% dan 3%, sedangkan UMMB sebesar 29% dan 17%, sehingga harga dari SPM lebih murah daripada UMMB (BATAN, 2005). Bahan yang digunakan dalam pembuatan SPM antara lain bungkil kedelai, ampas kecap, daun gamal dan bubur bayi afkir, urea, mineral makro dan mikro (BATAN, 2005). SPM juga memiliki kelebihan yaitu di dalamnya terkandung imbuhan pakan yang dapat berperan dalam proses metabolisme dalam tubuh ternak. Kelebihan lainnya adalah adanya protein by-pass yang dapat langsung dimanfaatkan oleh ternak untuk memenuhi kebutuhan proteinnya.
Tabel 8. Komposisi Kimia Suplemen Pakan Multinutrien
Komposisi Kimia SPM
Bahan Kering (%) 89,30
Bahan Organik (%) 92,20
Protein Kasar (%) 28,80
Serat Kasar (%) 17,20
Lemak Kasar (%) 4,70
Ca (%) 1,50
P (%) 0,40
S (%) 0,70
Fe (%) 1,70
Cu (ppm) 2259
Zn (ppm) 1650
12 Suplemen Kaya Nutrien (SKN)
SKN yang telah dibuat sebelumnya pada penelitian Sulistyo (2008) dan Wahyuni (2008) merupakan teknik pengkayaan dari SPM yang telah dikembangkan oleh BATAN. Bahan-bahan yang digunakan dalam pembuatan SKN ini antara lain molases, onggok, bekatul, ampas tahu, ampas kecap, kapur, tepung tulang, urea, campuran mineral, garam dapur, ampas teh, daun kembang sepatu dan kunyit. SKN ini memiliki beberapa kelebihan dibandingkan SPM yaitu memiliki kandungan protein kasar yang cukup tinggi sebesar 20,55% (Tabel 9). Selain itu SKN mengandung protein by-pass, agen defaunasi, dan mineral organik.
Tabel 9. Komposisi Kimia Suplemen Kaya Nutrien
Komposisi Kimia SKN1) SKN2) BETN (Sutardi, 2003 dalam Noviana, 2004)
**) TDN dihitung berdasarkan rumus Harris et al., (1972) 1)
SKN hasil pengkayaan SPM
2)
SKN penelitian
Sumber : 1) Hasil Analisa Pusat Penelitian Sumberdaya Hayati dan Bioteknologi IPB, 2007
2)
Hasil Perhitungan berdasarkan data Sutardi (1980)
13 proteintidak terdegradasi (protein bypass) atau Rumen Undegradable Protein (RUP), agen defaunasi, dan campuran mineral.
Bahan Baku Suplemen Kaya Nutrien sebagai Sumber Energi
Suplemen Kaya Nutrien (SKN) disusun dengan menggunakan bahan lokal dengan produksi yang cukup tinggi dan mudah didapat. Bahan makanan yang digunakan berupa campuran sumber energi, sumber protein dan sumber mineral. Berdasarkan kandungan nutrisinya, bahan pakan dapat dikelompokkan menjadi bahan baku sumber energi, bahan baku sumber protein baik protein hewani maupun protein nabati, bahan pakan sumber mineral, sumber vitamin, dan bahan pakan imbuhan seperti antibiotika dan hormon (Sofyan et al., 2000). Bahan sumber energi adalah bahan yang dapat menyediakan energi lebih banyak bagi ternak dibanding zat makanan lain. Bahan pakan yang digolongkan sebagai sumber energi adalah bahan-bahan pakan yang berasal dari hijauan maupun konsentrat (biji-bijian, kacang-kacangan, buah-buahan, umbi-umbian, dan limbah sisa penggilingan) dengan kandungan protein kasar (PK) kurang dari 20% dan kandungan serat kasar (SK) kurang dari 18% (Sukria dan Krisnan, 2009). Bahan sumber energi yang dapat digunakan dalam SKN yaitu dedak padi dan molases.
Dedak Padi
Beras merupakan komoditas yang sangat penting di Indonesia karena beras merupakan makanan pokok sebagian besar penduduk Indonesia. Dalam proses penggilingan padi menjadi beras, selain akan menghasilkan kulit beras atau sekam sekitar 15–20%, juga akan menghasilkan dedak padi yang merupakan kulit ari hasil dari penyosohan sekitar 8–12% dan menir sekitar 5% (Widowati, 2001). Persentase hasil samping tergantung kecepatan penggilingan, jenis padi dan faktor-faktor lain. Menurut NRC (2000), dedak padi mengandung PK 14,4%, dan SK 13%.
14 Molases
Molases merupakan sumber energi yang murah karena mengandung gula sebanyak 50%, baik dalam bentuk sukrosa 20–30% maupun dalam bentuk gula pereduksi 10–30%. Gula-gula pereduksi tersebut sangat mudah dicerna dan dapat langsung diserap oleh darah dan digunakan untuk pembakaran untuk keperluan energi. Molases mangandung 2,5–4,5% PK, separuh dari protein tersebut merupakan protein yang dapat dicerna. Berbagai asam amino yang banyak terdapat dalam molases adalah : aspartat, glutamat, pyrolidin karboksilat, asparagin, lisin dan alanin. Di samping itu, molases sangat kaya akan mineral. Kadar abu molases antara 2,5–7% sebagai karbonat. Sebagian besar dari kadar abu tersebut terdiri dari kalium 30–50% (sebagai K2O), besi dengan kadar 0,4–7% (sebagai Fe2O3), dan fosfor 0,5–2,5% (P2O5). Kadar kalsium molases empat kali lebih besar daripada susu. Kadar vitamin, khususnya vitamin-vitamin yang tahan panas dan basa (CaOH2), relatif sangat tinggi di dalam molases. Dalam satu cup molases mengandung biotin 48,00 mg, asam pantotenat 780,00 mg, riboflavin 0,57 mg dan niacin 6,0 mg (Winarno, 1982).
Menurut Perry et al. (2004), tujuan pemberian molases di dalam ransum ternak sapi, domba, dan kuda yaitu:
1. Untuk memperbaiki palatabilitas ransum 2. Untuk memperbaiki aktivitas mikroba rumen 3. Untuk mengurangi kadar kotoran
4. Sebagai pengikat untuk pellet 5. Sebagai sumber energi
Bahan Baku Suplemen Kaya Nutrien sebagai Sumber Protein
15 Daun Singkong (Manihot utilissima)
Potensi daun singkong sebagai pakan ternak di Indonesia cukup besar. Daun singkong adalah bahan pakan sumber protein, kandungan protein rata-rata adalah 21% bervariasi dari 16,1–39,9% tergantung varietas, umur panen, kesuburan tanah dan iklim. Daun singkong sumber mineral yang baik, kaya akan Ca, Mg, Mn dan Zn, serta kaya akan vitamin C, vitamin A dan riboflavin (Ravindran, 1992). Kandungan zat makanan daun singkong berdasarkan bahan kering adalah protein kasar (PK) 23,20%, lemak kasar (LK) 4,80%, abu 7,80%, BETN 42,30%, Ca 0,37%, P 0,58%, lisin 7,11%, metionin dan sisti 2,53%, treonin 4,70% dan triptopan 1,09% (Khajerern dan Khajerern, 1992).
Daun Lamtoro (Leucaena leucocephala)
Daun lamtoro telah banyak digunakan untuk meningkatkan produktivitas ternak di daerah tropis (Khamseekhiew et al., 2001). Tanaman lamtoro dapat diberikan kepada ternak berupa hijauan segar, kering, tepung, silase dan pelet. Hijauan lamtoro sangat baik sebagai pakan ternak. Hal ini dikarenakan daun lamtoro kaya akan protein, karoten, vitamin dan mineral (Rajendran et al., 2001). Daun lamtoro dapat digunakan sebagai bahan penyusun ransum karena kandungan protein yang cukup tinggi. Kandungan protein lamtoro berkisar 25–32% dari BK dan energi bruto berkisar antara 4237–4915 kalori/gram. Hartadi et al. (2005) juga mengatakan bahwa kandungan nutrien lamtoro adalah PK 23,70%, SK 18%, LK 5,80%, Ca 1,40%, dan P 0,21%. Dengan demikian lamtoro merupakan sumber protein dan sumber energi (Marlina dan Askar, 1998). Lebih lanjut ditambahkan oleh Tambunan et al. (1997) bahwa daun lamtoro mempunyai kualitas protein yang baik karena asam aminonya berimbang cukup baik. Efektif degradasi PK daun lamtoro sebesar 51,63% (Afzalani et al., 2000).
Daun lamtoro merupakan penyedia protein dan sebagai protein by pass untuk meningkatkan jumlah protein yang lolos dari degradasi rumen sehingga jumlah protein yang diserap di usus halus semakin meningkat. Daun lamtoro mengandung protein yang relatif rendah tingkat pemecahannya di dalam rumen yang merupakan sumber protein yang bagus untuk ternak ruminansia.
16 dalam lamtoro sebanyak 3–5% (Soebarinoto et al., 1991). Mimosin merupakan zat anti nutrien yang berada dalam bahan pakan, dimana apabila dikonsumsi oleh ternak dapat menyebabkan penurunan performan ternak tersebut (Widodo, 2005). McDonald et al. (2002) juga menyatakan bahwa apabila mimosin diberikan kepada ruminansia dalam kadar yang tinggi dapat menjadi racun bagi mikroba rumen sehingga dapat pula menurunkan produksi asam amino. Ternak ruminansia yang mengkonsumsi mimosin pada level tinggi dapat menunjukkan gejala kehilangan bulu (Tomaszewska et al., 1993). Efek negatif mimosin pada ruminansia tidak universal. Pada daerah beriklim sedang efek mimosin lebih sering terjadi daripada ternak di daerah tropis (Soebarinoto et al., 1991).
Untuk memperkecil kandungan mimosin dalam daun lamtoro dapat dilakukan dengan mengeringkan di bawah sinar matahari (Tambunan et al., 1997). Dengan pengeringan, kadar mimosin yang terkandung dalam daun lamtoro akan berkurang, sehingga dapat diberikan sebagai pakan ternak (Suprayitno, 1981).
Daun Turi (Sesbania grandiflora)
Ghol (1975) menyatakan bahwa daun turi merupakan tanaman kacang-kacangan yang dapat menyediakan hijauan dengan kualitas yang baik dan cepat tumbuh. Daun turi yang masih muda disukai oleh ayam, sapi dan domba, terutama pada saat musim kering dimana pada waktu itu umumnya hanya tersedia jerami atau rumput kering bagi ternak. Daun turi mengandung PK sebesar 20–40%, sedangkan kandungan SK dan LK berturut-turut sebesar 10–20% dan 2–7%. Selain kandungan protein yang tinggi, daun turi juga merupakan sumber mineral terutama Ca dan P yang besarnya berturut-turut 1–2,5% dan 0,2–0,5%. Di dalam daun turi ternyata banyak mengandung racun yang dikenal dengan nama saponin (Duke,1979). Dalam jumlah besar, saponin dapat memberikan pengaruh negatif terhadap legum (Wina dan Syaghiar, 1991).
Tepung Ikan
17 sehingga sering disubstitusi dengan meat and bone meal (MBM) (Indartono, 2003). Tepung ikan dari dalam negeri umumnya yang digunakan sebagai bahan mentah adalah yang berasal dari limbah pengolahan ikan menjadi ikan kalengan dan ikan limbah, bukan ikan segar seperti halnya tepung ikan impor, dan hanya sebagian kecil saja yang menggunakan ikan segar (Sultan, 2002).
Sisa-sisa atau limbah pengolahan ikan menjadi ikan kalengan seperti ikan tuna atau ikan sardin dan pengolahan udang beku pada umumnya terdiri dari kepala, ekor, isi perut, tulang, kulit dan sirip ikan atau udang; ikan limbah adalah ikan afkir berupa ikan kecil-kecil yang tidak dikonsumsi dan biasanya bercampur dengan kepiting-kepiting dan udang-udang kecil yang dikenal dengan sebutan ikan runcah atau ikan sampah; sedangkan ikan sisa adalah ikan yang tidak terjual di pasaran pada waktu pelelangan atau surplus hasil tangkapan pada musim puncak. Tidak digunakannya ikan segar dalam proses pengolahan menjadi tepung ikan di dalam negeri karena ikan segar masih merupakan sumber protein utama untuk dikonsumsi sebagian besar rakyat Indonesia. Selain itu penggunaan ikan segar dalam proses pengolahan menjadi tepung ikan harus bersaing dengan keperluan penggunaan lain seperti pembuatan ikan asin, kerupuk, kecap, terasi, bakso, dan lain-lain (Sultan, 2002).
Dalam proses pengolahan tepung ikan, bahan mentah yang digunakan akan berpengaruh terhadap kualitas hasil tepung ikan. Bila bahan mentah yang digunakan adalah berasal dari ikan segar, ikan diperlakukan dengan dengan baik serta dijaga dari pencemaran sebelum diolah maupun dalam proses pengumpulannya maka tepung ikan yang dihasilkan juga akan berkualitas baik. Sebaliknya bila bahan mentah yang digunakan berasal dari ikan yang tidak segar, ikan sisa atau limbah ikan, kualitas tepung ikan yang dihasilkan akan berkurang. Dengan demikian, kualitas tepung ikan yang dihasilkan dari proses pengolahan dalam negeri pada umumnya lebih rendah dari kualitas tepung ikan impor (Sultan, 2002).
18 Cairan Rumen
Rumen merupakan tabung besar dengan berbagai kantong yang menyimpan dan mencampur pakan hasil fermentasi mikroba. Kondisi dalam rumen adalah anaerobik dan hanya mikroorganisme yang paling sesuai dapat hidup di dalamnya. Tekanan osmosis dalam rumen mirip dengan tekanan aliran darah dan suhunya 38– 42 ºC. Cairan rumen berfungsi sebagai buffer dan membantu mempertahankan pH tetap pada nilai 6,8 (Sutardi, 1977). Perkembangan bakteri rumen terjadi karena adanya kontaminasi dari lingkungan dan kontak langsung induknya sehingga perkembangan populasi bakteri rumen akan terus meningkat seiring dengan bertambahnya umur ternak. Pemberian hijauan dan pakan berserat tinggi pada ternak ruminansia akan menstimulasi perkembangan rumen (Hobson dan Stewart, 1992).
Rumen sapi atau ternak ruminansia lainnya merupakan organ pencernaan yang selain tempat penampungan sementara makanan dalam jumlah banyak, rumen juga dihuni oleh berbagai macam mikroba (Karma, 2000). Di dalam rumen, makanan akan mengalami pencernaan oleh mikroba yang dikenal dengan pencernaan fermentatif, terutama dalam mendegradasi dinding sel tanaman (Lee et al., 2000).
Mikroba Rumen
Mikroba rumen termasuk di dalamnya sejumlah besar spesies bakteri, protozoa dan fungi membuat habitat rumen sangat komplek. Populasi mikroba dalam ekosistem sangat bervariasi antar individu maupun jenis hewan, tergantung jenis makanan yang diberikan karena hal itu akan berpengaruh terhadap efisiensi ruminan dalam menggunakan makanannya (Kamra, 2005).
Penghuni terbesar dalam cairan rumen adalah bakteri yaitu 1010–1011 sel/ml cairan rumen dan populasi terbesar kedua diduduki oleh protozoa yang populasinya mencapai 104–106 sel/ml cairan rumen. Jumlah tersebut dapat menjadi lebih besar karena sebagian besar mikroba tidak dikulturkan (Kamra, 2005). Sama seperti yang diungkapkan McDonald et al. (2002) bahwa protozoa merupakan mikroorganisme yang ada dalam rumen dengan jumlah lebih sedikit dibandingkan jumlah bakteri yaitu sekitar 1 juta/ml, tetapi dari segi jumlah biomassa protozoa lebih besar dibandingkan bakteri (Ogimoto dan Imai, 1981).
19 biji-bijian. Berbagai enzim dibutuhkan untuk mencerna makanan tersebut (Moharrery dan Das, 2001). Enzim-enzim disekresikan oleh mikroba ke dalam cairan rumen untuk mendegradasi partikel makanan (Kamra, 2005). Dengan demikian isi rumen terdiri atas padatan yang berasal dari bahan-bahan yang dimakan, dan cairan yang berisi mikroba rumen, enzim-enzim pencernaan yang disekresi oleh mikroba dan zat-zat makanan hasil perombakan mikroba rumen dan enzim serta vitamin-vitamin dan mineral-mineral yang larut dalam cairan rumen
Interaksi antar mikroba rumen terjadi sangat kompleks dan masing-masing mempunyai syarat tersendiri untuk tumbuh dan berkembang, misalnya bakteri selulolitik tidak dapat tumbuh pada pH cairan rumen lebih rendah dari 6,0 dan jumlahnya meningkat pada makanan yang kandungan serat kasarnya tinggi, hal ini terjadi sebaliknya pada bakteri amilolitik dan protozoa (Widiawati, 1992).
Faktor utama yang mempengaruhi pertumbuhan dan aktifitas populasi mikroba rumen adalah temperatur, pH, kapasitas buffer, tekanan osmotik, kandungan BK dan potensial oksidasi reduksi (Dehority, 2004). Pola pertumbuhan bakteri dan protozoa rumen dipengaruhi oleh pola fermentasi yang ditunjukkan oleh proporsi molar VFA dan pH rumen. Perkembangan populasi mikroba rumen secara umum ditentukan oleh tipe makanan yang dikonsumsi ternak (Arora, 1989), bangsa ternak, intensitas pemberian pakan, waktu pengambilan cairan rumen, konsentrasi NH3 (Preston dan Leng, 1987; Karimullah, 1991; Hakim, 2002), VFA dan pH rumen (Hungate, 1966), dan sifat predasi protozoa (Arora, 1989).
Bakteri Rumen
Flora atau bakteri rumen sebagian besar bersifat anaerob sejati, sedangkan yang bersifat anaerob fakultatif dan aerob terdapat dalam jumlah kecil (Ogimoto dan Imai, 1981). Menurut Jouany (1991), ada tiga lokasi yang membedakan bakteri berdasarkan letaknya, yaitu bakteri yang hidup menempel pada partikel makanan dalam rumen, bakteri yang menempel pada dinding epithelium rumen serta bakteri yang hidup bebas dalam cairan rumen. Preston dan Leng (1987) menyatakan bahwa jumlah bakteri yang menempel pada partikel makanan paling besar yaitu sekitar 70% dari total bakteri rumen.
20 menjadi asam organik dan dapat dimanfaatkan oleh ternak ruminansia. Bakteri menempel pada partikel pakan kasar dan perlahan-lahan mengikis material tercerna. Bakteri rumen memiliki fungsi yang sangat penting terhadap fermentasi serat dan tanaman berpolimer (Arora, 1989). Bakteri dipercaya berperan dominan dalam mendegradasi serat (Lee et al., 2000). Bakteri mengurai karbohidrat polimer dalam pakan menjadi senyawa sederhana seperti asam lemak dan alkohol dari selulosa, amilum, fruktosan dan xylan (Schlegel, 1994). Dalam pencernaan protein, bakteri lebih responsif dibandingkan protozoa (Moharrery dan Das, 2001).
Protozoa Rumen
Fardiaz (1992) menyatakan bahwa protozoa merupakan golongan protista tinggi yang mempunyai sifat lebih menyerupai hewan daripada tanaman atau yang biasa dikenal dengan eukariotik. Berbeda dengan bakteri yang digolongkan ke dalam prokariotik dan memiliki struktur yang lebih sederhana. Perbedaan antara eukariotik dan prokariotik terletak pada inti selnya. Eukariotik mempunyai inti sel sejati yaitu suatu struktur yang dikelilingi membran inti (nukleus) di mana di dalamnya terdapat kromosom. Di dalam nukleus terdapat nuklelous yang mempunyai kandungan RNA sangat tinggi. Nuklelous merupakan tempat sintesis RNA ribosom, sedangkan prokariotik tidak mempunyai inti sejati dan komponen keturunannya terdapat di dalam molekul DNA tunggal yang terletak bebas dalam sitoplasma.
Protozoa bersifat anaerob, apabila kadar oksigen atau pH rumen tinggi, maka protozoa tidak dapat membentuk cyte untuk mempertahankan diri dari lingkungan yang jelek, sehingga dengan cepat akan mati (Arora, 1989). Pada ruminansia, protozoa yang bersilia berkembang di dalam rumen dan membantu pencernaan zat-zat makanan dari rumput-rumputan yang kaya akan serat kasar. Berbeda dengan bakteri, protozoa lebih banyak mencerna pati sedangkan bakteri lebih berperan dalam mencerna serat (Bansi, 2001).
21 1989). Sebagian besar protozoa memakan bakteri untuk memperoleh sumber nitrogen dan mengubah protein bakteri menjadi protein protozoa, bersamaan dengan itu memperoleh tambahan sumber protein dan pati dari ingesta rumen. Dalam mendegradasi serat, protozoa hanya berperan sekitar 25–30 persen (Lee et al., 2000).
22 MATERI DAN METODE
Lokasi dan Waktu
Penelitian ini dilaksanakan dari bulan November 2010 sampai Juni 2011 di Laboratorium Biokimia, Fisiologi, dan Mikrobiologi Nutrisi, Departemen Ilmu Nutrisi dan Teknologi Pakan, Fakultas Peternakan, Institut Pertanian Bogor.
Materi Ransum
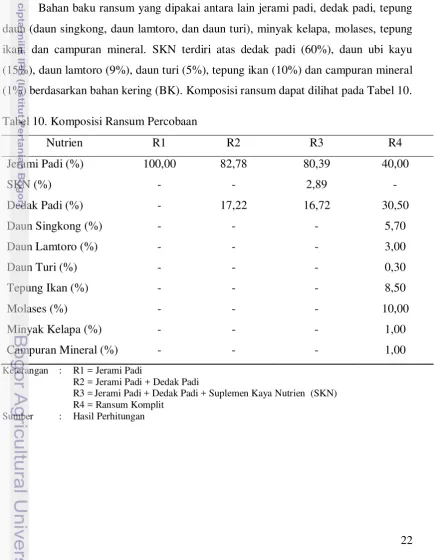
Bahan baku ransum yang dipakai antara lain jerami padi, dedak padi, tepung daun (daun singkong, daun lamtoro, dan daun turi), minyak kelapa, molases, tepung ikan, dan campuran mineral. SKN terdiri atas dedak padi (60%), daun ubi kayu (15%), daun lamtoro (9%), daun turi (5%), tepung ikan (10%) dan campuran mineral (1%) berdasarkan bahan kering (BK). Komposisi ransum dapat dilihat pada Tabel 10.
Tabel 10. Komposisi Ransum Percobaan
Nutrien R1 R2 R3 R4
Jerami Padi (%) 100,00 82,78 80,39 40,00
SKN (%) - - 2,89 -
Dedak Padi (%) - 17,22 16,72 30,50
Daun Singkong (%) - - - 5,70
Daun Lamtoro (%) - - - 3,00
Daun Turi (%) - - - 0,30
Tepung Ikan (%) - - - 8,50
Molases (%) - - - 10,00
Minyak Kelapa (%) - - - 1,00
Campuran Mineral (%) - - - 1,00
Keterangan : R1 = Jerami Padi
R2 = Jerami Padi + Dedak Padi
R3 = Jerami Padi + Dedak Padi + Suplemen Kaya Nutrien (SKN) R4 = Ransum Komplit
23 Cairan Rumen
Cairan rumen yang digunakan adalah cairan rumen yang berasal dari rumah potong hewan (RPH) di Fakultas Peternakan IPB dan di Bubulak.
Alat
Peralatan yang digunakan antara lain spoit, botol film, tutup karet ventilasi, pipet, gelas piala, labu Erlenmeyer, bulp, pipet Mohr, tabung fermentor, tabung pengencer, tabung Hungate, inkubator, tube roller, shaker water bath, autoclave, mikroskop, counting chamber (haemacytometer),cover glass, plastik kemasan, dan label.
Bahan
Bahan yang digunakan antara lain ransum berbasis jerami padi yang telah digiling (saringan berukuran 1,0 mm) dengan kadar air + 10%, larutan McDougall dengan temperatur 39 ºC, pH 6,5–6,9 (pH diturunkan dengan cara mengocok dengan gas CO2), cairan rumen segar dengan suhu 39 ºC, Trifanblue Formalsalin (TBFS), akuades, tepung Brain Heart Infusion (BHI), Carboxyl Methyl Celluloce (CMC), pati, casein, media pengencer, HCl 1%, NaCl 1%, dan alkohol 70%.
Prosedur Pengambilan Cairan Rumen
Termos diisi dengan air panas hingga mencapai suhu 39 ºC. Air di dalam termos tidak boleh dibuang hingga cairan rumen didapatkan. Cairan rumen diambil dari rumah potong hewan laboratorium Lapang Fakultas Peternakan IPB. Isi rumen diambil dan disaring dengan menggunakan kain penyaring, kemudian dimasukkan ke dalam termos yang sebelumnya sudah dibuang air panasnya. Cairan rumen dalam termos tersebut segera dibawa ke laboratorium Biokimia, Fisiologi, dan Mikrobiologi Nutrisi.
Pembuatan Media Pengenceran (Bryant dan Burkey, 1953)
24 hingga berubah warna dari biru menjadi bening. Campuran tersebut kemudian dimasukkan ke dalam tabung reaksi sebanyak 1 ml dan diotoklaf.
Pembuatan Larutan McDougal
Sebanyak 5 liter air destilasi dimasukkan ke dalam labu takar yang bervolume 6 liter dan dimasukkan bahan-bahan sebagai berikut: NaHCO3 sebesar 9,8 gram; Na2HPO4.7H2O sebesar 3,708 gram; KCl sebesar 0,97 gram; NaCl sebesar 0,47 gram; MgSO4.7H2O sebesar 0,12 gram; CaCl2.2H2O sebesar 0,053 gram. CaCl2.2H2O ditambahkan paling akhir setelah bahan lainnya larut sempurna. Leher labu dicuci dengan air destilasi hingga permukaan air mencapai tanda tera. Selanjutnya campuran dikocok dengan gas CO2 perlahan-lahan dengan melewatkannya dengan tujuan menurunkan pH hingga mencapai pH 6,8. Untuk tujuan ini diperlukan waktu paling sedikitnya 3 jam. Pemeriksaan pH dilakukan dan larutan dihangatkan sebanyak yang diperlukan hingga 37 ºC, jika perlu dikocok kembali dengan CO2 hingga pH mencapai 6,8 dengan catatan pH diturunkan sebelum larutan dihangatkan menjadi 37 ºC.
Pembuatan Media Stok Bakteri
Media stok dibuat dengan menambahkan media putih sebesar 4,85 ml dengan gliserol 80% sebesar 0,1 ml.
Percobaan in vitro
25 Perhitungan Populasi Protozoa
Perhitungan populasi protozoa dilakukan dengan meneteskan sampel (2 tetes) yang telah dicampur dengan larutan garam formalin (TFBS) dengan rasio 1:1 pada counting chamber (haemacytometer). Larutan TFBS dibuat dari campuran formalin 4% ditambah larutan garam NaCl fisiologis 0,9% dalam 100 ml larutan. Protozoa yang dihitung adalah total dari protozoa yang terdapat dalam counting chamber dengan ketebalan 0,1 mm, luas kotak terkecil 0,0625 mm2 yang terdapat 16 kotak dan jumlah kotak yang dibaca sebanyak 5 kotak. Perhitungan populasi protozoa dilakukan dengan mikroskop pada pembesaran 40 kali. Populasi protozoa dapat dihitung dengan rumus:
Keterangan:
C = Jumlah protozoa terhitung dalam counting chamber FP = Faktor Pengenceran
Perhitungan Bakteri Total, Proteolitik, Amilolitik dan Selulolitik
Populasi bakteri dihitung dengan metode pencacahan koloni bakteri hidup (Ogimoto dan Imai, 1981). Prinsip perhitungannya adalah cairan rumen diencerkan secara berseri lalu dibiakkan dalam tabung Hungate.
Media tumbuhBHI digunakan untuk menghitung populasi bakteri total. Media BHI dibuat dengan cara mencampur tepung BHI dengan bahan sumber nutrien mikroba lainnya, kemudian dimasukkan ke dalam botol yang telah diotoklaf. Campuran tersebut dipanaskan perlahan-lahan sambil dialiri gas CO2 sampai terjadi perubahan warna dari coklat menjadi merah dan berubah lagi menjadi coklat muda, lalu didinginkan. Selanjutnya media dimasukkan ke dalam tabung Hungate masing-masing sebanyak 5 ml yang sebelumnya telah diisi agar Bacto sebanyak 0,15 g, kemudian media disterilkan dalam otoklaf (suhu 121 ºC, 15 menit, tekanan 1,2Kgf/cm3). Media yang siap digunakan untuk pembiakan bakteri, dimasukkan ke dalam penangas air (suhu 47 ºC).
26 tumbuh untuk bakteri proteolitik ditambah dengan kasein, bakteri amilolitik ditambah dengan pati dan untuk bakteri dan selulolitik ditambah dengan Carboxyl Methyl Celluloce (CMC).
Menurut Ogimoto dan Imai (1981), contoh cairan rumen yang akan dikulturkan diencerkan terlebih dahulu dengan media pengencer. Pengenceran dilakukan sebagai berikut: 0,05 ml cairan rumen dimasukkan ke dalam 4,95 ml media pengencer. Selanjutnya diambil kembali 0,05 ml lalu dimasukkan ke dalam 4,95 ml media pengencer berikutnya, perlakuan tersebut dilakukan sampai 4 kali (4 seri tabung). Selanjutnya dari masing-masing seri tabung pengenceran diambil sebanyak 0,1 ml lalu ditransfer ke media agar lalu diputar sambil dialiri air, sehingga media dapat memadat secara merata pada dinding tabung dalam. Tabung selanjutnya diinkubasi selama 2–3 hari. Populasi bakteri dapat dihitung dengan rumus:
Populasi bakteri = n x 10x/0,05 x 0,1 CFU/ml Keterangan:
n = jumlah koloni yang terdapat pada tabung seri pengenceran ke-x
Rancangan dan Analisis Data Perlakuan
Perlakuan diterapkan untuk meningkatkan penggunaan ransum yang berbasis jerami padi dengan penambahan SKN. Penelitian ini menggunakan 4 perlakuan pakan (faktor A), yaitu :
R1 = Jerami Padi (100%)
R2 = Jerami Padi (82,78%) + Dedak Padi (17,22%)
R3 = Jerami Padi (80,39%) + Dedak Padi (16,72% ) + Suplemen Kaya Nutrien (2,89%)
R4 = Ransum Komplit (100%)
Keempat ransum perlakuan tersebut diinkubasikan pada waktu inkubasi (faktor B)1, 3 dan 5 jam.
Model
27 Yijk = µ + τi + αj+k + jk +ijk
Keterangan :
Yijk = nilai pengamatan kelompok ke-i, perlakuan ke-j = nilai rataan umum
i = pengaruh kelompok (cairan rumen) ke-i αj = pengaruh perlakuan ransum ke-j
k = pengaruh perlakuan waktu inkubasi ke-k
jk = pengaruh interaksi perlakuan ransum ke-j dan waktu inkubasi ke-k
ijk = galat percobaan untuk kelompok ke-i, perlakuan ransum ke-jdan perlakuan
waktu inkubasi ke-k
Peubah
Peubah yang diamati dalam penelitian ini adalah sebagai berikut : 1. Populasi protozoa total
28 HASIL DAN PEMBAHASAN
Kandungan Zat Makanan Ransum Percobaan
Semua bahan pakan yang digunakan di dalam ransum, SKN dan pakan komplit berasal dari bahan yang mudah ditemukan dan dibudidayakan. Kualitas nutrisi jerami padi yang digunakan dalam penelitian ini cukup rendah terutama kandungan proteinnya hanya 4,15% BK dan kandungan serat kasarnya yang tinggi sebesar 32,5% BK. Kandungan PK jerami padi yang digunakan dalam penelitian ini (Tabel 11) lebih rendah dibandingkan dengan hasil penelitian yang dilaporkan Prasetiyono et al. (2007) yaitu sebesar 5,06% BK. Lebih rendahnya kandungan protein jerami padi dalam penelitian ini dapat disebabkan oleh waktu panen, musim, dan kesuburan tanah yang berbeda.
Tabel 11. Kandungan Zat Makanan Pakan Penelitian
Kandungan Kimia Bahan Jerami Padi Dedak Padi SKN Konsentrat*
Kadar Air (%) 12,50 9,00 31,26 22,09
Sumber : Hasil analisis laboratorium PAU IPB (2011)
*=Hasil analisis Laboratorium Nutrisi Ternak Perah (2011)
Keterangan : 1) TDN dihitung dengan rumus TDN = 25,6+0,53 PK+1,7 L – 0,474 SK +0,732 BETN
29 fisik, kimiawi, maupun melalui perlakuan suplementasi dengan bahan makanan lain yang mampu meningkatkan daya guna jerami padi sebagai pakan ternak, sehingga ransum komplit perlu ditambahkan dengan pakan penguat seperti SKN.
SKN dibuat untuk mensubstitusi dan mengkoreksi zat-zat makanan jerami padi atau konsentrat komersial yang ketersediaannya kurang di dalam ransum sehingga terjadi perbaikan nutrisi pada pakan berbasis jerami padi. Keadaan ini diharapkan dapat meningkatkan produktivitas ternak sehingga mampu meningkatkan keuntungan bagi peternak. Hal ini dikarenakan SKN merupakan sumber protein, energi dan mineral yang memiliki keserasian dan keseimbangan gizi baik jumlah protein (14,62% BK) dan energi TDN (54,41% BK) (Tabel 11) yang cukup untuk memenuhi kebutuhan ternak.
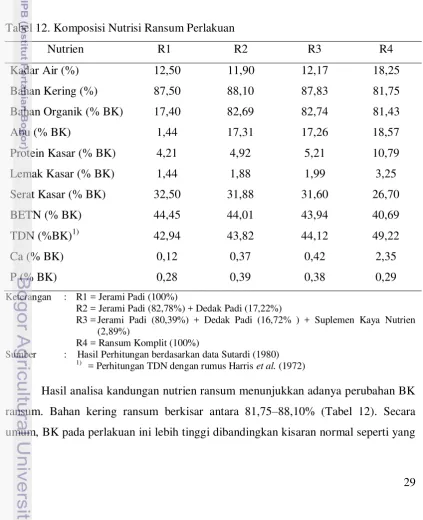
Tabel 12. Komposisi Nutrisi Ransum Perlakuan
Nutrien R1 R2 R3 R4
= Perhitungan TDN dengan rumus Harris et al. (1972)
30 disebutkan Prasetiyono et al. (2007) yaitu 76,93%. Hal tersebut dapat diakibatkan sampel yang digunakan pada penelitian ini adalah sampel kering.
Kandungan abu dalam penelitian ini berkisar antara 17,40–18,57% BK. Kandungan abu tertinggi terdapat pada ransum komplit (R4) yang diduga disebabkan oleh penggunaan molases. Molases mengandung abu sebesar 10,4% (Tillman et al.,1997) sehingga meningkatkan kandungan abu ransum komplit.Sedangkan SK semakin menurun seiring dengan berkurangnya kandungan jerami padi dalam ransum sehingga menyebabkan kandungan bahan organik (BO) semakin bertambah kecuali pada R4. Sebaliknya, kandungan LK pada ransum ini meningkat seiring berkurangnya kandungan jerami padi. Kandungan PK dan TDN dalam penelitian ini masing-masing berkisar antara 4,21–10,79% BK dan 42,94–49,22% BK.
Penggunaan suplemen berupa dedak padi dan SKN (R3) yang diberikan secara terpisah dapat meningkatkan kadar PK ransum berbasis jerami padi. Peningkatan yang lebih besar dalam kadar PK ransum terjadi pada pakan komplit (R4). Kandungan PK ransum penelitian meningkat seiring dengan berkurangnya kandungan jerami padi dalam ransum dan meningkatnya penggunaan SKN. Penurunan kadar jerami padi dalam pakan komplit (R4) dapat memperbaiki kandungan nutrisi ransum dan cukup untuk memenuhi kebutuhan nutrisi ternak sapi pedaging dengan bobot badan 500 kg yaitu untuk PK yang berkisar antara 8–9%, namun belum memenuhi untuk kebutuhan TDN sebesar 61% (Parakkasi,1999).
Peningkatan protein ransum ini disebabkan oleh kandungan PK dalam SKN (14,62% BK) lebih tinggi dibandingkan dengan kandungan PK dalam dedak padi (13,00% BK). Tingginya kandungan PK dalam SKN dikarenakan terdapatnya pakan sumber-sumber protein yang digunakan dalam SKN seperti tepung daun (daun singkong, daun lamtoro, daun turi), dan tepung ikan. Tepung daun dan tepung ikan merupakan pakan sumber protein yang ideal bagi ruminansia karena merupakan protein tidak terdegradasi atau Rumen Undegradable Protein (RUP). Sumber protein ideal bagi ruminansia adalah yang dapat menunjang sintesis protein mikroba yang optimal, sebagian dapat terhindar dari degradasi, dan mempunyai nilai biologis yang tinggi (Lu et al., 1982).
31 diduga sebagai akibat dari komposisi jerami padi pada ransum tersebut sebesar 40% lebih rendah dibandingkan perlakuan yang lain yang komposisi jerami padinya di atas 80%. Penurunan kadar SK tersebut menyebabkan kecernaan dan nilai penggunaan ransum R3 dan R4 meningkat.
Populasi Mikroba Rumen
Dalam penelitian ini, analisis in vitro di laboratorium bertujuan untuk mempelajari efektifitas penambahan SKN dalam ransum komplit ditinjau dari populasi bakteri dan protozoa rumen. Analisis populasi mikroba mencerminkan banyaknya biomassa mikroba yang merupakan sumber protein yang berkualitas tinggi bagi hewan induk semang.
Populasi Protozoa
Protozoa berperan dalam pola fermentasi rumen dengan cara mencerna partikel-partikel pati dengan lambat sehingga kadar asam lemak atsiri rendah, selain itu protozoa juga memangsa bakteri untuk memenuhi kebutuhan hidupnya karena kemampuan yang sangat rendah untuk mensintesis asam amino dan vitamin B komplek (Arora, 1989).
Hasil sidik ragam menunjukkan bahwa perlakuan ransum tidak mempengaruhi populasi protozoa, demikian juga dengan perlakuan waktu inkubasi maupun interaksi keduanya. Populasi rataan protozoa dari ransum R1, R2, R3, dan R4 secara umum lebih rendah dibandingkan blanko (Tabel 13).
32 Tabel 13. Populasi Protozoa Rumen dari Ransum Berbasis Jerami Padi pada Waktu
Inkubasi yang Berbeda
Rataan±SD 2,81±0,70 1,79±0,50 1,63±0,61 1,53±0,55 0,65±0,57 1,68±0,10
Keterangan : R1 = Jerami Padi (100%)
R2 = Jerami Padi (82,78%) + Dedak Padi (17,22%)
R3 = Jerami Padi (80,39%) + Dedak Padi (16,72% ) + Suplemen Kaya Nutrien (2,89%)
R4 = Ransum Komplit (100%)
Penurunan populasi protozoa juga berkaitan dengan produksi VFA total. Pada waktu inkubasi 3 jam populasi protozoa lebih tinggi dari 5 jam. Hasil ini sejalan dengan hasil rataan konsentrasi VFA yang dilaporkan oleh Saputra (2011) dimana produksi VFA akan meningkat pada 3 jam inkubasi sebagai akibat dari pertumbuhan mikroba yang sudah mulai meningkat. Pertumbuhan mikroba yang sudah baik, khususnya protozoa akan berpengaruh terhadap proses fermentasi pati menjadi VFA.
Bakteri Total
Mikroorganisme yang ada di dalam rumen dapat hidup dan melakukan aktivitasnya apabila kondisi lingkungannya mendukung. Populasi bakteri rumen total merupakan salah satu cerminan metabolisme yang terjadi di dalam rumen. Bakteri dapat hidup optimum pada pH 5,5–7,0 dalam kondisi tanpa oksigen, suhu antara 39– 40 ºC dan adanya produk fermentasi pada konsentrasi sedang (Hungate, 1966).
33
Rataan±SD 5,24±0,17 5,64±0,65 5,44±0,53 5,51±0,84 5,15±0,61 5,39±0,16
Keterangan : R1 = Jerami Padi (100%)
R2 = Jerami Padi (82,78%) + Dedak Padi (17,22%)
R3 = Jerami Padi (80,39%) + Dedak Padi (16,72% ) + Suplemen Kaya Nutrien (2,89%)
R4 = Ransum Komplit (100%)
Populasi bakteri total tidak berbeda nyata antar perlakuan dalam ransum. Ditinjau dari populasi bakterinya, cairan rumen segar yang dicobakan hanya memiliki 105 koloni/ml, padahal menurut Kamra (2005), bakteri dalam rumen dapat mencapai 1010–1011 koloni/ml. Hal tersebut dapat terjadi karena beberapa faktor :
(1) Cairan rumen yang digunakan sebagai sampel pada penelitian berasal dari dua rumah potong hewan yang berbeda sebagai akibat daripemberian pakan dan penanganan ternak sebelum pemotongan yang mempengaruhi kondisi cairan rumen dan populasi mikroba rumen. Seperti yang diungkapkan Kamra (2005) bahwa populasi mikroba dalam ekosistem rumen sangat bervariasi antar individu ataupun jenis hewan, tergantung jenis makanan yang diberikan karena hal itu akan berpengaruh terhadap efisiensi ruminan dalam menggunakan makanannya;
34 (3) Mikroba rumen dapat mentolerir antinutrisi dalam ransum. Zat antinutrisi seperti tanin dan saponin disintesis oleh tanaman untuk melindungi diri dari serangan mikroba, oleh karena itu zat antinutrisi ini memiliki aktivitas antimikroba (Kamra, 2005). Saponin mempunyai kemampuan untuk membentuk ikatan dengan kolesterol yang ada pada membran sel eukariotik, tetapi tidak pada sel prokariotik, sehingga adanya saponin hanya akan mempengaruhi populasi protozoa tanpa menghambat pertumbuhan bakteri (Wina et al., 2005c). Sedangkan tanin merupakan senyawa poliphenolic yang mampu mengikat protein dan membentuk senyawa komplek (Makkar, 2003). Secara umum tanin mempunyai pengaruh menurunkan penggunaan pakan secara in vivo, terutama penggunaan protein dan menurunkan berbagai aktivititas enzim. Selain itu tanin juga menurunkan serangan mikrobial terhadap pakan (Suhartati, 2005);
(4) Sumber protein pada ransum komplit dan SKN merupakan protein tidak terdegradasi atau Rumen Undegradable Protein (RUP) di rumen yang berasal dari tepung ikan dan tepung daun (daun singkong, lamtoro, dan turi). Proses pengolahan tepung daun dan tepung ikan dengan cara pengeringan mengakibatkan protein mengalami denaturasi atau terjadi perubahan struktur alaminya (pemanasan mengakibatkan protein membuka struktur aslinya) yang mengakibatkan terjadinya perubahan kimiawi dan kelarutan proteinnya (McDonald et al., 1982) sehingga protein sulit didegradasi yang mengakibatkan konsentrasi NH3 yang dihasilkan rendah.
Lama waktu inkubasi juga tidak mempengaruhi populasi bakteri total. Secara umum terjadi penurunan jumlah populasi pada waktu inkubasi 3 jam. Pertumbuhan bakteri secara umum melalui empat tahap yaitu tahap persiapan, logaritmik (pertumbuhan yang meningkat tajam), stasioner (pertumbuhan yang kecil), dan tahap kematian dimana populasi akan menurun (Ermentina, 1998). Akan tetapi grafik pertumbuhan yang dimiliki bakteri total tidak menunjukkan kecenderungan yang normal. Church (1979) menyatakan bahwa makanan adalah faktor terpenting yang mempengaruhi jumlah dan proporsi relatif dari spesies yang berbeda dalam rumen. Mikroba akan beradaptasi dengan substratnya untuk mencapai keseimbangan baru.
35 dapat tumbuh sebagaimana seharusnya. Akan tetapi ketika harus mencerna makanan yang sulit dicerna seperti dinding sel yang berkadar serat tinggi, bakteri harus melakukan adaptasi. Pada masa adaptasi ini, ada bakteri yang mampu beradaptasi dan adapula yang gagal. Semakin tinggi jumlah bakteri yang tidak mampu beradaptasi makin tajam penurunan populasi yang terjadi. Bakteri yang mampu beradaptasi ini akan berkembang kembali dan membentuk tahap-tahap pertumbuhan lagi (Ermentina, 1998).
Bakteri Proteolitik
Keberadaan mikroba rumen berperan dalam pemecahan pakan melalui proses fermentasi menyebabkan ternak ruminansia mampu mencerna pakan serat yang berkualitas rendah menjadi sesuatu yang dapat dimanfaatkan atau mempunyai nilai lebih sehingga kebutuhan asam-asam amino untuk nutrisi protein tidak sepenuhnya tergantung pada kualitas protein pakan yang diberikan (Sutardi, 1980).
Hasil sidik ragam menunjukkan bahwa perlakuan ransum tidak mempengaruhi populasi bakteri proteolitik, demikian juga dengan perlakuan waktu inkubasi maupun interaksi keduanya ( Tabel 15).
36 Tabel 15. Populasi Bakteri Proteolitik dari Ransum Berbasis Jerami Padi pada
Waktu Inkubasi yang Berbeda
Rataan±SD 4,81±0,34 5,48±0,35 5,37±0,68 4,78±0,45 5,61±0,37 5,21±0,15
Keterangan : R1 = Jerami Padi (100%)
R2 = Jerami Padi (82,78%) + Dedak Padi (17,22%)
R3 = Jerami Padi (80,39%) + Dedak Padi (16,72% ) + Suplemen Kaya Nutrien (2,89%)
R4 = Ransum Komplit (100%)
Lama waktu inkubasi juga tidak mempengaruhi populasi bakteri proteolitik. Pada waktu inkubasi1 jam, populasi bakteri proteolitik hampir sama untuk semua ransum perlakuan. Kecuali pada R4, populasi kemudian menurun pada waktu inkubasi 3 jam, dan meningkat saat waktu inkubasi 5 jam. Populasi bakteri proteolitik R1 mempunyai pola yang berseberangan dengan R4. Hal ini kemungkinan karena bakteri mengalami fase pertumbuhan yang berbeda sebagai akibat jenis dan ketersediaan makanan saat kondisi fermentasi, dan mikroba akan beradaptasi dengan substratnya untuk mencapai keseimbangan baru (Church, 1979; Ermentina, 1998). Penurunan populasi bakteri proteolitik ini berkorelasi negatif dengan konsentrasi NH3 dan VFA yang dilaporkan oleh Saputra (2011), semakin sedikit populasi bakteri proteolitik, konsentrasi NH3 dan VFA yang dihasilkan justru semakin banyak. Hal ini disebabkan VFA tidak digunakan oleh NH3 untuk sintesis protein mikroba.
Bakteri Amilolitik
37 Tabel 16. Populasi Bakteri Amilolitik dari Ransum Berbasis Jerami Padi pada
Waktu Inkubasi yang Berbeda
Rataan±SD 4,99±0,21 5,67±0,41 5,45±0,60 5,22±0,77 5,81±0,29 5,43±0,23
Keterangan : R1 = Jerami Padi (100%)
R2 = Jerami Padi (82,78%) + Dedak Padi (17,22%)
R3 = Jerami Padi (80,39%) + Dedak Padi (16,72% ) + Suplemen Kaya Nutrien (2,89%)
R4 = Ransum Komplit (100%)
Tidak terdapat perbedaan yang nyata dalam populasi bakteri amilolitik antar perlakuan dalam ransum walaupun populasi bakteri amilolitik semakin meningkat seiring dengan berkurangnya komposisi jerami padi dalam ransum. Beberapa faktor dapat mempengaruhi kondisi tersebut seperti (1) berbedanya cairan rumen yang digunakan sebagai akibat perbedaan ternak, pakan dan penanganan ternak sebelum pemotongan seperti yang dikemukakan oleh Kamra (2005); (2) penyediaan dedak padi (R2), SKN (R3) dan pakan komplit (R4) masih belum dapat menstimulasi pertumbuhan bakteri amilolitik; (3) penggunaan tepung daun dalam SKN yang mengandung berbagai antinutrisi dapat menghambat pertumbuhan bakteri amilolitik; dan (4) ketersediaan amonia yang terbatas sebagai sumber N untuk sintesis protein bakteri amilolitik sebagai akibat dari proses pengeringan tepung daun dan tepung ikan.
38 sedikit perbaikan dari penggunaan jerami padi. Penggunaan dedak padi sebagai suplemen (R2) juga menurunkan populasi bakteri amilolitik pada 3 jam waktu inkubasi yang tetap konstan pada 5 jam waktu inkubasi. SKN sebagai suplemen dalam R3 menurunkan populasi bakteri amilolitik dari waktu inkubasi 1 jam ke 3 jam, tetapi sedikit meningkatkan pada waktu inkubasi 5 jam. Penggunaan SKN yang semakin meningkat dan berkurangnya pemakaian jerami padi dalam R4 dapat menstimulasi pertumbuhan populasi bakteri amilolitik dari waktu inkubasi 1 jam ke 3 jam, lalu sedikit menurun pada waktu inkubasi 5 jam. Fluktuasi yang terjadi dalam populasi bakteri amilolitik tersebut berkaitan dengan jenis makanan, kemampuan mikroba beradaptasi dengan substrat yang tersedia dan karakteristik pertumbuhan mikroba (Church, 1979; Ermentina, 1998).
Bakteri Selulolitik
Bakteri selulolitik mempunyai enzim yang mampu menghancurkan karbohidrat yang sulit dicerna, seperti selulosa dan hemiselulosa. Berdasarkan hasil sidik ragam, populasi bakteri selulolitik dipengaruhi oleh waktu inkubasi (P<0,05), namun tidak dipengaruhi oleh perlakuan ransum maupun interaksi keduanya.
Populasi bakteri selulolitik dipengaruhi oleh waktu inkubasi (P<0,05). Uji kontras ortogonal menunjukkan bahwa populasi bakteri selulolitik pada waktu inkubasi 1 jam nyata (P<0,05) lebih tinggi dari waktu inkubasi 3 dan 5 jam (Tabel 17).
Tabel 17. Populasi Bakteri Selulolitik dari Ransum Berbasis Jerami Padi pada Waktu Inkubasi yang Berbeda
39
Superskrip yang berbeda pada baris yang sama berbeda pada (P<0,05)
Hasil uji kontras ortogonal menunjukkan hubungan populasi bakteri selulolitik dengan waktu inkubasi mengikuti persamaan polinomial Y= 1,187x2 – 7,903x + 36,41. Dari persamaan tersebut diketahui bahwa populasi bakteri selulolitik mengalami penurunan pada waktu inkubasi 3 jam. Hal ini tidak sejalan dengan konsentrasi VFA yang meningkat pada 3 jam inkubasi. Church (1979) menyatakan bahwa konsentrasi VFA total sangat berhubungan dengan populasi mikroba rumen, khususnya bakteri selulolitik dan amilolitik. Tingginya populasi bakteri selulolitik pada waktu inkubasi 1 jam disebabkan tingginya populasi bakteri selulolitik dalam cairan rumen. Bakteri selulolitik mulai beradaptasi terhadap pakan pada waktu inkubasi 3 jam sehingga terjadi penurunan. Adanya kandungan serat kasar yang tinggi pada jerami padi tidak menjamin populasi bakteri selulolitik yang tinggi. Hal ini dikarenakan bakteri selulolitik memiliki fase pertumbuhan yang lambat, meskipun zat makanan yang dibutuhkan tersedia. Hal ini sebagaimana ditunjukkan oleh data populasi bakteri selulolitik yang meningkat pada waktu inkubasi 5 jam. Fase pertumbuhan bakteri selulolitik yang lambat juga didukung oleh penelitian yang dilakukan Ulya (2007) dimana populasi bakteri selulolitik akan meningkat pada waktu inkubasi 9 dan 12 jam.
40 KESIMPULAN DAN SARAN
Kesimpulan
Populasi bakteri selulolitik lebih dipengaruhi oleh waktu inkubasi, sementara itu pemakaian suplemen kaya nutrien dan pakan komplit pada ransum berbasis jerami padi belum dapat memberikan pengaruh yang baik terhadap populasi protozoa dan bakteri rumen.
Saran