IDENTIFIKASI Ganoderma spp. MENGGUNAKAN TEKNIK
DNA BARCODING
DHANIAR ASTRI
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
ABSTRAK
DHANIAR ASTRI. Identifikasi
Ganoderma
spp. Menggunakan Teknik
DNA
Barcoding
. Dibimbing oleh SURYANI dan HAYATI MINARSIH.
Kelapa sawit merupakan salah satu komoditas perkebunan yang sangat
penting. Salah satu masalah yang dihadapi pada perkebunan kelapa sawit saat ini
adalah masalah serangan penyakit, yaitu Busuk Pangkal Batang (BPB) akibat
serangan
Ganoderma
spp.
Jika tanaman kelapa sawit sudah terserang BPB, maka
cepat atau lambat tanaman akan mati dan mengalami penurunan produksi secara
nyata. Dalam upaya pengendalian serangan Ganoderma yang efektif, maka
diperlukan suatu teknik pengendalian serangan Ganoderma melalui identifikasi
Ganoderma spp.
yang lebih akurat. Tujuan penelitian ini adalah mencari primer
spesifik untuk identifikasi
Ganoderma
spp. yang menyerang tanaman perkebunan
dan tanaman pelindungnya. Metode dilakukan secara molekuler dengan
menggunakan teknik
DNA Barcoding
. Berdasarkan hasil penelitian yang telah
dilakukan, diketahui bahwa pasangan primer ITS1_F/ITS4_B memiliki presentase
kemampuan yang tinggi dalam mengidentifikasi isolat
Ganoderma
spp.
dibandingkan dengan pasangan primer yang lain, yaitu pasangan primer
BenA_F/ BenA_R, Gan_ITS1_F/Gan_ITS1_R, CO1_PenF1/CO1_PenR1,
CO1_B_F/CO1_B_R, dan Gan_ITS2_F/Gan_ITS2_R. Pasangan primer
ITS1_F/ITS4_B memiliki potensi sebagai
DNA barcode
untuk identifikasi
spesies-spesies Ganoderma.
ABSTRACT
DHANIAR ASTRI. Identification of
Ganoderma
spp. using DNA Barcoding
Technique. Under the direction of SURYANI and HAYATI MINARSIH
Oil palm is one of comodity plantation which is very important. One of the
problems faced by the oil palm plantations are disease, namely Basal Stem Rot
(BSR) which caused by
Ganoderma
spp. If plants were attacked by BSR, sooner
or later, they would die and the production will decreased significantly.
Consequently, in order to have an effective control of Ganoderma, an accurate
identification is needed. This could be achieved by molecular techniques through
DNA Barcoding method. The aim of this study was to is to find a specific primer
for the identification of
Ganoderma
spp. that attack estate crops and their shade
trees. Based on this research, it showed that the primer pair ITS1_F/ITS4_B has a
high percentage of the ability to identify isolates of
Ganoderma
spp. compared
with the other primer pairs, which are primer BenA F/ BenA_R,
Gan_ITS1_F/Gan_ITS1_R, CO1_PenF1/CO1_PenR1, CO1_B_F/ CO1_B_R, and
Gan_ITS2_F/Gan_ITS2_R. The primer pair ITS1_F/ITS4_B had a potential as a
DNA barcode for identification of Ganoderma species.
IDENTIFIKASI Ganoderma spp. MENGGUNAKAN TEKNIK
DNA BARCODING
DHANIAR ASTRI
Skripsi
sebagai salah satu syarat memperoleh gelar
Sarjana Sains pada
Departemen Biokimia
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
Judul Skripsi
: Identifikasi
Ganoderma
spp. Menggunakan Teknik
DNA
Barcoding
Nama
: Dhaniar Astri
NIM
: G84070036
Disetujui
Komisi Pembimbing
Dr. Suryani, M.Sc
Dr. Ir. Hayati Minarsih, M.Sc
Ketua
Anggota
Diketahui,
Dr. Ir. I Made Artika, M.App.Sc.
Ketua Departemen Biokimia
PRAKATA
Alhamdulillah, puji dan syukur penulis panjatkan ke hadirat Allah SWT
atas karunia dan hidayah-Nya sehingga penulis dapat menyelesaikan penelitian ini
dengan baik. Penelitian ini berjudul Identifikasi
Ganoderma
spp. Menggunakan
Teknik
DNA Barcoding
. Kegiatan penelitian dilaksanakan dari bulan April hingga
Oktober 2011 dan bertempat di Laboratorium Biologi Molekuler dan Rekayasa
Genetika, Balai Penelitian Bioteknologi Perkebunan Indonesia (BPBPI), Jalan
Taman Kencana No.1 Bogor.
Penulis mengucapkan terima kasih kepada semua pihak yang telah
membantu selama kegiatan penelitian ini berlangsung, yaitu Dr. Suryani, M.Sc
selaku pembimbing utama dan Dr. Ir. Hayati Minarsih, M.Sc selaku pembimbing
lapangan yang telah memberikan saran, kritik, dan bimbingannya. Penulis juga
mengucapkan terima kasih kepada Mbak Niyyah Fitranty serta segenap staf di
Laboratorium Biologi Molekuler dan Rekayasa Genetika, Balai Penelitian
Bioteknologi Perkebunan Indonesia atas peran dan kerjasamanya dalam
membantu pelaksanaan penelitian ini. Ucapan terima kasih juga penulis tujukan
kepada orang tua, kakak, Eka Febrianti, Jatu Rukmi Mulyaningtyas, Ayu Arthuria
Rizqiyanti atas doa, dukungan, dan bantuan bagi penulis.
Bogor, Maret 2012
RIWAYAT HIDUP
Penulis dilahirkan di Jakarta, 5 Desember 1988 dari ayah Ateng Zaelani dan
ibu Sri Wahyuni. Penulis merupakan anak kedua dari dua bersaudara.
Pendidikan penulis dimulai dari SDN Larangan Utara 03 Tangerang,
kemudian dilanjutkan pendidikan ke SMPN 206 Jakarta Barat. Pada tahun 2007,
penulis lulus dari SMAN 90 Jakarta Selatan dan pada tahun yang sama lulus
seleksi masuk IPB melalui jalur Undangan Seleksi Masuk IPB (USMI). Penulis
memilih mayor Departemen Biokimia, Fakultas Matematika dan Ilmu
Pengetahuan Alam.
Selama mengikuti perkuliahan, penulis aktif dalam beberapa organisasi,
diantaranya menjadi salah satu staf divisi Bioanalisis dalam Himpunan Profesi
Community Research and Education of Biochemistry
(CREBs) pada tahun
2008/2009 dan bendahara umum Dewan Perwakilan Mahasiswa (DPM) FMIPA
IPB pada tahun 2009/2010. Penulis juga aktif mengikuti kepanitiaan, diantaranya,
staf divisi Publikasi, Dokumentasi, dan Dekorasi (PDD) Seminar Kesehatan dan
Keselamatan Kerja (K3) Laboratorium Biokimia IPB pada tahun 2008, ketua
divisi Publikasi, Dokumentasi, dan Dekorasi (PDD) Seminar Kanker IPB pada
tahun 2009, sekretaris Sidang Umum Dewan Perwakilan Mahasiswa (DPM)
FMIPA IPB pada tahun 2009, serta bendahara Pemilihan Raya Dewan Perwakilan
Mahasiswa (DPM) IPB pada tahun 2010.
DAFTAR ISI
Halaman
DAFTAR TABEL ... viii
DAFTAR GAMBAR ... ix
DAFTAR LAMPIRAN ... ix
PENDAHULUAN ... 1
TINJAUAN PUSTAKA
Ganoderma
spp
.
... 1
DNA Barcoding
... 2
Isolasi DNA ... 3
Polymerase Chain Reaction (PCR) ... 3
Teknik Rekombinasi DNA ... 4
BAHAN DAN METODE
Bahan dan Alat ... 4
Metode Percobaan ... 5
HASIL DAN PEMBAHASAN
Kultivasi Isolat Murni
Ganoderma
spp. dari Tubuh Buah ... 9
Kualitas dan Kuantitas DNA Kromosom
Ganoderma
spp. ... 9
Desain Primer, Optimasi Primer terhadap Isolat
Ganoderma
spp. dan
Amplifikasi DNA dengan Metode PCR ... 10
Ekstraksi (Elusi), Purifikasi Fragmen DNA dan Pengklonan Gen ... 11
PCR Koloni ... 12
Plasmid Rekombinan ... 12
Analisis Sekuens (
sequencing
) ... 13
SIMPULAN DAN SARAN
Simpulan ... 15
Saran ... 15
DAFTAR PUSTAKA ... 15
DAFTAR TABEL
Halaman
1 Primer yang digunakan untuk amplifikasi sampel DNA
Ganoderma
spp. ... 7
2 Data pengukuran nilai absorbansi DNA dengan spektrofotometer UV-VIS
pada panjang gelombang 260 dan 280 nm serta konsentrasi DNA . ... 10
3 Primer yang berhasil mengidentifikasi isolat
Ganoderma
spp ... 14
DAFTAR GAMBAR
Halaman
1 Tubuh buah
Ganoderma
spp. ... 2
2 Miselium isolat
Ganoderma
spp. pada media PDA ... 9
3 Elektroforegram kualitas DNA hasil isolasi ... 10
4 Elektroforegram hasil optimasi primer BenA_F/BenA_R ... 10
5 Elektroforegram hasil optimasi primer ITS1_F/ITS4_B ... 11
6 Elektroforegram hasil optimasi primer Gan_ITS1_F / Gan_ITS1_R ... 11
7 Elektroforegram hasil amplifikasi dengan primer BenA_F/BenA_R,
ITS1_F/ITS4_B, dan Gan_ITS1_F/Gan_ITS1_R ... 11
8 Elektroforegram hasil PCR koloni primer BenA_F/BenA_R ... 12
9 Elektroforegram hasil PCR koloni primer ITS1_F/ITS4_B ... 12
10 Elektroforegram hasil PCR koloni primer Gan_ITS1_F/Gan_ITS1_R ... 12
11 Elektroforegram hasil isolasi plasmid rekombinan primer BenA F/R ... 13
12 Elektroforegram hasil isolasi plasmid rekombinan primer ITS1_F/ITS4_B 13
13 Elektroforegram hasil isolasi plasmid rekombinan primer Gan_ITS1_F/
Gan_ITS1_R ... 13
DAFTAR LAMPIRAN
Halaman
1 Skema isolasi DNA ... 19
2 Isolat
Ganoderma
spp. dan primer yang digunakan untuk
sequencing
. ... 20
3 Peta marka seleksi vektor pGEM T-Easy ... 21
4 Hasil pengurutan basa nukelotida (
sequencing
) isolat
Ganoderma
spp. .... 22
5 Hasil analisis identifikasi sekuen
Ganoderma
spp. menggunakan program
BLAST ... 26
PENDAHULUAN
Kelapa sawit merupakan salah satu komoditas perkebunan yang sangat penting. Masalah yang dihadapi di perkebunan kelapa sawit saat ini adalah masalah serangan penyakit.. Salah satu penyakit penting yang menjangkiti pohon kelapa sawit adalah penyakit Busuk Pangkal Batang (BPB) akibat serangan Ganoderma spp. Pohon kelapa sawit yang diserang penyakit ini menyebabkan kerugian dari dua segi, yaitu mengurangi jumlah tandan dan berat buah serta mematikan pohon kelapa sawit tersebut (Ditjen Perkebunan 2011). Menurut Taniwiryono (2011), serangan penyakit BPB pada perkebunan kelapa sawit khususnya di wilayah Sumatera Utara sudah berada pada kondisi yang mengkhawatirkan. Berdasarkan contoh kasus sensus yang dilakukan pada salah satu perkebunan kelapa sawit di wilayah Sumatera, serangan penyakit BPB mencapai 50% dalam satu hektar lahan. Tanaman kelapa sawit yang terserang berumur 14 tahun pada generasi ke-3 dan ke-4. Jika tanaman kelapa sawit sudah terserang BPB maka cepat atau lambat tanaman akan menjumpai kematiannya.
Ganoderma spp. merupakan jamur yang dapat membentuk tubuh buah. Ganoderma
spp. diklasifikasikan ke dalam filum Fungi, kelas Basidiomycetes, subkelas Homobasidiomycetes, ordo Agaricales, famili Polyporaceae, genus Ganoderma, dan spesies Ganodermaboninense (Alexopoulos
et al. 1996). Penyakit BPB yang disebabkan oleh serangan Ganoderma spp. pada kelapa sawit mampu mengakibatkan kematian tanaman lebih dari 80% populasi tanaman pada satu hamparan (Taniwiryono 2011). Sementara itu, serangan Ganoderma spp. baru diketahui ketika tingkat infeksi sudah kritis dan tanaman sudah sulit diselamatkan (Mohd Su’ud et al. 2007; Minarsih et al.
2011). Kondisi inilah yang menjadikan penyakit BPB pada kelapa sawit sebagai penyakit terpenting yang harus segera dikendalikan (Taniwiryono 2011). Hal yang paling merugikan lagi adalah serangan
Ganoderma spp. yang terjadi pada tanaman sangat sulit dideteksi. Hal ini disebabkan oleh gejala yang mirip dengan serangan penyakit perakaran lainnya termasuk juga mirip dengan gejala kekeringan (Mohd Su’ud et al. 2007; Minarsih et al. 2011). Berdasarkan kenyataan-kenyataan yang terjadi di atas, maka diperlukan suatu teknik pengendalian serangan Ganoderma yang
efektif melalui analisis identifikasi
Ganoderma spp. secara molekuler. Analisis identifikasi Ganoderma spp. yang tumbuh pada tanaman perkebunan, seperti kakao, kopi, beserta tanaman pelindungnya dari berbagai wilayah di Indonesia, dilakukan melalui pendekatan molekuler dengan menggunakan teknik DNA Barcoding. Teknik DNA Barcoding dapat mengidentifikasi dan membedakan suatu organisme mulai tahap spesies hingga sub spesies. Keunggulan teknik DNA Barcoding, yaitu dapat digunakan untuk identifikasi dan karakterisasi berbagai spesies yang tidak dapat dibedakan secara morfologi (Tudge 2000). Selain itu, teknik DNA Barcoding
juga dapat digunakan untuk identifikasi suatu organisme walaupun DNA dari organisme tersebut tidak dalam bentuk murni atau utuh, bahkan DNA yang sudah mengalami degradasi dan proses pengolahan pun dapat digunakan untuk analisis DNA Barcoding (Hajibabaei et al. 2006).
Tujuan penelitian ini adalah mengidentifikasi Ganoderma spp. menggunakan teknik DNA Barcoding. Sementara itu, hipotesis mengenai penelitian ini, yaitu dimungkinkan memperoleh pasangan primer spesifik sebagai DNA
Barcode yang dapat digunakan untuk
mengidentifikasi spesies Ganoderma. Penelitian mengenai identifikasi Ganoderma
spp. menggunakan teknik DNA Barcoding
juga bermanfaat dalam mencari primer yang spesifik sebagai DNA Barcode untuk identifikasi isolat Ganoderma spp. sehingga diharapkan dapat memecahkan permasalahan mengenai identifikasi dan karakterisasi berbagai spesies Ganoderma spp. yang menyebabkan BPB di perkebunan kelapa sawit di Indonesia. Identifikasi spesies Ganoderma spp. ini merupakan langkah awal dalam pengembangan teknologi perlindungan tanaman perkebunan dan tanaman pelindungnya dari serangan
Ganoderma spp. yang terjadi di Indonesia.
TINJAUAN PUSTAKA
Ganoderma spp.
Ganoderma spp. dikenal sebagai Ling Zhi di Cina dan Reishi di Jepang.
mengenai Ganoderma spp. baru berlangsung sekitar 25 tahun terakhir (Suryanto 2003). Menurut Alexopoulos et al. (1996)
Ganoderma spp. diklasifikasi ke dalam filum fungi, kelas Basidiomycetes, subkelas Homobasidiomycetes, ordo Agaricales, famili Polyporaceae, genus Ganoderma, dan spesies Ganoderma spp. Ganoderma spp. terdiri atas tubuh buah yang tebal, bergabus, berwarna kuning kemerahan pada awalnya dan berubah menjadi warna cokelat ketika telah masak (Chang & Miles 2004 dalam Sutiarna 2010). Fungi Ganoderma spp. merupakan anggota Basidiomycetes yang menyebabkan penyakit pada tanaman keras dengan kemampuan menguraikan lignin, selulosa, dan polisakarida (Widyastuti 2007).
Ganoderma spp. membentuk tubuh buah berupa piringan keras (Gambar 1) yang menempel pada pangkal batang tanaman, permukaan atas berwarna cokelat kemerahan sampai cokelat tua, licin dan tampak mengkilat seperti dilapisi oleh lilin, bagian tepinya yang aktif tumbuh berwarna lebih muda kemudian akan berubah warna ketika telah tua. Bagian permukaan bawahnya berwarna putih kekuningan yang tersusun dari jutaan pori-pori. Pada bagian inilah terdapat ribuan basidia yang tumbuh dan menghasilkan jutaan spora secara serentak. Spora yang diterbangkan oleh angin dan dibawa terbang oleh serangga diduga memiliki peranan penting dalam penularan penyakit dari tanaman satu ke tanaman lain pada jarak yang melebihi jarak perkembangan sistem perakaran, dan penularan penyakit dari tanaman yang sakit ke tanaman sehat yang berada di dekatnya. Hal ini terjadi melalui kontak akar (Taniwiryono & Panji 1999).
Ciri-ciri lain Ganoderma spp. adalah memiliki sebuah lapisan himenium yang terdiri atas struktur yang disebut basidium (suatu sel berbentuk tabung atau seperti pemukul bola yang mempunyai empat buah basidiospora di bagian luarnya). Himenium yang dimiliki dapat menutupi permukaan, berpori, tubuh buah berkayu, keras dan ulet, serta mempunyai lapisan-lapisan membran, permukaan atas tubuh buah (konus) rata dan halus, dan spora pipih di bagian bawahnya. Berbagai macam spesies Ganoderma, yang paling banyak menyebabkan penyakit adalah
Ganoderma lucidum yang menyebabkan
penyakit busuk hati dan busuk pangkal batang (Streets 1982), serta Ganoderma
pseudoferreum yang dikenal sebagai jamur akar merah (Semangun 1988).
Ganoderma spp. merupakan jamur
penyebab penyakit busuk akar yang menyebabkan kerusakan pada tanaman perkebunan. Serangan Ganoderma spp. baru diketahui ketika tingkat infeksi sudah lanjut dan tanaman sudah sulit untuk diselamatkan (Mohd Su’ud et al. 2007; Minarsih et al.
2011). Penyakit busuk akar yang disebabkan oleh serangan Ganoderma spp. akan muncul pada pohon dewasa karena perkembangannya sangat lambat sehingga baru akan terlihat setelah bertahun-tahun kemudian (Semangun 1988).
Gambar 1 Tubuh buah Ganoderma spp.
DNA Barcoding
Teknik DNA barcoding merupakan teknik analisis identifikasi yang menggunakan standarisasi universal pada suatu fragmen genomik untuk mewadahi pencarian dan identifikasi spesies (Hajibabaei et al 2006; Hebert et al 2003; Kress et al 2005; Savolainen et al 2005). Hebert et al (2003) telah melakukan penelitian dengan teknik DNA Barcoding
yang diujikan pada berbagai macam spesies hewan. Penelitian tersebut menunjukkan kemampuan fragmen pendek di dalam mitokondria, yaitu CO1. Fragmen pendek ini sangat efektif sebagai sekuen barcode secara universal, terutama untuk hewan.
Kebutuhan untuk melakukan standarisasi universal dalam identifikasi spesies sangat tinggi dengan munculnya berbagai masalah dalam metode identifikasi dan determinasi spesies yang ada saat ini. Permasalahan tersebut dapat berakibat pada kesamaan nama pada dua spesies yang berbeda, yang dapat dimungkinkan karena kesamaan morfologi. Selain itu, dapat juga berakibat pada perbedaan nama pada satu spesies yang memiliki tingkat kehidupan yang sulit untuk diidentifikasi secara kasat mata. Oleh karena itu, menurut Hebert et al.
identifikasi seperti ini dapat mempercepat proses analisis sehingga dapat dijadikan suatu teknologi yang ekonomis untuk identifikasi spesies. Selain itu, keunggulan teknik DNA Barcoding adalah dapat digunakan untuk identifikasi suatu organisme walaupun DNA dari organisme tersebut tidak dalam bentuk murni, bahkan DNA yang sudah mengalami degradasi dan proses pengolahan pun dapat digunakan untuk analisis DNA Barcoding (Hajibabaei
et al. 2006).
Seifert et al. (2007) menunjukkan sebuah hasil penelitian lain, yaitu terdapat suatu prospek pada identifikasi fungi dengan teknik DNA barcoding. Pada fungi, sistem
barcode digunakan pada daerah 400-600 dari subunit ribosom, daerah cistron Internal Transcribed Spacer (ITS), daerah β-tubulin A (BenA), dan 1-α (EF-1α), serta gen CO1. Penelitian Seifert et al. (2007) ini menggunakan Pennicilum sebagai contoh untuk melakukan DNA barcoding pada fungi.
Isolasi DNA
Tahapan umum dalam isolasi DNA terdiri dari lisis sel atau penghancuran dinding sel, pemisahan debris sel atau pemisahan DNA dari bahan padat seperti selulosa dan protein, serta penghilangan protein (deproteinase) (Ardiana 2009). Dinding sel pada tanaman dapat dilisis secara fisik dan enzimatis, sedangkan membran sel dilisis dengan penambahan detergen. Lisis secara fisik dilakukan pada suhu 4°C agar DNA tidak rusak. Detergen yang digunakan dapat berupa SDS (sodium
dodecyl sulfate) atau CTAB
(cethyltrimethylammonium bromide)
(Wilson & Walker 2000). Metode isolasi DNA yang menggunakan penambahan CTAB merupakan metode yang umum digunakan dalam ekstraksi DNA genom tanaman, terutama yang banyak mengandung polisakarida dan senyawa polifenol (Ardiana 2009). Pada proses isolasi DNA juga digunakan EDTA (ethylenediaminetetraacetic acid) yang berfungsi sebagai pengkelat Mg2+, yaitu senyawa ionik yang dibutuhkan oleh enzim deoksiribonuklease (DNase) (Wilson & Walker 2000).
Setelah asam nukleat dipisahkan (DNA dan RNA) dari sel, maka proses selanjutnya adalah pemurnian. Proses ini merupakan proses pemisahan DNA dan RNA dari pengotor berupa protein dan pecahan sel.
Oleh karena itu, perlu dilakukan ekstraksi fenol-kloroform. Penambahan larutan ekstraksi ini dilanjutkan dengan proses sentrifugasi yang akan membagi larutan menjadi fase organik dan fase air yang dipisahkan oleh lapisan protein. DNA dan RNA akan terdapat pada fase air, sementara fase organik akan berisi lipid dan pecahan sel. RNA dapat dihilangkan dengan penambahan enzim ribonuklease (RNase) atau sentrifugasi gradien CsCl. Langkah terakhir, yaitu menambahkan etanol absolut untuk memekatkan DNA yang diperoleh dan selanjutnya disimpan pada suhu -20°C hingga siap digunakan (Adam et al. 1986).
Polymerase Chain Reaction (PCR) Metode Polymerase Chain Reaction (PCR) digunakan untuk amplifikasi DNA. Reaksi berantai polimerase (PCR) adalah suatu metode enzimatis untuk melipatgandakan secara eksponensial suatu sekuen nukleotida tertentu dengan cara in
vitro. Metode ini pertama kali
dikembangkan pada tahun 1985 oleh Kary B. Mullis, seorang peneliti di perusahaan
CETUS Corporation. Metode ini sekarang telah banyak digunakan untuk berbagai macam manipulasi dan analisis genetik (Yuwono 2006).
Amplifikasi PCR diawali dengan denaturasi DNA cetakan sehingga rantai DNA yang berantai ganda akan terpisah menjadi rantai tunggal. Denaturasi DNA menggunakan suhu 95°C selama 1-2 menit. Setelah itu suhu diturunkan menjadi 55°C sehingga primer akan menempel (annealing) pada cetakan yang telah terpisah menjadi rantai tunggal. Primer bertugas membentuk jembatan hidrogen dengan cetakan pada daerah sekuen yang komplementer dengan sekuen primer. Suhu inkubasi dinaikkan menjadi 72°C selama 1.5 menit. Pada suhu ini DNA polimerase akan melakukan polimerisasi rantai DNA yang baru. Setelah terjadi polimerisasi, rantai DNA baru akan membentuk jembatan hidrogen dengan DNA cetakan dan DNA untai ganda akan terbentuk. Reaksi-reaksi tersebut akan berlangsung terus-menerus hingga 25-30 siklus sehingga pada akhir siklus akan didapatkan molekul-molekul DNA rantai ganda yang baru hasil polimerisasi dalam jumlah yang banyak (Reece 2004).
Polymerase Chain Reaction (PCR)
digunakan juga memiliki ujung sekuen DNA target yang akan dicetak dan umumnya mempunyai kandungan guanin dan sitosin sebesar 50-60%. Primer universal mengandung pengertian bahwa primer tersebut mampu mengamplifikasi sekuen nukleotida pada DNA cetakan secara umum sehingga dapat digunakan oleh berbagai macam DNA cetakan. Sementara itu, pada primer spesifik, kemampuan primer tersebut sangat spesifik dalam mengamplifikasi sekuen nukleotida pada DNA cetakan sehingga hanya dapat digunakan pada DNA cetakan tertentu saja dan tidak dapat digunakan secara umum (Reece 2004).
Metode PCR sangat sensitif sehingga fragmen DNA yang diamplifikasi dengan menggunakan PCR akan diperoleh perbanyakan sebesar 200.000 kali setelah dilakukan 20 siklus reaksi selama 220 menit. Hal ini menunjukkan bahwa pelipatgandaan suatu fragmen DNA dapat dilakukan secara cepat. Kelebihan lain dari metode PCR adalah dapat dilakukan dengan menggunakan DNA cetakan dalam jumlah sangat sedikit. Selain itu, DNA cetakan yang digunakan juga tidak perlu dimurnikan terlebih dahulu sehingga metode PCR dapat digunakan untuk melipatgandakan suatu sekuen DNA dalam genom bakteri hanya dengan mencampurkan kultur bakteri di dalam tabung PCR (Yuwono 2006).
Teknik Rekombinasi DNA Teknik rekombinasi DNA secara in vitro
terdiri atas enam tahap. Pertama, isolasi molekul DNA (gen) yang akan digunakan. Kedua, pemotongan DNA dengan menggunakan enzim endonuklease restriksi. Tahap ketiga adalah penyambungan molekul-molekul DNA dengan menggunakan enzim DNA ligase ke dalam suatu molekul DNA vektor. Tahap keempat adalah transformasi sel inang dengan DNA rekombinasi hasil ligasi. Tahap terakhir, yaitu analisis dan konfirmasi keberadaan DNA rekombinan di dalam sel inang serta karakterisasi fungsional gen yang diklon (Brown 2010 diacu dalam Saraswati 2010). Salah satu bagian terpenting dalam teknik rekombinasi DNA atau kloning gen adalah vektor (Chawla 2002). Vektor adalah molekul DNA yang dapat bereplikasi secara mandiri dan dapat digunakan sebagai pembawa molekul DNA lain yang tidak mampu bereplikasi sendiri di dalam sel. DNA target perlu disisipkan ke dalam vektor agar DNA tersebut dapat direplikasi di
dalam sel inang serta dapat diwariskan pada saat membelah diri. Vektor pengklonan dirancang dan dikembangkan secara khusus untuk digunakan pada sel inang tertentu (Artika 2008).
Setelah proses ligasi DNA asing dengan DNA vektor dilakukan, tahap selanjutnya adalah memasukkan DNA rekombinan ke dalam sel inang yang sesuai (Whitehouse 2009). Proses pemasukan molekul DNA rekombinan ke dalam sel inang bakteri dikenal dengan istilah transformasi (jika vektor yang digunakan adalah plasmid) atau transfeksi (jika vektor yang digunakan adalah virus) (Brown 2000). Plasmid akan mengubah sel inang ke dalam sifat-sifat tertentu, seperti resisten terhadap antibiotika tertentu (Artika 2008). Transformasi dapat dilakukan dengan teknik induksi kompetensi sel secara kimiawi diikuti kejutan panas (heat shock) atau teknik elektroporasi menggunakan kejutan arus listrik bertegangan tinggi dalam waktu singkat (Brown 2000). Cara elektroporasi lebih efisien dibandingkan kejut panas tetapi memerlukan peralatan khusus untuk membangkitkan aliran listrik bertegangan tinggi yang disebut elektroporator (Howe 1995).
Sel inang yang digunakan pada proses kloning gen biasanya adalah sel E.coli. Sel
E.coli ini telah dibuat menjadi kompeten sehingga bukan strain alami karena sel
E.coli tergolong tidak efisien dalam
menyerap DNA. Setelah plasmid dimasukkan ke dalam sel inang, maka dilakukan seleksi sel transforman dengan menumbuhkan sel hasil transformasi pada media tumbuh yang spesifik. Transforman ditumbuhkan pada media sesuai marka seleksi. Marka seleksi adalah gen yang memberi karakteristik baru pada sel transforman yang tidak dimiliki oleh sel non-transforman. Sel transforman akan mampu tumbuh pada media sesuai marka seleksi, tetapi sel non-transforman akan mati (Liu 2007).
BAHAN DAN METODE
Bahan dan Alat
Bahan-bahan yang digunakan pada proses kultur isolat Ganoderma spp., yaitu isolat Ganoderma spp. (Lampiran 2), bubuk
DiftoTM Potato Dextrose Agar (PDA), tripton, ekstrak ragi, NaCl, dan agar bakto.
larutan setiltrimetilamonium bromida (CTAB), serbuk polivinilpolipirolidon (PVPP), nitrogen cair, es batu, akuades steril, larutan β-merkaptoetanol 1%, larutan kloroform : isoamilalkohol (24:1), larutan isopropanol dingin, buffer Tris-HCl EDTA (TE), natrium asetat, etanol absolut, dan etanol 70%.
Bahan-bahan yang digunakan untuk amplifikasi DNA, yaitu DNA cetakan, bufer PCR, molecular water (MW), 6 pasang primer (CO1_PenF1 / CO1_PenR1; CO1_B_F / CO1_B_R; BenA_F / BenA_R; ITS 1_F / ITS4_B; Gan_ITS1_F / Gan_ITS1_R; Gan_ITS2_F / Gan_ITS2_R), enzim Taq Polimerase, dan dNTPs 10 mM. Bahan-bahan yang diperlukan untuk elektroforesis gel agarosa adalah bubuk agarosa (Fermentas), bufer Tris-Borat EDTA (TBE) 0.5x, larutan etidium bromida (EtBr) 5 µg/100ml, loading buffer
(bromfenol biru 2.5%, sukrosa 40%), dan penanda molekuler (marker) DNA1 kb plus ladder (Invitrogen)
Bahan-bahan yang digunakan pada pengklonan gen adalah DNA target, vektor (plasmid) pGEM-T Easy (Promega), T4 Ligase, molecular water (MW), sel E.coli
kompeten, antibiotik ampisilin, isopropil-β -D-1-tiogalaktopiranosida (IPTG), 5-bromo-4-kloro-3-indol-β-D galaktopiranosida (X-Gal), dan media Luria Bertani (LB) agar.
Bahan-bahan yang digunakan pada elusi dan pemurnian adalah QIAquick Gel Extraction Kit (Qiagen). Sementara itu, untuk isolasi plasmid menggunakan
GeneJET Plasmid Miniprep Kit (Fermentas). Alat-alat yang digunakan adalah tabung Eppendorf, pipet mikro (0.1 µL, 10 µL, 100 µL, dan 1000 µL) dan tip,cawan Petri, labu Erlenmeyer, sudip, bunsen, tusuk gigi steril, gelas ukur, gelas piala, Laminar Air Flow
(LAF), autoklaf, mikrosentrifus Eppendorf 5417R dengan diameter rotor 10 cm, seperangkat alat elektroforesis, adaptor 100 volt, transiluminator ultraviolet (UV) T2201 (Sigma), program software geldoc, mesin PCR (ESCO Swift max), DNA speed vacum
110 savant, transluminator ultraviolet (UV) T2201 (Sigma), neraca analitik digital, oven, inkubator, lemari es dan penangas air.
Metode
Kultivasi Isolat Murni Ganoderma spp. dari Tubuh Buah
Media yang dibuat terdiri atas media padat dan media cair. Media padat yang
digunakan, yaitu Potato Dextrose Agar
(PDA). Sebanyak 10 gram bubuk PDA ditimbang, kemudian dilarutkan dengan akuades sebanyak 250 mL pada labu Erlenmeyer dan dipanaskan hingga mendidih. Kemudian larutan media PDA ditutup rapat dengan kapas dan alumunium foil. Selanjutnya disterilisasi dalam autoklaf selama kurang lebih 15 menit pada suhu 121°C dan tekanan 1 atm. Larutan media yang sudah disterilisasi di dalam autoklaf kemudian ditambahkan antibiotik streptomisin. Setelah itu dituang ke dalam cawan Petri yang sudah steril dan didiamkan hingga padat. Penuangan media ke dalam cawan Petri dilakukan di dalam laminar. Sementara itu, media cair yang digunakan adalah kaldu ekstrak malt (MEB). Sebanyak 4 gram ekstrak malt dan 4 gram pepton ditimbang, dilarutkan dengan akuades 200 mL, dan dipanaskan di dalam labu Erlenmeyer. Setelah itu dituang ke dalam tabung kecil, ditutup rapat dengan alumunium foil. Selanjutnya disterilisasi dalam autoklaf selama kurang lebih 15 menit pada suhu 121°C dan tekanan 1 atm. Kemudian ditambahkan antibiotik streptomisin sebelum digunakan.
Selanjutnya, isolat murni Ganoderma
spp. dari tubuh buah dikulturkan dengan keadaan steril di dalam Laminar Air Flow
(LAF). Tubuh buah Ganoderma spp. dipatahkan dengan tangan pada bagian tengah, kemudian diambil jaringan tubuh buah yang terletak di bagian tengah sebanyak satu cuplikan dengan menggunakan pinset steril. Selanjutnya diinokulasikan pada media PDA yang telah disiapkan sebelumnya. Biakan akan tumbuh setelah kurang lebih 1-2 minggu. Setelah 1-2 minggu, biakan murni yang tumbuh dipindahkan ke dalam agar miring untuk koleksi dan ke dalam media PDA baru. Pemindahan biakan ke dalam media PDA baru bertujuan untuk peremajaan. Setelah biakan tumbuh banyak, biakan pada media PDA dipotong-potong kecil dan dipindahkan ke dalam media cair (MEB). Selanjutnya diinkubasi selama kurang lebih 1 minggu. Miselium akan tumbuh lebat dan dipanen untuk keperluan isolasi DNA.
Isolasi DNA Genom Ganoderma spp. (Orozco-Castillo et al. 1994)
miselium beku, kemudian digerus dengan mortar dingin dan ditambahkan serbuk PVPP 0.1 gram. Selama penggerusan, ditambah nitrogen cair secara terus menerus untuk menjaga temperatur agar DNA tidak rusak. Penggerusan dihentikan ketika sampel telah menjadi serbuk halus dan selanjutnya dimasukkan ke dalam tabung Eppendorf. Setelah itu, sampel ditambahkan campuran 1 mL buffer ekstraksi dan 0.1 mL larutan β -merkaptoetanol 1% yang hangat serta dicampur merata dengan membolak-balikkan tabung Eppendorf. Kemudian campuran diletakkan di dalam penangas air selama 30 menit dengan suhu 65°C. Setelah 30 menit, campuran didinginkan pada suhu kamar selama 5 menit dan ditambahkan dengan larutan kloroform : isoamilalkohol (24:1) sebanyak 1 mL. Selanjutnya dicampur merata dengan membolak-balikkan tabung Eppendorf.
Tahap selanjutnya, yaitu sentrifugasi selama 10 menit dengan kecepatan 1355.2 g. Sentrifugasi akan menghasilkan dua lapisan cairan, namun hanya lapisan bagian atas saja (supernatan) yang diambil dan dipindahkan ke dalam tabung Eppendorf baru. Supernatan kemudian ditambahkan dengan larutan kloroform : isoamilalkohol (24:1) sebanyak 1 mL dan disentrifugasi selama 10 menit dengan kecepatan 1355.2 g. Sentrifugasi menghasilkan dua lapisan lagi, lapisan bagian atas diambil dan dipindahkan ke dalam tabung Eppendorf baru.
Setelah itu ditambahkan dengan larutan isopropanol dingin sebanyak 1 volume (1 mL) dan dikocok hingga homogen serta disimpan pada suhu 4°C selama 30 menit. Campuran disentrifugasi selama 10 menit dengan kecepatan 1355.2 g, kemudian pelet dikeringkan dan dilarutkan dengan 100 µL buffer TE, 10 µL CH3COONa 3 M pH 5.2,
dan 250 µL etanol absolut. Campuran dikocok hingga homogen. Kemudian disimpan di dalam lemari es (suhu -20°C) selama 30 menit atau semalam. Setelah beku, kemudian sentrifugasi kembali selama 10 menit dengan kecepatan 1612.8 g dan suhu 4°C. Pelet DNA diambil dan ditambahkan dengan etanol 70% sebanyak 100 µL, kemudian ditambahkan dengan 30
µL nuclease-free water (NFW) serta
disimpan dalam lemari pembeku.
Pengukuran Kuantitas dan Kualitas DNA (Sambrook & Russell 2001)
Pengukuran kuantitas DNA menggunakan metode spektrofotometri
dengan alat spektrofotometer UV. Sampel DNA yang sebelumnya telah diisolasi diencerkan hingga 100x dengan menambahkan 297 µL molecular water
(MW) ke dalam 3 µL sampel DNA. Selanjutnya, diukur absorbansinya pada panjang gelombang 230, 260 dan 280 nm. Tingkat kemurnian sampel dapat diperkirakan dengan menghitung nilai rasio serapan pada panjang gelombang 260 nm dan 280 nm, sedangkan konsentrasi sampel diperoleh dari hasil perkalian nilai serapan pada panjang gelombang 260 nm dengan faktor koreksi dan faktor pengenceran.
Sementara itu, pengukuran kualitas DNA menggunakan metode elektroforesis gel agarosa 1%. Sebanyak 0.6 gram agarosa dicampur dengan bufer TBE 0.5x sebanyak 60 mL. Setelah dicampurkan, kemudian dilarutkan sambil dipanaskan di dalam oven selama 90 detik. Setelah itu, cairan didiamkan hingga hangat dan diberi larutan EtBr sebanyak 3 µL. Cairan agarosa diaduk merata, dituang ke dalam cetakan sumur dan didiamkan hingga membeku dan padat. Setelah gel agarosa membeku dan padat, gel agarosa dipindahkan ke dalam bak elektroforesis yang telah berisi larutan bufer TBE 0.5x. Hasil isolasi DNA sebanyak 5 µL
dan loading buffer sebanyak 1 µL
diinjeksikan satu per satu ke dalam sumur gel agarosa 1%. Pada bagian ujung sumur gel agarosa dimasukkan juga penanda molekuler (marker) dengan ukuran1 kb plus
ladder sebanyak 0.8 µL. Kemudian dialirkan arus dan diberi tegangan sebesar 75 V. Migrasi dapat ditunggu selama kurang lebih 45-60 menit. Setelah proses migrasi selesai, maka diamati dan dianalisis pola pita DNA isolat Ganoderma spp. pada gel agarosa dengan menggunakan transluminator ultraviolet (UV) T2201 (Sigma). Hasil pengamatan pola pita DNA dilihat melalui perangkat lunak Geldoc.
Desain Primer, Optimasi Primer terhadap Isolat Ganoderma spp., dan Amplifikasi DNA dengan Metode PCR (Wilson &Walker 2000)
Tahap ini bertujuan untuk mendesain primer yang akan digunakan pada amplifikasi DNA isolat Ganoderma spp. melalui analisis secara in silico dengan program PRIMER3
(http://frodo.wi.mit.edu/primer3/) atau
merupakan hasil desain peneliti di Balai Penelitian dan Bioteknologi Perkebunan Indonesia (BPBPI) (Minarsih 2011) dan ada pula yang dirujuk dari beberapa jurnal
penelitian serta website (http://www.boldsystems.org). Pilihan primer yang digunakan ada enam pasang
(forward dan reverse) seperti yang
tercantum pada Tabel 1.
Optimasi primer berfungsi untuk menguji kecocokan pasangan primer (forward dan reverse) dengan sampel DNA
Ganoderma spp. melalui teknik Polymerase
Chain Reaction (PCR). DNA isolat
Ganoderma spp. dengan konsentrasi 100 ng diambil sebanyak 1 µL dan ditampung di dalam tabung Eppendorf yang berbeda-beda. Kemudian masing-masing tabung Eppendorf yang berisi DNA isolat
Ganoderma spp. ditambahkan campuran
reaksi dengan komposisi bufer 2.5 µL, dNTPs 0.5 µL, sepasang primer masing-masing 1 µL, enzim Taq polimerase 0.3 µL, dan ddH2O 18.70 µL. Total volume
campuran dalam satu tabung Eppendorf adalah 25 µL.
Setelah campuran merata, kemudian dimasukkan ke dalam mesin PCR. Mula- mula dilakukan optimasi suhu penempelan (annealing) primer dengan menggunakan metode PCR gradien. Suhu annealing yang digunakan pada masing-masing pasangan primer adalah CO1_PenF1/CO1_PenR1 (50.4; 52.4; 54.4; 56.4; 58.4)°C, CO1_B_F/CO1_B_R (49; 51; 53; 55; 57; 59) °C, BenA_F/BenA_R (53; 55; 57; 59; 61)°C, ITS1_F/ITS4_B (50; 52; 54; 56; 58; 60; 62)°C, Gan_ITS1_F/Gan_ITS1_R (48; 50; 52; 54; 56; 58)°C, Gan_ITS2_F/Gan_ITS2_R (48; 50; 52; 54; 56; 58)°C. Amplifikasi DNA dilakukan sebanyak 30 siklus dengan dengan program predenaturasi pada suhu 94°C selama 5 menit, denaturasi pada suhu 94°C selama 45
detik, penempelan primer (annealing) pada suhu yang telah disebutkan di atas selama 45 detik, pemanjangan primer (extension) pada suhu 72°C selama 2 menit, dan pasca pemanjangan pada suhu 72°C selama 5 menit. Selanjutnya produk PCR diverifikasi pada elektroforesis gel agarosa 1%.
Ekstraksi (Elusi) dan Purifikasi Fragmen DNA (Metode QIAquick Gel Extraction Kit), dan Pengklonan Gen (Sambrook & Russell 2001)
Tahap ini bertujuan untuk memperoleh DNA yang terpisah dari komponen-komponen lainnya (pengotor) seperti RNA, protein, karbohidrat, dan larutan, sehingga diperoleh suatu DNA murni untuk kepentingan analisis berikutnya. Gel yang telah dielektroforesis dan menunjukkan hasil elektroforegram positif, kemudian dipotong-potong menjadi potongan-potongan kecil pada bagian yang terdapat fragmen DNA. Gel tersebut disimpan dalam tabung Eppendorf berukuran 2 ml. Kemudian ditimbang dan diusahakan agar bobot gel tidak melebihi 400 mg. Elusi menggunakan metode QIAquick Gel Extraction Kit (Qiagen) dengan bantuan mikrosentrifus.
Selanjutnya, kolom QIAquick dibilas dengan etanol dan disentrifus kembali dengan kecepatan 1892,8 g selama 1 menit. Pembilasan dengan etanol berfungsi untuk mencuci DNA dari senyawa-senyawa lain seperti lemak, protein, dan lain-lain. Setelah itu, etanol dibuang dan tabung Eppendorf yang hanya berisi benang-benang DNA disentrifus kembali dengan kecepatan dan waktu yang sama untuk mengeringkan kolom QIAquick. Kemudian ditambahkan bufer elusi sebanyak 15 µl untuk mengelusi cairan yang berada di dalam kolom
QIAquick dan didiamkan selama 1 menit serta disentrifus lagi. Selanjutnya, cairan yang terkumpul dapat disimpan di dalam lemari es dan dapat dianalisis dengan elektroforesis gel agarosa untuk membuktikan DNA telah bersih dari pengotor.
Sementara itu, langkah-langkah untuk melakukan kloning gen terdiri atas ligasi
insert (DNA target) ke dalam vektor
(plasmid), transformasi DNA plasmid ke dalam sel kompeten, dan seleksi sel yang membawa plasmid berisi gen target. Tahap pertama, yaitu ligasi. Campuran untuk ligasi
insert ke dalam vektor (plasmid) berisi komposisi insert sebanyak 3.0 µL, vektor (plasmid) pGEMT-Easy (Promega) sebanyak 0.5 µL, T4 Ligase 1.0 µL, bufer sebanyak 5.0 µL, dan MW 0.5 µL. Pencampuran harus dilakukan dalam kondisi dingin (di dalam es). Setelah dicampur, kemudian diinkubasi selama semalam pada suhu 4°C.
Transformasi DNA plasmid ke dalam sel
E.coli dilakukan dengan memasukkan DNA plasmid sebanyak 10 µL hasil ligasi ke dalam 200 µL sel kompeten dan dicampur merata secara perlahan serta disimpan di dalam es selama 30 menit. Kemudian diberi perlakuan heat shock (kejut panas) di dalam penangas air dengan suhu 42°C selama 50 detik tanpa digoyang sedikit pun. Setelah itu, didinginkan kembali di dalam es selama 10 menit. Selanjutnya, ditambahkan campuran 800 µL medium LB dan glukosa 20 mM tanpa antibiotik. Kemudian diinkubasi pada suhu 37°C sambil digoyang dengan kecepatan 50 rpm selama 1 jam. Setelah itu, sebanyak 100 µL suspensi sel transforman dipindahkan pada media LA yang telah diberi antibiotik ampisilin 100 ppm, IPTG, dan X-Gal pada cawan Petri dan diratakan. Selanjutnya diinkubasi pada suhu 37°C selama semalam. Setelah semalam, maka akan muncul koloni putih dan biru pada media LA.
PCR Koloni
Koloni putih yang tumbuh kemudian dipindahkan ke dalam media LA dengan menggunakan tusuk gigi steril untuk membuat duplikat koloni. Selanjutnya diinkubasi selama semalam pada suhu 37°C. Selain itu, koloni putih yang tumbuh juga dipindahkan ke dalam tabung Eppendorf yang berisi MW sebanyak 10 µl. Pemindahan koloni dilakukan secara steril di dalam laminar. Adapun komponen reaksi yang perlu dipersiapkan adalah sebagai berikut: bufer PCR 1.5 µl, dNTPs 0.3 µl, sepasang primer spesifik masing-masing 0.15 µl, enzim Taq polimerase 0.15 µl, dan MW 2.75 µl.
Tahap pertama PCR adalah program lisis 96°C selama 5 menit, 50°C selama 1 menit 30 detik, 96°C selama 1 menit 30 detik, 45°C selama 1 menit 30 detik, 96°C selama 1 menit, 40°C selama 1 menit. Program PCR dihentikan sejenak untuk penambahan komponen reaksi sebanyak 5 µl ke dalam masing-masing tabung Eppendorf. Kemudian program PCR dilanjutkan dengan program PCR biasa, yaitu predenaturasi pada suhu 94°C selama 5 menit, denaturasi pada suhu 94°C selama 45 detik, penempelan primer (annealing) pada suhu yang telah disebutkan di atas selama 45 detik, pemanjangan primer (extension) pada suhu 72°C selama 2 menit, dan pasca pemanjangan pada suhu 72°C selama 5 menit. Setelah perbanyakan koloni transforman selesai, dilakukan pengujian dengan elektroforesis gel agarosa 1%.
Isolasi Plasmid Rekombinan (Kit Fermentas)
Langkah pertama sebelum diisolasi adalah kultur plasmid. Duplikat koloni putih yang telah terbentuk pada media LA selanjutnya dikultur ke dalam media LB cair yang telah diberi ampisilin sebanyak 100 ppm. Koloni putih tersebut diambil dengan ujung tusuk gigi steril dan dimasukkan ke dalam media LB cair. Setelah itu, ditutup rapat dan diinkubasi sambil digoyangselama 16 jam. Setelah 16 jam, maka koloni akan tumbuh dan plasmidnya dapat diisolasi.
larutan lisis sebanyak 250 µL dan tabung Eppendorf dikocok secara perlahan dengan membolak-balikkan tabung Eppendorf sebanyak 4-6 kali. Selanjutnya, ditambah lagi dengan 350 µL larutan netralisasi dan tabung Eppendorf dikocok secara perlahan seperti sebelumnya. Kemudian disentrifus selama 5 menit dengan kecepatan 1892,8 g. Supernatannya yang terbentuk dituang ke dalam kolom GeneJET spin dan disentrifus kembali selama 1 menit dengan kecepatan 1892,8 g. Setelah itu, ditambah larutan pencuci sebanyak 500 µl dan disentrifus selama 30-60 detik. Cairan yang tersaring oleh kolom kemudian dibuang. Hal ini dilakukan sebanyak dua kali. Selanjutnya, kolom kosong yang telah dibuang cairannya, disentrifus kembali untuk menghilangkan sisa-sisa cairan yang masih ada. Langkah terakhir, yaitu kolom dipindahkan ke dalam tabung Eppendorf baru dan ditambah dengan 50 µl buffer elusi 10 mM Tris-HCl pH 8,5. Selanjutnya diinkubasi selama 2 menit pada suhu ruang dan disentrifus selama 2 menit pada kecepatan 1892,8 g. Supernatannya yang terbentuk merupakan hasil isolasi plasmid rekombinan dan disekuen untuk dianalisis keragaman genetiknya.
Analisis Sekuens (sequencing)
Fragmen DNA plasmid Ganoderma spp. yang telah diisolasi, kemudian diurutkan basa nukleotida (sequencing) di Balai Lembaga Ilmu Pengetahuan Indonesia (LIPI) Cibinong dan 1st BASE menggunakan primer universal (T7). Hasil
sequencing kemudian diidentifikasi
spesiesnya dengan menggunakan program
BLAST pada NCBI (http://www.ncbi.nlm.nih.gov) dan BOLD
System (http://www.boldsystems.org). Setelah isolat Ganoderma spp. berhasil diidentifikasi, tahap selanjutnya adalah analisis primer yang paling efektif untuk dijadikan primer spesifik dalam mengidentifikasi Ganoderma spp. Hal ini dapat dilihat dari tingkat akurasi setiap pasang primer dalam mengidentifikasi spesies Ganoderma.
HASIL DAN PEMBAHASAN
Kultivasi Isolat Murni Ganoderma spp. dari Tubuh Buah
Miselium isolat Ganoderma spp. tumbuh menyebar pada media PDA dalam cawan Petri. Miselium berwarna putih dan
berbentuk seperti benang-benang halus seperti yang ditunjukkan pada Gambar 2. Miselium ini diperoleh dari penanaman satu cuplikan jaringan tubuh buah Ganoderma
spp. pada media PDA selama kurang lebih 1-2 minggu.
Selanjutnya, setelah 1-2 minggu, miselium berwarna putih yang tumbuh banyak pada media PDA di dalam cawan Petri dikulturkan kembali ke dalam media MEB cair. Tujuannya memperbanyak sampel untuk keperluan isolasi DNA selanjutnya. Setelah satu minggu, maka tumbuh miselium berwarna putih yang menyebar pada media MEB cair di dalam cawan Petri. Miselium ini kemudian dipanen untuk keperluan isolasi DNA.
Gambar 2 Miselium isolat Ganoderma spp. pada media PDA
Kualitas dan Kuantitas DNA Kromosom
Ganoderma spp.
Keberhasilan tahap isolasi ditunjukkan dengan kualitas DNA, yaitu dengan visualisasi elektroforesis gel agarosa 1% pada paparan sinar lampu UV. Gambar 3 menunjukkan pola pita DNA yang terbentuk tebal dan tidak terfragmentasi.
Sementara itu, kuantitas DNA diukur dengan menggunakan spektrofotometer UV-VIS pada panjang gelombang serapan 260 dan 280. Selanjutnya nilai absorbansi 1,000 dari hasil pembacaan dengan spektrofotometer setara dengan 50 µg/ml (Brown 2000; Wilson & Walker 2000). Nilai ini merupakan faktor konversi sehingga diperoleh rumus untuk menghitung konsentrasi DNA, yaitu A260 x faktor
konversi x faktor pengenceran.
Selain itu, tingkat kemurnian DNA juga dapat dilihat berdasarkan rasio serapan pada panjang gelombang 260 nm dan 280 nm (A260 : A280). Berdasarkan Wilson & Walker
(2000), suatu DNA memiliki kemurnian yang tinggi apabila rasio serapan A260 : A280
Pada Tabel 2, sampel DNA isolat
Ganoderma spp. menunjukkan tingkat
kemurnian yang beragam. Tingkat kemurnian DNA yang tinggi ditunjukkan oleh sampel PL5, PL6 dan T21. Ketiga sampel isolat DNA Ganoderma spp. ini memiliki rasio serapan A260 : A280 > 1.8.
Sementara itu, pada sampel isolat DNA
Ganoderma spp. yang lain (PL7, T9,T20, Laras, Raja, Bek, Tin, Lipi) memiliki kemurnian DNA yang rendah. Hal ini ditunjukkan dengan rasio serapan A260 :
A280 < 1.8. Walaupun DNA hasil isolasi
memiliki tingkat kemurnian yang rendah, namun DNA tersebut masih dapat dijadikan sampel untuk proses selanjutnya, yaitu amplifikasi dengan menggunakan metode PCR. Hal inilah yang menjadi keunggulan dari metode PCR, yaitu komponen sampel DNA dalam jumlah sedikit saja sebagai DNA cetakan, tetap dapat digunakan untuk proses amplifikasi (Bintang 2010). Oleh karena itu, sebelas sampel isolat DNA
Ganoderma spp. yang berhasil diisolasi dapat digunakan untuk amplifikasi menggunakan metode PCR.
Gambar 3 Elektroforegram kualitas DNA hasil isolasi. (1) sampel DNA PL5, (2) PL6, (3) PL7, (4) T9, (5) T20, (6) T21, (7) Laras, (8) Raja, (9) Bek, (10) Tin, (11) & (12) Lipi, (M) marker 1 kb plus
DNA ladder
Tabel 2 Hasil uji kuantitatif DNA pada panjang gelombang 260 dan 280 nm serta konsentrasi DNA (µg/ml)
Desain Primer, Optimasi Primer terhadap Isolat Ganoderma spp., dan Amplifikasi DNA dengan Metode PCR Suhu annealing (penempelan) pada enam pasang primer masing-masing dioptimasi terlebih dahulu sebelum digunakan untuk amplifikasi DNA. Tujuannya untuk mencari suhu annealing yang optimum untuk proses amplifikasi DNA pada sampel Ganoderma
spp. Berdasarkan hasil optimasi dengan berbagai suhu annealing, maka pasangan primer CO1_PenF1/CO1_PenR1, pasangan primer CO1_B_F/CO1_B_R, dan pasangan primer Gan_ITS2_F/Gan_ITS2_R tidak dapat mengamplifikasi DNA karena tidak terbentuk pola pita DNA pada gel agarosa 1% (data tidak ditampilkan). Hal ini berarti ketiga pasang primer tersebut tidak komplemen dengan sampel DNA
Ganoderma spp. sehingga tidak digunakan pada proses amplifikasi DNA selanjutnya. Sementara itu, pasangan primer BenA_F/BenA_R memiliki ukuran 348 pb dengan suhu annealing yang optimum sebesar 58°C (Gambar 4). Selanjutnya pasangan primer ITS1_F/ITS4_B memiliki ukuran 800 pb dengan suhu annealing yang optimum sebesar 58°C (Gambar 5). Pasangan primer Gan_ITS1_F /Gan_ITS1_R memiliki ukuran 200 pb dengan suhu
annealing yang optimum sebesar 55°C
(Gambar 6). Oleh karena itu, ketiga pasang primer tersebut dapat digunakan untuk amplifikasi sampel DNA Ganoderma spp. dengan suhu annealing yang berbeda-beda.
Gambar 4 Elektroforegram hasil optimasi primer BenA_F/BenA_R. (1) kontrol negatif, (2) 53°C, (3) 55°C, (4) 57°C, (5) 59°C, (6) 61°C, (M) marker 1 kb plus
Gambar 5 Elektroforegram hasil optimasi primer ITS1_F/ITS4_B. (1) kontrol negatif, (2) 50°C, (3) 52°C, (4) 54°C, (5) 56°C, (6) 58°C, (7) 60°C, (8) 62°C, (M) marker 1 kb plus DNA ladder
Gambar 6 Elektroforegram hasil optimasi primer Gan_ITS1_F / Gan_ITS1_R. (1) kontrol negatif, (2) 48°C, (3) 50°C, (4) 52°C, (5) 54°C, (6) 56°C, (7) 58°C, (M) marker 1 kb plus
DNA ladder
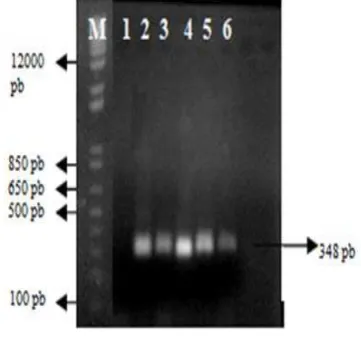
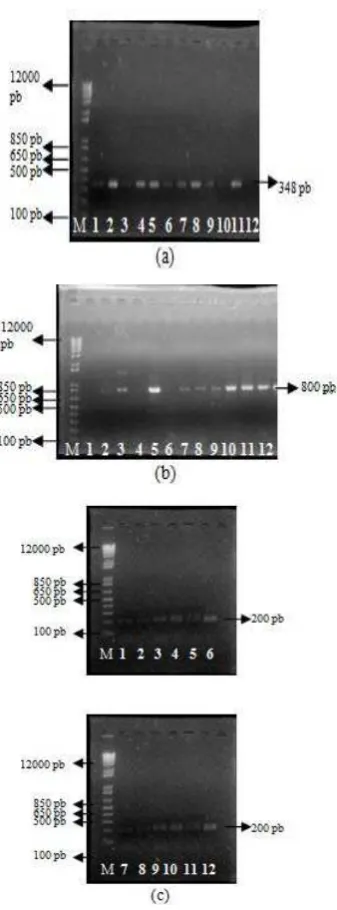
Pada Gambar 7 berkode (a), (b), dan (c) menunjukkan hasil amplifikasi primer BenA_F/BenA_R, ITS1_F/ITS4_B, dan Gan_ITS1_F / Gan_ITS1_R terhadap 11 sampel DNA dari isolat Ganoderma spp. Berdasarkan elektroforegram hasil amplifikasi terhadap 11 sampel DNA isolat
Ganoderma spp., maka terlihat beberapa pola pita DNA yang terang dan tebal dengan ukuran masing-masing secara berurutan adalah 348 pb, 800 pb, dan 200 pb. Pola pita DNA yang terang dan tebal ini kemudian dilakukan proses elusi (ekstraksi) untuk membersihkan dari segala pengotor DNA, seperti RNA, protein, dan lain-lain.
Gambar 7 Elektroforegram hasil amplifikasi dengan primer (a) BenA_F/BenA_R, (b) ITS1_F/ITS4_B, (c) Gan_ITS1_F / Gan_ITS1_R dan
masing-masing sampel DNA (1) PL5, (2) PL6, (3) PL7, (4) T9, (5) T20, (6) T21, (7) Laras, (8) Raja, (9) Bek, (10) Tin, (11) & (12) Lipi, (M) marker 1 kb plus
DNA ladder
Ekstraksi, Purifikasi Fragmen DNA, dan Pengklonan Gen
RNA. Hasil ekstraksi dan purifikasi ini kemudian disisipkan ke dalam vektor pGEM T Easy dan ditransformasikan ke dalam sel
E.coli (kloning gen). Hasil elusi (ekstraksi) dan purifikasi pada tahap sebelumnya, disisipkan ke dalam vektor pGEM T Easy (ligasi). Setelah itu, dilakukan proses transformasi ke dalam sel kompeten E.coli
dengan diberikan perlakuan kejut panas (heat shock) pada suhu 42°C.
Metode seleksi sel transforman yang digunakan untuk menyeleksi sel E.coli berisi plasmid rekombinan adalah metode koloni putih-biru. Bakteri E.coli yang diduga membawa gen target ditumbuhkan dalam media Luria Bertani agar (LA). Media ini mengandung antibiotik ampisilin 100 ppm, IPTG, dan X-Gal. Koloni yang mengandung sisipan (insert) akan merusak dinding gen lacZ sehingga tidak akan sanggup lagi memetabolisme X-gal yang menjadi substrat gen lacZ dan akan nampak sebagai koloni putih pada pelat agar. Sedangkan koloni yang tidak tersisipi gen target, maka gen lacZ yang fungsional akan memetabolisme X-gal sehingga akan nampak sebagai koloni berwarna biru. Penambahan ampisilin dilakukan karena vektor yang digunakan (pGEM T-Easy) memiliki marka seleksi resisten terhadap ampisilin (Lampiran 3). Koloni putih ini selanjutnya diduplikasi untuk diperbanyak jumlah sisipannya.
PCR Koloni
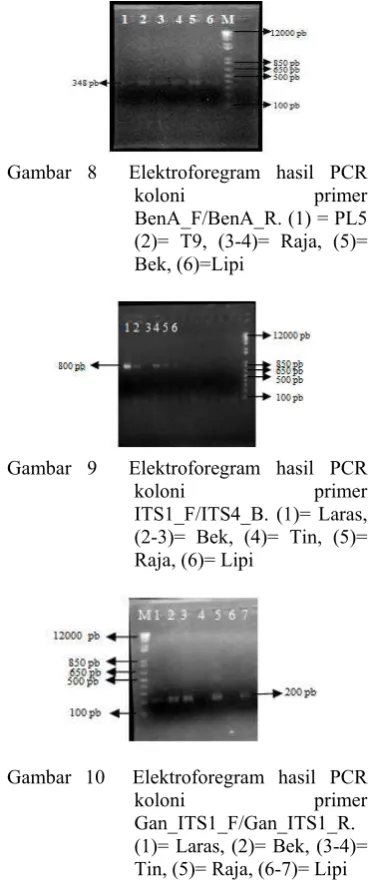
Koloni bakteri yang tumbuh dikonfirmasi dengan menggunakan teknik PCR koloni. Hal ini bertujuan agar dapat dipastikan bahwa koloni bakteri tersebut mengandung plasmid rekombinan. PCR koloni menggunakan tiga pasang primer spesifik, yaitu BenA_F / BenA_R, ITS1_F / ITS4_B, dan Gan_ITS1_F / Gan_ITS1_R. Gambar 8 menunjukkan hasil PCR koloni untuk primer BenA_F/BenA_R dengan ukuran 348 pb. Gambar 9 menunjukkan hasil PCR koloni untuk primer ITS1_F / ITS4_B dengan ukuran 800 pb dan Gambar 10 menunjukkan hasil PCR koloni untuk primer Gan_ITS1_F / Gan_ITS1_R dengan ukuran 200 pb.
Setelah proses konfirmasi PCR koloni, koloni rekombinan dikulturkan di dalam media Luria Bertani (LB) cair. Media LB cair ini berfungsi sebagai pemberi nutrisi agar sel dapat mengalami perbanyakan. Komposisi media LB terdiri atas ekstrak khamir yang berfungsi sebagai sumber gula, nitrogen, dan nutrisi organik dan anorganik,
tripton berfungsi menyediakan asam amino, serta NaCl sebagai penyedia garam. Selanjutnya koloni yang dikultur ke dalam media LB cair, diinkubasi semalam hingga tumbuh dengan mengamati warna media LB cair yang keruh. Langkah berikutnya, yaitu isolasi DNA plasmid di dalam koloni rekombinan sel E.coli.
Gambar 8 Elektroforegram hasil PCR
koloni primer BenA_F/BenA_R.(1) = PL5
(2)= T9, (3-4)= Raja, (5)= Bek, (6)=Lipi
Gambar 9 Elektroforegram hasil PCR
koloni primer ITS1_F/ITS4_B. (1)= Laras,
(2-3)= Bek, (4)= Tin, (5)= Raja, (6)= Lipi
Gambar 10 Elektroforegram hasil PCR
koloni primer Gan_ITS1_F/Gan_ITS1_R.
(1)= Laras, (2)= Bek, (3-4)= Tin, (5)= Raja, (6-7)= Lipi
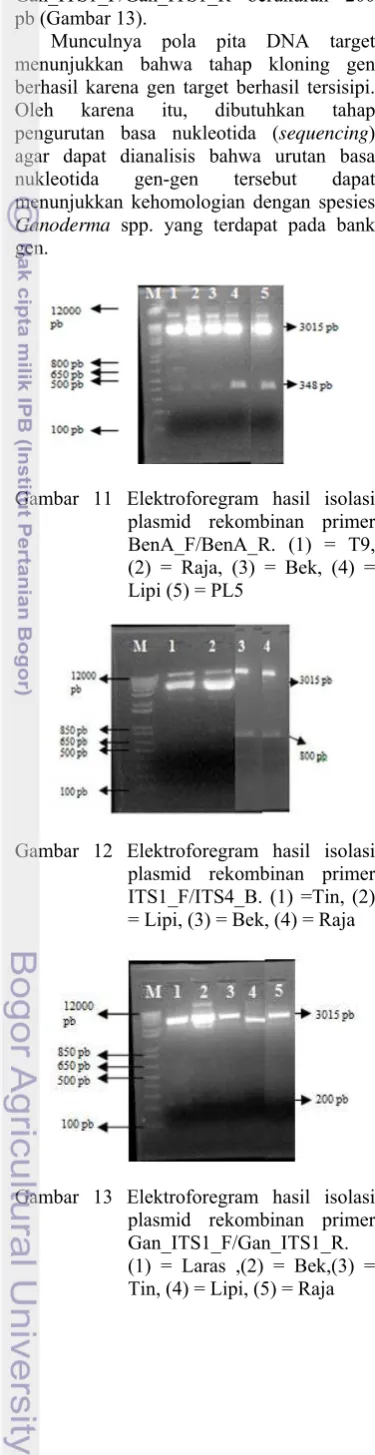
Plasmid Rekombinan
berukuran 800 pb (Gambar 12), dan primer Gan_ITS1_F/Gan_ITS1_R berukuran 200 pb (Gambar 13).
Munculnya pola pita DNA target menunjukkan bahwa tahap kloning gen berhasil karena gen target berhasil tersisipi. Oleh karena itu, dibutuhkan tahap pengurutan basa nukleotida (sequencing) agar dapat dianalisis bahwa urutan basa nukleotida gen-gen tersebut dapat menunjukkan kehomologian dengan spesies
Ganoderma spp. yang terdapat pada bank gen.
Gambar 11 Elektroforegram hasil isolasi plasmid rekombinan primer BenA_F/BenA_R. (1) = T9, (2) = Raja, (3) = Bek, (4) = Lipi (5) = PL5
Gambar 12 Elektroforegram hasil isolasi plasmid rekombinan primer ITS1_F/ITS4_B. (1) =Tin, (2) = Lipi, (3) = Bek, (4) = Raja
Gambar 13 Elektroforegram hasil isolasi plasmid rekombinan primer Gan_ITS1_F/Gan_ITS1_R. (1) = Laras ,(2) = Bek,(3) = Tin, (4) = Lipi, (5) = Raja
Analisis Sekuens
Tahap analisis sekuen merupakan tahap yang paling krusial dan membutuhkan waktu serta biaya yang tidak sedikit. Oleh karena itu, hanya beberapa isolat Ganoderma spp. saja yang dikirim ke Balai LIPI dan 1st BASE Pte Ltd (www.base-asia.com) untuk dilakukan sequencing. Isolat Ganoderma
spp. yang tercantum pada Lampiran 2 dipilih karena kualitasnya yang paling baik diantara isolat Ganoderma spp lainnya. Hal ini diketahui melalui tahap amplifikasi DNA dan kloning gen. Berdasarkan tahap pengurutan basa nukleotida (sequencing), maka diperoleh urutan basa nukleotida seperti yang tercantum pada Lampiran 4a-4d.
Urutan basa nukelotida (sekuen) selanjutnya dianalisis dengan program BLAST pada NCBI untuk menunjukkan kehomologian dengan spesies Ganoderma
spp. yang terdapat pada bank gen. Basic
Local Alignment Search Tool (BLAST)
merupakan program untuk mencari dan menganalisis kehomologian sekuen suatu organisme. Program ini dikelola oleh
National Center for Biotechnology
Information (NCBI)
(http://www.ncbi.nlm.nih.gov/BLAST/) (Ye Jian et.al 2006; Altschul et.al 1990). Selain dengan program BLAST, analisis identifikasi juga dilakukan dengan menggunakan program The Barcode of Life
Data System (BOLD System)
(http://www.boldsystems.org). BOLD System merupakan suatu wadah baru yang menampung informasi-informasi mengenai bioinformatika dari berbagai spesies yang disajikan dalam bentuk pustaka barcode. Bukan hanya itu, BOLD System juga menyediakan sistem analisis sekuen barcode
berdasarkan daerah pendek untuk identifikasi suatu spesies. Daerah pendek tersebut diklasifikasikan berdasarkan pengelompokkan hewan (CO1), tumbuhan (rbcL dan matK), dan fungi (ITS) (Ratnasingham & Hebert 2007).
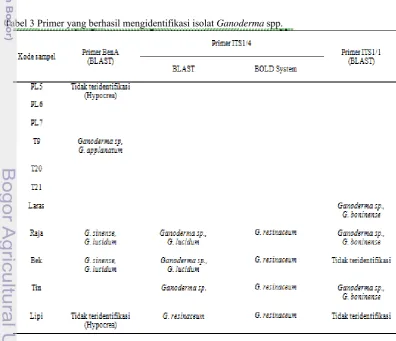
Ganoderma spp. berhasil diidentifikasi (Tabel 3). Hal ini terlihat dari nilai identitas yang menunjukkan tingkat homologi berkisar pada nilai > 90% (Lampiran 5a-5j). Pada Tabel 3 beberapa isolat Ganoderma
spp. juga menunjukkan hasil yang homolog dengan dua spesies berbeda yang teridentifikasi. Hal ini karena kedua spesies tersebut memiliki persentase identitas yang sama besar. Selanjutnya, pada identifikasi isolat Ganoderma spp. dengan primer ITS1_F/ITS4_B digunakan dua program analisis identifikasi, yaitu BLAST dan BOLD System. Hasil analisis kedua program tersebut mengidentifikasi Ganoderma spp. sebagai dua spesies yang berbeda (Tabel 3). Kemudian, untuk isolat yang berasal dari LIPI, semula isolat Ganoderma ini diidentifikasi oleh LIPI sebagai Ganoderma lucidum. Namun, setelah dianalisisis dengan menggunakan program BLAST dan BOLD System, sampel DNA yang berasal dari LIPI tersebut teridentifikasi sebagai Ganoderma resinaceum (Tabel 3). Sementara itu, isolat
Ganoderma spp. yang tidak berhasil
teridentifikasi dimungkinkan karena kesalahan pada saat proses sequencing. Hasil kromatogram yang terbaca dengan alat
sequencer tidak bagus karena memiliki noise
yang banyak sehingga pada saat analisis identifikasi dengan menggunakan program BLAST ataupun BOLD System, isolat tersebut tidak dapat diidentifikasi sebagai spesies Ganoderma.
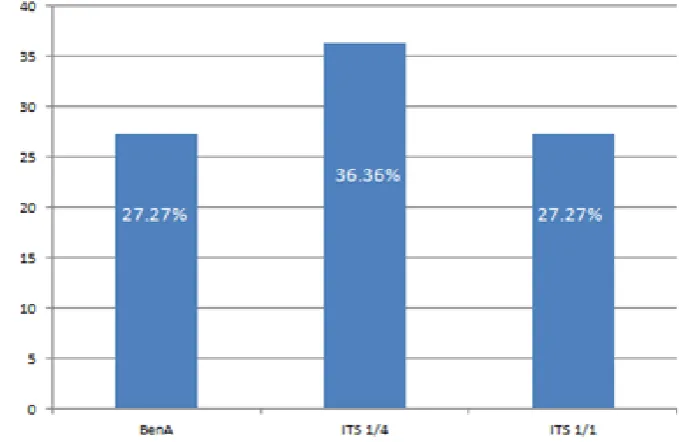
Berdasarkan ketiga pasang primer yang berhasil mengidentifikasi isolat Ganoderma
spp. dapat disimpulkan bahwa primer ITS1_F/ITS4_B yang paling banyak mengidentifikasi isolat Ganoderma spp. (Gambar 14). Diagram tersebut menunjukkan bahwa primer ITS1_F/ITS4_B memiliki persentase kemampuan dalam mengidentifikasi isolat Ganoderma spp. sebesar 36.36%, primer BenA_F/BenA_R dan primer Gan_ITS1_F/ Gan_ITS1_R sebesar 27.27%. Kemudian dari ketiga pasang primer yang berhasil mengidentifikasi isolat Ganoderma spp., primer ITS1_F/ITS4_B dan primer Gan_ITS1_F/ Gan_ITS1_R mampu mengidentifikasi isolat Bek sebagai
Ganoderma sp. Sementara itu, isolat
Ganoderma lain menunjukkan tingkat akurasi yang rendah karena dengan tiga pasang primer yang berbeda tidak dapat diidentifikasi sebagai spesies yang sama.
Gambar 14 Keberhasilan primer dalam mengidentifikasi isolat Ganoderma spp.
SIMPULAN DAN SARAN
Simpulan
Pasangan primer ITS1_F/ITS4_B memiliki persentase kemampuan yang tertinggi dalam mengidentifikasi isolat
Ganoderma spp., dibandingkan dengan
pasangan primer BenA_F/BenA_R,
Gan_ITS1_F/ Gan_ITS1_R, CO1_PenF1/CO1_PenR1,
CO1_B_F/CO1_B_R, dan Gan_ITS2_F/ Gan_ITS2_R. Oleh karena itu, pasangan primer ITS1_F/ITS4_B mempunyai potensi sebagai DNA Barcode untuk identifikasi spesies Ganoderma.
Saran
Saran yang dapat diberikan yaitu perlu dilakukan pengulangan tahap sequencing
pada beberapa sampel DNA Ganoderma
spp. yang memiliki hasil sekuen kurang baik agar dapat menunjukkan kualitas sequencing
yang baik. Selain itu, perlu mendesain kembali primer dengan daerah identifikasi yang lebih bervariasi sehingga dapat mengidentifikasi Ganoderma hingga tahap subspesies.
DAFTAR PUSTAKA
Adam RLP, Knowler JT, Leader DP. 1986.
Thiyae Biochemistry of The Nucleic
Acid, Tenth Edition. New York:
Chapman and Hall.
Alexopoulos CJ, Mims CW, Blackwell M. 1996. Introductory Micology. New York: John Willey & Sons, Inc.
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. 1990. Basic local alignment search tool. J Mol Biol. 215: 403-410.
Ardiana DW. 2009. Teknik isolasi DNA genom tanaman pepaya dan jeruk dengan menggunakan modifikasi buffer CTAB.
J Teknik Pertanian 14:12-16.
Artika IM. 2008. Teknologi Asam Nukleat dan Protein. Bogor: Institut Pertanian Bogor.
Bintang M. 2010. Teknik Penelitian Biokimia. Jakarta: Erlangga.
Brown TA. 2000. Essential Molecular Biology: a Practical Approach. New York: Oxford University Press, Inc.
Brown TA. 2010. Gene Cloning and DNA Analysis: An Introduction. 6th edition. West Sussex: John Wiley & Sons.
Effect, and Enviromental Impact. 2nd Edition. New York: CRC Press
Chawla HS. 2002. Introduction to Plant
Biotechnology. 2th edition. New
Hampshire: Science Publishers.
[Ditjen Perkebunan] Direktorat Jenderal Perkebunan. 2011. Ancaman penyakit busuk pangkal batang pada tanaman
kelapa sawit. http://ditjenbun.deptan.go.id/perlindunga
n/ [14 Desember 2011]
Hajibabaei M et al. 2006. A minimalist barcode can identify a specimen whose DNA is degraded. J Compilation Blackwell Publishing. 6: 959-964
Hebert PDN, Cywinska A, Ball SL, DeWaard JR. 2003. Biological identifications through DNA barcodes.
Proceedings of the Royal Society of London Series B, Biological Sciences. 270: 313- 321.
Howe C. 1995. Gene Cloning and Manipulation. Cambridge: Cambridge University Press.
Kress WJ, Wurdack KJ, Zimmer EA, Weight LA, Janzen DH. 2005. Use of DNA barcodes to identify flowering plants. Proceedings of the National Academy of Sciences, USA. 102: 8369-8374.
Liu SQ. 2007. Bioregenerative Engineering: Principles and Applications. New Jersey: John Wiley & Sons.
Minarsih H, NP Dyah, TW Darmono, Herliyana EN. 2011. Analisis keragaman genetic Ganoderma spp. yang berasosiasi dengan tanaman kakao dan tanaman pelindungnya menggunakan Random
Amplified Polymorphic DNA (RAPD).
Menara Perkebunan. 79(1): 6-14.
Mohd Su’ud M, PIA Loonis & AS Idris. 2007. Towards automatic recognition and grading of Ganoderma infection pattern using fuzzy systems. Int J Biomed Sci. 2: 1306-1216.
Orozco-Castillo C et al. 1994. Detection of genetic diversity and selective gene introgression in coffee using RAPD
markers. Theor. Appl. Genet. 87: 934-940.
Ratnasingham S, Hebert PDN. 2007. BOLD: The barcode of life data system. J Compilation Blackwell Publishing.
Reece RJ. 2004. Analysis of Genes and
Genomes. Manchester: Manchester
University Press.
Sambrook J, Russell DW. 2001. Molecular
Cloning a Laboratory Manual. New
York: Cold Spring Harbor Laboratory Press.
Savolainen V, Cowan RS, Vogler AP, Roderick GK, Lane R. 2005. Towards writing the encyclopaedia of life: an introduction to DNA barcoding.
Philosophical Transactions of The Royal Society of London. Series B, Biological Sciences. 360: 1805-1811.
Seifert KA, Samson RA, deWaard JR, Houbraken J, levesque CA, Moncalvo JM, Seize GL, Hebert PDN. 2007. Prospect for fungus identification using CO1 DNA barcodes, with Penicillium as a test case. Proceedings of the National Academy of Sciences, USA. 104: 3901-3906.
Semangun H. 1988. Penyakit-penyakit
Tanaman Perkebunan di Indonesia.
Yogyakarta: Gajah Mada University Press.
Streets RB. 1982. Diagnosis Penyakit Tanaman. Imam Santoso, penerjemah. Terjemahan dari: Diagnosis of Plant Diseases. Jakarta: PT Gede Jaya.
Suryanto D. 2003. Melihat Keanekaragaman Organisme melalui Beberapa Teknik Genetika Molekuler. [Skripsi]. Sumatera Utara: Program Sarjana, Universitas Sumatera Utara.
Sutiarna A. 2010. Suplementasi Ganoderma lucidum, kromium organik dan kedelai sangrai pada pakan sapi laktasi yang dievaluasi dari aspek fisiologis dan imunitas ternak [Skripsi]. Bogor: Program Sarjana, Institut Pertanian Bogor.
bagi manusia. Di dalam: Teknis Bioteknologi Perkebunan Untuk Praktek.
Prosiding Pertemuan Ilmiah Tahunan; Bogor, 5-6 Mei 1999. Bogor: Perhimpunan Mikrobiologi Indonesia Cabang Bogor. Hlm 31-37.
Taniwiryono D. 2011. Ganoderma sebagai patogen penyakit tanaman dan bahan baku obat tradisional. Simposium Nasional dan Lokakarya Ganoderma; IPB International Convention Center, 2-3 November.
Tudge C. 2000. The Variety Of Life. New York: Oxford University Press.
Widyastuti SM. 2007. Peran Trichoderma spp. dalam Revitalisasi Kehutanan di Indonesia. Yogyakarta: Gadjah Mada University Press.
Wilson K, Walker J. 2000. Principles and Techniques of Practical Biochemistry, Fifth Edition. Cambridge: Cambridge University Press.
Whitehouse DB. 2009. Genes and Genomes. Di dalam: Walker JM, Rapley R, (eds). 2009. Molecular Biology and Biotechnology. 5th edition. Cambridge: The Royal Society of Chemistry.
Ye J, McGinnis S, Madden TL. 2006. BLAST: Improvements for better sequence analysis. Nucleic Acid Research 34.
Yuwono T. 2006. Teori dan Aplikasi Polymerase Chain Reaction.
Lampiran 1 Skema isolasi DNA (Orozco-Castillo
et al.
1994)
Miselium digerus & ditambah 1 mL buffer ekstraksi & 0.1 mL larutan
β
-merkaptoetanol 1% hangat, dicampur hingga merata
Campuran diletakkan di dalam penangas air selama 30 menit, suhu 65°C,
didinginkan 5 menit, ditambahkan larutan kloroform : isoamilalkohol
(24:1) sebanyak 1 mL, sentrifugasi 1355,2 g selama 10 menit
Lapisan atas diambil, ditambah larutan kloroform : isoamilalkohol
(24:1) 1 mL, sentrifugasi 1355,2 g selama 10 menit
Lapisan atas diambil, ditambah larutan isopropanol dingin 1 mL,
sentrifugasi 10 menit 1355,2 g selama 10 menit, pelet dikeringkan
Pelet dilarutkan dengan 100 µl buffer TE, 10 µl CH
3COONa 3 M pH
5.2 & 250 µl etanol absolut
Dikocok hingga homogen, disimpan di dalam lemari beku (suhu
-20°C) selama 30 menit, sentrifugasi 1612,8 g, 10 menit suhu 4°C
Pelet DNA diambil dan ditambahkan dengan etanol 70% sebanyak
100 µL
Lampiran 2 Isolat
Ganoderma
spp. dan primer yang digunakan untuk
sequencing
Isolat
Ganoderma
spp.
No Kode Tumbuhan inang Asal wilayah
1. PL5 Sengon Palembang
2. PL6 Sengon Palembang
3. PL7 Sengon Palembang
4. T9 Sengon Tasikmalaya
5. T20 Sengon Tasikmalaya
6. T21 Sengon Tasikmalaya
7. Laras Kelapa sawit Medan
8. Raja Kelapa sawit Medan
9. Bek Kelapa sawit Lampung
10. Tin Kelapa sawit Medan
11. Lipi - LIPI
Primer yang digunakan untuk
sequencing
Kode
sampel Primer BenA Primer ITS1/4 Primer ITS1/1
PL5 9
PL6 PL7
T9 9
T20 T21
Laras 9
Raja 9 9 9
Bek 9 9 9
Tin 9 9
Lampiran 4a Hasil pengurutan basa nukelotida (
sequencing
) isolat
Ganoderma
spp. (warna merah = primer yang digunakan untuk amplifikasi
DNA, warna biru = penambahan sekuen nukleotida)
PL5 (Ben A_F/ BenA_R) : 342 pb
CATGGGTACCCTCCTGATCTCCAAGATCCGCGAGGAGTTCCCCGACCGAATGATGGCC ACCTTCTCCGTCGTGCCATCCCCCAAGGTGTCCGACACCGTCGTTGAGCCCTACAACG CCACCCTCTCCGTCCACCAGCTGGTCGAGAACTCCGACGAGACCTTCTGCATTGACAA CGAGGCTCTCTACGACATCTGCATGCGCACCCTCAAGCTGTCCAACCCTGCCTACGGT GACCTGAACTACCTCGTCTCCGCTGTCATGTCCGGCATCACCACCTGTCTGCGATTCCC CGGTCAGCTTAACTCTGATCTGCGCAAGCTGGCTGTCAACATGGTTCCCTT
Bek (Ben A_F/ BenA_R) : 389 pb
GTTTCCAATGCGGGGTTGCCGCTGCATGGTGGCCGCGGGGATTGGGATTCAGGGGTAC CCTCCGGATCTCGGGATTATTGAAGAATGCCCGATCGTATGATGGGAGTTTCTCCGCC TGTCCCCTCGCCGAAGGTGTCCGACACTGTCGTCGAGCCCTACAACGCGACGCTCTCT GTACACCAGCTCGTTGAGAACTCGGACCAGACCTTCTGTATCGACAACGAGGCGCTCT ACGACATCTGCTTCAGGACGCTTAAGCTCACCACGCCCACATATGGTGACCTCAACCA CCTCGTCTCGATCGTCATGTCGGGTATCACGACTTGCTTGCGCTTCCCTGGTCAGCTCA ACTCGGACCTCAGGAAGTTGGCTGTCAACATGGTTCCCTT
Raja (Ben A_F/ BenA_R) : 342 pb
CATGGGTACCCTCCTGATCTCCGGGATTAGTGAAGAATACCCCGATCGGATGATGGCC ACCTTCTCCGTTGTCCCCTCGCCGAAGGTGTCCGACACTGTCGTCGAGCCCTACAACG CGACGCTCTCTGTTCACCAGCTCGTTGAGAACTCGGACCAGACCTTCTGTATCGACAA CGAGGCGCTCTACGACATCTGCTTCAGGACGCTTAAGCTCACCACGCCCACATATGGT GACCTCAACCACCTCGTGTCCATCGTCATGTCGGGTATCACGACTTGCTTGCGCTTCCC TGGTCAGCTCAACTCGGACCTCCGGAAGTTGGCTGTCAACATGGTTCCCTT
T9 (Ben A_F/ BenA_R) : 198 pb
TCGAGTTCTGACTGGGTTGTAGCTGGCCTTCCGAGGCATGTGCACGCCCTGCTCATCC ACTCTACACCTGTGCACTTACTGTGGGTTTACGGGTCGTGAAACGGGCTCGTTTATTC GGGCTTGTTGAGCGCACTTTGTTGCCTGCGTTTATCACAAACTCTATAAAGTATCAGA ATGTGTATTGCGATGTAACGCATC
Lipi (BenA_F/BenA_R) : 390 pb
TCACGCCGGGTTGGGGCGGGCTCTCCTGTATGGCCGCAGGGATGGCGGCCGGGGTTC CCTCCTGGTCTCCGGGATCCGTGTGGAGTGCCCGGACCGAATGATGGGCGCCTTCTCC GTCGTGCCATCCCCCAAGGTGTCCGACACCGGCGTTGAGCCCTACAACGCCGCCCTCT CCGTCCACCAGCTGGTCGAGAACTCCGACGAGACCTTCTGCATTGACAACGAGGCTCT CTACGACATCTGCATGCGCACCCTCAGGCTGTCCAACCCTGCCTACGGTGACCTGAAC TACCTCGTCTCCGCTGTCATGTCCGGCATCACCACCTGTCTGCGATTCCCCGGTCAGCT TAACTCTGATCTGCGCAAGCTGGCTGTCAACATGGTTCCCTT
Raja (Gan_ITS_1_F/ Gan_ITS1_R) : 202 pb
TCGAGTTTTGACTGGGTTGTAGCTGGCCTTCCGAGGCATCGTGCACGCCCTGCTCATC CACTCTACACCTGTGCACTTACTGTGGGTTATAGATCGTGTGGAGCGAGCTCGTTCGT TTGACGAGTTCGCGAAGCGCGTCTGTGCCTGCGTTTTATCACAAACACTATAAAGTAT TAGAATGTGTATTGCGATGTAACGCATC
Laras (Gan_ITS_1_F/ Gan_ITS1_R) : 147 pb
Lampiran 4b Hasil pengurutan basa nukelotida (
sequencing
) isolat
Ganoderma
spp. (warna merah = primer yang digunakan untuk amplifikasi
DNA) (lanjutan)
Tin (Gan_ITS_1_F/ Gan_ITS1_R) : 202 pb
TCGAGTTTTGACTGGGTTGTAGCTGGTTTTTCGAGGCATCGTGCACGCCCTGCTCATCC ACTCTACACCTGTGCACTTACTGTGGGTTATAGATCGTGTGGAGCGAGCTCGTTCGTTT GACGAGTTCGCGAAGCGCGTCTGTGCCTGCGTTTTATCACAAACACTATAAAGTATTA GAATGTGTATTGCGATGTAACGCATC
Lipi (Gan_ITS_1_F/ Gan_ITS1_R) : 994 pb
TTTGGGGTCGGGACTGGGGAGGGCTGGCAGATGTGGGGGCTGGGAGGCGTTCGCTGT AGAGCTTGAGGTTCTATAGTGTGTCCTAAATAGCTTGGCGTAATCATGGTCATAGCTG TTTCCTGTGTGAAATTGTTATCCGCTCACAATTCCACACAACATACGAGCCGGAAGCA TAAAGTGTAAAGCCTGGGGTGCCTAATGAGTGAGCTAACTCACATTAATTGCGTTGCG CTCACTGCCCGCTTTCCAGTCGGGAAACCTGTCGTGCCAGCTGCATTAATGAATCGGC CAACGCGCGGGGAGAGGCGGTTTGCGTATTGGGCGCTCTTCCGCTTCCTCGCTCACTG ACTCGCTGCGCTCGGTCGTTCGGCTGCGGCGAGCGGTATCAGCTCACTCAAAGGCGGT AATACGGTTATCCACAGAATCAGGGGATAACGCAGGAAAGAACATGTGAGCAAAAG GCCAGCAAAAGGCCAGGAACCGTAAAAAGGCCGCGTTGCTGGCGTTTTTCCATAGGC TCCGCCCCCCTGACGAGCATCAAAAAAAATCGACGCTCAAGTCAGAGGTGGCGAAAC CCGACAGGGACTATAACAGATACCAGGCGTTTCCCCCTGGAAGCTCCCTCGTGCGCTC TCCTGTTCCGACCCTGCCGCTTACCGGATACCTGTCCGCCTTTCTCCCTTCGGGGAAGC GTGGCGCTTTCTCATAGCTCACGCTGTAAGGTATCTCAGTTCGGTGTAGGGTCGGTTC GCTCCAAGGCTGGGCTGTGTGCACGAACTCCTCCGTTTCAGCCCGGACCGCTGCGCCT TATCCCGGTAACTTATTCGTTCTTTGAGTTCCCACCCGGGTAGAACACGACTTTATTCG CCCACTGGCAGCAGCCACTGGGTTAACAGGGAATTAGCAGAGCGGAGGTATTGTAAG CCGGTCCTACAAAGTCTTGGAGTGGTGGCCCTTACTTACGGTACCTAGAGAACAGTTA TTTGGATATCCG
Bek (Gan_ITS_1_F/ Gan_ITS1_R) : 1023 pb
Lampiran 4c Hasil pengurutan basa nukelotida (
sequencing
) isolat
Ganoderma
spp. (warna merah = primer yang digunakan untuk amplifikasi
DNA) (lanjutan)
Raja (ITS 1_F/ ITS4_B) : 831 pb