www.elsevier.nlrlocateraqua-online
The nutritional value of Palmaria mollis cultured
under different light intensities and water exchange
rates for juvenile red abalone Haliotis rufescens
Gunther Rosen, Chris J. Langdon
), Ford Evans
Hatfield Marine Science Center and Department of Fisheries and Wildlife, Oregon State UniÕersity, Newport,
OR 97365, USA
Accepted 21 October 1999
Abstract
A growth experiment with juvenile red abalone was conducted to determine whether the
Ž .
nutritional quality of red macroalga dulse Palmaria mollis was dependent on dulse culture Ž y1.
conditions. Dulse culture conditions differed by seawater volume exchange rate 1, 6, or 35 d
Ž y1.
and supplemental illumination 0 or 24 h d , creating a variety of dulse morphologies, biochemical compositions, and palatabilities for red abalone. The results of this study agreed with those of previous studies, showing dulse to be of high nutritional quality for abalone, with a relatively high protein content when compared with other macroalgae. Specific growth rates
ŽSGR of abalone fed on all dulse diets were higher than those of abalone fed on kelp Nereocystis. Ž .
luetkeana . In general, abalone grew faster on dulse cultured with supplemental illumination.
Ž .
Protein content of dulse 10.85–18.22% dry wt. , however, was directly dependent on seawater volume exchange per day, with highest protein levels observed in dulse cultured with 35 volume exchanges dy1. We suggest that other variables apart from biochemical composition, such as
epiphytic diatoms and morphological differences, contributed to the higher growth rates of abalone fed on light-supplemented diets.q2000 Elsevier Science B.V. All rights reserved.
Keywords: Haliotis; Dulse; Co-culture; Polyculture; Growth; Feeding rate; Protein; Diet; Macroalga
1. Introduction
Culture of red abalone in the United States has traditionally been based on abalone fed a diet of Macrocystis spp. or Nereocystis luetkeana harvested from coastal kelp
)Corresponding author. Tel.:q1-541-867-0231; fax:q1-541-867-0105.
Ž .
E-mail address: [email protected] C.J. Langdon .
0044-8486r00r$ - see front matterq2000 Elsevier Science B.V. All rights reserved. Ž .
( ) G. Rosen et al.rAquaculture 185 2000 121–136
122
Ž .
beds Hahn, 1989 . The absence of a reliable supply of these seaweeds outside of
Ž .
California Norman-Boudreau, 1988 , as well as the economic and environmental
Ž .
considerations in harvesting large quantities of kelp Mercer et al., 1993 , have prevented the expansion of abalone farming to other coastal areas. Co-culture of the red seaweed, Palmaria mollis, and red abalone, at Oregon State University, Hatfield Marine
Ž .
Science Center HMSC , has shown promise as an efficient method for abalone culture. This land-based approach will allow abalone aquaculture to be widely practiced in the Pacific Northwest. By growing the two commercially important species together, seawater-pumping costs can be reduced due to the seaweed’s ability to remove toxic ammonia excreted by abalone. In addition, dulse provides a nutritious food source for abalone.
P. mollis has been shown to be a fast-growing and effective species for removal of
Ž .
ammonia, nitrate, and phosphate from aquaculture effluents Levin, 1991 . The alga also
Ž .
has been shown to support juvenile abalone shell length SL growth rates as high as 3.9
y1 Ž .
mm month Evans, unpublished data . In a study of 56 species of marine algae occurring off the coast of Japan, a similar algal species, P. palmata, resulted in the
Ž .
highest monthly weight gain 51% of body weight , or an SL increase of 2.97 mm
y1 Ž .
month , when fed to H. discus hannaii Uki et al., 1986 . P. palmata is considered to be a well-balanced feed for abalone, with protein, lipid, and carbohydrate levels of 15,
Ž .
3–5, and 20–30%, respectively Mercer et al., 1993 .
The primary diet of red abalone in northern California waters consists of N.
Ž .
luetkeana, a brown macroalga commonly known as bull kelp Hahn, 1989 . When given
Ž .
a choice, red abalone prefer brown seaweed to red Tenore, 1976 . Like most marine algae, bull kelp is relatively poor in nutritional value and has a high moisture content ŽBritz, 1996 . Its abundance in the red abalone’s natural habitat and the soft texture of its.
Ž
fronds, however, may explain why it is a preferred diet of abalone McShane et al., .
1994 . Bull kelp also tends to possess low concentrations of polyphenolic compounds, which have been shown to deter grazing, making it particularly attractive to herbivores ŽWinter and Estes, 1992 . Ebert and Houk 1989 reported a high feeding rate of 16.8%. Ž . of mean body weight dy1 for red abalone fed this alga, but only a modest SL increase of
1.6 mm monthy1.
Ž .
Evans and Langdon in press investigated the effect of different dulse–abalone co-culture conditions on growth rate and maximum stocking density of red abalone.
Ž y1. Ž
Seawater volume exchange rate 1, 6, or 35 d and light exposure 0 or 24 h
y1.
supplemental illumination d resulted in different dulse growth rates and morpholo-gies, creating potential changes in food quality for abalone. It is conceivable that
Ž .
experimental culture conditions reported by Evans and Langdon in press resulted in dulse that differed in nutritional value for abalone because protein content of macroalgae
Ž .
is often affected by both light intensity Lapointe and Tenore, 1981 and seawater flow
Ž .
rate Subander et al., 1993 . Dulse morphology and texture may also affect its food quality because abalone have been shown to have preference for soft macroalgal thalli ŽWinter and Estes, 1992; McShane et al., 1994; Stepto and Cook, 1996 ..
Ž The objective of this study was to determine the effects of culture conditions 1, 6, or
y1 y1.
Ž .
Langdon in press in that abalone were fed on the different algal types separately from the co-culture system; therefore, culture conditions for dulse did not have a direct influence on abalone growth. A diet of wild-harvested bull kelp, N. luetkeana, was used as a control diet.
2. Materials and methods
2.1. Experimental organisms
Ž .
Juvenile red abalone 11 mm SL"0.3 mm were supplied by The Cultured Abalone, Santa Barbara, California, where they were reared on a diet consisting of kelp,
Macrocystis pyrifera. P. mollis was originally obtained from Fidalgo Island,
Washing-Ž ton, for culture at HMSC, Newport, Oregon. Dulse was cultured in 110-l tanks 0.155
2 . y1
m surface area at a stocking density of 9 g wet wt. l . Dulse culture conditions varied
Ž y1.
by seawater exchange rate 1, 6, or 35 seawater volume exchanges d and light
Ž y1.
exposure 0 or 24 h supplemental illumination d . Supplemental illumination was supplied by 1000-W metal halide bulbs, which provided 21–24 mol photon my2 dy1 of
Ž .
photosynthetically active radiation PAR at the water surface. Average daily PAR supplied by sunlight alone ranged from 29 mol my2 dy1 in July to 7 mol my2 dy1 in
September. Seawater for the dulse cultures was sand-filtered and UV-disinfected. All Ž
dulse cultures were fertilized to excess on a continual basis with nitrogen added as
. Ž . Ž
NaNO3 and phosphate added as NaH PO2 4 as described by Evans and Langdon in .
press . Cultures had been established for approximately 1 year before the start of this
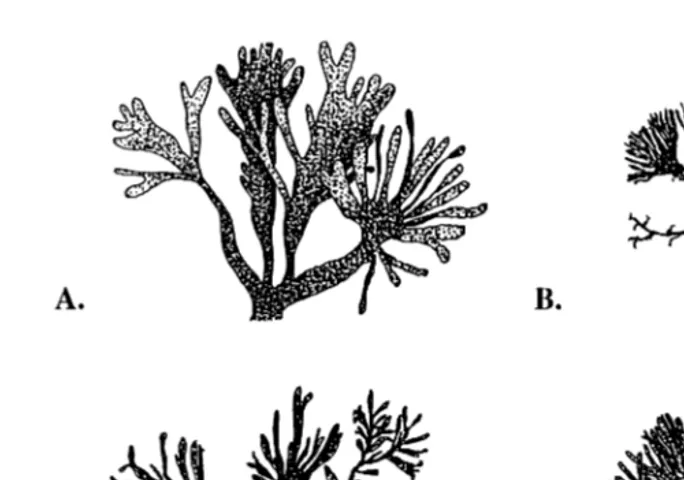
Ž .
study, during which time morphological differences became apparent Fig. 1 . Tempera-tures of dulse culTempera-tures during the experimental period ranged from 17.28C to 19.08C.
Ž .
The control diet of bull kelp N. luetkeana , was collected weekly from the north jetty at Yaquina Bay, Newport, Oregon.
2.2. Experimental design
Ž .
The abalone feeding experiment was conducted over an 8-week 56 days period from July to September, 1997, at HMSC. Dietary treatments consisted of dulse cultured under one of six different combinations of seawater exchange rate and light exposure ŽTable 1 , while the seventh treatment consisted of fronds cut into 1 in. squares. Ž .
Ž .
removed from freshly collected N. luetkeana bull kelp . Treatments were replicated fourfold.
High-density polyethylene containers with a 370-ml seawater capacity and 221 cm2
of submerged surface area served as experimental chambers, each of which housed 10 abalone. The containers were covered with a black plastic film to inhibit diatom growth
Ž
as well as to promote maximum feeding by juvenile abalone Ebert and Houk, 1984; .
( ) G. Rosen et al.rAquaculture 185 2000 121–136
124
Ž . Ž
Fig. 1. Morphologies of dulse P. mollis after 1 year of culture under different light 0 or 24 h light
y1. Ž y1.
supplementation d and seawater exchange rate 1 or 35 vols. d combinations. Broad thalli were typical
Ž . Ž .
of dulse in the 24 hr35=treatment A , narrow thalli were typical of dulse in the 0 hr1=treatment D , and
Ž . Ž .
intermediate thalli between A and D were typical of the 0 hr35=treatment C . Very fine thalli were typical
Ž .
of slow-growing dulse in the 24 hr1=treatment B .
approximately 100 ml miny1
, or 389 volume exchanges dy1
. The containers were randomly distributed in a flow-through water bath. The containers were checked daily to ensure food availability and to monitor water flows. Seawater temperature was recorded
Ž .
at 2-h intervals with a temperature logger StowAway, Onset, MA, USA . Average
Ž .
seawater temperature was 13.6"1.58C "1 standard deviation, ranges11.8–18.58C during the experimental period. Salinity of incoming water was automatically measured every 6 min by an OMEGA non-contact conductivity sensor, model CDCN-108.
y1 Ž
Average salinity was 31.3"1.2 g l "1 standard deviation, ranges25.1–33.1 g
y1.
l during the experimental period.
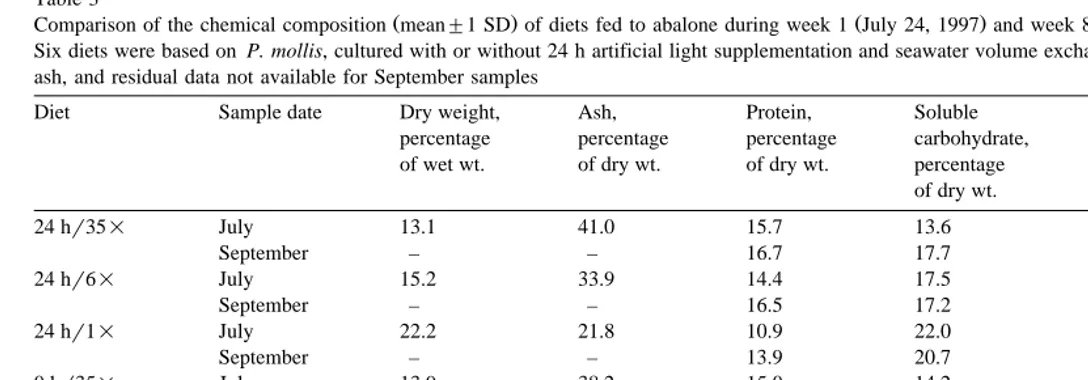
Table 1
Experimental dietary treatments tested with juvenile red abalone
Treatment Mean initial Diet Dulse culture method Frond Presence
a b
abalone size Supplemental light Exchange rate morphology of epiphytes
y1 y1
5 10.76"0.12 P. mollis 24 6 medium moderate
6 10.96"0.12 P. mollis 24 35 wide moderate
c
7 11.49"0.14 N. luetkeana – – wide low
a Ž
Supplemental light refers to number of hours of exposure to artificial lighting 21.1–24.2 mol photons
y2 y1. Ž y2 y1.
m d in addition to ambient light 7.0 mol m d . b
Exchange rate expressed as the number of total volume seawater exchanges of 110-l culture vessels over a 24-h period.
c
N. luetkeana was collected at the north jetty of the Yaquina estuary, Newport, Oregon.
2.3. Measurements
SL was recorded on day 1 and on day 56 of the study by measuring the longest axis
Ž .
of the shell to the nearest 0.1 mm with calipers. Mean SL increase for the experimental
y1 Ž .
period was reported in mm d for each treatment. Specific growth rate SGR , feed
Ž . Ž .
conversion efficiency FCE , and daily feed consumption DFC were measured and reported using ash-free dry weights of both abalone tissue and algae, unless otherwise stated. SGR, expressed as percent change in natural log of the ash-free dry wt. dy1
, was calculated using the formula:
SGRs100U
Ž
ln Wfyln Wi.
rdŽ .
1where W was the final mean ash-free dry weight of tissue, W was the initial meanf i
Ž .
ash-free dry weight of tissue, and d was the experimental period days .
Ž .
Initial wet weights were determined after blotting-dry the abalone see above . Four groups of 10 abalone, randomly sampled from the same population used for the growth experiment, were weighed at the beginning of the experiment for determination of initial wet tissue weight, dry tissue weight, and ash-free dry tissue weight. Tissues were dried at 608C for 24 h to determine dry weight. Dried abalone tissues were subsequently placed in a muffle furnace for 24 h at 4208C for ash weight determination. Ash-free dry weights of tissue were calculated by subtracting ash weights from dry tissue weights. Final weights of abalone used in the study were calculated using the same procedures. Dry weights and ash contents of macroalgal diets were also determined in this manner. FCE, used as a measure of how efficiently the diet was used for growth, was calculated using the formula:
FCEs100U
W yW r F yF 2
Ž
f i.
Ž
g u.
Ž .
Ž .
where W was the final ash-free dry weight g of abalone tissue, W was the initialf i
Ž . Ž .
( ) G. Rosen et al.rAquaculture 185 2000 121–136
126
Ž .
and F was the ash-free dry weight g of food uneaten for the whole experimentalu
period.
The DFC rate was estimated as the ash-free dry weight of algae consumed per day,
Ž y1.
expressed as a percentage of the ash-free dry tissue weight of abalone %BW d ,
Ž .
according to a formula modified from Britz et al. 1997 :
DFCs
Ž
FyR. Ž
r WUT.
Ž .
3Ž y1 y1.
where DFC was the daily consumption g algae g abalone d , F was the ash-free Ž .
dry weight g of feed offered over the experimental period, R was the ash-free dry Ž .
weight g of remaining feed collected over the experimental period, W was the average
Ž .
weight of abalone ash-free dry weight of tissue over the experimental period, and T
Ž .
was the experimental period days .
2.4. Biochemical composition
Protein, carbohydrate, and lipid content of the seaweed diets were analyzed using
Ž .
methods modified from Mann and Gallager 1985 . At the beginning and end of the experiment, one sample was collected for each diet by randomly sampling dulse from each of the culture conditions. The samples were stored aty808C until analysis. Frozen
y3 Ž
samples were freeze-dried for 24 h at less than 133=10 mbar LABCONCO .
freeze-dryer . Freeze-dried samples were subsequently pulverized in a Fritsch planetary micromill and stored in glass scintillation vials at 48C until analysis.
Protein was precipitated from 25 to 30 mg sub-samples of each diet with 5% wrv
Ž .
trichloroacetic acid TCA . Protein was then extracted from the precipitate by heating at 608C for 30 min in 5 ml of a solution of 2% wrv Na CO dissolved in 0.1 M NaOH,2 3 followed by a second extraction with 5 ml 1 M NaOH under the same temperature conditions. Supernatants from the two extractions were combined. The concentration of
Ž .
dissolved protein was determined using the BCA protein assay method Pierce , in which 100 ml of the protein solution was mixed with 2 ml of BCA reagent mixture, and the absorbance of the resulting solution was determined at 562 nm after incubation at
Ž .
378C for 30 min. Bovine serum albumin BSA was used for construction of a standard curve.
Soluble carbohydrate was determined in 2–3 mg freeze-dried, powdered macroalgae
Ž .
samples by extraction with 10 ml 0.1N H SO for 1 h Morgan and Simpson, 1981a .2 4
Ž .
The phenol–sulfuric acid method described by Dubois et al. 1956 was subsequently used to determine soluble carbohydrate by reading absorbance at 490 nm, and compar-ing absorbances with glucose standards.
Lipids were extracted from 25 to 30 mg sub-samples of the prepared samples with
Ž .
chloroform and methanol as described by Mann and Gallager 1985 and weights
Ž .
gravimetrically determined. Fish oil controls Zapata Haynie, feed grade menhaden oil were used to correct for weight changes during lipid extraction and weighing.
2.5. Statistical analyses
Ž .
Single-factor ANOVA P-0.05 was used to determine whether there was a dietary treatment effect on SL, SGR, FCE, and DFC. When null hypotheses were rejected,
Ž .
Tukey’s HSD multiple range tests P-0.05 were used to determine significant differences among treatments. Transformation of data was deemed unnecessary after
Ž testing for homogeneity of variances by box plot analysis. Two-factor ANOVA P
-.
0.05 was used to determine overall effects of light and water volume exchange rate on algal food quality for abalone. Simple linear regressions were used to determine if significant relationships existed between specific variables.
3. Results
3.1. Abalone growth and surÕiÕal
Ž .
Abalone survival was high Table 2 , except in one of the replicates with N.
luetkeana, which was eliminated from the analysis. The remaining containers exhibited
no significant difference in survival and all abalone appeared healthy upon visual inspection at termination of the study.
Most of the difference in dulse nutritional value was attributed to light
supplementa-Ž .
tion ANOVA; P-0.01 , as water exchange rate alone had no effect on abalone growth ŽANOVA; P)0.05 . There was also an interaction effect ANOVA; P. Ž -0.05 between. light and exchange rate on the nutritional value of dulse for abalone.
Ž
Shell growth differed among abalone fed on the seven dietary treatments ANOVA;
. y1
All of the dulse diets resulted in higher SGR than that of abalone fed on N.
Ž .
luetkeana, although differences were not always significant Table 2 . SGR of dulse-fed y1 Ž . y1 Ž .
abalone ranged from 1.23% d 0 hr1= to 1.42% d 24 hr35= . As with SL data, Tukey’s test indicated that both the 24 hr35= and 24 hr1= diets supported
Ž .
greater growth than the 0 hr1=diet Tukey’s HSD test; P-0.01 .
3.2. Feed conÕersion efficiency
Ž . Ž
Among the dulse treatments, FCE ranged from 18.2% 0 hr1= to 22.6% 24
. Ž .
hr35= Table 2 . Two-factor ANOVA indicated that both water volume exchange
Ž . Ž .
rate P-0.001 and light supplementation P-0.05 had an effect on FCE. Water Ž
volume exchange rate and light supplementation also had an interaction effect ANOVA;
. Ž
P-0.01 on FCE. FCE of abalone fed on the 24 hr35= diet was higher Tukey’s; .
P-0.001 than for abalone fed on all other diets, indicating that abalone converted this
Ž .
()
G.
Rosen
et
al.
r
Aquaculture
185
2000
121
–
136
128
Table 2
Ž . Ž . Ž . Ž . Ž .
Survival, shell length SL increase, specific growth rate SGR , feed conversion efficiency FCE , and daily feed consumption DFC mean"1 SD of juvenile red
Ž
abalone after 8 weeks of feeding on experimental dulse diets. Values in the same column followed by a common letter are not significantly different Tukey’s HSD
.
test; P)0.05
Ž
Calculations are based on either ash-free dry weights or wet weights of abalone tissue and algae. P. mollis diets differed by their culture conditions hours of
.
supplemental illumination, h, and number of seawater volume exchanges per day,=. ns4 for the dulse treatments, while ns3 for the N. luetkeana treatment.
y1 y1
Ž . Ž . Ž . Ž . Ž .
Diet Survival % SL increase mm d SGR % d FCE % DFC %
Dry wt. Wet wt. Dry wt. Wet wt. Dry wt. Wet wt.
24 h, 35= 100 62.81"3.49 a 1.42"0.07 a 1.40"0.08 a 22.60"0.66 a 13.51"0.59 cd 5.68"0.18 b 10.64"0.33 b 24 h, 1= 100 61.21"2.56 a 1.41"0.04 a 1.38"0.04 ab 18.68"0.35 b 24.90"0.98 a 6.83"0.24 a 5.70"0.23 d 24 h, 6= 97.5 59.78"5.03 ab 1.33"0.08 ab 1.29"0.07 ab 19.60"0.85 b 15.15"0.34 c 6.06"0.41 ab 8.71"0.60 c 0 h, 6= 100 56.22"3.98 ab 1.36"0.03 ab 1.33"0.12 ab 20.08"1.66 b 14.33"0.41 d 6.07"0.59 ab 9.49"0.65 bc 0 h, 35= 100 54.60"3.50 ab 1.27"0.09 ab 1.22"0.07 ab 19.55"0.93 b 12.91"0.35 d 5.73"0.34 b 9.64"0.62 bc 0 h, 1= 100 51.56"3.49 b 1.23"0.10 b 1.23"0.12 b 18.19"0.53 b 19.38"1.30 b 5.91"0.46 b 6.47"0.51 d
comparable to most FCE values of abalone fed on dulse diets when expressed in terms of ash-free dry weight, but was lower than for all dulse diets when calculated on a wet
Ž . Ž .
weight basis Tukey’s; P-0.001 Table 2 .
3.3. Daily feed consumption
Feed consumption ranged from 5.42 %BW dy1 for abalone fed on N. luetkeana to
y1 Ž .
6.83 %BW d for abalone fed on the 24 hr1= dulse diet Table 2 . Two-factor ANOVA indicated that light did not have an effect on ash-free dry weight consumption
Ž .
of dulse ANOVA; P)0.05 , but the effect of water volume exchange rate on DFC was significant. There was also a significant interaction effect between light and exchange
Ž .
rate on DFC. The highest DFC 24 hr1= differed from those of abalone fed on 24
Ž .
hr35=, 0 hr35=, and 0 hr1=dulse diets Tukey’s HSD test; P-0.01 . Although all dulse diets, on an ash-free dry weight basis, resulted in higher DFC rates than for abalone fed N. luetkeana, on a wet weight basis, Nereocystis was consumed at about twice the rate as the 6=and 35=dulse diets and three times the rate as the 1=dulse
Ž .
diets Table 2 .
3.4. Chemical composition of abalone diets
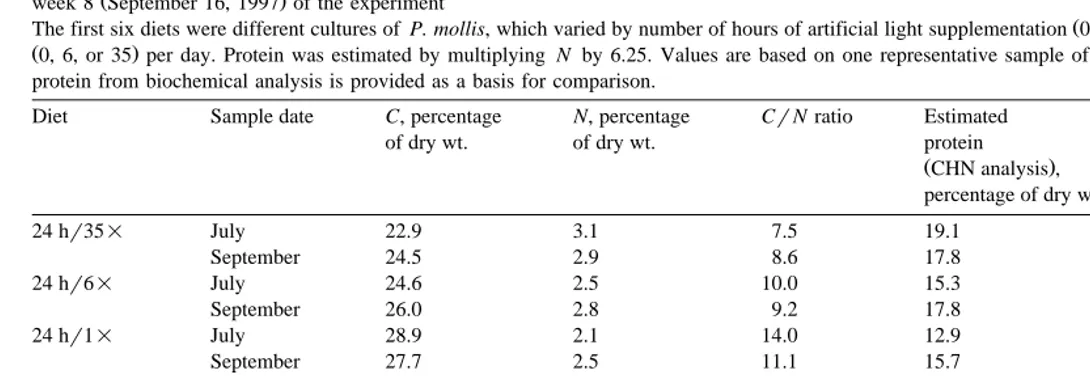
Ž Protein content of dulse samples was lower earlier in the study than at the end Table
. Ž . Ž .
3 , ranging from 10.9% 24 hr1= to 15.7% 24 hr35= of dry weight in July, and
Ž . Ž .
13.9% 24 hr1= to 18.2% 0 hr6= in September. The general trend was similar at both dates, however, with protein content primarily affected by water exchange rate. This was especially evident in the July samples, in which protein increased with
Ž .
increasing number of seawater volume exchanges from 1 to 35 per day. Likewise, September samples exhibited relatively low protein in the 1=diets, but the difference
Ž .
between 6=and 35=samples was not as apparent Table 3 . Light conditions did not appear to have an effect on protein content in July, but protein was somewhat higher in the absence of supplemental illumination in September. Protein content of wild N.
Ž .
luetkeana was comparable to that of dulse diets in July 13.5% , but was the lowest
Ž . Ž .
among all diets tested in September 10.8% Table 3 . In July, a strong positive Ž 2
correlation between protein and ash content of dulse diets was observed r s0.95, .
ns6 . Ash data were not available for September samples; thus, no comparison was possible.
By multiplying measured N values by 6.25, CHN analysis resulted in similar protein
Ž .
estimates to those obtained biochemical analyses Table 4 . In July, dulse protein
Ž . Ž .
estimates ranged from 12.9% 24 hr1= to 19.1% 24 hr35= , while in September,
Ž . Ž .
they ranged from 15.7% 24 hr1= to 19.7% 0 hr6= . CrN ratios decreased with
Ž .
increasing exchange rate in both July and September Table 4 .
In September, there appeared to be a relationship between protein content of dulse
Ž 2 .
and SGR of abalone in the 0 h light treatments r s0.88, ns3 , but not in the 24 h
Ž 2 .
light treatments r s0.12, ns3 . The relationship between protein and SGR was not
Ž 2 . Ž 2 .
()
G.
Rosen
et
al.
r
Aquaculture
185
2000
121
–
136
130
Table 3
Ž . Ž . Ž .
Comparison of the chemical composition mean"1 SD of diets fed to abalone during week 1 July 24, 1997 and week 8 September 16, 1997 of the experiment Six diets were based on P. mollis, cultured with or without 24 h artificial light supplementation and seawater volume exchange rates of 1, 6, or 35 dy1. Dry weight, ash, and residual data not available for September samples
Diet Sample date Dry weight, Ash, Protein, Soluble Lipid, Residual,
percentage percentage percentage carbohydrate, percentage percentage
of wet wt. of dry wt. of dry wt. percentage of dry wt. of dry wt.
of dry wt.
24 hr35= July 13.1 41.0 15.7 13.6 2.8 26.9
September – – 16.7 17.7 3.2 –
24 hr6= July 15.2 33.9 14.4 17.5 3.3 31.9
September – – 16.5 17.2 2.7 –
24 hr1= July 22.2 21.8 10.9 22.0 2.4 43.0
September – – 13.9 20.7 2.3 –
0 hr35= July 13.9 38.2 15.0 14.2 3.0 29.6
September – – 17.1 15.1 3.1 –
0 hr6= July 14.4 36.0 14.1 18.2 2.3 29.4
September – – 18.2 17.9 2.6 –
0 hr1= July 17.9 26.7 11.2 22.3 3.0 36.8
September – – 15.3 24.6 2.4 –
N. luetkeana July 8.5 51.6 13.5 1.9 2.5 30.5
()
G.
Rosen
et
al.
r
Aquaculture
185
2000
121
–
136
131
Ž . Ž . Ž .
Comparison of the total carbon C , nitrogen N , CrN ratios, and estimated protein contents of the seven diets fed to abalone during week 1 July 24, 1997 and
Ž .
week 8 September 16, 1997 of the experiment
Ž .
The first six diets were different cultures of P. mollis, which varied by number of hours of artificial light supplementation 0 or 24 h and seawater volume exchanges
Ž0, 6, or 35 per day. Protein was estimated by multiplying N by 6.25. Values are based on one representative sample of food that was fed to abalone. Estimated.
protein from biochemical analysis is provided as a basis for comparison.
Diet Sample date C, percentage N, percentage CrN ratio Estimated Estimated
of dry wt. of dry wt. protein protein
ŽCHN analysis ,. Žbiochemical analysis ,.
percentage of dry wt. percentage of dry wt.
24 hr35= July 22.9 3.1 7.5 19.1 15.7
September 24.5 2.9 8.6 17.8 16.7
24 hr6= July 24.6 2.5 10.0 15.3 14.4
September 26.0 2.8 9.2 17.8 16.5
24 hr1= July 28.9 2.1 14.0 12.9 10.9
September 27.7 2.5 11.1 15.7 13.9
0 hr35= July 22.3 2.9 7.6 18.4 15.0
September 24.5 3.1 7.8 19.6 17.1
0 hr6= July 24.7 2.6 9.6 16.1 14.1
September 26.1 3.2 8.3 19.7 18.2
0 hr1= July 27.2 2.1 13.0 13.1 11.2
September 28.6 2.5 11.3 15.8 15.3
N. luetkeana July 20.9 3.0 7.0 18.7 13.5
( ) G. Rosen et al.rAquaculture 185 2000 121–136
132
Soluble carbohydrate content of dulse was inversely correlated with protein content,
Ž 2 . Ž
particularly in July dulse samples r s0.93, ns6 . Levels ranged from 13.6% 24
. Ž . Ž .
hr35= to 22.3% 0 hr1= of dry weight in July, and 15.1% 0 hr35= to 24.6% Ž0 hr1=. in September Table 3 . Soluble carbohydrate extracted from N. luetkeanaŽ . samples was much lower than that for dulse, with 1.9% present in July and 1.6% present
Ž .
in September Table 3 . Samples of N. luetkeana in July contained 30.5% residual material that was likely largely composed of insoluble carbohydrate. Residual matter in
Ž . Ž . Ž .
dulse ranged from 26.9% 24 hr35= to 43.0% 24 hr1= in July Table 3 . Data for September residual material were unavailable due to the lack of ash weights for these samples.
Ž . Ž .
Lipid ranged from 2.3% 0 hr6= to 3.3% 24 hr6= of dry dulse weight in July,
Ž . Ž . Ž .
and 2.3% 24 hr1= to 3.2% 24 hr35= in September Table 3 . Although lipid content increased with exchange rate in September, no such relationship existed in July ŽTable 3 . Samples of N. luetkeana collected from the wild contained 2.5% lipid in July.
Ž .
and 2.2% in September Table 3 .
Ž . Ž .
Ash content of dulse in July ranged from 21.8% 24 hr1= to 41.0 % 24 hr35= ,
Ž . Ž
and increased with increasing exchange rate of seawater Table 3 . Dry weight per-.
centage of wet weight , on the other hand, decreased as flow rate increased, and ranged from 13.1% of wet algae weight in the 24 hr35=dulse diet to 22.2% in the 24 hr1=
Ž .
diet Table 3 .
4. Discussion
Ž
Even though red abalone generally prefer brown seaweed Tenore, 1976; Hahn, .
1989 , dulse was readily accepted by juvenile abalone and supported better growth rates
Ž .
than N. luetkeana Table 2 . In addition, the results of this study indicate that culture conditions of dulse had a significant effect on daily dulse consumption rate, FCE, and growth rate of abalone. Increases in SL and SGR of abalone indicate that dulse cultured under 24 h supplemental illumination had higher nutritional value for abalone than dulse cultured with only ambient light. Biochemical composition of the diets, however, was more dependent on water volume exchange rate, suggesting that other variables, such as presence of epiphytes and frond morphology, may have contributed to the observed differences in food value for abalone.
The 24 hr35= diet resulted in the greatest SL increase, SGR, and FCE, but the
Ž .
lowest consumption rate Table 2 , suggesting that relatively small quantities were required to achieve relatively fast growth. High protein content of dulse cultured with
Ž y1.
the maximum seawater volume exchange rate 35 exchanges d may have been partially responsible for the good growth of abalone fed on the 24 hr35= dulse ŽShpigel et al., 1999 . However, abalone fed on the 0 h. r35= diet, which had comparable protein contents to the 24 hr35= dulse diet, converted food relatively inefficiently and grew at a slower rate.
apparently responsible for the relatively high SGR and SL increase observed for abalone Ž fed on the 24 hr1=diet, which had the lowest protein content of all dulse diets Table
. Ž .
3 . DFC based on ash-free dry weights of abalone fed the 24 hr1= diet were significantly higher than for any of the other diets, suggesting that the abalone in this treatment attempted to make up for a nutritionally sub-optimal diet by higher consump-tion rates.
Ž .
The higher DFC based on ash-free dry weights of abalone fed the low protein 24 hr1=diet indicates that nutritional value is not necessarily positively correlated with abalone consumption rates. Dietary preference of abalone is believed to be based on many factors including morphology, texture, odor, and presence of unpalatable
chemi-Ž
cals, in addition to nutritional composition McShane et al., 1994; Fleming, 1995; . Marsden and Williams, 1996; Rivero and Viana, 1996; Stepto and Cook, 1996 . Although animals in this study were not given a choice, it has been shown that
Ž .
herbivores generally grow better on diets that they prefer Jensen, 1983 .
The presence of epiphytic diatoms, which tend to occur in dulse cultures with high
Ž .
irradiance Morgan and Simpson, 1981b , may have affected the nutritional value and palatability of the dulse. Several studies have shown that abalone and other herbivorous
Ž
gastropods grow best when fed mixed algal diets Watanabe, 1984; Mercer et al., 1993; .
Stuart and Brown, 1994 . In addition, diatoms are the preferred diet of abalone less than
Ž .
10 mm in SL Hahn, 1989; Kawamura et al., 1995 . Diatoms may have been more attractive andror easier to digest than dulse for the juvenile abalone. The relatively high
Ž .
DFC on an ash-free dry weight basis of abalone fed on the 24 hr1= diet, where epiphytes were particularly evident, may indicate a higher palatability for this mixed diet.
Ž .
The biochemical composition of P. mollis Table 3 was generally comparable to Ž
reported values of the similar alga, P. palmata Morgan and Simpson, 1981b; Mercer et .
al., 1993 . Protein contents between 10.9% and 18.2% of dry weight, depending on culture conditions, are high for macroalgae. CHN analysis resulted in similar, but slightly higher, protein estimates compared to biochemical determination of contents. It is likely that CHN analysis overestimated true protein content due to inclusion of other nitrogen-containing compounds in the algal tissue samples, such as nitrate.
Reduced protein content of dulse cultured with one volume exchange of seawater dy1
ŽTable 3 suggests that limited flow conditions deprived the algae of naturally occurring. inorganic nutrients necessary for protein synthesis. The strong positive correlation between protein and ash content of dulse in this study also implies that low flow cultures were not receiving adequate trace elements typically found in ash. The positive relationship between protein and ash content in dulse was also observed by Morgan and
Ž .
Simpson 1981a,b . Protein content in September samples was generally higher in
Ž . Ž .
cultures without supplemental illumination Table 3 . Morgan and Simpson 1981b found that plants exposed to low irradiance levels accumulated nitrogen and mineral
Ž .
elements ash due to their restricted growth. This relationship, however, was not observed in July dulse samples, indicating that other environmental factors besides light affected protein content.
Ž .
Although soluble carbohydrate levels Table 3 were lower than those reported for P.
Ž . Ž .
( ) G. Rosen et al.rAquaculture 185 2000 121–136
134
Ž 2 .
soluble carbohydrate was apparent in July r s0.92, ns6 . This relationship has been
Ž .
reported for P. palmata as well as for other macroalgae Morgan and Simpson, 1981b . It is suspected that insoluble carbohydrate was the primary component of the residual material of dulse samples after accounting for ash, protein, lipid, and soluble
carbo-Ž hydrate. The low soluble carbohydrate levels observed in N. luetkeana samples Table
.
3 suggest that the residual material contains primarily insoluble carbohydrates, as this
Ž .
species has been reported to consist of 39.8% total carbohydrate Hahn, 1989 .
Lipid content appeared to be positively correlated with flow rate in September, but the absence of such a relationship in July suggests that seasonal factors also influenced
Ž .
lipid content Table 3 . It is interesting to note, however, that lipid levels were within the
Ž . Ž .
range reported for P. palmata 0.2–3.8% of dry weight Morgan et al., 1980 . Ž
The wide fronds of the algae cultured with high water exchange rates 35 volume
y1.
exchanges d had relatively high moisture content compared to dulse cultured with only one volume exchange of seawater dy1. Calculations based on wet weight revealed
Ž .
that DFC was relatively high for abalone fed on 35=diets Table 2 , which could be attributed to high dulse moisture content. Conversely, the low ash-free dry weight DFC for 35=diets suggests that less of the dry weight of this diet was required by abalone to meet their daily feed requirements, possibly due to higher protein levels of these diets. Although abalone performed better when fed on dulse diets compared with kelp, growth rates of dulse-fed abalone observed in this study were lower compared to those
Ž .
reported in other studies Uki et al., 1986; Evans and Langdon, in press . This may be
Ž .
have been due to relatively low seawater temperatures in the study 13.68C"1.58C . Juvenile red abalone have been shown to display most rapid growth in the temperature
Ž .
range of 15–188C Leighton, 1974 . Higher temperatures likely would have stimulated increased feeding rates and may have resulted in larger differences in growth rates of abalone fed on the experimental diets.
In summary, this study showed that growth, FCE, and feed consumption rates of red abalone fed on dulse were affected by conditions used to culture the dulse. Culture conditions also resulted in different proportions of ash, protein, lipid, and carbohydrate content of dulse, which were primarily dependent on seawater exchange rate rather than light supplementation. Faster growth of abalone fed on dulse that had been cultured with 24 h light supplementation suggests that light played a role in increasing the nutritional quality of dulse. Epiphytic diatom growth as well as morphological differences of dulse supplemented with light may have contributed to the faster growth of abalone fed on these diets.
Acknowledgements
number AQ 96.077-7319-05 from the National Coastal Resource Research and Develop-ment Institute.
References
Britz, P.J., 1996. The suitability of selected protein sources for inclusion in formulated diets for the South African abalone, Haliotis midae. Aquaculture 140, 63–73.
Britz, P.J., Hecht, T., Mangold, S., 1997. Effect of temperature on growth, feed consumption and nutritional indices of Haliotis midae fed a formulated diet. Aquaculture 152, 191–203.
Dubois, M., Gilles, K.A., Hamilton, J.K., Pebers, P.A., Smith, F., 1956. Colorimetric method for determina-tion of sugars and related substances. Anal. Chem. 28, 350–356.
Ebert, E.E., Houk, J.L., 1984. Elements and innovations in the cultivation of red abalone Haliotis rufescens. Aquaculture 39, 375–392.
Ebert, E.E., Houk, J.L., 1989. Abalone cultivation methods used at the California Department of Fish and
Ž .
Game’s Marine Resources Laboratory. In: Hahn, K. Ed. , Handbook of Culture of Abalone and Other Marine Gastropods. CRC Press, Boca Raton, pp. 242–244.
Fleming, A.E., 1995. Growth, intake, feed conversion efficiency and chemosensory preference of the Australian abalone, Haliotis rubra. Aquaculture 132, 297–311.
Ž .
Hahn, K.O., 1989. Nutrition and growth of abalone. In: Hahn, K. Ed. , Handbook of Culture of Abalone and Other Marine Gastropods. CRC Press, Boca Raton, pp. 135–156.
Ž
Jensen, K.R., 1983. Factors affecting feeding selectivity in herbivorous ascoglossa Mollusca:
Opistho-.
branchia . J. Exp. Mar. Biol. Ecol. 66, 135–148.
Kawamura, T., Saido, T., Takami, H., Yamashita, Y., 1995. Dietary value of benthic diatoms for the growth of post-larval abalone Haliotis discus hannai. J. Exp. Mar. Biol. Ecol. 194, 189–199.
Lapointe, B.E., Tenore, K.R., 1981. Experimental outdoor studies with UlÕa fasciata: 1. Interaction of light and nitrogen on nutrient uptake, growth, and biochemical composition. J. Exp. Mar. Biol. Ecol. 53, 135–152.
Leighton, D.L., 1974. The influence of temperature on larval and juvenile growth in three species of southern California abalones. Fishery Bulletin 72, 1137–1145.
Levin, J.M., 1991. Land-based polyculture of marine macroalgae and Pacific salmon. M.S. thesis, Oregon State University, Corvallis, Oregon.
Mann, R., Gallager, S.M., 1985. Physiological and biochemical energetics of larvae of Teredo naÕalis L. and
Ž . Ž .
Banki gouldi Bartsch Bivalvia: Teredinidae . J. Exp. Mar. Biol. Ecol. 85, 211–228.
Marsden, I.D., Williams, P.M.J., 1996. Factors affecting the grazing rate of the New Zealand abalone Haliotis
iris Martyn. J. Shellfish Res. 5, 401–406.
McShane, P.E., Gorfine, H.K., Knuckey, I.A., 1994. Factors influencing food selection in the abalone Haliotis
Ž .
rubra Mollusca: Gastropoda . J. Exp. Mar. Biol. Ecol. 176, 27–37.
Mercer, J.P., Mai, K.S., Donlon, J., 1993. Comparative studies on the nutrition of two species of abalone,
Haliotis tuberculata Linnaeus and Haliotis discuss hannai Ino: I. Effects of algal diets on growth and
biochemical composition. Invertebr. Reprod. Dev. 23, 75–88.
Morgan, K.C., Simpson, F.J., 1981a. The cultivation of Palmaria palmata. Effect of light intensity and nitrate supply on growth and chemical composition. Bot. Mar. 24, 273–277.
Morgan, K.C., Simpson, F.J., 1981b. The cultivation of Palmaria palmata. Effect of light intensity and temperature on growth and chemical composition. Bot. Mar. 24, 547–552.
Morgan, K.C., Wright, J.L.C., Simpson, F.J., 1980. Review of chemical constituents of the red alga Palmaria
Ž .
palmata dulse . Econ. Bot. 34, 27–50.
Norman-Boudreau, K., 1988. Abalone nutrition and the potential role of purified diets. J. Shellfish Res. 7, 564. Rivero, L.E., Viana, M.T., 1996. Effect of pH, water stability and toughness of artificial diets on the
palatability for juvenile abalone Haliotis fulgens. Aquaculture 144, 353–362.
( ) G. Rosen et al.rAquaculture 185 2000 121–136
136
Stepto, N.K., Cook, P.A., 1996. Feeding preferences of the juvenile South African abalone Haliotis midae
ŽLinnaeus, 1758 . J. Shellfish Res. 15, 653–657..
Ž .
Stuart, M.D., Brown, M.T., 1994. Growth and diet of cultivated black-footed abalone, Haliotis iris Martyn . Aquaculture 127, 329–337.
Subander, A., Petrell, R.J., Harrison, P.J., 1993. Laminaria culture for reduction of dissolved inorganic nitrogen in salmon farm effluent. J. Appl. Phycol. 5, 455–463.
Tenore, K.R., 1976. Food chain dynamics of abalone in a polyculture system. Aquaculture 8, 23–27.
Ž .
Tutschulte, T.C., Connell, J.H., 1988. Feeding behavior and algal food of three species of abalone Haliotis in southern California. Mar. Ecol. Prog. Ser. 49, 57–64.
Uki, N., Sugiura, M., Watanabe, T., 1986. Dietary value of seaweeds occurring on the Pacific Coast of Tohoku for growth of the abalone Haliotis discus hannai. Bull. Jpn. Soc. Fish. 52, 257–266.
Ž
Watanabe, J.M., 1984. Food preference, food quality and diets of three herbivorous gastropods Trochidae:
.
Tegula in a temperate kelp forest habitat. Oecologia 62, 47–52.
Winter, F.C., Estes, J.A., 1992. Experimental evidence for the effects of polyphenolic compounds from
Ž .
Dictyoneurum californicum Ruprecht Phaeophyta: Laminariales on feeding rate and growth in the red