METODE PENELITIAN
Waktu dan Tempat
Penelitian dilaksanakan pada bulan November 2016 sampai Maret 2017.
Pengambilan sampel dilakukan di Desa Lubuk Kertang Kecamatan Brandan Barat
Kabupaten Langkat Provinsi Sumatera Utara, analisis laboratorium dilakukan di
SSPL (socfindoseed production and laboratory) Socfindo Desa Martebing Dolok
Masihul Kabupaten Serdang Bedagai Sumatera Utara dan di Laboratorium
Fitokimia Fakultas Farmasi, Universitas Sumatera Utara, Medan. Penelitian
dilaksanakan selama lima bulan dengan waktu kegiatan yang diperlukan untuk
penelitian adalah sebagai berikut: (a) Orientasi lapangan, (b) Pengumpulan bahan,
(c) Analisis data, dan (d) Penyusunan dan penulisan.
Alat dan Bahan
Alat yang digunakan dalam penelitian ini adalah micro pipette, pipette
tips, botol kocok, botol ekstrak, kertas saring, corong, aluminium foil, water bath
(incubator), silica gel 60-TLC, RP-18 silica gel TLC/2D TLC (two dimentional
thin layer chromathography), timbangan analitik, oven,mesin penghalus/grinding,
magnet batangan, hair dryer, oven, chamber, scanner Canon PIXMA E400, dan
alat tulis.
Bahan yang digunakan adalah bunga dan buah Hibiscus tilliaceus,
Melastoma candidum, dan Ricinus communis, pelarut chloroform, metanol,
n-hexane, etanol, KOH, aquades, toluene, etyl acetat, aceton, standard dolichol dan
Prosedur Penelitian 1. Pengumpulan sampel
Bunga dan buah H. tilliaceus, M. candidum, dan R. communis yang berada
di Desa Lubuk Kertang Kecamatan Brandan Barat Kabupaten Langkat Provinsi
Sumatera Utara dikumpulkan lalu sampel segar disimpan pada suhu -80oC sampai
digunakan.
2. Isolasi polyisoprenoid alkohol
Prosedur untuk isolasi polyisoprenoid seperti yang dideskripsikan oleh
Sagami et al, (1992). Bunga dan buah dikeringkan pada suhu 60-75 oC selama 1-2
hari. Jaringan bunga dan buah (masing-masing 2 g) dihancurkan menjadi bubuk
halus dan direndam dalam 30 ml pelarut kloroform/metanol (2:1 vol/vol) selama
48 jam. Ekstrak lipid dari bunga dan buah disaponifikasi pada suhu 65 oC selama
24 jam dalam konsentrasi 0,45 g KOH, 2 ml etanol, dan 2 ml aquades. NSL (non
saponifiable lipid) setiap jaringan kemudian dilarutkan dengan pelarut heksan
dan siap untuk dianalisis.
3. Analisis dengan dua dimensi kromatografi
Dimensi pertama TLC dilakukan selama 60 menit di atas silika gel (20x3
cm) dengan sistem pelarut toluen-etil asetat (9:1). Tepi longitudinal dari dimensi
pertama TLC dengan lebar 1 cm dan zona konsentrasi dari fase reverse RP-18
TLC yang dijepit dengan cara menggunakan dua magnetik batangan (4.0 x 1.1 x
0.8 cm) dengan menghadap setiap fase gel. Plat TLC yang terikat kemudian
dikembangkan tegak lurus ke dimensi pertama untuk transfer dolichol dan
Dimensi dua fase reverse RP-18 silika gel TLC dilakukan dengan pelarut
aseton selama sekitar 30 menit. Posisi polyisoprenoid alkohol dipisahkan dan
dikembangkan oleh silica gel TLC dua dimensi, kemudian diidentifikasi dan
divisualisasikan dengan uap yodium (iodine vapor). Gambar kromatografi
diperoleh dan discan.
Gambar 1. Metode kromatografi lapis tipis (TLC) untuk memisahkan polyprenol dan
dolichol
(Sagami et al, 1992).
4. Analisis Data
Konsentrasi dolichol dan polyprenol yang terdeteksi pada HPTLC RP-18
diukur menggunakan ImageJ 1.46r (Schneider et al., 2012) dengan standard
dolichol dan polyprenol sebagai referensi/acuannya yang digunakan untuk
HASIL DAN PEMBAHASAN
Hasil
Penelitian dilakukan untuk mengetahui distribusi kandungan senyawa
polyisoprenoid pada organ generatif, yaitu bunga dan buah tiga jenis mangrove
ikutan Langkat dan menghitung panjang rantai polyisoprenoid dengan cara
mamisahkannya menjadi dolichol dan polyprenol dengan menggunakan 2D-TLC
(two-dimentional thin layer chromatography) (Sagami et al., 1992). Tabel 1
menunjukkan hasil analisis total lipid dan kehadiran famili dolichol dan
polyprenol pada organ generatif jenis H. tilliaceus, M. candidum, dan
R. communis.
Dari Tabel 1 dapat kita lihat bahwa nilai total lipid tiga jenis mangrove
ikutan pada jaringan bunga berkisar antara 558,2-708,3 mg g-1 dw (dw = dry
weight/berat kering) dengan nilai rata-rata 625,57 mg g-1 berat kering. Nilai total
lipid terkecil terdapat pada bunga H. tilliaceus dengan nilai 558,2 mg g-1 berat
kering dan yang terbesar terdapat pada bunga R. communis dengan nilai 708,3
mg/g berat kering. Sedangkan pada jaringan buah, nilai total lipid berkisar antara
534,65-656,5 mg g-1 berat kering dengan nilai rata-rata 591,05 mg g-1 berat kering.
Dimana nilai total lipid terkecil dan terbesar terdapat pada jenis yang sama seperti
pada jaringan bunga dengan nilai sebesar 534,65 mg g-1 berat kering pada buah
H. tilliaceus dan 656,5 mg g-1 berat kering pada buah R. communis. Nilai ini
diperoleh dari nilai berat akhir ekstraksi bunga dan buah yang telah dihaluskan
dengan konsentrasi berat sampel 2 g di masing-masing jenis melalui serangkaian
kegiatan seperti yang telah dijelaskan di bab metode penelitian, yaitu pada isolasi
Tabel 1. Nilai total lipid dan distribusi kehadiran dolichol dan polyprenol pada tiga jenis mangrove ikutan H. tilliaceus, M. candidum, dan R. communis.
nd = not detected atau tidak terdeteksi, dw = dry weight atau berat kering
Jumlah kandungan polyisoprenoid pada jaringan bunga dan buah tiga jenis
mangrove ikutan adalah berkisar antara 13-103 mg g-1 berat kering (Tabel 1).
Adapun jumlah/nilai yang terkecil adalah pada jaringan buah jenis R. communis
(13 mg g-1 berat kering) dan yang terbesar adalah pada jaringan bunga jenis
H. tilliaceus (103 mg g-1 berat kering). Nilai ini diperoleh dari hasil penjumlahan
nilai kandungan senyawa dolichol dan polyprenol.
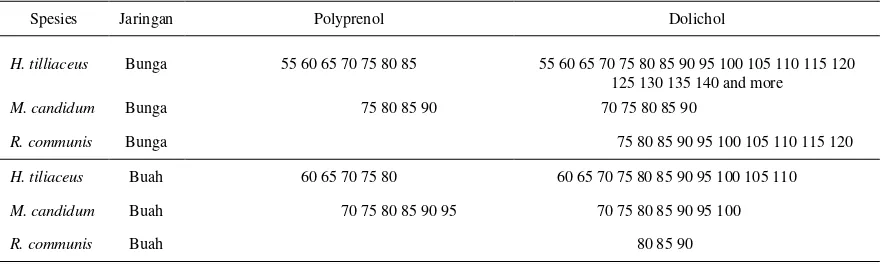
Tabel 2. Panjang rantai karbon polyprenol dan dolichol pada tiga jenis mangrove ikutan
H. tilliaceus, M. candidum, dan R. communis*
*Angka mengacu pada panjang rantai karbon polyisoprenoid alkohol.
Spesies Jari- Lipid total Polyiso
-prenoid Polyprenol Dolichol % di total lipid % di polyisoprenoid Tipe
ngan (mg/g dw) (mg/g dw) (mg/g) (mg/g)
Polyiso-prenoid Polyprenol Dolichol Polyprenol Dolichol
H. tiliaceus Bunga 558.2 103 17.8 85.2 18.4 3.2 15.3 17.3 82.7 II
Spesies Jaringan Polyprenol Dolichol
A B C
D E F
Gambar 2. Kromatografi lapis tipis dua dimensi (2D-TLC) polyisoprenoid alkohol dari bunga H. tiliaceus (A), bunga M. candidum (B), bunga R. communis (C), buah H. tiliaceus (D), buah M. candidum (E), dan buah R. communis (F). Angka karbon mengacu pada panjang rantai karbon polyisoprenoid alkohol.
Tabel 2 dan Gambar 1 menggambarkan distribusi senyawa dolichol pada
jaringan bunga dengan panjang rantai karbon C55-C140, C70-C90, dan C75-C120
ditemukan pada masing-masing jenis H. tiliaceus, M. candidum, dan
R. communis. Begitu juga senyawa polyprenol dengan panjang rantai karbon
C55-C85 dan C75-C90 ditemukan pada masing-masing jenis M. candidum dan
H. tiliaceus. Sedangkan pada jaringan buah, dolichol dengan panjang rantai
karbon mulai dari C60-C80, C70-C100, dan C80-C90 ditemukan pada
masing-masing jenis H. tiliaceus, M. candidum, dan R. communis. Polyprenol dengan
panjang rantai karbon C60-C80 dan C70-C100 ditemukan pada jenis H. tiliaceus
dan M. candidum. Dari Tabel 2 juga menunjukkan bahwa dolichol mendominasi
dan C80-C90. Adapun senyawa polyprenol tidak ditemukan baik pada jaringan
bunga maupun buah jenis R. communis.
Keluarga dolichol hadir dengan dua dominasi rantai panjang C80-C85
(Gambar 2) terdeteksi pada jaringan bunga R. communis. Pada jaringan bunga
H. tiliaceus juga terdapat keluarga dolichol yang hadir dengan tiga dominasi
rantai panjang C100-C110 tergantung pada jenis dan jaringan tanaman mangrove
(Tabel 2).
Distribusi jaringan bunga dan buah jenis H. tiliaceus dan M. candidum
termasuk ke dalam klasifikasi tipe II (Tabel 1) dimana dolichol dan polyprenol
hadir pada masing-masing jenis ini, sedangkan R. communis baik jaringan bunga
maupun buah termasuk kedalam klasifikasi tipe I dimana dolichol mendominasi
dibandingkan polyprenol. Hal ini merujuk kepada penelitian yang dilakukan
sebelumnya oleh Basyuni et al (2016).
Pembahasan
Teknik pemisahan polyisoprenoid dengan metode kromatografi sudah
dilakukan dan diteliti puluhan tahun sebelumnya. Banyak para peneliti mencoba
melakukannya dengan metode kromatografi dengan mengaplikasikan performa
tinggi kromatografi lapis tipis (high-performance thin layer chromatography
(HPTLC)). Sagami et al (1992) telah menemukan metode untuk memisahkan
dolichol dan polyprenol yang lebih mudah pada jaringan tanaman Ginkgo biloba
dengan sistem two-plate TLC (thin layer chromatography) yang mudah untuk
dilakukan. Dalam penelitian kali ini pun mencoba menemukan distribusi
polyisoprenoid pada jaringan bunga dan buah H. tiliaceus, M. candidum, dan
Berdasarkan strukturnya polyisoprenoid alkohol dibagi menjadi dolichol
dan polyprenol (Basyuni et al, 2016; Kania et al, 2011). Pada tiga jenis mangrove
ikutan R. communis, M. candidum, dan H. tiliaceus senyawa ini hadir dengan
rantai panjang yang berbeda-beda berdasarkan jaringannya dan terdistribusi di
masing-masing jenis. Rantai panjang polyisoprenoid telah terjadi di berbagai
jaringan tanaman (Swiezewska dan Danikiewicz, 2005). Senyawa ini juga hadir
pada jaringan bunga dan buah Philesia magellanica (Strzalka et al, 2009), bunga
tanaman karet (Tateyama et al, 1999), buah-buahan dan minuman (Muceniece et
al, 2016), bunga Magnolia (Swiezewska dan Danikiewicz, 2005), serta
terdistribusi luas di jaringan daun dan akar tanaman mangrove (Basyuni et al,
2016; 2017).
Tabel 2 merangkum distribusi dan kehadiran polyprenol dan dolichol yang
ada di tiga jenis mangrove ikutan H. tiliaceus, M. candidum, dan R. communis.
Senyawa dolichol dengan panjang rantai C55-C140, C70-C90, dan C75-C120
ditemukan di jaringan bunga masing-masing jenis dan dengan panjang rantai
karbon C60-C110, C70-C105, dan C80-C90 ditemukan di jaringan buah
masing-masing jenis. Dolichol ditemukan pada semua jenis baik pada jaringan bunga
maupun buah. Hal ini menandakan bahwa dolichol aktif berperan dan terdistribusi
luas di tanaman mangrove. Sama halnya dengan temuan Basyuni et al (2016)
bahwa dolichol mendominasi di jaringan daun dan akar sembilan jenis mangrove
meskipun beberapa famili polyprenol juga hadir. Walaupun faktanya senyawa
dolichol terutama hadir dan melimpah pada jaringan hewan, ragi, jamur, dan
uniseluler eukariot (Rip et al, 1985). Namun temuan ini berbeda dengan temuan
dolichol melainkan hanya rantai panjang polyprenol (C80-C120). Polyprenol juga
mendominasi pada bunga Magnolia dengan dominasi rantai panjang C55
(Swiezewska dan Danikiewicz, 2005) serta bunga dan buah P. magellanica
dengan dominasi rantai panjang C80 dan C100 (Strzalka et al, 2009).
Panjang rantai karbon dolichol pada bunga H. tiliaceus (Gambar 2C) lebih
panjang dibanding rantai polyprenol, juga sama terjadi pada buahnya (Gambar
2E). Namun pada bunga dan buah M. candidum (Gambar 2B dan 2E) perbedaan
panjang rantai karbon dolichol dan polyprenol tidak jauh berbeda. Pengamatan ini
menunjukkan bahwa jalur biosintesis rantai pendek polyprenol, menengah
polyprenol, rantai panjang polyprenol dan dolichol diatur secara berbeda di
kerajaan tanaman termasuk tanaman mangrove (Basyuni et al, 2016).
Senyawa polyprenol dengan panjang rantai karbon C55-C85 dan C75-C90
ditemukan pada masing-masing jaringan bunga M. candidum dan H. tiliaceus.
Sedangkan dengan panjang rantai karbon C60-C80 dan C70-C100 ditemukan
pada jaringan buahnya. Kehadiran polyprenol pada jaringan bunga telah diteliti
sebelumnya oleh Tateyama (1999) bahwa terdapat rantai polyprenol (C80-C120)
pada bunga H. brasiliensis. Sama halnya, polyprenol juga ditemukan di jaringan
bunga dan buah P. magellanica (Strzalka et al, 2009), serta pada buah-buahan dan
minuman, seperti teh, kopi, dan anggur (wine) (Muceniece et al, 2016). Dalam
dunia tanaman, polyprenol umumnya dominan pada daun (jaringan fotosintesis)
dengan rasio melimpah dibandingkan dolichol (Swiezewska dan Danikiewicz,
2005) namun juga sering dijumpai hadir pada jaringan bunga dan buah tanaman
Dari Tabel 2 dan Gambar 2A dan 2D terlihat bahwa pada jaringan bunga
dan buah R. communis (dengan panjang rantai karbon C75-C120 dan C80-C90)
tidak terdapat senyawa polyprenol. Pola distribusi ini berbanding terbalik dengan
studi kasus pada jaringan bunga dan buah P. magellanica (Strzalka et al, 2009)
dan pada bunga Magnolia (Swiezewska dan Danikiewicz, 2005) dimana hanya
senyawa polyprenol ditemukan. Kejadian ini dapat diduga disebabkan oleh faktor
lingkungan, seperti yang dinyatakan oleh Surmacz et al (2011) bahwa akumulasi
berlebihan polyprenol/dolichol biasanya dipertimbangkan sebagai indikator dari
asosiasi kandungan lipid dengan kondisi stress/cekaman tertentu, namun
walaupun begitu mekanisme kesangkutpautan ini masih belum jelas.
Dari hasil kromatografi di atas (Gambar 1 dan Tabel 2) menggambarkan
bahwa dolichol dan polyprenol di temukan pada bunga dan buah M. candidum
dan H. tiliaceus dengan perbedaan panjang rantai yang beragam, sedangkan pada
bunga dan buah R. communis hanya ditemukan senyawa dolichol saja. Perbedaan
panjang rantai polyisoprenoid pun dapat berbeda pada tiap-tiap jaringan. Telah
disarankan oleh Tateyama (1999) bahwasanya rantai panjang dolichol bervariasi
dari jaringan ke jaringan bahkan untuk spesies yang sama, dan muncul untuk
membentuk dominasi keluarga yang berbeda dengan spesies molekul. Polyprenol
juga terjadi sebagai satu atau dua famili polyprenol tergantung pada jenis tanaman
dan jaringan.
Senyawa polyisoprenoid tersebar luas dalam dunia tanaman dengan
kehadiran dan distribusi yang beragam. Dolichol dan polyprenol juga diketahui
sebagai penanda penuaan (umur) dan mungkin juga dapat dipertimbangkan
jaringan diketahui memiliki fungsi masing-masing walaupun fungsinya pada
dunia tanaman masih belum jelas (Basyuni et al, 2016). Namun sangat
disayangkan sekali adalah bahwa analisis dolichol dan polyprenol pada jaringan
bunga dan buah tanaman mangrove belum pernah tercatat pada temuan-temuan
sebelumnya.
Perbedaan panjang rantai polyisoprenoid yang terjadi pada R. communis,
M. candidum, dan H. tiliaceus (Tabel 2 dan Gambar 2) diatas dapat diakibatkan
oleh beberapa faktor, diantaranya adalah peningkatan umur (penuaan jaringan)
(Swiezewska et al, 2005), cekaman salinitas (Oku et al, 2003), perbedaan jaringan
(Tateyama et al, 1999; Surmacz dan Swiezewska, 2011) dan cahaya (Swiezewska
et al, 2005; Basyuni et al, 2012).
Basyuni et al (2016) telah membagi distribusi dolichol dan polyprenol
yang diklasifikasikan menjadi 3 tipe (I, II, dan III). Pada tipe I adalah dolichol
mendominsi dibanding polyprenol (lebih dari 90%) yang terdeteksi pada jaringan
bunga dan buah R. communis (Tabel 1) dengan kehadiran dolichol mencapai
100% pada kedua jaringan tersebut. Dan tipe II adalah senyawa dolichol dan
polyprenol sama-sama hadir yang terdeteksi pada jaringan bunga dan buah
M. candidum dan H. tiliaceus. Polyprenol berjumlah 26,2% dan dolichol 73.8%
pada jaringan bunga M. candidum dan pada buahnya jumlah polyprenol adalah
39,0% dan dolichol 61,0%. Sedangkan pada jaringan bunga H. tiliaceus,
polyprenol berjumlah 17,3% dan dolichol sebesar 82,7% dan pada buahnya
jumlah polyprenol adalah 47, 4% dan dolichol 52,6%. Namun dalam studi ini,
tidak terdapat tipe III karena tidak ada satupun jaringan yang menunjukkan
Dari Tabel 2 dapat diakumulasikan bahwa dolichol mendominasi di setiap
jaringan bunga dan buah tiga jenis mangrove ikutan. Dolichol bertindak sebagai
lipid pembawa gula dalam biosintesis N-glikoprotein. Oleh karena itu terjadinya
jumlah dolichol yang lebih besar bahkan di jaringan bunga dan buah tanaman
mangrove mengindikasikan bahwa polyprenol reduktase mungkin aktif di bunga
dan buah mangrove (Cantagrel et al, 2010).
Analisis polyisoprenoid di bunga dan buah tanaman mangrove ini
menunjukkan bahwa komponen utama polyisoprenoid bukan senyawa polyprenol
tetapi dolichol. Biasanya dalam dunia tanaman (terutama jaringan daun)
polyprenol terdeteksi melimpah konsentrasinya dibanding dolichol (Tateyama
et al, 1999). Dan sebaliknya di dunia hewan (terutama jaringan hati) dolichol
adalah polyisoprenoid utama dan hanya beberapa senyawa polyprenol ditemukan
(Sagami et al, 1992).
Oleh karena itu, terjadinya jumlah dan konsentrasi dolichol yang lebih
besar daripada senyawa polyprenol di jaringan bunga dan buah tiga jenis
mangrove ikutan menyarankan bahwa polyprenol reduktase yang berfungsi
mengkatalisis reduksi polyprenol menjadi dolichol mungkin aktif di jaringan
bunga dan buah mangrove. Walaupun demikian, belum terdapat banyak penelitian
mengenai senyawa polyisoprenoid pada jaringan bunga dan buah tanaman
mangrove. Oleh sebab itu kemungkinan ini masih belum cukup kuat dibandingkan
dengan temuan-temuan mengenai polyisoprenoid di daun dan akar tanaman
KESIMPULAN DAN SARAN
Distribusi dolichol dan polyprenol ditemukan pada tanaman mangrove
bervariasi tergantung pada jaringannya. Hal ini terjadi pada jaringan bunga dan
buah tiga jenis mangrove ikutan H. tiliaceus, M. candidum, dan R. communis.
Dolichol dengan panjang rantai C55-C140, C70-C90, dan C75-C120 ditemukan
pada jaringan bunga dan panjang rantai C60-C8110, C70-C105, dan C80-C90
ditemukan pada jaringan buah. Sedangkan senyawa polyprenol dengan panjang
rantai C55-C85 dan C75-C90 ditemukan pada jaringan bunga M. candidum dan
H. tiliaceus dan panjang rantai C60-C80 dan C70-C100 ditemukan pada jaringan
buahnya. Adapun pada jenis R. communis tidak ditemukan satu pun keluarga
senyawa polyprenol pada jaringan bunga maupun buahnya.
Distribusi dolichol dan polyprenol pada jaringan bunga dan buah tiga jenis
mangrove ikutan H. tiliaceus, M. candidum, dan R. communis dibagi menjadi dua
tipe (I dan II). Tipe I (dolichol mendominasi dibanding polyprenol) termasuk di
dalamnya adalah bunga dan buah R. communis. Tipe II (kedua dolichol dan
polyprenol sama-sama hadir) termasuk di dalamnya bunga dan buah M. candidum
dan H. tiliaceus.
Saran
Sebaiknya perlu diketahui perbedaan umur sampel bunga dan buah setiap
jenis pada penelitian ini. Dan perlu adanya perluasan studi mengenai kandungan