Summary We examined relationships between stem diame-ter, sapwood area, leaf area and transpiration in a 15-year-old mountain ash (Eucalyptus regnans F. Muell.) forest containing silver wattle (Acacia dealbata Link.) as a suppressed overstory species and mountain hickory (Acacia frigescens J.H. Willis) as an understory species. Stem diameter explained 93% of the variation in leaf area, 96% of the variation in sapwood area and 88% of the variation in mean daily spring transpiration in 19 mountain ash trees. In seven silver wattle trees, stem diameter explained 87% of the variation in sapwood area but was a poor predictor of the other variables. When transpiration measure-ments from individual trees were scaled up to a plot basis, using stem diameter values for 164 mountain ash trees and 124 silver wattle trees, mean daily spring transpiration rates of the two species were 2.3 and 0.6 mm day−1, respectively. The leaf area index of the plot was estimated directly by destructive sam-pling, and indirectly with an LAI-2000 plant canopy analyzer and by hemispherical canopy photography. All three methods gave similar results.
Keywords: Acacia dealbata, Acacia frigescens, Eucalyptus regnans, heat pulse method, leaf area index, mountain hickory, silver wattle.
Introduction
Water for the city of Melbourne is obtained from 155,000 ha of forested catchments in the central highlands of Victoria, Australia. Mountain ash (Eucalyptus regnans F. Muell.) forests cover just under half this area, but yield 80% of the streamflow because they grow on the higher rainfall sites.
In 1939, about 80% of the extensive old-growth mountain ash forest in Melbourne’s water supply catchments was killed in bushfires. It regenerated naturally with dense re-growth mountain ash forest. The widespread destruction and regenera-tion of the forests was followed by a prolonged decline in streamflows. In the three decades following the fires, mean annual streamflows from five catchments were 25% less than
the pre-1939 mean (Langford 1976). The reduction in water yield after the fire in each catchment was proportional to the percentage of mountain ash forest burnt (Langford 1976, Kuczera 1987). The maximum decline in mean annual stream-flow occurred 27 years after the fires and was equivalent to 6 mm for each 1% of catchment converted from old-growth to re-growth (Kuczera 1987).
There have been many studies of the Melbourne water supply area. Field experiments have examined stand-age-de-pendent rates of fog drip (O’Connell and O’Shaughnessy 1975), rainfall interception (Langford and O’Shaughnessy 1978) and evapotranspiration (Legge 1980, Dunn and Connor 1993, Jayasuriya et al. 1993). Fog drip was shown to be less than 1% of gross precipitation and did not vary significantly between re-growth and old-growth stands. Although Langford and O’Shaughnessy (1978) found that interception in 34-year-old re-growth mountain ash forest was lower than in a 160-year-old mature mountain ash forest, a recent, more comprehensive study has determined that interception peaks at 26% of gross rainfall at age 30 years and steadily declines to 17% by age 240 years (S. Haydon, Melbourne Water, personal communication). However, changes in interception explained only about 25% of the changes in streamflow resulting from conversion of old-growth forests to a re-growth state.
Although the causal mechanisms of the water yield response in the Melbourne water supply area have not been fully eluci-dated, it is now assumed that differences in evapotranspiration are the prime cause of the stand age--water yield relationship in mountain ash forests. Recent studies by Dunn and Connor (1993) and Jayasuriya et al. (1993) emphasize the role of changing leaf area and sapwood area in determining changes in forest evapotranspiration. However, no studies have pro-vided quantitative data linking leaf area, sapwood area and transpiration in mountain ash forests.
As a first step in elucidating the factors underlying the stand age--water yield relationship in mountain ash forests, we have examined relationships between stem diameter, sapwood area, leaf area and transpiration in a 15-year-old mountain ash forest
Relationships between stem diameter, sapwood area, leaf area and
transpiration in a young mountain ash forest
R. A. VERTESSY,
1,4R. G. BENYON,
2,4S. K. O’SULLIVAN
3,4and P. R. GRIBBEN
3,41 CSIRO Division of Water Resources, GPO Box 1666, Canberra, ACT 2601, Australia
2 Melbourne Water, Box 4342, Melbourne, Victoria 3001, Australia
3 Monash University, Wellington Road, Clayton, Victoria 3168, Australia
4 Cooperative Research Center for Catchment Hydrology, Wellington Road, Clayton, Victoria 3168, Australia
Received October 20, 1994
and obtained evidence that similar relationships could be de-rived for older stands. We determined leaf area by destructively sampling individual trees. Because such sampling is impracti-cal in older forests with large trees, we also evaluated two methods for indirectly measuring leaf area index and show that these can be used in older stands of mountain ash.
Materials and methods
Site description
The study site was located in the North Maroondah experimen-tal area (Figure 1), at an elevation of 890 m, with a mean annual rainfall of 1713 mm. Soils in the area are deep krasnozems of high permeability and high water-holding capacity (Langford and O’Shaughnessy 1977). The even-aged stand of mountain ash was regenerated from direct seeding after clear-cutting in 1978. As a result of natural thinning, the stocking density of mountain ash trees decreased from 8500 stems ha−1 in 1983 to 656 stems ha−1 at the time of the study in the spring of 1993. Silver wattle (Acacia dealbata Link.) is a subdominant canopy species, present at a stocking density of 496 stems ha−1. The understory is mostly mountain hickory (Acacia frigescens J.H. Willis), rising to a height of 3 m, and assorted ferns reaching about 1 m in height.
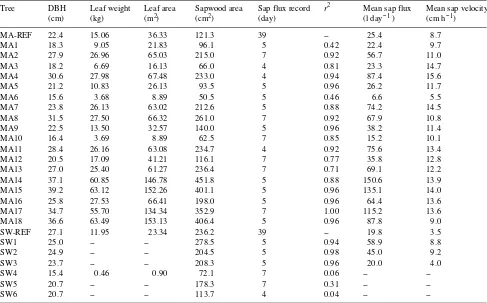
A 50 × 50 m plot was established, containing 164 mountain ash and 124 silver wattle trees, with mean heights of 28 and 24 m, respectively. The stem diameter at breast height over bark (DBH) of each tree was measured. Mean DBH values for the mountain ash and silver wattle trees were 20.4 and 15.6 cm, respectively. Minimum DBH values were 9.6 and 10.0 cm, and maximum DBH values were 39.2 and 27.1 cm, respectively. Nineteen mountain ash and seven silver wattle trees, spanning the range of DBH values in the plot, were selected for detailed examination. For each of these trees, we determined the mean daily sapflow velocity, mean daily transpiration rate, DBH,
height, total leaf weight, total leaf area and sapwood cross-sec-tional area (Table 1).
Transpiration measurements
Mean daily minimum and maximum temperatures for the period of record were 4.4 and 12.8 °C, respectively, slightly lower than the long-term October--November averages of 7.3 and 13.4 °C (Langford and O’Shaughnessy 1977). Total solar radiation varied between 3.1 and 25.1 MJ m−2 day−1, with a mean value of 13.9 MJ m−2 day−1. This mean daily value is similar to the local October--November average of 13.2 MJ m−2 day−1 (Langford and O’Shaughnessy 1977).
Over a 39-day period between September and November 1993, measurements of half-hourly transpiration were made on the sample trees by the heat pulse technique (Cohen et al. 1985, Green and Clothier 1988, Hatton and Vertessy 1990, Olbrich 1991, Dunn and Connor 1993). Half-hourly measure-ments of sapflow velocities were multiplied by sapwood cross-sectional area to compute transpiration flux in individual trees. Dunn and Connor (1993) validated this technique in young mountain ash trees and showed that no significant wound effect takes place after implanting sensor probes.
Six sapflow sensor units (Greenspan Technology, Warwick, Queensland) were deployed for a period of 39 days using a roaming sensor technique. One sensor unit was allocated to a reference mountain ash tree (MA-REF) and another to a refer-ence silver wattle tree (SW-REF) for the entire 39-day meas-urement period. The other four sensor units were moved from tree to tree every 4 to 7 days (Table 1). At all times, three roaming sensors were deployed in mountain ash trees and one in a silver wattle tree. Measurements of total daily transpira-tion obtained from the roaming sensors were regressed against those obtained with the reference sensors. The resulting regres-sion equations were used to predict total daily transpiration in the roaming sensor trees outside the period of measurement in
those trees. This enabled us to estimate total daily transpiration for each sample tree over the entire 39-day measurement period. No transpiration measurements were made in the un-derstory.
Fractions of wood, water and air in the sapwood were determined gravimetrically on one 5-mm diameter increment core from each tree at the end of the transpiration experiment. In the mountain ash trees, the wood fraction varied between 0.26 and 0.34 (mean = 0.29), whereas the water fraction varied between 0.55 and 0.67 (mean = 0.60). In the silver wattle trees, the wood fraction varied between 0.31 and 0.37 (mean = 0.32), and the water fraction varied between 0.28 and 0.49 (mean = 0.36). Hence, the silver wattle trees had a much larger air fraction in the sapwood than the mountain ash trees.
Nondestructive measurement of leaf area index
An LAI-2000 plant canopy analyzer (PCA) (Li-Cor Inc., Lin-coln, NE) and hemispherical canopy photographs were used to estimate the leaf area index (LAI) of the 50 × 50 m plot. Both methods combine the leaf area of the overstory and understory species to give a total LAI value for the plot.
Leaf area index was computed from the PCA measurements using a canopy gap fraction model (Welles and Norman 1991). We deployed two PCAs; one device was left in a clearing (1.5 km from the site) to record light conditions automatically every 15 s, whereas the other PCA was used manually beneath the canopy in the experimental plot. All PCA measurements were made over a 360° field of view, under cloudy (diffuse
radiation dominated) conditions or at dawn or dusk when there was no direct sunlight on the canopy. We only used readings from the four inner rings of the sensor, thereby confining the PCA’s field of view to a radius of 41 m.
Three sampling protocols (random, transect and fixed posi-tion) were used in the application of the PCA to the experimen-tal plot. In the random sampling protocol, the PCA operator moved around the plot making readings at random locations. These measurements were repeated on four different occasions to include a variety of sky conditions. In the transect sampling protocol, the PCA operator made measurements at roughly 1-m intervals along two 50-m transects laid out across the plot and two 70-m transects running along the diagonals of the plot. One of the 50-m transects was measured twice and the other 50-m transect was measured three times. In the fixed position sampling protocol, the PCA operator made measurements at six fixed points in the plot on six occasions.
Multiple hemispherical photographs were taken at the same six points within the plot. We used Ilford pan F (50 ASA) film in a NIKON FG camera, fitted with a Nikkor 8 mm fisheye lens. All photographs were taken under cloudy conditions to ensure even back-lighting. Each negative was digitized to provide a 512 × 512 8-bit image. The images were analyzed to determine the percent canopy gap as a function of zenith and azimuth angles. The PISCES program (Jupp et al. 1980) was used to compute the generalized foliage area index (GFAI), which is analogous to LAI, from the canopy gap fraction data. Although the scanned images extended to a zenith angle of
Table 1. Parameter values for all sample trees. The value of r2 is relative to mean daily transpiration in reference trees MA-REF and SW-REF.
Tree DBH Leaf weight Leaf area Sapwood area Sap flux record r2 Mean sap flux Mean sap velocity
(cm) (kg) (m2) (cm2) (day) (l day−1 ) (cm h−1)
MA-REF 22.4 15.06 36.33 121.3 39 -- 25.4 8.7
MA1 18.3 9.05 21.83 96.1 5 0.42 22.4 9.7
MA2 27.9 26.96 65.03 215.0 7 0.92 56.7 11.0
MA3 18.2 6.69 16.13 66.0 4 0.81 23.3 14.7
MA4 30.6 27.98 67.48 233.0 4 0.94 87.4 15.6
MA5 21.2 10.83 26.13 93.5 5 0.96 26.2 11.7
MA6 15.6 3.68 8.89 50.5 5 0.46 6.6 5.5
MA7 23.8 26.13 63.02 212.6 5 0.88 74.2 14.5
MA8 31.5 27.50 66.32 261.0 7 0.92 67.9 10.8
MA9 22.5 13.50 32.57 140.0 5 0.96 38.2 11.4
MA10 16.4 3.69 8.89 62.5 7 0.85 15.2 10.1
MA11 28.4 26.16 63.08 234.7 4 0.92 75.6 13.4
MA12 20.5 17.09 41.21 116.1 7 0.77 35.8 12.8
MA13 27.0 25.40 61.27 236.4 7 0.71 69.1 12.2
MA14 37.1 60.85 146.78 451.8 5 0.88 150.6 13.9
MA15 39.2 63.12 152.26 401.1 5 0.96 135.1 14.0
MA16 25.8 27.53 66.41 198.0 5 0.96 64.4 13.6
MA17 34.7 55.70 134.34 352.9 7 1.00 115.2 13.6
MA18 36.6 63.49 153.13 406.4 5 0.96 87.8 9.0
SW-REF 27.1 11.95 23.34 236.2 39 -- 19.8 3.5
SW1 25.0 -- -- 278.5 5 0.94 58.9 8.8
SW2 24.9 -- -- 204.5 5 0.98 45.0 9.2
SW3 23.7 -- -- 208.3 5 0.96 20.0 4.0
SW4 15.4 0.46 0.90 72.1 7 0.06 --
--SW5 20.7 -- -- 178.3 7 0.31 --
--84.3°, the foliage was so dense that measurements beyond 65° had little influence on the analysis; hence, the hemispherical photographic and PCA methods had similar fields of view. Destructive measurement of leaf area index
After completion of the transpiration and PCA measurements, nineteen mountain ash trees and two silver wattle trees were felled. The total leaf mass from each tree was harvested and weighed. The height of each tree was measured and stem discs were cut at breast height (1.3 m above ground level) for measurements of sapwood cross-sectional area.
Five circular subplots, each with a radius of 5 m, were established within the 50 × 50 m plot, covering a total area of 392.75 m2, or 15.7% of the experimental plot. Leaves were stripped and weighed from all mountain hickory trees located within each subplot.
Leaf subsamples from each of the three tree species were weighed, then immediately fed through a planimeter to obtain a leaf area to weight ratio for each species. It was difficult to obtain reliable leaf area estimates for the silver wattle trees because the fragile pinnate leaves were easily dislodged when the trees hit the ground and were awkward to feed through the planimeter. Leaf area estimates were therefore obtained for only two of the seven sample silver wattle trees, including the largest and one of the smallest trees in the plot.
Sapwood area measurements
Sapwood thickness and sapwood cross-sectional area of each sample tree were determined from measurements on several 5-mm diameter increment cores, taken at breast height in each of the sample trees before felling. The sapwood width meas-urements were used to determine appropriate depths for im-planting heat pulse sensor probes. After felling, more accurate measurements of sapwood area were made on wood discs, cut at breast height. Measurements of bark thickness, sapwood thickness and heartwood diameter at up to 15 points around each wood disc were used to compute sapwood area. These determinations were used in the transpiration calculations.
In the silver wattle trees, the heartwood and sapwood were clearly distinguishable on the basis of color. To identify sap-wood in the mountain ash increment cores, it was necessary to hold each core up against a bright light. The open vessels of the sapwood were clearly visible as pin-points of light, whereas the blocked vessels of the heartwood did not transmit light. The heartwood in the mountain ash discs could be easily distinguished after a day or two of exposure to air because the heartwood became brown, whereas the sapwood remained straw colored. In mountain ash trees, the no-flow boundary detected with the sapflow sensor corresponded to the inde-pendent estimates of the sapwood--heartwood transition.
Results
Direct leaf area index estimates
Leaf area estimates for 19 mountain ash trees were based on the wet weight of leaves collected from each tree, multiplied
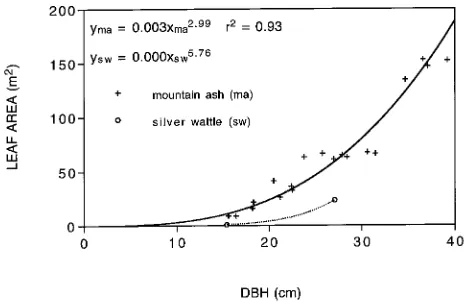
by the measured leaf area/leaf wet weight ratio of 2.412 m2 kg−1 (Table 1). To scale up individual tree leaf areas to a plot LAI value, a regression equation was developed that related leaf area to stem diameter (Figure 2). Because a linear fit gave an undesirable y-intercept value, we applied a power function that passed through the origin and hence reflected the growth process. The r2 value of 0.93 (n = 19) indicated a strong association between stem diameter and leaf area. This regres-sion equation was evaluated for all 164 mountain ash trees in the plot, resulting in a total mountain ash leaf area of 5342 m2, or an LAI of 2.1.
Leaf wet weights were determined for two silver wattle trees (Table 1), and these were multiplied by the measured leaf area/leaf wet weight ratio of 1.953 m2 kg−1 to determine leaf area for both trees. Silver wattle leaf area was regressed against stem diameter, based on the regression equation used for the mountain ash trees. Although the power function was fitted to only two data points, we believe the fitting was valid because the function passed through the origin, and data points for the largest tree and one of the smallest trees in the plot were included. The small diameter silver wattle trees supported little leaf area and were beginning to die out as a result of over-top-ping by the mountain ash canopy. The large silver wattle trees occurred in canopy gaps and were therefore less affected by competition with mountain ash trees. The regression equation relating silver wattle leaf area to stem diameter was used to estimate total leaf area for all 124 silver wattle trees in the 50 × 50 m plot. This yielded an estimate of 290.23 m2 of leaf area, equivalent to an LAI of 0.1.
The leaf area/leaf wet weight ratio for the mountain hickory was 3.646 m2 kg−1. The total leaf mass harvested from five circular plots was 194.38 kg, equivalent to a total leaf area of 708.72 m2, or an LAI of 1.8.
Indirect leaf area index estimates
Table 2 lists means of all of the PCA-based LAI estimates made for the 50 × 50 m plot using the random, transect and fixed position sampling protocols. For each sampling protocol, multiple determinations were made under varying irradiances
to obtain an average value of LAI. Mean LAI values for the random, transect and fixed position sampling protocols were 4.1, 4.4 and 4.1, respectively, and the mean of all PCA readings obtained from all sampling protocols was 4.2. None of the sampling protocols was more accurate or reproducible than the others.
Fixed-position LAI estimates based on the PCA and photo-graphic methods are compared in Table 3. For the PCA method, we list the mean and range of LAI values measured for each plot. For the photographic method, we list the LAI determination and error margin for a single photograph from each plot. With the exception of Plot 3, the individual fixed-po-sition LAI estimates obtained by the PCA and photographic methods differed by 0.5 or less. The mean photograph-based LAI estimate for the plot was 4.4, which is almost the same as the mean fixed-position estimate of 4.1 obtained with the PCA. Although we applied the PCA under ideal light conditions, LAI determinations for some plots varied by 1.0 or more between samplings. Error bounds for the photograph-based
LAI estimates were large for individual photographs (up to 2.1 units) but small for the lumped plot value (only 0.3).
Sapwood area estimates
Sapwood areas in the mountain ash trees ranged between 50.5 and 451.8 cm2 and averaged 207.8 cm2, whereas silver wattle sapwood areas ranged between 72.1 and 278.5 cm2 and aver-aged 184.5 cm2 (Table 1). Sapwood areas were reliably pre-dicted from stem diameter measurements in both species, yielding r2 values of 0.96 and 0.87, respectively (Figure 3). Regression equations for the two species were similar, though statistically different.
The sapwood areas for all mountain ash and silver wattle trees in the plot were estimated from regression equations for each species. Total sapwood area for the 164 mountain ash trees in the plot was 8.04 m2 ha−1. Total sapwood area for the 124 silver wattle trees in the plot was 4.16 m2 ha−1, or 34.1% of the combined overstory sapwood area. There was a strong relationship between sapwood area and leaf area in the moun-tain ash trees (r2 = 0.96, n = 19) (Figure 4). There were insufficient leaf area measurements for the silver wattle trees to make a similar comparison.
Figure 3. Stem diameter (DBH) versus sapwood area for the sampled mountain ash and silver wattle trees.
Figure 4. Sapwood area versus leaf area for the sampled mountain ash trees.
Table 2. Various LAI estimates for the 50 × 50 m plot obtained using the PCA; n = number of readings per sample.
Sampling protocol Run identifier n LAI
Random 1 228 3.6
Random 2 200 4.5
Random 3 60 4.5
Random 4 200 3.8
Transect (east) 5 50 4.4
Transect (east) 6 50 4.2
Transect (north) 7 55 4.4
Transect (north) 8 55 5.0
Transect (north) 9 55 4.5
Transect (both diagonals) 10 130 3.9
Fixed position 11 6 3.8
Fixed position 12 6 4.3
Fixed position 13 6 4.1
Fixed position 14 6 4.4
Fixed position 15 6 4.0
Fixed position 16 6 3.8
Total/mean 1--16 1119 4.2
Table 3. Fixed position LAI estimates for the six circular plots ob-tained using the PCA and hemispherical canopy photography.
Plot LAI Range LAI Error
no. (PCA) (PCA) (photo) (photo)
1 3.2 2.9--3.5 3.7 ± 0.8
2 4.1 3.6--4.7 4.0 ± 0.6
3 4.5 4.1--4.7 6.1 ± 1.3
4 4.2 3.8--4.8 3.7 ± 1.4
5 4.3 4.0--4.6 4.2 ± 1.0
6 4.1 3.2--4.6 4.4 ± 2.1
Transpiration estimates in the sample trees
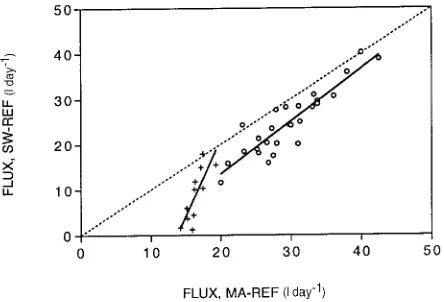
Over the 39-day measurement period, transpiration in tree MA-REF varied between 9 and 43 l day−1 , and the mean daily value was 25 l day−1 (Figure 5). In tree SW-REF, transpiration varied between 1 and 40 l day−1 , with a mean daily value of 20 l day−1 (Figure 5). Tree SW-REF had a larger diameter than tree MA-REF (27.1 versus 22.4 cm) and greater sapwood area (236.2 versus 121.3 cm2), but a lower leaf area (23.34 versus 36.33 m2). Despite these differences, daily transpiration rates in the two trees were similar, except that when transpiration in tree MA-REF dropped below 20 l day−1, transpiration in tree SW-REF declined sharply (Figure 6). On these days, radiation was usually below average, and wind speeds were significantly lower than normal. The silver wattle canopy was on average about 4 m below that of the mountain ash and would have been poorly ventilated on days of low wind speed, so the results suggest uncoupling of the silver wattle canopy from the atmos-pheric boundary layer during these times.
For periods of concurrent record, daily transpiration values for the other 18 mountain ash trees were regressed against daily transpiration values for tree MA-REF. Because the statis-tical association between daily transpiration totals in the dif-ferent mountain ash trees was strong, with r2 values exceeding 0.9 in over half the cases and exceeding 0.7 in all but two cases
(Table 1), we were able to estimate daily transpiration in the other 18 mountain ash trees for the entire 39-day period. The mean daily transpiration for these trees varied between 6.6 (MA6) and 150.6 l day−1 (MA14), with a mean of 62 l day−1 (Table 1).
Daily transpiration values for tree SW-REF were regressed against daily transpiration values for the other six silver wattle trees sampled during periods of concurrent record. Although trees SW1, SW2 and SW4 displayed daily transpiration behav-ior similar to tree SW-REF (r2 = 0.9), trees SW3, SW5 and SW6 did not. This difference may be attributable to the rapid thinning of the silver wattle trees in the plot as the mountain ash canopy began to over-top the silver wattle trees. With the exception of tree SW4, all of the positive results came from the larger silver wattle trees. Consequently, robust transpiration totals for the 39-day sampling period were derived for only four of the seven silver wattle trees sampled. For these four trees, mean daily transpiration varied between 19.8 (SW-REF) and 58.9 l day−1 (SW1), with a mean of 35.9 l day−1 (Table 1). There was considerable variation in mean daily sapflow velocity among the 19 mountain ash trees sampled, with values ranging between 5.5 (MA6) and 15.6 cm h−1 (MA4), and averaging 11.9 cm h−1 (Table 1). There was no systematic relationship between stem diameter and mean daily sapflow velocity in the mountain ash trees. Mean daily sapflow veloci-ties for the four silver wattle trees varied between 3.5 (SW-REF) and 9.2 cm h−1 (SW2), and averaged 6.4 cm h−1 (Table 1). This mean daily value was just over half that served for the mountain ash trees, but almost twice that ob-served for mature A. frigescens (Dunn and Connor 1993).
Mean daily transpiration in the mountain ash trees was strongly related to both DBH (r2 = 0.88, n = 19) and leaf area (r2 = 0.91, n = 19) (Figures 7 and 8). The relationship between leaf area and transpiration was almost linear (cf. Running and Coughlan 1988, Hatton et al. 1992, Band et al. 1993, Vertessy et al. 1993). However, the larger trees appeared to have a slightly lower leaf area efficiency than the smaller trees, even though they experienced higher radiation and wind speeds, and lower humidity than the smaller trees. This finding is consis-tent with observations by Yoder et al. (1994) that large trees tend to experience plumbing problems and consequent sto-matal limitations more than small trees.
Figure 5. Daily transpiration in the two reference trees (MA-REF and SW-REF) over the 39-day sampling period.
Plot transpiration
The mean daily transpiration for all 164 mountain ash trees in the experimental plot was calculated to be 5740 l day−1, or 2.3 mm day−1 when expressed on an area basis. Although we could not obtain a statistically significant relationship between mean daily transpiration and DBH in the silver wattle trees, by multiplying the total sapwood area (1.04 m2) by the mean daily sapflow velocity (6.4 cm h−1), we obtained a mean daily transpiration estimate of 1597 l day−1, or 0.6 mm day−1. The estimate of plot transpiration for the silver wattle trees is less robust than the estimate for mountain ash, because the estimate of mean sapflow velocity was based on only four trees and these were amongst the largest silver wattle trees in the plot. However, we observed no systematic relationship between DBH and sapflow velocity in the mountain ash trees. For the entire 39-day measurement period, the mean daily transpira-tion rate for the mountain ash and silver wattle trees combined was 2.9 mm day−1 .
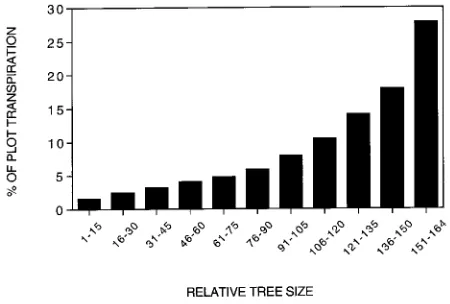
Transpiration among the mountain ash trees in the experi-mental plot was highly skewed. When the relative mean daily transpiration (expressed as a percentage of the plot total) of mountain ash trees was ranked on the basis of DBH, the 14
largest mountain ash trees in the plot (occupying 22% of the basal area) accounted for 28% of the mountain ash transpira-tion, whereas the 90 smallest mountain ash trees in the plot (also occupying 22% of the basal area) were responsible for only 22% of the mountain ash transpiration (Figure 9). This finding emphasizes the need to sample a spectrum of tree sizes when attempting to estimate plot water use by the heat pulse method.
Discussion
In the mountain ash, we found strong statistical associations between stem diameters and all other stand parameters ana-lyzed. Stem diameter measurements explained 93% of the variation in leaf area, 96% of the variation in sapwood area and 88% of the variation in mean daily transpiration. Because stem diameters are easy to measure, it is possible to make reliable assessments of related stand parameters in large plots. We did not test whether the same statistical associations hold for older mountain ash stands.
A strong association between sapwood area and leaf area has been noted in other eucalypts (Brack et al. 1985, Moore 1993, Hatton et al. 1995). Although Hatton and Wu (1995) cautioned that leaf area can decline quickly in response to drought, whereas sapwood area remains stable, this effect is probably not significant in mountain ash trees because they are rarely drought stressed. However, it is not known whether leaf area varies temporally in these forests in response to seasonal changes in irradiance or temperature, or both.
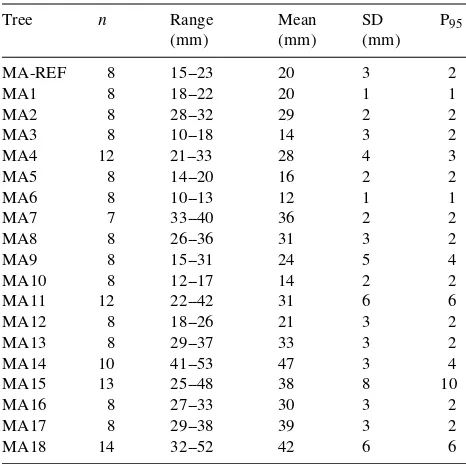
When estimating tree transpiration using the heat pulse technique, accurate measurement of sapwood area is essential. Even in young mountain ash trees that appear symmetrical, we found considerable variation in the width of the sapwood at breast height within individual trees (Table 4). Thus, a single sample from MA15 may have produced an estimated sapwood width anywhere between 25 and 48 mm, whereas the true mean was 38 mm. In most of the 19 mountain ash trees analyzed, at least two or three core samples would be needed to give an estimate of the mean sapwood width within 5 mm
Figure 8. Leaf area versus mean daily transpiration for the sampled mountain ash trees.
Figure 9. Relative transpiration by mountain ash trees of different sizes in the 50 × 50 m plot. Class 151-164 contains the 14 largest trees in the plot.
of the true mean with 95% confidence (Table 4). Our stem disc analyses indicated that sapwood width was more variable in the large-diameter trees than in the small-diameter trees. Be-cause of slight buttressing, wider sapwood occurred at the buttresses, whereas the narrower sapwood occurred between the buttresses.
Dunn and Connor (1993) reported sapwood area values of 6.74, 6.09, 4.23 and 4.04 m2 ha−1 for 50-, 90-, 150- and 230-year-old mountain ash stands, respectively. Our plot-based estimate of mountain ash sapwood area of 8.04 m2 ha−1 therefore lends support to Dunn and Connor’s (1993) hypothe-sis that sapwood area declines with age in mountain ash for-ests. Our data also support Dunn and Connor’s (1993) contention that mean daily sapflow velocity in mountain ash trees appears to be independent of stand age. Over a spring--summer--autumn sampling period, they estimated mean daily sapflow velocities of 11.5, 11.4, 9.9 and 11.8 cm h−1 for 50-, 90-, 150- and 230-year-old mountain ash stands, respectively. In our 15-year-old stand, we estimated a mean daily sapflow velocity of 11.9 cm h−1. This implies that spring transpiration in our stand was almost twice that of a 230-year-old stand.
The strong relationship between stem diameter and transpi-ration in mountain ash permits calculations to be made on the implications of forest thinning for catchment water balance. We estimate that a 50% basal area reduction would reduce mean daily (spring) transpiration by 58% if the biggest trees were felled, or by 42% if the smaller trees were felled. Leaf area (and hence rainfall interception) would be reduced by 61 or 39%, depending on whether the biggest or smallest moun-tain ash trees were removed (Figure 2). These data imply that naturally occurring water yield deficits in re-growth mountain
ash forests could be significantly off-set by selectively felling the largest trees every few years after re-growth commences. However, recent forest thinning experiments in mountain ash forests suggest that instantaneous reductions in transpiration and interception would not be converted entirely into stream-flow gains (Jayasuriya et al. 1993).
We note that our plot transpiration estimates are deficient in two key respects if they are to be used in water balance calculations. First, they are only valid for spring conditions. To estimate annual transpiration from stem diameter measure-ments, it would be necessary to repeat sapflow measurements in the trees at different times of the year. Second, we did not measure the transpiration rate of the mountain hickory under-story, though we have reason to believe that this is small, despite its significant contribution to plot leaf area. In a mature (230-year-old) mountain ash forest with a well developed mountain hickory understory and low density mountain ash overstory (72 stems ha−1), Dunn and Connor (1993) estimated that the understory was responsible for about 27% of the total forest transpiration during summer. However, in a soil water balance study carried out in denser 38-year-old re-growth forest, Langford and O’Shaughnessy (1979) concluded that the woody understory contributed little to soil water depletion over a summer period. Although 45% of the total plot leaf area was associated with the understory at our site, we suspect that transpiration from this layer would be low because of intense shading from the overstory trees and minimal turbulent trans-fer between the understory and overstory layers.
Based on destructive sampling, the leaf area indices of the mountain ash, silver wattle and mountain hickory vegetation were 2.1, 0.1 and 1.8, respectively, yielding a total plot LAI value of 4.0. The PCA and photographic methods yielded values of 4.2 and 4.4, respectively, thus overestimating LAI by 5 to 10%. This difference is small and could be attributable to errors in any of the estimation techniques. However, the degree and direction of difference are consistent with other studies that have compared direct and indirect estimates of LAI in forests; these studies have concluded that slight overestimation arises in indirect methods because tree stems and branches account for between 4 and 12% of light interception in forests (Neumann et al. 1989, Chason et al. 1991, Lang et al. 1991).
Leaf area is difficult to determine directly in older mountain ash trees because of their great size (50--80 m high), thus indirect LAI estimation techniques, such as the PCA, are required. The PCA is simple and rapid to apply and does not require knowledge of the light extinction coefficient for the stand, as is the case with quantum sensors (Pierce and Running 1988, Rich et al. 1993). However, the estimates of LAI made with the PCA varied with sampling protocol and irradiance at the time of measurement (Tables 2 and 3). Also, we do not know how well the PCA performs in older stands where canopy structure differs from that in young stands.
Acknowledgments
This study was funded by the Cooperative Research Center for Catch-ment Hydrology. Melbourne Water and the Victorian DepartCatch-ment of Conservation and Natural Resources assisted with several aspects of Table 4. Variability of sapwood widths within individual mountain ash
trees; n = number of samples, SD = standard deviation, P95 = number
of samples needed to estimate mean sapwood thickness within 95% confidence limits of 5 mm.
the field experiment. We thank Kim Whitford of the West Australian Department of Conservation and Land Management for loaning us photographic equipment and Dr. Barry Harper of Wollongong Univer-sity for analyzing the canopy photographs.
References
Band, L.E., P. Patterson, R. Nemani and S.W. Running. 1993. Forest ecosystem processes at the watershed scale: incorporating hillslope hydrology. Agric. For. Meteorol. 63:93--126.
Brack, C.L., M.P. Dawson and A.M. Gill. 1985. Bark, leaf and sap-wood dimensions in Eucalyptus. Aust. For. Res. 15:1--7.
Chason, J.W., D.D. Baldocchi and M.A. Huston. 1991. A comparison of direct and indirect methods for estimating canopy leaf area. Agric. For. Meteorol. 57:107--128.
Cohen, Y., T.A. Black and F.M. Kelliher. 1985. Determination of sap flow in Douglas-fir trees using the heat pulse technique. Can. J. For. Res. 15:422--428.
Dunn, G.M. and D.J. Connor. 1993. An analysis of sap flow in mountain ash (Eucalyptus regnans) forests of different age. Tree Physiol. 13:321--336.
Green, S.R. and B.W. Clothier. 1988. Water use of kiwifruit vines and apple trees by the heat-pulse technique. J. Exp. Bot. 39:115--123. Hatton, T.J. and R.A. Vertessy. 1990. Improved sampling procedures
for the robust estimation of sapflow in Pinus radiata by the heat pulse method. Hydrol. Proc. 4:289--298.
Hatton, T.J., J. Walker, W. Dawes and F.X. Dunin. 1992. Simulations of hydroecological responses to elevated CO2 at the catchment scale. Aust. J. Bot. 40:679--696.
Hatton, T.J., S.J. Moore and P.H. Reece. 1995. Estimating stand transpiration in a Eucalyptus populnea woodland with the heat pulse method: measurement errors and sampling strategies. Tree Physiol. 15:219--227.
Hatton, T.J. and H. Wu. 1995. A scaling theory to extrapolate individ-ual tree water use to stand water use. Hydrol. Proc. In press. Jayasuriya, M.D.A., G. Dunn, R. Benyon and P.J. O’Shaughnessy.
1993. Some factors affecting water yield from mountain ash ( Euca-lyptus regnans) dominated forests in south-east Australia. J. Hydrol. 150:345--367.
Jupp, D.L.B., M.C. Anderson, G.M. Adomeit and S.J. Witts. 1980. PISCES----A computer program for analysing hemispherical can-opy photographs. CSIRO, Australia Division of Land Use Re-search, Technical Memorandum 80/23, 22 p.
Kuczera, G.A. 1987. Prediction of water yield reductions following a bushfire in ash-mixed species eucalypt forest. J. Hydrol. 94:215--236.
Lang, A.R.G., R.E. McMurtrie and M.L. Benson. 1991. Validity of surface area indices of Pinus radiata estimated from transmittance of the sun’s beam. Agric. For. Meteorol. 57:157--170.
Langford, K.J. 1976. Change in yield of water following a bushfire in a forest of Eucalyptus regnans. J. Hydrol. 29:87--114.
Langford, K.J. and P.J. O’Shaughnessy. 1977. First progress report, North Maroondah. Melbourne and Metropolitan Board of Works, Catchment Hydrology Research Report Number MMBW-W-0005, 340 p.
Langford, K.J. and P.J. O’Shaughnessy. 1978. A study of canopy interception in native forests and conifer plantations. Melbourne and Metropolitan Board of Works, Catchment Hydrology Research Report Number MMBW-W-0007, 88 p.
Langford, K.J. and P.J. O’Shaughnessy. 1979. Soil moisture depletion rates in 1939 regrowth mountain ash over a summer drying period. Melbourne and Metropolitan Board of Works, Catchment Hydrol-ogy Research Report Number MMBW-W-0008, 161 p.
Legge, N.J. 1980. Aspects of transpiration in mountain ash Eucalyptus regnans F. Muell. Ph.D. Thesis. Latrobe University, Victoria, Aus-tralia, 164 p.
Moore, S. 1993. Estimating stand transpiration in a Eucalyptus popul-nea woodland. Honors Thesis. Australian National University, Can-berra, Australia, 98 p.
Neumann, H.H., G. Den Hartog and R.H. Shaw. 1989. Leaf area measurements based on hemispheric photographs and leaf-litter collection in a deciduous forest during autumn leaf-fall. Agric. For. Meteorol. 45:325--345.
O’Connell, M.J. and P.J. O’Shaughnessy. 1975. The Wallaby Creek fog drip study. Melbourne and Metropolitan Board of Works, Catchment Hydrology Research Report Number MMBW-W-0004, 66 p.
Olbrich, B.W. 1991. The verification of the heat pulse velocity tech-nique for measuring sap flow in Eucalyptus grandis. Can. J. For. Res. 21:836--841.
Pierce, L.L. and S.W. Running. 1988. Rapid estimation of coniferous forest leaf area index using a portable integrating radiometer. Ecol-ogy 69:1762--1767.
Rich, P.M., D.B. Clark, D.A. Clark and S.F. Oberbauer. 1993. Long-term study of solar radiation regimes in a tropical wet forest using quantum sensors and hemispherical photography. Agric. For. Me-teorol. 65:107--127.
Running, S.W. and J.C. Coughlan. 1988. A general model of forest ecosystem processes for regional applications. I. Hydrologic bal-ance, canopy gas exchange and primary production processes. Ecol. Model. 42:125--154.
Vertessy, R.A., T.J. Hatton, P.J. O’Shaughnessy and M.D.A. Jayas-uriya. 1993. Predicting water yield from a mountain ash forest catchment using a terrain analysis based catchment model. J. Hy-drol. 150:665--700.
Welles, J.M. and J.M. Norman. 1991. Instrument for indirect measure-ment of canopy architecture. Agron. J. 83:818--825.