Summary Naturally seeded Scots pine (Pinus sylvestris L.) trees, age 25--30 years, were subjected to two soil-nitrogen-supply regimes and to elevated atmospheric CO2
concentra-tions by the branch-in-bag method from April 15 to September 15 for two or three years. Gas exchange in detached shoots was measured in a diffuse radiation field. Seven parameters associ-ated with photosynthetic performance and two describing stomatal conductance were determined to assess the effects of treatments on photosynthetic components. An elevated concen-tration of CO2 did not lead to a significant downward regulation
in maximum carboxylation rate (Vcmax) or maximum electron
transport rate (Jmax), but it significantly decreased
light-satu-rated stomatal conductance (gsat) and increased minimum
stomatal conductance (gmin). Light-saturated rates of CO2
as-similation were higher (24--31%) in shoots grown and measured at elevated CO2 concentration than in shoots grown and
meas-ured at ambient CO2 concentration, regardless of treatment
time or nitrogen-supply regime. High soil-nitrogen supply significantly increased photosynthetic capacity, corresponding to significant increases in Vcmax and Jmax. However, the
com-bined elevated CO2 + high nitrogen-supply treatment did not
enhance the photosynthetic response above that observed in the elevated CO2 treatment alone.
Keywords: carboxylation rate, electron transport rate, Pinus sylvestris, stomatal conductance.
Introduction
Photosynthetic responses of plants to elevated atmospheric concentrations of CO2 are complex and depend on many
physi-ological and environmental variables. Despite numerous stud-ies on photosynthetic responses to elevated atmospheric CO2
concentrations, many uncertainties remain (Cure and Acock 1986, Ziska et al. 1990, 1991, Idso et al. 1991, Pettersson et al. 1992, Dufrêne et al. 1993, Gunderson et al. 1993).
Depending on growth conditions and experimental meth-ods, responses of leaf-level photosynthesis to elevated CO2
concentrations may reflect combined changes in biochemical capacity and leaf morphology. Results from gas exchange and metabolic studies (Farquhar and von Caemmerer 1982, Sharkey 1985, von Caemmerer and Edmondson 1986) suggest that changes in the biochemical capacity of photosynthesis can
be induced by Rubisco activity, the rate of RuP2 regeneration
or triosephosphate consumption (Sage et al. 1989, Sage 1990, Stitt 1991). In addition, plants grown under inadequate nutri-tional supplies are often unable to respond to elevated atmos-pheric CO2 concentrations (Eamus and Jarvis 1989, Evans
1989, Arp 1991). Morphological changes in response to ele-vated CO2 may include accumulation of carbohydrates as a
result of inadequate sink demands for photosynthates (Clough et al. 1981, Sage et al. 1989), changes in leaf thickness and changes in mesophyll cell number (Vu et al. 1989). In princi-ple, well-established models can be used to quantify modifica-tions in these key photosynthetic components (Harley et al. 1992, McKee and Woodward 1994, Mitchell et al. 1995).
The response of trees to elevated CO2 concentrations may be
expected to mirror that of other C3 species; however, forest tree
longevity raises questions about long-term effects about which little is known. For example, it is not known whether short-term enhancement of photosynthesis by elevated CO2
concen-tration can be sustained over several growing seasons. For several reasons, most CO2 enrichment experiments have been
confined to short-term studies on juvenile trees (Eamus and Jarvis 1989), but it is not known whether these observations can be extrapolated to mature trees (Cregg et al. 1989). The branch-in-bag method makes it possible to examine the effects of elevated CO2 concentrations on mature trees without the
technical difficulties and expense of subjecting entire trees to the CO2 treatment (Barton et al. 1993, Dufrêne et al. 1993).
The validity of branch-in-bag experiments is based on the finding that branches are relatively autonomous with respect to their carbon budget during the growing season. The assump-tion that branches do not import carbon but only export it to woody tissues and roots is considered valid for large branches after initial shoot elongation has ceased (Sprugel et al. 1991).
We measured gas exchange in shoots of Scots pine (Pinus sylvestris L.) trees grown in two atmospheric CO2
concentra-tions and two soil-nitrogen regimes for two to three years. Specifically, we determined: (1) how long-term elevation of CO2 concentration affects photosynthetic performance; (2)
whether a high soil-nitrogen supply enhances the response of photosynthesis to elevated atmospheric CO2 concentrations;
(3) whether different lengths of exposure (two or three years) to elevated CO2 concentration lead to differences in
photosyn-Photosynthetic responses of Scots pine to elevated CO
2
and nitrogen
supply: results of a branch-in-bag experiment
SEPPO KELLOMÄKI and KAI-YUN WANG
University of Joensuu, Faculty of Forestry, P.O.Box 111, FIN-80101 Joensuu, Finland
Received February 28, 1996
thetic capacity; and (4) whether there is a coupled relationship between modifications in photosynthetic components and leaf nitrogen concentration. The main physiological parameters concerning photosynthetic processes were determined by fit-ting the data to a set of photosynthetic models and to an empirical model of stomatal conductance.
Materials and methods
Experimental design
The experiment was established in a naturally seeded stand of Scots pine near the Mekrijärvi Research Station (62°47′ N, 30°58′ E, 145 m elevation), University of Joensuu, Finland. The soil on the site is sandy with a water retention of 7 mm at field capacity and 3 mm at the wilting point for the top 30-cm layer of soil. The mean density of the pure Scots pine stand was about 2500 stems per hectare. The trees were 25--30 years old and had a mean height of 4--5 m.
Three branches of the third whorl from the stem apex on each tree were chosen for the treatments: one branch was placed in a bag and subjected to an elevated CO2 concentration
(Bagged + CO2); one branch was placed in a bag and subjected
to ambient CO2 concentration (Bagged); and one branch was
left as an unbagged control (Unbagged). Starting in early spring of 1993, six sample trees representing six replicates of each treatment were exposed to the CO2 treatments. In early
spring of 1994, a similar experiment was set up with 12 trees per CO2 treatment. Within each CO2 treatment, half of the trees
were grown under normal soil conditions of the site (Normal-N), whereas the remaining trees received an improved nitrogen supply (High-N); i.e., 0.7 kg of pure nitrogen in the form NH4NO3 was added to a 4 × 4 m area around each tree.
The design of the branch bags was similar to that described by Barton et al. (1993). Briefly, the bags consisted of a cylin-drical wire frame covered with a polyethylene sleeve which was made of PVC plastic (thickness 0.4 mm, protected from deterioration by UV radiation) and had a volume of 0.379 m3 (length 1.65 m, diameter 0.6 m). The sleeve material prevented only a small fraction of radiation (approximately 10%), within the spectral range 400--800 nm, from entering the bags, and had only a small impact on the spectral composition of visible radiation.
Air was supplied to each bag through a 10-cm-diameter duct. Air in the duct was driven by a blower fan (maximum air flow = 190 dm3 s−1) that was mounted in a weather-proof housing. A butterfly valve was positioned in the duct, close to the housing, to enable manual setting of the air flow rate which was usually 0.02--0.025 m−3 s−1.
The CO2 concentration inside the bags providing the
ele-vated CO2 treatment was increased to 670--820 µmol mol−1
from April 15 to September 15, by injecting pure CO2 into the
duct (Figure 1). The CO2 concentration inside the bags was
monitored with a CO2 analyzer (Model 7MB1300-0BA00,
Siemens, Germany).
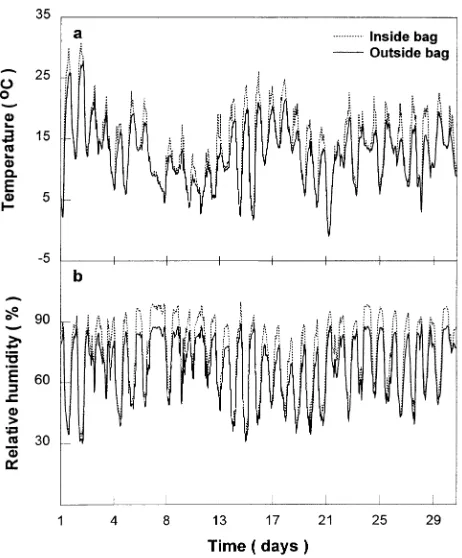
Relative humidity and temperature inside and outside the bags were recorded separately with a probe combining a
tem-perature sensor and a Humicap relative humidity sensor (Model HMP131Y, Vaisala, Vantaa, Finland). During most of the day, temperature and relative humidity inside the branch bags were only slightly higher than ambient values (Figures 2a and 2b) except during short periods of intense sunshine, when tem-peratures inside the bags exceeded ambient by 4--6 °C (about 10% of the time) and relative humidity inside the bags ex-ceeded ambient by 10--15% (about 20% of the time). Figure 1. Frequency distribution of atmospheric CO2 concentration in
bags supplied with CO2-enriched air. The plot is based on hourly
means of 30-s readings. Data are for the period April 15 to September 15, 1995.
Measurement and analysis of gas exchange
Measurements of CO2 exchange in a diffuse radiation field
were conducted in the laboratory with an automatic open-gas-exchange system as described by Wang (1996).
A one-year-old shoot was excised under water and trans-ported to the laboratory. After the shoot had been recut under water, a 10-cm-long section of shoot with only 30 needle fascicles (surplus needles had been thinned evenly one week before conducting the measurements to minimize shoot struc-ture effects) was sealed in the assimilation cuvette. The base of the shoot was inserted in a tube providing a constant water supply throughout the measurements.
Gas-exchange measurements were conducted on two shoots from each branch bag through July and August 1995. Photo-synthetic parameters were first estimated on the basis of a single branch bag. Then the mean of the estimated parameters from the six replicated branch bags representing the same treatment was used to analyze the results. During all measure-ments, air temperature in the assimilation cuvette was main-tained at 20 ± 0.5 °C and water vapor pressure deficit remained below 0.6 kPa, except during measurements to estimate sto-matal conductance parameters. Gas-exchange parameters were calculated by the method of von Caemmerer and Far-quhar (1981). Rate of net assimilation (An) was based on
projected needle area measured with a leaf-area meter (LI-3100, Li-Cor, Inc., Lincoln, NE).
Immediately after the gas-exchange measurements, all need-les were removed from the shoot, after which their projected area, fresh weight, and weight after drying at 75 °C were determined. In addition, their nitrogen concentration was de-termined by the micro-Kjeldahl method.
Estimation of photosynthetic parameters
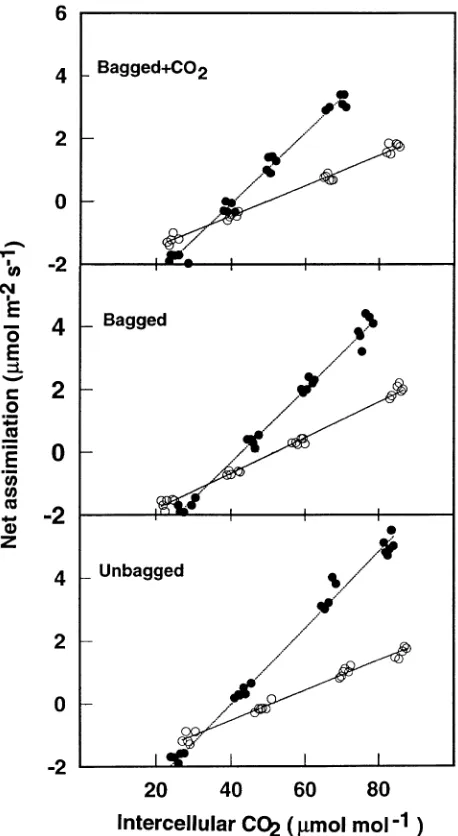
Three sets of response curves were obtained for each shoot. First, slopes of An versus the calculated intercellular
concentra-tion of CO2 (An/Ci curve) at photon flux densities (Qp) of 150
and 700 µmol m−2 s−1 were determined by least-squares linear regression. Respiration rate in light (Rd) and CO2
compensa-tion point in the absence of nonphotorespiratory respiracompensa-tion (Γ∗) were estimated from the intercept of the two slopes, as described by Brooks and Farquhar (1985). Representative ex-amples of An/Ci curves, measured at the two Qp values used to
estimate Rd and Γ*, are shown in Figure 3.
Then An/Ci curves were measured at nearly saturating Qp
(1500 µmol m−2 s−1). Based on the estimated Rd and Γ*,
Equa-tion 1 was fitted to each An/Ci curve. The maximum RuP2
-satu-rated rate of carboxylation (Vcmax) was estimated for each
branch bag:
Anc=
Vcmax(Ci−Γ∗)
Ci+ Kc(1 + Oi/Ko)
− Rd, (1)
where Anc is RuP2-saturated rate of CO2 assimilation, Oi is
intercellular concentration of O2 (198 µmol mol−1), and Kc and
Ko are the Michaelis constants for carboxylation and
oxygena-tion, respectively. Values of Kc (358.6) and Ko (255.9) at 20 °C
were derived from the measurements as described by Badger and Collatz (1977).
Finally, An/Qp curves were measured at a CO2 concentration
of 2000 ± 10 µmol mol−1. Three parameters, maximum rate of electron transport (Jmax), effectivity factor for use of light (q)
and convexity factor of light-response curve (θ) were esti-mated in Equation 2 by the nonlinear least-squares regression techniques described by Ziegler-Jöns and Selinger (1987): Figure 3. Representative partial An/Ci curves measured at two
irrad-iances, 150 µmol m−2 s−1 (s) and 700 µmol m−2 s−1 (d)air tempera-ture at 20 ± 0.5 °C and vapor pressure deficit less than 0.6 kPa, for shoots grown under different environmental conditions (Bagged + CO2, Bagged, and Unbagged) for three growing seasons. Slopes were
fitted by least-squares linear regression. The CO2 compensation point,
Γ*, in the absence of nonphotorespiratory respiration and respiration rate in light, Rd, were estimated by solving for the intercept of the two
Anj=
Jmax+ qQp−[(Jmax + qQp)2− 4θqQpJmax]0.5
9θ − Rd, (2)
where Anj is the RuP2-regeneration limited rate of CO2
assimi-lation.
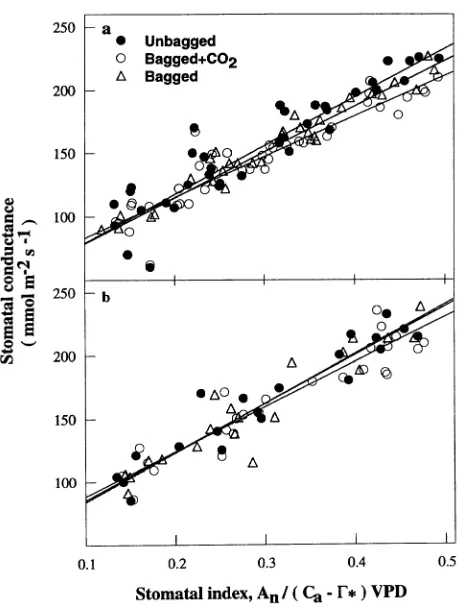
Estimate of stomatal parameters
The model of Ball et al. (1987), modified by Leuning (1990, 1995) and Collatz et al. (1991), was applied to describe the response of stomatal conductance, gs, to variations in net
assimilation rate (An), leaf-to-air vapor pressure difference
(Ds), and CO2 mole fraction (Cs) at the leaf surface:
gs= gmin+ g1
1000 An
Ds(Cs−Γ∗)
, (3)
where gmin is minimum stomatal conductance to water vapor
and g1 is an empirical coefficient. We assumed: (1) Cs equals
the CO2 mole fraction inside the cuvette (Ca); (2) units are
corrected by the factor 1000 when all units in Equation 3 coincide with those presented in the Appendix; (3) Ds is the
difference between saturated vapor pressure at air temperature and actual vapor pressure inside the cuvette, because leaf temperature maintained a relatively constant boundary con-ductance that was fairly large (750--800 mmol m−2 s−1). Two stomatal parameters, gmin and g1, were estimated by fitting
Equation 3 to observations within the ranges: 0 < Qp < 1500
µmol m−2 s−1, 50 < Cs < 1400 µmol mol−1, and 0.3 < Ds <
2.0 kPa (Figure 4).
Statistical analysis
Responses of photosynthesis to environmental factors were analyzed in terms of the parameters of the fitted function. Analysis of variance was applied to test the effects of growth conditions. First, effects of CO2 concentration and
soil-nitro-gen supply on each parameter were analyzed (multifactor ANOVA) using the SPSS/PC software package (SPSS, Chi-cago, IL) based on shoots grown in bags, assuming that signifi-cant responses were attributable to the aforementioned treatments, and not to some unknown bagging effects. Treat-ment time was analyzed as repeated measures during the two growth seasons. Finally, one-way ANOVA was applied to test for the effect of bagging.
Results
Maximum RuP2-saturated rate of carboxylation, Vcmax, and
maximum rate of electron transport, Jmax
Both Vcmax and Jmax were significantly affected by the High-N
treatment (Tables 1 and 2). Compared to the Normal-N treat-ment, the High-N treatment increased Vcmax and Jmax on
aver-age by 33 and 11%, respectively. Regardless of duration of exposure, elevated CO2 alone did not significantly affect
Vcmax or Jmax, although Vcmax was depressed 11% after the third
growing season. Effects of elevated CO2 on Vcmax and Jmax
were not enhanced by High-N (Table 2). Bagging resulted in only a slight decrease in Jmax (Table 1).
Carbon dioxide compensation point in the absence of dark respiration, Γ*, and respiration rates
The CO2 compensation point (Γ*), and respiration rates in the
light (Rd) and the dark (Rn) were significantly affected by
elevated CO2 (Tables 1 and 2). Compared to the bagged
treat-ment alone, elevated CO2 concentration decreased Γ* on
aver-age by 9%, and increased Rd and Rn by 22 and 18%,
respectively. Furthermore, only in the case of Rn was this
increase significantly enhanced by both High-N and treatment time. The High-N treatment resulted in a slight increase in the CO2-induced increase in Rd. There was no effect of bagging on
any of the three parameters; however, treatment time led to a significant increase (17%) in Rn (Table 1).
Figure 4. Stomatal conductance, gs, plotted against the stomatal index
(An/[(Cs−Γ∗)Ds]) of Equation 3 for shoots of Scots pine grown in
different environmental conditions (Bagged + CO2, Bagged, and
‘‘Effectivity factor’’ for the use of light, q, and convexity factor of light-response curve, θ
Values of θ were significantly increased 23% by High-N, 13% by elevated CO2 concentration, 14% by time of treatment, and
13% by the bagged treatment (Tables 1 and 2). In contrast, values of q were significantly increased (by 13%) only by the High-N treatment. The bagged treatment caused a 2% reduc-tion in values of q.
Minimum stomatal conductance, gmin, and the empirical
coefficient, g1
Compared with the bagged treatment alone, elevated CO2
significantly increased gmin by 12% on average, but this
in-crease was not enhanced by High-N (Tables 1 and 2). Both time of treatment and the bagged treatment significantly in-creased gmin by 13 and 20%, respectively. Treatment effects on
g1 were the opposite of those observed on gmin. Thus, elevated
CO2 significantly decreased g1 by 10%, whereas High-N
sig-nificantly increased g1 by 11%, and the bagged treatment and
treatment time caused slight decreases in g1 (Table 2).
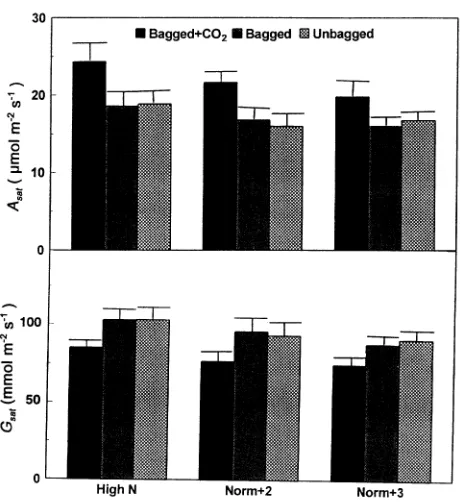
Light-saturated rate of assimilation, Asat, and stomatal
conductance, gsat
Figure 5 presents Asat and gsat as functions of the treatments.
Compared to bagged shoots alone, elevated CO2 concentration
increased Asat by 31, 29 and 24%, and decreased gsat by 17, 19
and 16% separately for shoots grown in High-N), Normal-N after two growth seasons (Norm + 2) and Normal-N after three growth seasons (Norm + 3). However, there were no signifi-cant differences in Asat or gsat between bagged shoots and
unbagged shoots.
Nitrogen concentration of needles
Regardless of CO2 concentration and treatment time, High-N
resulted in a significant increase (P < 0.01) in area-based needle nitrogen concentration, but not in mass-based needle nitrogen concentration (Figure 6). Elevated CO2 treatment
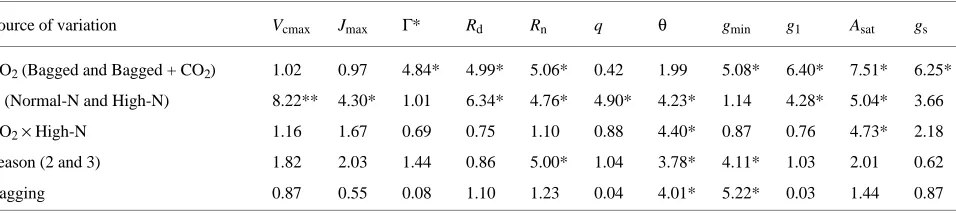
decreased needle nitrogen concentration, but the decrease was only significant (P < 0.05) for mass-based needle nitrogen concentration. Neither the bagged treatment nor treatment Table 2. Summary of F-values from multifactor ANOVA for the effects of growth CO2 regime and soil-nitrogen supply on photosynthetic
parameters, assuming that significant responses were due to CO2 and nitrogen, and not to some unknown bagging effect. The environmental
treatments are CO2 concentration (ambient [CO2 ] (Bagged) and elevated [CO2] (Bagged + CO2)), and soil-nitrogen supply (normal nitrogen supply (Normal-N) and high soil-nitrogen supply (High-N)). Treatment times were treated as repeated measures during two growth seasons. Bagging effects were analyzed by one-way ANOVA. Least significant differences (LSD) was used for the range test. Legend: * P < 0.05; **P < 0.001.
Source of variation Vcmax Jmax Γ* Rd Rn q θ gmin g1 Asat gs
CO2 (Bagged and Bagged + CO2) 1.02 0.97 4.84* 4.99* 5.06* 0.42 1.99 5.08* 6.40* 7.51* 6.25*
N (Normal-N and High-N) 8.22** 4.30* 1.01 6.34* 4.76* 4.90* 4.23* 1.14 4.28* 5.04* 3.66
CO2× High-N 1.16 1.67 0.69 0.75 1.10 0.88 4.40* 0.87 0.76 4.73* 2.18
Season (2 and 3) 1.82 2.03 1.44 0.86 5.00* 1.04 3.78* 4.11* 1.03 2.01 0.62
Bagging 0.87 0.55 0.08 1.10 1.23 0.04 4.01* 5.22* 0.03 1.44 0.87 Table 1. Parameter estimates as functions of treatments. Data are means ± SE of estimated values from six replicated branch bags per treatment. ‘‘Bagged + CO2’’ are bagged shoots with CO2 concentration of 700 µmol mol--1; ‘‘Bagged’’ are bagged shoots with CO2 concentration of 350 µmol mol --1 and ‘‘Unbagged’’ are unbagged shoots with CO2 concentration of 350 µmol mol --1.
Treatment Vcmax Jmax Γ* Rd Rn q θ gmin g1
After two growing seasons with High-N
Bagged + CO2 60.3 ± 2.7 172.6 ± 6.1 32.11 ± 2.4 1.090 ± 0.06 1.678 ± 0.04 0.388 ± 0.03 0.87 ± 0.05 53.7 ± 1.3 3.55 ± 0.20
Bagged 59.8 ± 1.4 169.7 ± 4.5 35.07 ± 1.8 0.918 ± 0.04 1.311 ± 0.07 0.396 ± 0.04 0.74 ± 0.03 47.9 ± 2.1 3.83 ± 0.30 Unbagged 61.6 ± 2.2 177.1 ± 4.2 35.22 ± 2.1 0.929 ± 0.07 1.347 ± 0.06 0.401 ± 0.06 0.72 ± 0.07 44.6 ± 1.4 3.91 ± 0.34
After two growing seasons with Normal-N
Bagged + CO2 43.7± 2.1 156.3 ± 6.3 31.86 ± 2.2 0.874 ± 0.07 1.203 ± 0.04 0.349 ± 0.02 0.70 ± 0.04 52.8 ± 2.0 3.08 ± 0.26 Bagged 47.8 ± 2.7 152.8 ± 7.4 34.79 ± 1.9 0.788 ± 0.05 1.112 ± 0.05 0.347 ± 0.03 0.60 ± 0.03 47.2 ± 1.7 3.57 ± 0.21 Unbagged 45.5 ± 3.2 158.4 ± 4.1 35.00 ± 1.7 0.780 ± 0.08 1.147 ± 0.04 0.350 ± 0.05 0.59 ± 0.08 42.3 ± 1.8 3.56 ± 0.27
After three growing seasons with Normal-N
Bagged + CO2 39.7 ± 3.7 145.2 ± 9.3 30.84 ± 2.4 0.963 ± 0.04 1.474 ± 0.06 0.320 ± 0.07 0.76 ± 0.06 59.6 ± 2.6 2.90 ± 0.23
time led to a significant change in needle nitrogen concentra-tion.
Discussion
We assessed whether long-term treatment with elevated CO2
concentration and high soil-nitrogen supply modifies the photosynthetic capacity of Scots pine. We applied the branch-in-bag technique (Barton et al. 1993), where single branches in the same whorl are subjected to CO2 treatments or are used to
control the effect of the bag itself. By this means it is possible to reduce the effects of genetic and physiological status of the tree on the results. This technique also facilitates study of the effects of elevated CO2 concentration on physiological and
developmental processes in mature trees. However, despite these advantages, the technique subjects only a small fraction of the tree to elevated CO2 concentrations, and subsequent
source--sink relationships could differ greatly from those of whole trees under natural conditions.
Because of the complex structure of a Scots pine shoot, irradiation of needle surfaces is not uniform (Oker-Blom et al. 1992), and the needle area receiving direct radiation is often difficult to determine (Leverenz and Jarvis 1978, Carter and
Smith 1985). We attempted to overcome these technical diffi-culties by using shoots with relative few needles (i.e., needles were thinned evenly to give 30-needle fascicles per sample shoot) so that no point on the needle surface was shaded in all directions under a uniform diffuse radiation field. Therefore, the photon flux density incident on the needle surface was assumed to be equal to the irradiance incident on any surface in the diffuse radiation field (Oker-Blom et al. 1992). Further, the projected area of needles can be measured directly in the laboratory.
The total radiation in the spectral range 400--800 nm enter-ing the bag was about 90% of that outside the bag, implyenter-ing that bagged branches did not have to acclimate to a substan-tially altered radiation regime. Therefore, bag effects on some photosynthetic parameters (Table 1) are probably associated with increases in temperature or relative humidity (Figure 1); however, bag effects did not lead to significant differences in the acclimation of key parameters such as Vcmax and Jmax.
Although branches were exposed to the elevated-CO2
treat-ment from April 15 to September 15, a period longer than the average growing season outside the bags, there was no signifi-cant downward regulation in the capacity for CO2
carboxyla-tion (Vcmax) or maximum electron transport (Jmax) regardless of
Figure 5. Light-saturated rate of assimilation, Asat, and stomatal
con-ductance, gsat, as a function of treatment. Value is the mean of
meas-urements from six replicated bags per treatment. Measuring conditions: photon flux density of 1500 µmol m−2 s−1, vapor pressure deficit of 0.4 kPa and air temperature of 20 °C for all treatments; CO2
concentrations, Ca, of 350 µmol mol−1 for Bagged and Unbagged shoots; 700 µmol mol−1 for Bagged + CO2 shoots. High-N = high
soil-nitrogen supply after two growth seasons; Norm + 2 = normal soil-nitrogen content after two growth seasons; Norm + 3 = normal soil-nitrogen content after three growth seasons.
soil-nitrogen supply and treatment time (Besford and Hand 1989, Sage et al. 1989, Harley et al. 1992, Nie and Long 1992, Van Oosten et al. 1992) (Tables 1 and 2). When measured at the CO2 concentration in which the shoots were grown, the
light-saturated rate of CO2 assimilation was higher (24--31%)
in shoots grown at elevated CO2 concentration than in shoots
grown at current ambient CO2 concentration (Figure 5).
Reasons for this difference in response could be that: (i) only a small part of each tree was subjected to elevated CO2
concen-trations; (ii) the trees were in a stage of rapid growth (a big ‘‘sink’’ in other parts of the trees and a high demand for carbohydrates would have accompanied (i) and (ii)); (iii) com-pared to trees grown in a pot, relatively unrestricted root growth of field-grown trees may help prevent sink limitation and end-product inhibition; and (iv) the presence of active sinks for recently assimilated metabolites may also be impor-tant for maintenance of photosynthetic capacity (Stitt 1991, Thomas and Strain 1991). The finding that Scots pine can sustain increased capacity for carboxylation after long-term exposure to elevated CO2 concentrations suggests that this
species will exhibit increased growth and carbon storage with future increases in atmospheric CO2 concentration.
Values of Γ* were similar among treatments, except in the elevated CO2 treatment (Table 1). This absence of treatment
effects was expected because the value of Γ* depends mainly on the kinetic characteristics of Rubisco (Laisk 1977, Jordan and Ogren 1981, von Caemmerer et al. 1994), which are unlikely to change rapidly (Long and Drake 1991). The aver-age Γ* value of 33.54 µmol mol−1 is close to the value of 35.5 µmol mol−1 found by Brooks and Farquhar (1985) for Spi-nacia oleracea L. leaves. The relatively low value of Γ* in the elevated CO2 treatment may have resulted from using the
x coordinate (intercellular CO2 partial pressure) at the intercept
of the two slopes as the value of Γ* (see Figure 2). This estimate ignores the effect of conductance for CO2 transfer
from substomatal cavities to the sites of carboxylation (gw), on
the value of Γ* (see Brooks and Farquhar 1985 and von Caemmerer et al. 1994, for detailed discussion on measure-ments of Γ*). If valid, this explanation implies that the de-crease in Γ* in response to the elevated CO2 treatment was
associated with changes in gw and Rd.
Our results showing a significant increase in Rd with
ele-vated CO2 and high soil-nitrogen supply, are consistent with
the results of McKee and Woodward (1994), but differ from Thomas et al. (1993) who found no effect. Studies on the long-term effect of elevated CO2 on dark respiration (Rn) have
shown increases (Hrubec et al. 1985, Poorter et al. 1988), decreases (Reuveni and Gale 1985, Bunce 1990, Wullschleger et al. 1992) or no effect (Gifford et al. 1985, Hrubec et al. 1985) depending on species, age, plant part, or reference units. Wang et al. (1996) have shown that the direction of change in Rn of
Scots pine shoots grown in open-top chambers in elevated CO2
depends on both growing conditions and temperature during measurements.
Many studies (e.g., Sharp et al. 1984, Brooks and Farquhar 1985) have shown that Rd is correlated with Rn. We observed
that different treatments induced similar changes in Rd and Rn
(Table 1) resulting in relatively small variations (0.58--0.72) in the ratio of Rd to Rn. The mean ratio value of 0.68 was higher
than that of 0.34 estimated by Brooks and Farquhar (1985) for Spinacia oleracea, but still within the range (0.25--1.0) meas-ured by others (Peisker et al. 1981, Azcón-Bieto and Osmond 1983).
The elevated CO2 treatment decreased the slope of stomatal
functions (g1 in Equation 3), but increased minimum stomatal
conductance (gmin; Table 1, Figure 4) (cf. Harley et al. 1992).
This finding is consistent with the measured decrease in light-saturated stomatal conductance in shoots grown in bags and elevated CO2 (Figure 5), and implies that elevated CO2 could
limit stomata adaptability to changing environmental condi-tions.
Because leaf nitrogen is a major component of Rubisco (Evans 1989), it is often closely correlated with photosynthetic performance (Brix 1981, Matyssek and Schulze 1987). Al-though there is not a direct relationship between leaf nitrogen concentration and soil nitrogen content, fertilization usually results in increased foliar nitrogen concentrations in Scots pine, perhaps because pine forests commonly occur on nutri-ent-poor forest soils (Miller et al. 1979, Sheriff et al. 1986, Näsholm and Ericsson 1990, Mitchell and Hinckley 1993). In response to nitrogen fertilization, the increase in area-based needle nitrogen concentration was closely coupled with the increase in light-saturated assimilation rate (Figures 5 and 6), regardless of growing CO2 concentration. Similar responses
have been reported by Mitchell and Hinckley (1993) for Pseudotsuga menziesii (Mirb.) Franco.
Long-term exposure to elevated CO2 often alters
photosyn-thetic capacity as a result of nitrogen reallocation from Ru-bisco to other photosynthetic components, or to nonphoto-synthetic processes (Evans 1989, Newton 1991, Stitt 1991, Conroy 1992, Tissue et al. 1993). Such acclimation is variable because it depends on several factors. Compared to the Normal-N, High-N increased maximum carboxylation rate more than maximum electron transport rate (33 versus 11%), indicating that high soil-nitrogen supply had a greater effect on carboxy-lation rate than on electron transport capacity (cf. Evans and Terashima 1987, Tissue et al. 1993). However, a comparison of the Bagged shoots and Bagged + CO2 shoots indicated that
increasing soil-nitrogen supply did not significantly increase the magnitude of photosynthetic response to elevated CO2
concentration, implying that photosynthetic response to ele-vated CO2 was not limited by soil-nitrogen content at the site.
An analytical model is only as good as the set of assump-tions underlying it. Our parameter estimates were based on simplified model assumptions: Rubisco was assumed to be fully activated; and Ci was assumed to equal the C
concentra-tion at the site of fixaconcentra-tion, which ignores possible liquid resis-tances between the intercellular space and carboxylation site. The error resulting from this assumption has been estimated by Farquhar and von Caemmerer (1982) (see also Harley et al. 1985). We believe, however, that this approach represents an advance in our ability to understand CO2 exchange kinetics at
ecophysiological comparisons among plants acclimated to dif-ferent environments.
We conclude that two or three years of exposure to elevated CO2 concentrations did not significantly reduce the
photosyn-thetic capacity of Scots pine. However, elevated CO2
concen-trations decreased light-saturated stomatal conductance. A high soil-nitrogen supply significantly increased the photosyn-thetic capacity in Scots pine as a result of significant increases in maximum carboxylation rate, Vcmax and maximum electron
transport rate, Jmax. However, high soil-nitrogen supply did not
increase photosynthetic response to elevated CO2
concentra-tion.
Acknowledgments
This study is part of a research project entitled ‘‘Likely Impact of Elevating CO2 and Temperature on European Forests’’ under the coor-dination of Prof. Paul Jarvis, University of Edinburgh. The study was funded through the Finnish Climate Change Study Programme (SILMU), and the University of Joensuu. We express our appreciation to Jorma Aho for providing the facilities at the Mekrijärvi Research Station; thanks also to Matti Lemettinen and Alpo Hassinen for devel-oping and maintaining the experimental infra-structure.
References
Arp, W.J. 1991. Effects of source--sink relations to photosynthetic acclimation to elevated CO2. Plant Cell Environ. 14:869--875. Azcòn-Bieto, J. and C.B. Osmond. 1983. Relationship between
pho-tosynthesis and respiration. The effect of carbohydrate status on the rate of CO2 production by respiration in darkened and illuminated
wheat leaves. Plant Physiol. 71:574--581.
Badger, M. and G.J. Collatz. 1977. Studies on the kinetic machine of ribulose 1,5-bisphosphate carboxylase and oxygenase reaction, with particular reference to the effect of temperature on kinetic parameters. Carnegie Institute, Washington Yearbook, 76:355--361. Ball, J.T., I.E. Woodrow and J.A. Berry. 1987. A model predicting stomatal conductance and contribution to the control of photosyn-thesis under different environment conditions. In Progress in Pho-tosynthesis Research. Ed. I. Biggins. Proc. VII International Congress on Photosynthesis, Vol. IV. Martinus Nijhoff, Dordrecht, pp 221--224.
Barton, C.V.M., H.S.J. Lee and P.G. Jarvis. 1993. A branch bag and CO2 control system for long-term CO2 enrichment of mature Sitka
spruce (Picea sitchensis (Bong.) Carr.). Plant Cell Environ. 16:1139--1148.
Besford, R.T. and D.W. Hand. 1989. The effects of CO2 enrichment and nitrogen oxides on some Calvin cycle enzymes and nitrate reductase in glasshouse lettuce. J. Exp. Bot. 40:329--336. Brix, H. 1981. Effects of nitrogen fertilizer source and application
rates on foliar nitrogen concentration, photosynthesis, and growth of Douglas-fir. Can. J. For. Res. 11:775--780.
Brooks, A. and G.D. Farquhar. 1985. Effect of temperature on the CO2/O2 specificity of ribulose-1,5-bisphosphate
carboxylase/oxy-genase and the rate of respiration in the light. Estimates from gas-exchange measurements on spinach. Planta 165:397--406. Bunce, J.A. 1990. Short- and long-term inhibition of respiratory carbon
dioxide efflux by elevated carbon dioxide. Ann. Bot. 65:637--642.
Carter, G.A. and W.K. Smith. 1985. Influence of shoot structure on light interception and photosynthesis in conifers. Plant Physiol. 79:1038--1043.
Clough, J.M., M.M. Peat and P.J. Kramer. 1981. Effect of high atmos-pheric CO2 and sink size on rates of photosynthesis of a soybean cultivar. Plant Physiol. 67:1007--1010.
Collatz, G.J., J.T. Ball, C. Grivet and J.A. Berry. 1991. Physiological and environmental regulation of stomatal conductance, photosyn-thesis and transpiration: a model that includes a laminar boundary layer. Agric. For. Meteorol. 54:107--136.
Conroy, J.P. 1992. Influence of elevated atmosphere CO2 concentra-tions on plant nutrition. Aust. J. Bot. 40:445--456.
Cregg, B.M., J.E. Halpin, P.M. Dougherty and R.O. Teskey. 1989. Comparative physiology and morphology of seedling and mature forest trees. In Air Pollution Effects on Forest Vegetation. Eds. R.D. Noble, J.L. Martin and K.F. Jensen. John Wiley and Sons, Chiches-ter, pp 21--50.
Cure, J.D. and B. Acock. 1986. Crop responses to carbon dioxide doubling: a literature survey. Agric. For. Meteorol. 38:127--145. Dufrêne, E., J.Y. Pontailler and B. Saugier. 1993. A branch bag
technique for simultaneous CO2 enrichment and assimilation
meas-urements on beech (Fagus sylvatica L.). Plant Cell Environ. 16:1131--1138.
Eamus, D. and P.G. Jarvis. 1989. The direct effects of increase in the global atmospheric CO2 concentration on natural and commercial
temperate trees and forests. Adv. Ecol. Res. 19:1--55.
Evans, J.R. 1989. Photosynthesis and nitrogen relationships in leaves of C3 plants. Oecologia 78:9--19.
Evans, J.R. and I. Terashima. 1987. Effects of nitrogen nutrition on electron transport components and photosynthesis in spinach. Aust. J. Plant Physiol. 14:59--68.
Farquhar, G.D. and S. von Caemmerer. 1982. Modelling of photosyn-thetic response to environmental conditions. In Encyclopedia of Plant Physiology, Vol. 12B. Eds. O.L. Lange, P.S. Nobel, C.B. Osmond and H. Ziegler. Springer-Verlag, Heidelberg, New York, pp 549--587. Gifford, R.M., H. Lambers and J.I.L. Morison. 1985. Respiration of
crop species under CO2 enrichment. Physiol. Plant. 63:351--356.
Gunderson, C.A., R.J. Norby and S.D. Wullschleger. 1993. Foliar gas exchange responses of two deciduous hardwoods during 3 years of growth in elevated CO2: no loss of photosynthetic enhancement.
Plant Cell Environ. 16:797--807.
Harley, P.C., J.A. Weber and D.M. Gates. 1985. Interactive effects of light, leaf temperature, CO2 and O2 on photosynthesis in soybean.
Planta 165:249--263.
Harley, P.C., R.B. Thomas, J.F. Reynolds and B.R. Strain. 1992. Modelling photosynthesis of cotton grown in elevated CO2. Plant Cell Environ. 15:271--282.
Hrubec, T.C., J.M. Robinson and R.P. Donaldson. 1985. Effects of CO2 enrichment and carbohydrate content on the dark respiration of
soybeans. Plant Physiol. 79:684--689.
Idso, S.B., B.A. Kimball and S.G. Allen. 1991. Net photosynthesis of sour orange trees maintained in atmospheres of ambient and ele-vated CO2 concentration. Agric. For. Meteorol. 54:95--101.
Jordan, D.B. and W.L. Ogren. 1981. Species variation in the specificity of ribulose bisphosphate carboxylase/oxygenase. Nature 291:513--423. Laisk, A.K. 1977. Kinetics of photosynthesis and photorespiration in
C3-plants. Nauka, Moscow, pp 123−146.
Leuning, R. 1990. Modelling stomatal behaviour and photosynthesis of Eucalyptus grandis. Aust. J. Plant Physiol. 7:159--175. Leuning, R. 1995. A critical appraisal of a combined
Leverenz, J.W. and P.G. Jarvis. 1978. Photosynthesis of Sitka spruce (Picea sitchensis (Bong.) Carr.) VIII. The effect of light flux density and direction on rate of net photosynthesis and stomatal conduc-tance of needles. J. Appl. Ecol. 16:919--932.
Long, S.P. and B.G. Drake. 1991. Effects of the long-term elevation of CO2 concentration in the field on the quantum yield of photosynthe-sis of the C3 sedge, Scirpus olneyi. Plant Physiol. 96:221--226. Matyssek, R. and E.-D. Schulze. 1987. Heterosis in hybrid larch
(Larix decidua x leptolepsis). II. The role of leaf characteristics. Trees 1:225--231.
McKee, I.F. and F.I. Woodward. 1994. The effect of growth at elevated CO2 concentrations on photosynthesis in wheat. Plant Cell Environ.
17:853--859.
Miller, H.G., J.M. Cooper and J.D. Miller. 1979. Nutrient cycles in pine and their adaptations to poor soils. Can. J. For. Res. 9:19--26. Mitchell, A.K. and T.M. Hinckley. 1993. Effects of foliar nitrogen concentration on photosynthesis and water use efficiency in Douglas-fir. Tree Physiol. 12:403--410.
Mitchell, R.A.C., D.W. Lawlor, V.J. Mitchell, C.L. Gibbard, E.M. White and J.R. Poter. 1995. Effects of elevated CO2 concentration
and increased temperature on winter wheat: test of ARCWHEAT1 simulation model. Plant Cell Environ. 18:736--748.
Näsholm, T. and A. Ericsson. 1990. Seasonal changes in amino acids, protein and total nitrogen in needles of fertilized Scots pine trees. Tree Physiol. 6:267--281.
Newton, P.C.D. 1991. Direct effects of increasing carbon dioxide on pasture plants and communities. N.Z. J. Agric. Res. 34:1--24. Nie, G.Y. and S.P. Long. 1992. The effect of prolonged growth in
elevated CO2 concentrations in the field on the amounts of different
leaf proteins. In Research in Photosynthesis, IV. Ed. N. Murata. Kluwer Academic Publishers, Amsterdam, pp 855--858.
Nonaka, M. 1974. Improved paint coating for photometric integrators. Lighting Res. Tech. 6:30--31.
Oker-Blom, P., T. Lahti and H. Smolander. 1992. Photosynthesis of a Scots pine shoot: a comparison of two models of shoot photosynthe-sis in direct and diffuse radiation fields. Tree Physiol. 10:111--125. Peisker, M., I. Tichá and J. Èatsky. 1981. Ontogenetic changes in the
internal limitations to bean-leaf photosynthesis. 7. Interpretation of the linear correlation between CO2 compensation concentration and CO2 evolution in darkness. Photosynthetica 15:161--168.
Pettersson, R. and A.J.S. McDonald. 1992. Effects of elevated carbon dioxide concentration on photosynthesis and growth of small birch plants (Betula pendula Roth.) at optimal nutrition. Plant Cell Envi-ron. 15:911--919.
Poorter, H., S. Pot and H. Lambers. 1988. The effect of an elevated atmospheric CO2 concentration on growth, photosynthesis and res-piration of Plantago major. Physiol. Plant. 73:553--559.
Reuveni, J. and J. Gale. 1985. The effect of high levels of carbon dioxide on dark respiration and growth of plants. Plant Cell Envi-ron. 8:623--628.
Sage, R.F. 1990. A model describing the regulation of ribulose-1,5-bisphosphate carboxylase, electron transport and triosephosphate use in response to light intensity and CO2 in C3 plants. Plant
Physiol. 94:1728--1734.
Sage, R.F., T.D. Sharkey and J.R. Seemann. 1989. Acclimation of photosynthesis to elevated CO2 in five C3 species. Plant Physiol.
89:590--596.
Sharkey, T.D. 1985. O2-insensitive photosynthesis in C3 plants. Its occurrence and a possible explanation. Plant Physiol. 78:71--75.
Sharp, R.E., M.A. Matthews and J.S. Boyer. 1984. Kok effect and the quantum yield of photosynthesis. Light partially inhibits dark res-piration. Plant Physiol. 75:95--101.
Sheriff, D.W., E.K.S. Nambiar and D.N. Fife. 1986. Relationships between nutrient status, carbon assimilation and water use effi-ciency in Pinus radiata needles. Tree Physiol. 2:73--88.
Sprugel, D.G., T.M. Hinckley and W. Schnap. 1991. The theory and practice of branch autonomy. Annu. Rev. Ecol. Sys. 22:309--334. Stitt, M. 1991. Rising CO2 levels and their potential significance for
carbon flow in photosynthetic cells. Plant Cell Environ. 14:741--762. Thomas, R.B., C.D. Reid, R. Ybema and B.R. Strain. 1993. Growth and maintenance components of leaf respiration of cotton grown in elevated carbon dioxide partial pressure. Plant Cell Environ. 16:539--546.
Thomas, R.B. and B.R. Strain. 1991. Root restriction as a factor in photosynthetic acclimation of cotton seedlings grown in elevated carbon dioxide. Plant Physiol. 96:627--634.
Tissue, D.T., R.B. Thomas and B.R. Strain. 1993. Long-term effects of elevated CO2 and nutrients on photosynthesis and rubisco in
loblolly pine seedlings. Plant Cell Environ. 16:859--865.
Van Oosten, J.-J., D. Afif and P. Dizengremel. 1992. Long-term effects of a CO2-enriched atmosphere on enzymes of the primary carbon
metabolism of spruce trees. Plant Physiol. Biochem. 30:541--547. von Caemmerer, S. and D.L. Edmondson. 1986. Relationship between
steady-state gas exchange, in vivo ribulose bisphosphate carboxy-lase activity and some carbon reduction cycle intermediates in Raphanus sativus. Aust. J. Plant Physiol. 13:669--688.
von Caemmerer, S. and G.D. Farquhar. 1981. Some relationships between the biochemistry of photosynthesis and the gas exchange of leaves. Planta 153:376--387.
von Caemmerer, S., J.R. Evans, G.S. Hudson, and T.J. Andrews. 1994. The kinetics of ribulose-1,5-bisphosphate carboxylase/oxygenase in vivo inferred from measurements of photosynthesis in leaves of transgenic tobacco. Planta 195:88--97.
Vu, J.C.V., L.H. Allen, Jr. and G. Bowes. 1989. Leaf ultrastructure, carbohydrates and protein of soybeans grown under CO2
enrich-ment. Environ. Exp. Bot. 29:141--147.
Wang, K.-Y. 1996. Apparent quantum yield in Scots pine after four years of exposure to elevated temperature and CO2. Photosynthetica
32:339−353.
Wang, K.-Y., S. Kellomäki and K. Laitinen. 1996. Acclimation of photosynthetic parameters in Scots pine after three years exposure to elevated temperature and CO2. Agric. For. Meteorol.
82:195--219.
Wullschleger, S.D., R.J. Norby and C.A. Gunderson. 1992. Growth and maintenance respiration in leaves of Liriodendron tulipifera L. exposed to long-term carbon dioxide enrichment in the field. New Phytol. 121:515--523.
Ziegler-Jöns, A., D. Knoppik and H. Selinger. 1987. Characteristics of the CO2 exchange of wheat ears. In Progress in Photosynthesis
Research. Ed. J. Biggins. Martinus Nijhoff Publ., Dordrecht, Bos-ton, Lancaster, pp 233--236.
Ziska, L.H., B.G. Drake and S. Chamberlain. 1990. Long-term photo-synthetic response in single leaves of a C3 and C4 salt marsh species
grown at elevated CO2 in situ. Oecologia 83:469--472.
Appendix
Symbols Unit Definition
An µmol m−2 s−1 Net assimilation rate of foliage per unit leaf area (projected area)
Anc µmol m−2 s−1 Rubisco-limited rate of net assimilation
Anj µmol m−2 s−1 RuP2-limited rate of net assimilation
Asat µmol m−2 s−1 Light-saturated rate of assimilation
Ca µmol mol−1 Molar fraction of CO2 in the air inside cuvette
Cs µmol mol−1 Molar fraction of CO2 at surface of leaf
Ci µmol mol−1 Intercellular concentration of CO2
gmin mmolm−2 s−1 Minimum stomatal conductance to water vapor
g1 dimensionless An empirical coefficient
gs mmol m−2 s−1 Stomatal conductance to water vapor
gsat mmol m−2 s−1 Light-saturated stomatal conductance to water vapor
Qp µmol m−2 s−1 Incident photon flux density
Jmax µmol m−2 s−1 Maximum rate of electron transport
Kc, Ko µmol mol−1 Michaelis constants for CO2, O2
Oi µmol mol--1 Intercellular concentration of O2
Rd µmol m−2 s--1 Respiration rate in light
Rn µmol m−2 s--1 Respiration rate in dark
Rubisco Ribulose 1,5-bisphosphate carboxylase/oxygenase
RuP2 dimensionless Ribulose 1,5-bisphosphate
Vcmax µmol m--2 s −1 Maximum RuP2 saturated rate of carboxylation
Ds kPa Leaf-to-air vapor pressure difference at leaf surface
Γ* µmol mol--1 CO2 compensation point in the absence of dark respiration