di Kabupaten Karo
Identification of Physiological Race of Phytophthora infestans in Potato
Production Center in Kabupaten Karo
Rosmayati
1, G. A. Wattimena
2, Meity Suradji Sinaga
2, M. Machmud
31 Pengajar di Departemen Budidaya Pertanian, Fakultas Pertanian USU, Medan–20155
2 Pengajar Institut Pertanian Bogor,
3Peneliti Balitbang Tanaman Pangan Bogor
Diterima 23 September 2005/Disetujui 4 Februari 2006
Abstract
Phytopthora infestans species has different virulent gene r, that comprise of 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 and its combination. The causal of late blight known unstabil in genetics there for the pathogen has many different physiological races. Especially in Indonesia, where syntetic fungicide has been irrasionally used for long time periods. An experiment has been done to identified the physiological races of P. infestans that attacked potatoes in Kabupaten Karo. The analysis were done by bioassay methods using 18 differential genotype cultivar. This experiments was arranged by Randomized Complete Design with 18 genoptype and 6 locations derived from inoculum from 3 Kecamatan; Kecamatan Brastagi, Kabanjahe and Merek. The results showed that P.infestans race found in Kabupaten Karo as race 1, 2, 3, 5, 6, 8, 11, 1.2, 1.3, 2.3, dan 1.2.4. In Brastagi, Merek and Kabanjahe race 1, 2, 3, 8, 1.2, 1.3, 2.3 and 1.2.4., Race 5, 6, and 11 only in Kecamatan Merek and Berastagi. Race 7 only in Kecamatan Merek. Di Kecamatan Berastagi , Merek and Kabanjahe were found race 1, 2, 3, 8, 1.2, 1.3, 2.3 and 1.2.4., Race 5, 6, and 11 only in Kecamatan Merek and Berastagi. Race 7 only in Kecamatan Merek.
Keywords: P. infestans, physiological race, Kabupaten Karo Abstrak
Spesies Phytophthora infestans dikenal mempunyai 11 r gen virulen yaitu, r 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 dan kombinasinya. Penyebab penyakit hawar daun ini dikenal tidak stabil, sehingga mempunyai banyak ras-ras fisiologis yang berbeda. Terutama di Indonesia, di mana penggunaan fungisida yang tidak rasional dan dalam waktu yang panjang. Percobaan ini bertujuan untuk mengidentifikasi ras fisiologis P. infestans yang menyerang pertanaman kentang di Kabupaten Karo dengan metoda bioassai, menggunakan kultivar kentang diferensial. Percobaan dilakukan dengan rancangan acak lengkap. Perlakuan yaitu kombinasi 18 genotipe diferensial dan 6 lokasi asal isolat dari 3 kecamatan, yaitu Kecamatan Berastagi, Kecamatan Kabanjahe, dan Kecamatan Merek. Hasil penelitian menunjukkan bahwa Ras P.
infestans yang dijumpai di Kabupaten Karo adalah ras 1, 2, 3, 5, 6, 8, 11, 1.2, 1.3, 2.3, dan
1.2.4. Di Kecamatan Berastagi, Merek, dan Kabanjahe ditemukan ras 1, 2, 3, 8, 1.2, 1.3, 2.3 dan 1.2.4. Ras 5, 6, dan 11 hanya ada di Kecamatan Merek dan Berastagi. Ras 7 hanya di jumpai di Kecamatan Merek.
Kata kunci: P. infestans, ras fisiologi, Kabupaten Karo
Pendahuluan
Penyakit hawar daun merupakan penyakit utama yang menyerang tanaman kentang di Kabupaten Karo. Penyakit ini tidak diketahui dengan jelas saat pertama
kali masuk ke Indonesia, diduga berasal dari bibit impor dari Eropa.
Kerugian yang ditimbulkan penyakit ini dapat mencapai 100% gagal panen, terutama pada musim penghujan dengan
kelembaban udara 90% (Sangoba dan Hakiza, 1999).
Dalam upaya mengatasi kerugian yang ditimbulkan penyakit ini, petani Karo secara intensif menggunakan fungisida dengan interval penggunaan 2–3 hari sekali bahkan dengan mencampurkan beberapa fungisida dengan bahan aktif yang berbeda (Tobing, et al., 2001). Petani tidak menyadari bahwa apa yang dilakukannya, akan menimbulkan dampak negatif yang besar terhadap lingkungan. Residu fungisida mencemari hasil panen kentang dan menimbulkan tekanan seleksi yang besar pada patogen. Patogen akan menjadi resisten terhadap fungisida dan terstimulasi membentuk keragaman genetik yang besar melalui pembentuk ras-ras baru (Bradshaw, et al., 1996).
Fry dan Goodwin (1995) menjelaskan bahwa epidemi hawar daun sangat sulit dikendalikan, karena tingginya keragaman genetik P. infestans. Menurut Deahl dan Jones (1997), perubahan genetik dari P.
infestans meliputi perubahan utama dari type mating dan ketahanan terhadap fungisida.
Selain itu perubahan keragaman genetik juga dapat terjadi karena mutasi, rekombinasi somatik, heterokariosis, dan paraseksualisme (Shaw, 1991).
Deahl dan Jones (1997) melaporkan bahwa lebih dari 13 genotipe yang berbeda didapat dari 556 isolat yang dianalisis selama periode tahun 1993 sampai 1996 di Kanada.
Hasil analisis ini memperlihatkan bahwa perubahan tidak hanya terjadi di dalam populasi, tetapi juga menunjukkan bagaimana genotipe US-8 (tipe A1) meningkat dengan cepat, dan genotipe ini sangat menonjol. Penelitian yang sama juga dilakukan di Kanada dan Kolombia pada tahun 1998, genotipe yang sangat menonjol juga US-8 dan US-11 (tipe A2), sehingga penggunaan berbagai kultivar yang dilepas secara periodik, tidak tahan terhadap penyakit ini (Deahl dan Jones, 1997).
Spesies P. infestans diketahui mempunyai 11 r gen virulen (r1– r11) dengan rekombinan- rekombinannya. Gen tahan pada inang berupa R dominan (R1–R11) telah dimasukkan ke dalam kentang budidaya, berasal dari tipe kentang liar Solanum demissum (Wastie, 1991).
Flor (1955) mengemukakan bahwa gen virulen patogen yang berkoresponden dan kompatibel dengan gen tahan pada inang, mengakibatkan tanaman inang menjadi rentan, sebaliknya bila gen virulen patogen bertemu dengan gen tahan inang yang tidak kompatibel mengakibatkan tanaman inang berespons tahan. Konsep Flor ini menunjukkan bahwa bila tanaman yang mengandung gen tahan R1 dapat terinfeksi oleh patogen, maka patogen yang menyerang tanaman tahan ini mengandung ras 1 atau kombinasi gen patogen yang mengandung ras 1, sebaliknya bila tanaman inang tidak terinfeksi patogen, maka patogen yang menyerang tanaman ini bukanlah patogen yang mengandung ras 1 (Van der Plank, 1984)
Banyak usaha sudah dilakukan untuk mengendalikan penyakit ini, salah satunya dengan menggunakan tanaman tahan, tetapi ketahanan tanaman inang mudah terpatahkan karena kehadiran ras patogen yang berbeda dengan gen tahan pada inang. Sampai saat ini belum diketahui jenis ras yang ada pada areal penanaman kentang di Kabupaten Karo. Identifikasi ras patogen perlu dilakukan untuk pengendalian penyakit hawar daun yang lebih ramah lingkungan, misalnya dengan menanam varietas kentang yang mengandung gen tahan yang tidak kompatibel dengan gen virulen patogen. Epidemik penyakit hawar daun bersifat polisiklik, oleh karena itu varietas tahan yang akan efektif mengendalikan penyakit ini harus bersifat tahan secara horizontal. Identifikasi ras P.
infestans dapat dilakukan secara molekuler,
biokimia, maupun bioassai.
Penelitian ini bertujuan untuk mengidentifikasi ras fisiologis P. infestans
yang menyerang pertanaman kentang di Kabupaten Karo dengan metoda bioassai menggunakan kultivar kentang diferensial.
Bahan dan Metoda
Tempat dan Waktu PenelitianPenelitian ini dilaksanakan di Laboratorium Hama dan Penyakit Tumbuhan, Fakultas Pertanian Universitas Sumatera Utara, Kebun Percobaan Tanaman Pangan Hutagadung, Kabupaten Karo. Percobaan berlangsung dari bulan September 2003 sampai Januari 2004.
Bahan Tanaman
Bahan tanaman yang diuji dalam percobaan ini adalah 18 genotipe diferensial mengandung gen tahan R yang sudah teridentifikasi, berasal dari CIP Peru yang merupakan koleksi Prof. G. A. Wattimena. Inokulum patogen P. infestans diisolasi dari daun kentang terserang hawar daun, diambil dari 3 kecamatan di Kabupaten Karo, yaitu Kecamatan Berastagi, Merek, dan Kabanjahe, dari setiap kecamatan diambil 2 lokasi untuk sampel. Daun sakit didesinfektan terlebih dahulu dengan NaOCl
konsentrasi 3%. Isolat ini kemudian dibilas dengan aquadest steril sebanyak 3 kali, selanjutnya potongan daun ditanam pada media V-8 juice agar dalam cawan petri. Isolat kemudian diinkubasikan pada suhu
150 C selama 6 hari, dan selanjutnya
dimurnikan dengan mengambil spora P.
infestans, kemudian isolat ini diperbanyak
kembali pada media V-8 jus agar.
Bibit yang digunakan merupakan hasil stek dari bibit yang telah diaklimatisasikan di rumah kasa, berasal dari bibit kecil hasil kultur jaringan. Bibit yang berumur 2 minggu hasil stek ditanam dalam polibag. Tiap polibag ditanam dua tanaman. Polibag berukuran 1 kg yang digunakan untuk tempat penanaman bibit, disusun di rumah kasa. Media tanam yang digunakan merupakan campuran tanah dan pupuk kandang 1:1. Sebagai pupuk dasar diberikan urea (5 g/tan), SP-36 (5 g/tan), dan KCl (5 g/tan). Pupuk SP-36 dan KCl diberikan sekaligus pada saat tanam, sedangkan pupuk urea diberikan bertahap yakni setengah bagian pada waktu tanam dan sisanya sebulan kemudian.
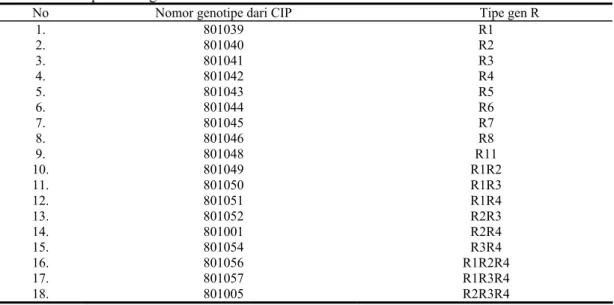
Tabel 1. Genotipe kentang diferensial asal CIP
No Nomor genotipe dari CIP Tipe gen R
1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 12. 13. 14. 15. 16. 17. 18. 801039 801040 801041 801042 801043 801044 801045 801046 801048 801049 801050 801051 801052 801001 801054 801056 801057 801005 R1 R2 R3 R4 R5 R6 R7 R8 R11 R1R2 R1R3 R1R4 R2R3 R2R4 R3R4 R1R2R4 R1R3R4 R2R3R4
Metoda Uji Ras Fisiologi P. infestans
Pengujian menggunakan metoda daun yang dipetik dan diuji di laboratorium. Pengujian dengan metoda ini lebih efektif dan efisien, karena patogen yang menginfeksi daun tanaman inang, merupakan patogen
P. infestans murni, di samping itu dapat
terhindar dari lingkungan yang tidak menguntungkan pertumbuhan patogen, serta lebih murah.
Daun-daun tanaman kentang diferensial yang akan digunakan dipetik dari tanaman pada umur 30 hari sesudah tanam, kemudian diletakkan di dalam cawan petri yang sudah diberi kertas saring basah. Selanjutnya sisi daun sebelah bawah yang menghadap ke atas diinokulasi dengan meneteskan suspensi inokulum dengan konsentrasi
2.5x104/ml sebanyak ± 0.2 ml, lalu
diinkubasikan pada suhu 200 C.
Peubah amatan dilakukan setiap hari setelah inokulasi.
Metoda Penelitian
Percobaan disusun menggunakan Rancangan Acak Lengkap (RAL). Perlakuan merupakan interaksi 18 genotipe diferensial (Tabel 1) dan 6 macam isolat, L1, L2, L3, L4, L5, L6 dengan 3 ulangan.
L1 = isolat Kecamatan Berastagi pada lokasi 1
L2 = isolat Kecamatan Berastagi pada
lokasi 2
L3 = isolat Kecamatan Merek pada lokasi
1
L4 = isolat Kecamatan Merek pada lokasi
2
L5 = isolat Kecamatan Kabanjahe pada
lokasi 1
L6 = isolat Kecamatan Kabanjahe pada
lokasi 2, sehingga menjadi 108 perlakuan.
Peubah yang diamati:
1. Periode laten (hari). Pengamatan dilakukan dengan menghitung jumlah hari sejak inokulasi sampai munculnya gejala awal penyakit.
2. Laju perkembangan bercak. Peubah ini dihitung dengan menggunakan rumus: r = Xj-Xi/tj-ti
Keterangan:
r = Laju perkembangan bercak
Xi = Diameter bercak pada pengamatan
ke i
Xj = Diameter bercak pada pengamatan
ke j
ti = Waktu pengamatan pada hari ke i
tj = Waktu pengamatan pada hari ke j
3. Diameter perkembangan bercak.
Diukur setiap hari dimulai segera setelah periode laten dapat ditentukan.
Data hasil pengamatan dianalisis ragam untuk melihat pengaruh perlakuan. Jika analisis nyata maka perbandingan pasangan antar-perlakuan dilakukan uji beda rataan dengan uji jarak berganda Duncan. Hubungan keeratan antar peubah dianalisis dengan analisis korelasi.
Hasil dan Pembahasan
Berdasarkan evaluasi dengan inang diferensial diketahui bahwa, di Kecamatan Berastagi terdapat ras 1, 2, 3, 5, 6, 8, 1.2, 1.3, 2.3 dan 1.2.4. Di antara 2 macam isolat yang digunakan untuk evaluasi lanjut diketahui ada perbedaan jenis ras asal
Kecamatan Berastagi. Ras 5 dan 6 hanya
terdapat pada lokasi 2 dan tidak pada lokasi 1. Di Kecamatan Merek ada ras 1, 2, 3, 5, 6, 7, 8, 11, 1.2, 1.3, 2.3, dan 1.2.4. Pada Kecamatan inipun ada perbedaan jenis ras antara lokasi 1 dengan lokasi 2. Pada lokasi 2 terdapat ras 7 sedangkan pada lokasi 1 tidak ada, sebaliknya pada lokasi 1 terdapat ras 2.3 sedangkan pada lokasi 2 tidak ada. Di Kecamatan Kabanjahe dijumpai ras 1, 2, 3, 5, 8, 11, 1.2, 1.3, dan 1.2.4. Ada perbedaan jenis ras pada 2 isolat yang digunakan. Isolat yang diambil dari lokasi 1
terdapat ras 5 dan 2.3sedangkan pada lokasi
terdapat di 3 Kecamatan sentra pertanaman kentang di Kabupaten Karo ini.
Periode laten adalah periode di antara inokulasi tanaman dan ekspresi gejala penyakit yang terlihat (Umaerus, 1970).
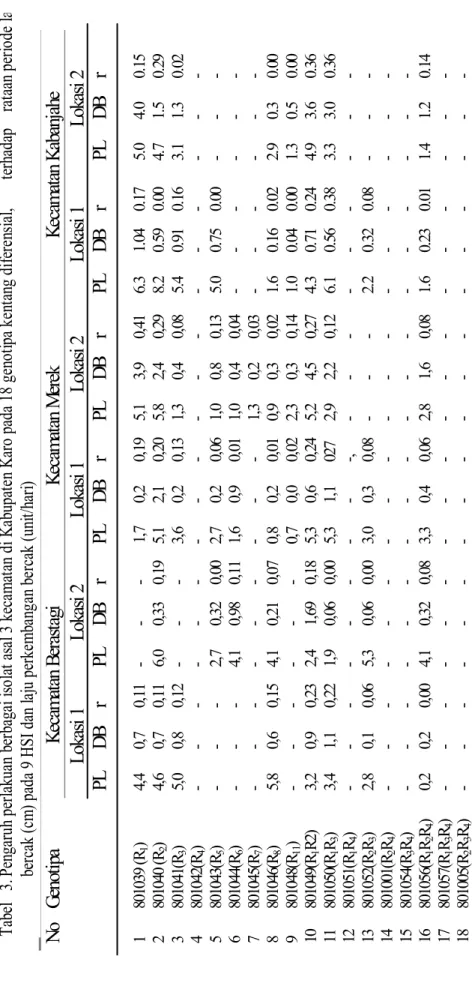
Periode laten ras 2pada genotipe 801040 di
6 lokasi yang dicobakan berkisar antara 4,6– 8,2 hari dengan rataan 5,7 hari, ras ini mempunyai periode laten yang terlama, kemudian ras 1.2 pada genotipe 801049 antara 2,4–5,3 hari dengan rataan 4,2 hari, ras 1.3 pada genotipe 801050 antara 1,9–6,1 hari dengan rataan 3,8 hari, diikuti ras 1 pada genotipe 801039 antara 1,7–6,3 hari dengan rataan 3,8 hari, ras 3 (801041) antara 1,3–5,4 hari dengan rataan 3,1 hari, ras 8 pada genotipe 801046 antara 0,8–5,8 hari dengan rataan 2,7 hari, ras 2.3 pada genotipe 801052 antara 2,2–5,3 hari dengan rataan 2,2 hari dan ras 1.2.3 pada genotipe 801056 antara 1,6–4,1 hari dengan rataan 2,2 hari. Sedangkan ras-ras yang lain periode latennya sangat kecil dengan rataan lebih kecil dari 2 hari yaitu pada genotipe 801043, 801044, dan 801045 (Tabel 3). Ras yang mempunyai periode laten panjang menggambarkan ras tersebut mempunyai tingkat virulensi yang rendah, sedangkan ras yang mempunyai periode laten pendek menggambarkan tingkat virulensi ras tinggi.
Dari Tabel 3 terlihat genotipe 801039 mempunyai laju diameter bercak yang terbesar yaitu 0,41 cm/hari di lokasi 2 kecamatan Merek, diikuti genotipe 801049, 801050, 801040, sebesar 0,27 sampai 0,36/hari, sedangkan genotipe 801041, 801043, 801044, 801045, 801046, 801048, 801052, dan 801056 mempunyai laju perkembangan bercak antara 0,01 sampai 0,18 unit/hari.
Peubah kejadian penyakit yang diamati dari hari ke-5 sampai ke-9 sesudah inokulasi, memperlihatkan pengaruh yang nyata karena perlakuan macam isolat dan 18 genotipe diferensial. Beberapa genotipe diferensial yang tidak terserang patogen di antaranya, 801042 (R4), 801051 (R1R4), 8010001 (R2R4), 801054 (R3R4), 801057 (R1R3R4), dan 801005 (R2R3R4), sedangkan
untuk genotipe 801040 (R2), 801041 (R3)
di lokasi 1 Kecamatan Berastagi pada hari ke-5 sudah 100% terinfeksi demikian juga dengan genotipe 801049 (R1R2) dilokasi 2 Kecamatan Kabanjahe. Pada hari ke-6 sesudah inokulasi, genotipe 801049 (R1R2) sudah hampir semuanya terinfeksi kecuali di lokasi 2 Kecamatan Berastagi dan lokasi 1 di Kecamatan Merek. Genotipe 801039 (R1) di lokasi 2 Kecamatan kabanjahe dan genotipa 801050 (R1R3) di lokasi 1 Kecamatan Berastagi pada hari ke-6 sudah terinfeksi 100%.
Tabel 2. Gen tahan pada genotipe diferensial yang kompatibel dengan gen virulen P. infestans berasal dari 3 kecamatan di sentra pertanaman kentang (Kecamatan Berastagi, Merek, dan Kabanjahe)
Genotipe dgn Gen R Berastagi
L1 L2 Merek L3 L4 Kabanjahe L5 L6 801039 (R1) 801040 (R2) 801041 (R3) 801042 (R4) 801043 (R5) 801044 (R6) 801045 (R7) 801046 (R8) 801048 (R11) 801049 (R1R2) 801050 (R1R3) 801051 (R1R4) 801052 (R2R3) 801001 (R2R4) 801054 (R3R4) 801056 (R1R2R4) 801057 (R1R3R4) 801005 (R2R3R4) + - + + + - - - - + - + - - + + - - + + + + - - + + - - - - + + - - - - + + + + + + - - + + + + - + + + + + + + + + - - + - - - - - + + - - - - + + + + + + - - + - - - - - + + + + + + + + - - + - - - - - + + - - - -
K
ecam
at
an
B
er
ast
ag
i
K
ecam
at
an
M
erek
K
ecam
at
an
K
ab
an
jah
e
Lo
ka
si 1
Lo
ka
si 2
Lo
ka
si 1
Lo
ka
si 2
Lo
ka
si 1
Lo
ka
si 2
No
G
en
ot
ip
a
PL
D
B
r
PL
D
B
r
PL
D
B
r
PL
D
B
r
PL
D
B
r
PL
D
B
r
1 8010 39 ( R1 ) 4, 4 0, 7 0, 11 - - - 1, 7 0, 2 0, 19 5, 1 3, 9 0, 41 6. 3 1. 04 0. 17 5. 0 4. 0 0 .15 2 8010 40 (R2 ) 4, 6 0, 7 0, 11 6, 0 0, 33 0, 19 5, 1 2 ,1 0, 20 5, 8 2, 4 0, 29 8. 2 0. 59 0. 00 4. 7 1. 5 0 .29 3 8010 41( R3 ) 5, 0 0, 8 0, 12 - - - 3, 6 0 ,2 0, 13 1, 3 0, 4 0, 08 5. 4 0. 91 0. 16 3. 1 1. 3 0 .02 4 8010 42( R4 ) - - - - - - -- -- - - - - - -5 8010 43( R5 ) - - - 2, 7 0, 32 0, 00 2, 7 0 ,2 0, 06 1, 0 0, 8 0, 13 5. 0 0. 75 0. 00 - - - 6 8010 44( R6 ) - - - 4, 1 0, 98 0, 11 1, 6 0 ,9 0, 01 1, 0 0, 4 0, 04 - - - - - - 7 8010 45( R7 ) - - - - - - -1, 3 0, 2 0, 03 - - - - -8 -8010 46( R8 ) 5, 8 0, 6 0, 15 4, 1 0, 21 0, 07 0, 8 0 ,2 0, 01 0, 9 0, 3 0, 02 1. 6 0. 16 0. 02 2. 9 0. 3 0 .00 9 8010 48( R11 ) - - - - - - 0, 7 0 ,0 0, 02 2, 3 0, 3 0, 14 1. 0 0. 04 0. 00 1. 3 0. 5 0 .00 10 8010 49( R1 R2 ) 3, 2 0, 9 0, 23 2, 4 1, 69 0, 18 5, 3 0 ,6 0, 24 5, 2 4, 5 0, 27 4. 3 0. 71 0. 24 4. 9 3. 6 0 .36 11 8010 50( R1 R3 ) 3, 4 1, 1 0, 22 1, 9 0, 06 0, 00 5, 3 1 ,1 02 7 2, 9 2, 2 0, 12 6. 1 0. 56 0. 38 3. 3 3. 0 0 .36 12 8010 51( R1 R4 ) - - - - - - -, - - - - - - - -13 8010 52( R2 R3 ) 2, 8 0, 1 0, 06 5, 3 0, 06 0, 00 3, 0 0 ,3 0, 08 - - - 2. 2 0. 32 0. 08 - - - 14 8010 01( R2 R4 ) - - - - - - -- -- - - - - - -15 8010 54( R3 R4 ) - - - - - - -- -- - - - - - -16 8010 56( R1 R2 R4 ) 0, 2 0, 2 0, 00 4, 1 0, 32 0, 08 3, 3 0 ,4 0, 06 2, 8 1, 6 0, 08 1. 6 0. 23 0. 01 1. 4 1. 2 0 .14 17 8010 57( R1 R3 R4 ) - - - - - - -- -- - - - - - -18 8010 05( R2 R3 R4 ) - - - - - - -- -- - - - - --Ta
be
l 3.
Pe
ng
ar
uh
pe
rla
kua
n
be
rb
ag
ai
is
ol
at
a
sa
l 3
ke
ca
m
at
an
d
i K
ab
upa
te
n
K
ar
o p
ada
18
ge
no
tip
a k
en
ta
ng d
ife
re
ns
ia
l,
te
rh
ad
ap
r
at
aan
p
er
io
de
la
be
rc
ak
(c
m
) pa
da
9
H
SI da
n
la
ju
pe
rke
m
ba
ng
an
b
er
ca
k (
un
it/
ha
ri)
0.00 10.00 20.00 30.00 40.00 50.00 60.00 70.00 80.00 90.00 100.00
5 hsi 6 hsi 7 hsi 8 hsi 9 hsi
Waktu Pengamatan (hari)
Kej adi an Penya ki t (% ) 80-39 80-40 80-41 80-42 80-43 80-44 80-45 80-46 80-48 80-49 80-50 80-51 80-52 80-01
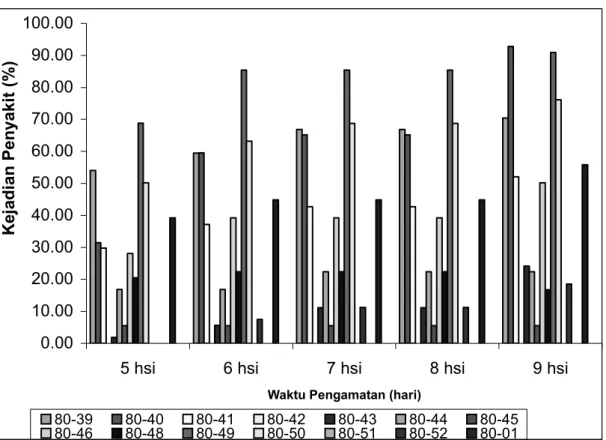
Gambar 1. Rataan kejadian penyakit pada setiap genotipa diferensial selama 9 hari pengamatan. hsi = hari sesudah inokulasi
Kejadian penyakit akan semakin meningkat dengan bertambahnya hari. Pada hari ke-9 sesudah inokulasi, genotipe 801040 (R1) serangan patogen sudah mencapai antara 78,5–100% untuk semua lokasi diikuti genotipe 801049 (R1R2) antara 56,2–100%, genotipe 801041 (R2) dan genotipe 801046 (R8) berkisar antara 33,3–100%, kecuali untuk lokasi 2 Kecamatan Berastagi genotipe 801041 tidak terserang patogen, hal yang sama juga diperoleh pada genotipe 801039 (R1). Sedangkan untuk genotipe 801050 (R1R3) serangan patogen untuk semua lokasi berkisar antara 11,1– 100%. Genotipe-genotipe 801043 (R5), 801044 (R6), 801048 (R11), 801052 (R2R3), 801056 (R1R2R4) serangan patogen berkisar antara 0–75%.
Berdasarkan semua peubah yang diamati akibat infeksi patogen pada
genotipe 801039 (R1), 801040 (R2), 801041 (R3), 801043 (R5), 801044 (R6), 801045 (R7), 801046 (R8), 801049 (R1R2), 801050 (R1R3), 801052 (R2R3), dan 801056 (R1R2R4), nampak bahwa di daerah Kecamatan Berastagi, Merek, dan Kabanjahe didapat ras 1, 2, 3, 5, 6, 7, 8, 1.2, 1.3, 2.3, dan 1.2.4, karena gen virulen patogen hanya akan menginfeksi tanaman yang mempunyai gen tahan (R) yang sama dengan gen virulen (r) yang ada pada patogen yang disebutkan dengan reaksi kompatibilitas. Sebaliknya bila tidak terjadi infeksi pada tanaman inang, maka gen virulen pada patogen tidak kompatibel dengan gen tahan (R) yang ada pada tanaman. Genotipe yang tidak mengalami infeksi sesudah diinokulasi dengan patogen
P. infestans adalah genotipe 801042 (R4),
(R3R4), 801057 (R1R3R4), 801005 (R2R3R4) berarti di daerah Kecamatan Berastagi, Kabanjahe, dan Merek tidak didapat ras 4 dengan kombinasinya kecuali kombinasi ras 1.2.4. Tingginya keragaman ras pada 3 kecamatan di Kabupaten Karo yaitu Kecamatan Berastagi, Kabanjahe, dan Merek dikarenakan pola penggunaan fungisida yang tidak memenuhi aturan dan anjuran yang sudah ditetapkan oleh pemerintah. Petani Karo dalam melakukan penyemprotan fungisida pada tanaman kentang yaitu dengan mencampur beberapa fungisida dalam satu larutan kemudian diaplikasikan ke tanaman dalam selang 2–3 hari sekali. Tingginya intensitas penggunaan fungisida ini menimbulkan tekanan seleksi yang tinggi terhadap patogen, sehingga menyebabkan patogen mengalami mutasi dan adaptasi, yang akhirnya menimbulkan keragaman ras yang besar. Di samping itu petani pada setiap kecamatan di Kabupaten Karo tidak selalu sama menggunakan jenis dan interval fungisida (hasil wawancara dengan petani), hal ini menyebabkan tekanan seleksi yang ditimbulkan berbeda-beda pada setiap kecamatan, oleh larena itu dapat dipahami bahwa jenis ras patogen pada setiap kecamatan juga berbeda. Kondisi seperti ini pernah dibuktikan oleh Suhardi (1982) dengan menggunakan 44 isolat patogen yang berasal dari Segunung, Lembang, Pengalengan, Cikajang, dan Cisurupan Jawa Barat yang diinokulasikan pada 12 genotipe tanaman kentang yang masing-masing terdiri dari r, R1, R2, R4, R5, R10, R11, R1R2, R1R3, R1R4, R1R2R3, dan R2R4 didapat bahwa terdapat ras 0, ras 1, ras 4, ras 5, ras 10, ras 11, ras 1.2, ras 1.3, ras 1.4, ras 1.2.3, dan ras 2.4. Selanjutnya Suhardi (1982) mengemukakan kebanyakan ras-ras tersebut tercampur dalam satu populasi isolat dan masing-masing isolat sering memiliki ras yang berbeda-beda. Keadaan inilah yang
mengakibatkan adanya perbedaan dalam penelitian dengan menggunakan isolat yang diambil dari lapangan. Ras 5, 6, dan 11 hanya ada di Kecamatan Merek dan Berastagi, sedangkan ras 7 hanya didapat pada Kecamatan Merek.
Pada penelitian ini, tidak dapat dideteksi ras 9 dan 10 karena tidak adanya genotipe diferensial yang mengandung gen
tahan R9 dan R10, namun kemungkinan ras
9 tidak ada karena ras 9 sangat rentan terhadap fungisida metalaksil. Sedangkan ras 10 berpeluang ada terbukti dengan
genotipe premier yang mengandung gen R10
hasil uji ketahanan di rumah kasa menunjukkan adanya sifat tahan. Untuk hal ini masih diperlukan pembuktian lanjut, apakah ketahanan pada genotipe premier karena virulensi ras 10 yang rendah atau karena ras 10 memang tidak ada.
Berdasarkan periode laten, diameter bercak, dan laju perkembangan bercak yang ditimbulkan, nampak ras 1.2 merupakan patogen yang paling virulen, diikuti oleh ras 1, 2, dan 1.3. Diameter bercak pada genotipe 801049 (R1R2) berkisar antara 3,6–4,5 cm, diikuti genotipe 801039 (R1)
(3,9–4,0 cm), 802040 (R2) (2,1–2,4 cm),
801050 (R1R3) (2,2–3,0 cm) dengan laju perkembangan bercak berkisar antara 0,15 sampai 0,36 unit/hari. Sedangkan pada genotipe 801041, 801043, 801044, 801045, 801046, 801048, 801052, dan 801056 diameter bercak yang ditimbulkan sekitar 2 cm untuk setiap asal isolat dengan laju perkembangan bercak berkisar antara 0,01 sampai 0,18 unit/hari. Berdasarkan periode laten dan diameter bercak yang ditimbulkan oleh patogen dari 6 macam isolat yang diuji, terlihat ras 1, 2, 1.2, dan 1.3 mempunyai tingkat virulensi yang tinggi. Di antara 6 macam isolat yang digunakan, isolat dari Kecamatan Merek lokasi 2 yang diambil untuk menguji tingkat ketahanan genotipe terhadap penyakit hawar daun, karena isolat
dari daerah ini mempunyai jenis ras yang paling banyak dengan tingkat virulensi yang paling tinggi untuk ras 1, 2, 1.2, dan 1.3.
Kesimpulan
Pada 6 areal pertanaman kentang di Kecamatan Berastagi, Merek, dan Kabanjahe ditemukan 12 jenis ras fisiologi
P. infestans. Di Kecamatan Berastagi, Merek,
dan Kabanjahe ditemukan ras 1, 2, 3, 8, 1.2, 1.3, 2.3, dan 1.2.4., sedangkan ras 5, 6, dan 11 hanya ada di Kecamatan Merek dan Berastagi dan ras 7 hanya dijumpai di Kecamatan Merek.
Di semua Kecamatan tidak terdapat ras 4 atau kombinasi dengan ras 4 (r1r4; r2r4; r3r4).
Ras 1, 2, 1.2, dan 1.3 merupakan ras yang paling virulen dan agresif, sedangkan ras 3, 5, 6, 7, 8, 2.3, dan 1.2.4 kurang virulen dan kurang agresif.
Daftar Pustaka
Bradshaw, J. E., H. E. Stewaar and G. R. MacKay. 1999. New approach to breeding for late blight resistance: Objectives, source, and technology.
In: Crissman L. and C.Lizarraga
(eds.). Late Blight A Threat to Global Food Security Vol 1. Proc. Global Food Initiative on Late Blight. Conf. March 16-19, 1999. Quito, Equador.
Deahl, K. L. and R. Jones. 1999. The Occurrence of Late Blight in North Amerika. In: Crissman L. and C.Lizarraga (eds.). Late Blight A Threat to Global Food Security Vol. I. Proc. Global Food Initiative on Late Bligth Conference. March 16-19, 1999. Quito, Equador.
Flor, H. H. (1955). Host-parasite interaction in Flax rust its genetics and other implications. Phytopath. Vol. 45: 680-685
Fry, W. E., S. B. Goodwin. 1995. Recent migration of Phytophthora infestans. Pp. 89–95. In Dowley LJ, Bannon E, Cooke LR, Keane T and O`Sullivan E (eds). Phytophthora infestans. Plant Physiol. 85:34–41.
Sangoba, T., J. J. Hakiza. 1999. The Current Status of Late Blight Caused by Phytophthora infestans in Africa with empasis on Eastern and Southern Africa. In: Crissman L. and C. Lizarraga (eds.). Late Blight A Threat to Global Food Security Vol. 1. Proc. Global Food Initiative on Late Bligth Conference. March 16–19, 1999. Quito, Equador.
Shaw, D. S. 1991. Genetics. In: Ingram D. S., P. H. William (eds). Advances in Plant Pathology, Vol. 7. Phytophthora
infestans, the Cause of Late Blight of
Potato. Academic Press, London. Suhardi. 1982. Beberapa Aspek Ekologi
Phytophthora infestans dan Respons
Tanaman Kentang Terhadapnya. Disertasi Program Pascasarjana IPB, Bogor. (tidak dipublikasikan)
Tobing, C., D. Bakti, Lisnawita. 2001. The research of insecticide marketing for vegetable and horticultural crops in Karo District Sumatera Utara. In: Coorporation Departement of Plant Protectin Faculty of Agriculture, University of North Sumatera with PT Pacific Chemicals Indonesia.
Umaerus, V. 1970. Studies on field resistance to Phytophthora infestans. Mechanisme of resistance and aplication to potato breeding. Z. Pflanzenzuchtg. 63:1–23.
Vanderplank, J. E. 1984. Disease Resistance in Plants. Second Edition. Academic Press, INC.
Wastie, R. L. 1991. Breeding for resistance.
In: Ingram D.S., P.H. Williams (eds)
Advances in Plant Pathology, Vol. 7.
Phytophthora infestans, the Cause of
Late Blight of Potato. Academic Press, London.