BAB IV
HASIL DAN PEMBAHASAN
4.1. Isolasi RNA Total
RNA total sengon diisolasi dengan reagen Trizol dari jaringan “xylem” batang sengon yang tua (berumur 5-10 tahun) dan bibit sengon yang berumur 3- 4 bulan. Kualitas RNA yang diisolasi dari batang yang tua tidak sebaik RNA yang diperoleh dari batang yang muda dan hampir sebagian besar terdegradasi. Degradasi RNA total ditandai dengan tidak adanya ke dua pita rRNA 28S dan rRNA 18S dan pada bagian bawah lajur migrasi RNA pada elektroforesis muncul pita tebal yang merupakan potongan-potongan RNA yang terdegradasi oleh nuklease (Lin, et. al., 1996). Karena kualitas RNA yang diperoleh akan sangat mempengaruhi optimalisasi sintesis cDNA, maka RNA yang digunakan untuk penelitian ini bersumber dan diisolasi dari bibit sengon dengan umur 3-4 bulan. Perbedaan kualitas RNA yang diperoleh kemungkinan disebabkan oleh karena aktivitas nuklease yang lebih tinggi pada batang sengon tua dibanding dengan sengon yang berumur 3-4 bulan. Selain itu, kandungan senyawa fenolik yang terdapat pada tanaman juga turut mempengaruhi tingkat keberhasilan isolasi RNA. Semakin tua umur pohon, maka kandungan zat ekstraktif, terutama senyawa-senyawa fenolik akan semakin besar. Berbeda dengan tanaman muda yang kandungan senyawa fenoliknya masih relatif sedikit, komponen ini akan mengganggu proses isolasi RNA dari batang kayu yang berumur lebih tua.
Hasil kuantifikasi RNA total dengan spektrofotometer pada panjang gelombang 260 nm menunjukkan bahwa konsentrasi RNA total yang diperoleh cukup baik dan memadai untuk sintesis cDNA. Konsentrasi RNA bervariasi dari 126 μg sampai 157 μg dengan variasi rasio OD260/OD280 antara 1.81-1.92. Nilai OD yang berada dalam kisaran range 1.8-2 menunjukkan kualitas RNA dengan kemurnian cukup tinggi (Manchester, 1996). Analisis RNA total melalui elektroforesis pada gel agarose menghasilkan integritas 2 pita RNA yang dominan dan seimbang, yaitu r RNA 28S dan 18S (Gambar 5).
1 2 3
Gambar 5. Integritas pita RNA 28S dan 18S dari RNA total Keterangan : Nomor 1-6: RNA total batang sengon umur 4 bulan 4.2. Sintesis cDNA
cDNA disintesis melalui proses transkripsi balik (riverse transcription) dengan menggunakan RNA total sebagai bahan cetakan. Primer yang digunakan untuk transkripsi balik adalah oligo d(T) sehingga hanya mRNA yang mengandung poli-A pada ujung 3’ yang akan disintesis menjadi cDNA, sedang rRNA dan tRNA yang tidak memiliki poli-A pada ujung 3’ tidak dapat membentuk cDNA. Hasil elektroforesis cDNA yang divisualisasikan dari gel agarose dibawah sinar UV menunjukkan bahwa proses sintesis cDNA berhasil dilakukan. Hal ini dapat dilihat dari kualitas pita cDNA sengon dibandingkan dengan kontrol RNA dari kit “ready to go RT-PCR” seperti pada Gambar 6.
Gambar 6. Sintesis cDNA dari sengon Keterangan :
1. Kontrol positif, 2. Kontrol negatif, 3. Hasil sintesis cDNA dari RNA 28S
18S 1 2 3 4 5 6
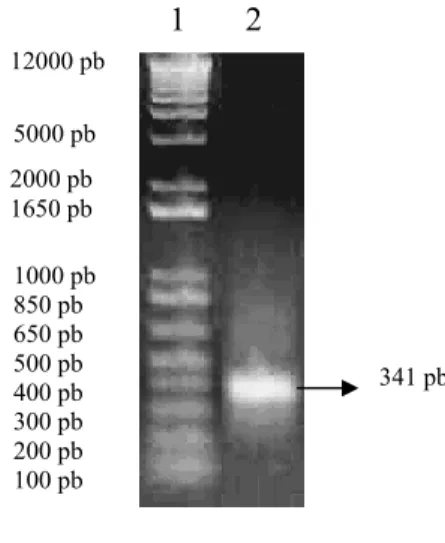
Proses PCR untuk mengamplifikasi cDNA menggunakan primer spesifik juga berhasil dilakukan dan menghasilkan fragmen dengan ukuran 341 pb. Fragmen cDNA yang berukuran 341 pb ini merupakan kandidat fragmen gen penyandi sukrosa sintase dari tanaman sengon (Gambar 7). Fragmen tersebut selanjutnya diisolasi dan dimurnikan dari gel agarose dan diintroduksikan ke E.coli.
Gambar 7. Pita hasil PCR cDNA Keterangan :
Nomor 1. Marker 1 Kb DNA plus Ladder; 2. Fragmen cDNA 4.3. Kloning Fragmen
Plasmid pGemT-Easy yang mengandung gen lac-Z digunakan sebagai vektor ligasi. Hasil ligasi fragmen sukrosa sintase dengan pGemT-Easy diintroduksikan ke bakteri E. coli galur DH5α dan disebar ke media seleksi yang
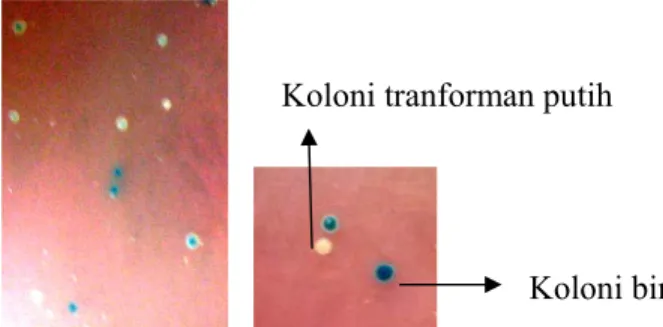
mengandung ampisilin, IPTG dan X-gal. Koloni E. coli yang mengandung transforman (plasmid rekombinan) diseleksi berdasarkan pembentukan koloni biru putih. Koloni E. coli yang berwarna biru tidak mengandung transforman, sehingga gen lacZ yang menyandi enzim β-galaktosidase masih aktif terekspresi dan mengubah substrat X-gal yang tidak berwarna menjadi biru. Koloni target adalah koloni putih yang mengandung transforman, dimana gen lacZ disisisipi oleh fragmen sukrosa sintase sehingga tidak terekspresi dan menyebabkan koloni tetap
341 pb 2000 pb 1650 pb 1000 pb 850 pb 650 pb 500 pb 400 pb 300 pb 200 pb 100 pb 12000 pb 5000 pb 1 2
Konfirmasi koloni putih dengan PCR dilakukan untuk mengetahui fragmen sukrosa sintase yang menyisip pada pGemT-Easy. PCR koloni yang menghasilkan pita DNA dengan ukuran 341 pb menunjukkan bahwa koloni putih adalah transforman yang disisipi fragmen sukrosa sintase. Ini berarti, fragmen sukrosa sintase berhasil dikloning ke vektor pGemT-Easy didalam E. coli galur DH5α.
Gambar 8. Koloni E. coli hasil transformasi berupa koloni putih 4.4. Verifikasi Sisipan dan Analisis Situs Pemotongan Enzim Restriksi
Hasil analisis peta restriksi fragmen DNA sengon menunjukkan bahwa pada fragmen tersebut tidak terdapat situs restriksi untuk enzim EcoR1 seperti pada plasmid pGemT-Easy. Hal ini tentu saja akan memudahkan pemisahan fragmen DNA sukrosa sintase sengon dari plasmid tersebut. Restriksi fragmen sukrosa sintase dengan enzim EcoR1 berhasil dilakukan. Hal dapat dilihat dari terbentuknya 2 pita spesifik, yaitu pita pada daerah 3015 pb yang spesifik untuk plasmid pGemT-Easy dan pita pada daerah 341 pb untuk fragmen sukrosa sintase (Gambar 9).
Gambar 9. Hasil restriksi fragmen sukrosa sintase dengan EcoR1; Nomor 1. Restriksi sukrosa sintase; 2. Marker 1 kb plus DNA ladder
3015 pb
341 pb
Koloni biru Koloni tranforman putih
1 2 1000 pb 850 pb 650 pb 500 pb 400 pb 300 pb 200 pb 100 pb 2000 pb 1650 pb
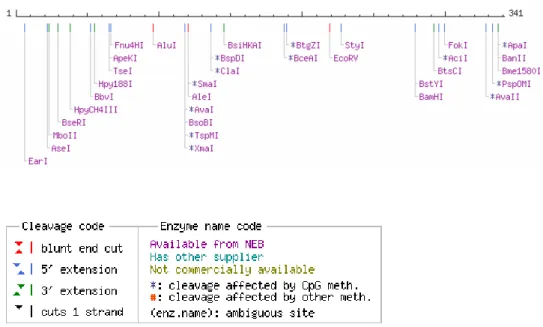
Analisis peta situs pemotongan enzim restriksi pada fragmen sukrosa sintase dibuat dengan program NEB Cutre (Gambar 10). Hasil pemetaan enzim restriksi menunjukkan bahwa fragmen sukrosa sintase tidak memiliki situs pemotongan yang sama dengan situs pemotongan seperti pada plasmid pGemT-Easy. Perbedaan situs tersebut akan memudahkan pemisahan sisipan fragmen DNA sukrosa sintase dari plasmid pGemT-easy. Peta restriksi sangat bermanfaat untuk menentukan enzim restriksi yang paling tepat untuk pembuatan kontruksi gen dan pustaka genom.
Gambar 10. Peta restriksi fragmen sukrosa sintase berdasarkan NEB Cutter
4.5. Pengurutan Fragmen Sukrosa Sintase
Hasil pengurutan fragmen Sukrosa sintase menghasilkan 341 pb nukleotida. Basa-basa tersebut menyandi 113 asam amino. Urutan basa nukleotida
9 18 27 36 45 54 5' GTG TTA AGG GTG AAG AGG AGA ATT AAT TTG TTC AAG ACT GTG ATA TTG AGA TGT --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- Val Leu Arg Val Lys Arg Arg Ile Asn Leu Phe Lys Thr Val Ile Leu Arg Cys
63 72 81 90 99 108 CCG AAA ATG GGG CTG CCG GGT GCT AAC TTG TTG TGC CAC ACA GCT TTC ACC CTT --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- Pro Lys Met Gly Leu Pro Gly Ala Asn Leu Leu Cys His Thr Ala Phe Thr Leu
117 126 135 144 153 162 CCC GTC CTC ACC CGG GTG GTT CAT GGT ATC GAT GTG CTC GAT CCC AAG TTA AAC --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- Pro Val Leu Thr Arg Val Val His Gly Ile Asp Val Leu Asp Pro Lys Leu Asn
171 180 189 198 207 216 ATT GTT CAC ACG GCG ATG TCA TGT GTA TTT ACT TCC CCT TCA AGA GAA GTT AGC --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- Ile Val His Thr Ala Met Ser Cys Val Phe Thr Ser Pro Ser Arg Glu Val Ser
225 234 243 252 261 270 AGA TAT CAG GGC CTT GGA AGT CGA TCT GGG AGA AGT TTT AGG AAG GGG AAG GAA --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- Arg Tyr Gln Gly Leu Gly Ser Arg Ser Gly Arg Ser Phe Arg Lys Gly Lys Glu
279 288 297 306 315 324 AAG AAA ATA GGA TCC GGG ATG AAA AGC GGA ACA AGG CAA ATG TTT TAT AGC AAG --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- Lys Lys Ile Gly Ser Gly Met Lys Ser Gly Thr Arg Gln Met Phe Tyr Ser Lys
333
GAA TGG ACC GGG CCC CA 3' --- --- --- --- --- -- Glu Trp Thr Gly Pro
DNA dan profil asam amino yang diperoleh dari hasil sekuensing DNA plasmid dapat dilihat pada Gambar 11 .
Gambar 11. Urutan basa nukleotida fragmen sukrosa sintase dan deduksi asam aminonya
4.6. Analisis Homologi Sukrosa Sintase
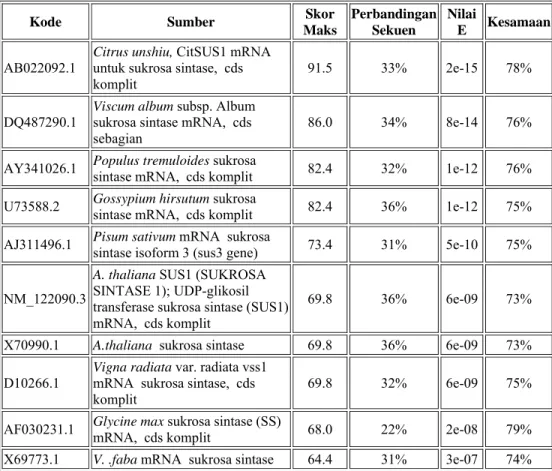
Urutan basa nukleotida fragmen sukrosa sintase dianalisis dan dibandingkan dengan database gen penyandi sukrosa sintase dari Bank DNA menggunakan program BLAST ( Basic Local Alligment Search Tool) untuk mengetahui identitas dan tingkat homologinya dengan fragmen dan gen yang telah diketahui (Tabel 2). BLAST adalah program penyejajaran urutan basa nukleotida atau asam amino berdasarkan skor tertinggi (Mount, 2001).
Tabel 2. Penyejajaran nilai kesamaan basa nukleotida sukrosa sintase dari beberapa tumbuhan (Blastn NCBI, 2007)
Kode Sumber Skor
Maks Perbandingan Sekuen Nilai E Kesamaan AB022092.1
Citrus unshiu, CitSUS1 mRNA
untuk sukrosa sintase, cds
komplit 91.5 33% 2e-15 78%
DQ487290.1 Viscum albumsukrosa sintase mRNA, cds subsp. Album
sebagian 86.0 34% 8e-14 76%
AY341026.1 Populus tremuloidessintase mRNA, cds komplit sukrosa 82.4 32% 1e-12 76%
U73588.2 Gossypium hirsutumsintase mRNA, cds komplit sukrosa 82.4 36% 1e-12 75%
AJ311496.1 Pisum sativumsintase isoform 3 (sus3 gene) mRNA sukrosa 73.4 31% 5e-10 75%
NM_122090.3
A. thaliana SUS1 (SUKROSA
SINTASE 1); UDP-glikosil transferase sukrosa sintase (SUS1) mRNA, cds komplit
69.8 36% 6e-09 73%
X70990.1 A.thaliana sukrosa sintase 69.8 36% 6e-09 73%
D10266.1 Vigna radiatamRNA sukrosa sintase, cds var. radiata vss1
komplit 69.8 32% 6e-09 75%
AF030231.1 Glycine maxmRNA, cds komplit sukrosa sintase (SS) 68.0 22% 2e-08 79% X69773.1 V. .faba mRNA sukrosa sintase 64.4 31% 3e-07 74%
Hasil BLAST basa nukleotida dengan program BLASTN menggunakan fragmen sukrosa sintase yang diperoleh menunjukkan bahwa fragmen sukrosa sintase sengon memiliki kesamaan urutan basa nukleotida dengan Glycine max, Citrus unshiu, Vigna radiata dan Arabidopsis thaliana dengan nilai kesamaan berurutan 79%, 78%, 75% dan 73%. Batas standar nilai E (expectacy) yang dapat digunakan sebagai batas terendah untuk tingkat kepercayaan pada nilai kesamaan dan homologi antar organisme adalah pangkat -4. Nilai E yang berada pada range pangkat -7 sampai -15 dari penyejajaran basa nukleotida sengon dibanding organisme lain seperti pada Tabel 2 menunjukkan bahwa tingkat homologi gen penyandi sukrosa sintase sengon cukup tinggi.
Selain BLAST berdasarkan basa nukleotida, juga dilakukan BLAST berdasarkan deduksi asam amino. Hasil BLAST asam amino yang menyandi
homologi yang mirip dengan Glycine max, Citrus unshiu, Vigna radiata dan Arabidopsis thaliana dengan nilai berurutan 54.3, 51.2, 54.3 dan 51.6. (Tabel 3) . Tabel 3. Penyejajaran deduksi protein sukrosa sintase dari beberapa
Tumbuhan (Blastp NCBI, 2007)
Kode Sumber Skor Nilai E
P13708 Glycine max 54.3 2e-06
Q01390 Vigna radiata 54.3 2e-06
ABP88869 Medicago sativa 53.9 3e-06
CAB40795.1 Medicago truncatula53.9 3e-06
CAA09910.1 Pisum sativum 53.9 3e-06
P31926 Vicia faba 53.9 3e-06
CAA50317.1 A. thaliana 51.6 1e-05
BAA89049.1 Citrus unshiu 51.2 2e-05
ABF50715.1 Viscum album 50.8 2e-05
AAD28641.1 Gossypium hirsutum 50.8 2e-05
ABB53602.1 Eucalyptus grandis 50.4 3e-05
AAR03498.1 Populus tremuloides 50.1 3e-05
Hasil BLAST dan penyejajaran menggunakan deduksi asam amino menunjukkan bahwa, gen penyandi sukrosa sintase yang diperoleh dari sengon belum komplit dan sekuen tersebut berada pada daerah tengah gen penyandi sukrosa sintase dari berbagai spesies yang telah berhasil diidentifikasi (Lampiran 2). Untuk itu, masih perlu dilakukan upaya untuk mendapatkan gen yang utuh.
4.7. Analisis Tingkat Kekerabatan
Tingkat kekerabatan sukrosa sintase sengon dibandingkan dengan sukrosa sintase dari berbagai spesies yang lain diuji berdasarkan penyejajaran deduksi asam amino. Perbandingan deduksi asam amino tersebut kemudian digunakan sebagai dasar pembuatan profil pohon pilogenetik gen sukrosa sintase dengan menggunakan program piranti DNASIS (Gambar 12). Analisis kekerabatan berdasarkan pengelompokan/kluster menunjukkan bahwa gen penyandi sukrosa sintase berada satu kluster dengan Glycine max, Vigna radiata dan lain-lain. Satu hal perlu ditelaah lebih lanjut adalah bahwa gen penyandi sukrosa sintase sengon
berada satu subkluster dengan Arabidopsis thaliana yang bukan merupakan jenis pohon berkayu. Hal ini mungkin terjadi karena gen penyandi sukrosa sintase yang diperoleh belum dalam bentuk sekuen yang utuh atau cDNA komplit.
Gambar 12. Nilai kekerabatan sukrosa sintase berbagai tumbuhan berdasarkan deduksi asam amino
4.8. Pembahasan Umum
Isolasi dan kloning gen penyandi sukrosa sintase dari sengon melalui teknik RT PCR yang telah dilakukan berhasil memperoleh 341 pb nukleotida. Fragmen cDNA yang diperoleh merupakan kandidat gen sukrosa sintase. Hal ini dapat dibuktikan berdasarkan uji tingkat homologi sekuen basa nukleotida dan asam amino fragmen sukrosa sintase yang diperoleh dibandingkan dengan gen penyandi sukrosa sintase dari organisme atau species yang lain menggunakan program BLAST.
Penelitian ini merupakan studi awal untuk memperbaiki kualitas kayu sengon melalui peningkatan deposisi selulosa. Untuk itu masih perlu dilakukan
Viscum album Citrus unshiu Gossypium hirsutum Eucalyptus di di Populus tremuloides A. thaliana Paraserianthes falcataria Medicago sativa Medicago truncatula Pisum sativum Vicia faba Glycine max Vigna radiata
sukrosa sintase kembali ke tanaman sengon. Gen yang diperoleh dari sengon masih berupa cDNA parsial. Gene establishment bertujuan untuk memperoleh pustaka cDNA yang utuh. Pustaka cDNA yang utuh dapat diperoleh melalui tahapan-tahapan seperti, penapisan pustaka cDNA dengan menggunakan fragmen sukrosa sintase sengon sebagai pelacak (probe) dan sebagai dasar desain primer untuk metode “DNA walking” seperti metode RACE (Rapid Amplified cDNA End). Keunggulan metode RACE adalah amplifikasi cDNA dapat dilakukan pada ujung 3’ atau 5’ sehingga dapat diperoleh sekuen yang utuh. Selanjutnya, gen yang diperoleh diekspresikan ke prokariot untuk melihat apakah protein sukrosa sintase tersebut secara fungsional dapat meningkatkan deposisi selulosa.
Kloning gen sukrosa sintase sengon yang telah diperoleh kembali sengon untuk peningkatan deposisi selulosa dapat dilakukan melalui dua pendekatan, yaitu kloning cDNA genomik dan kloning melalui perantaraan vektor plasmid. Kloning melalui cDNA genomik dapat dilakukan dengan cara penembakan partikel gen. Akan tetapi metode ini memiliki beberapa kelemahan, diantaranya adalah bahwa lokasi tempat gen akan menyisip bersifat random/acak sehingga tidak dapat diprediksi, jumlah “copy” besar kemungkinan banyak dan biayanya lebih mahal. Kloning melalui perantaraan vektor plasmid dapat dilakukan dengan cara transformasi ke Agrobacterium tumefaciens. Kloning melalui vektor plasmid relatif lebih menguntungkan dibanding kloning cDNA genomik karena DNA target dapat langsung menyisip ke DNA inti pada kromosom, jumlah “copy” lebih sedikit dan biayanya relatif lebih murah.