O
ne of the striking features emerging from the various plant genome sequencing projects is the extraordinary diversity of the cytochrome P450 superfamily of oxygenases in higher plants: ~300 genes are expected in the diminutive genome of Arabidopsis(http://drnelson.utmem.edu/CytochromeP450.html; http://ag.arizona.edu/p450/). It is now clear that P450s form the largest class of plant enzymes, and that several hundreds of P450 proteins are probably encoded by most plant species. Even if the number of P450 genes is not proportional to the size of each genome, this number might be larger in the case of polyploid crops. Some essential P450 functions are conserved among plant species1,2, including hormone, sterol and oxygenated fatty acid synthesis. Others, probably the majority, are involved in aspects of secondary metabolism that differ from plant to plant. Conse-quently, P450 number and substrate specificity also differs from plant to plant. This is one of the reasons for herbicide selectivity.
P450 signatures
Biochemical characteristics
P450s are heme proteins that use electrons from NADPH to cata-lyse the activation of molecular oxygen. The catacata-lysed reaction is usually a mono-oxygenation, with the formation of a molecule of water and an oxygenated product (Eqn 1), but other more atypical activities, such as dimerizations, isomerizations, dehydrations and reductions, have also been reported3,4
.
R-H 1O21NADPH 1 H 1→
R-OH 1H2O 1NADP 1
(1)
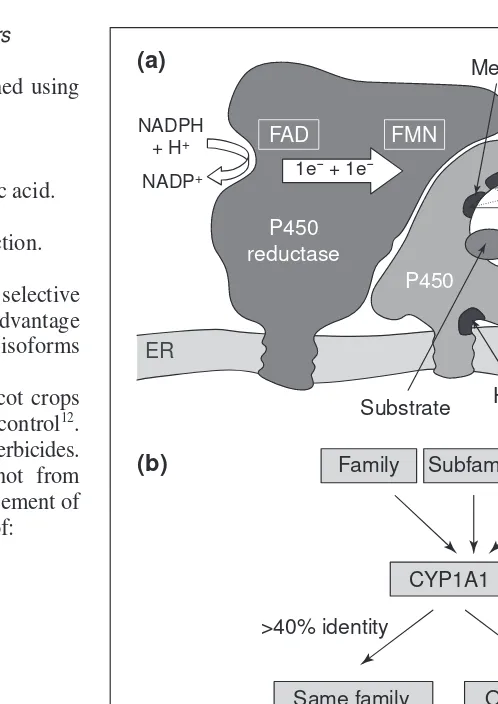
Electrons from NADPH are transferred one by one to P450s via FAD and FMN flavoproteins called cytochrome P450-reductases. Both plant P450s and their reductases are usually bound via their N-terminus to the cytoplasmic surface of the endoplasmic reticulum.
Catalysis is inhibited by carbon monoxide, which is a high-affinity ligand of the reduced enzyme and competes with oxygen. The reduced-carbon monoxide complex absorbs light at 450 nm. This is a major characteristic of P450 proteins (P450 means ‘pig-ment absorbing at 450 nm’). Carbon monoxide can be displaced by light, with a maximum efficiency at 450 nm.
Common structural features
P450 proteins have molecular masses ranging from 45 to 62 kDa, and might have as little as 16% amino acid identity. However, their overall tridimensional structure is conserved3,5
, as are a few residues
on both sides of the heme. A Phe-x-x-Gly-x-Arg-x-Cys-x-Gly motif near the C-terminus is the most conserved sequence among P450s (Fig. 1). It includes the cysteine that serves as a fifth ligand to the heme iron. Another Ala/Gly-Gly-x-Asp-/Glu-Thr-Thr/Ser consensus, located ~150 residues upstream, corresponds to the oxygen-binding and activation groove in the I helix on the distal side of the heme. A few other less conserved motifs, such as a Pro-Pro-x-Pro hinge near the N-terminus, or a Pro-Glu/Asp-Arg/His-Phe/Trp sequence between I helix and heme-binding cysteine are found in most P450 proteins.
Such conserved sequences, and their location, are considered to be signatures for the proteins deduced from P450 genes, as are the biochemical criteria for P450-dependent activities, such as: • O2-dependence.
• NADPH-dependence.
• Inhibition by carbon monoxide and reversion by light. • Endoplasmic reticulum location.
• Inhibition by antibodies directed against P450 reductases.
First hints of P450-dependent herbicide metabolism Formation of oxygenated metabolites
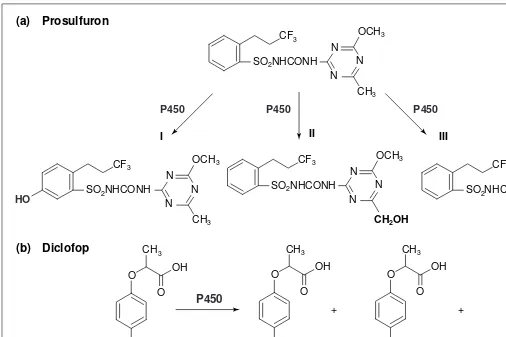
An indication that P450s might be involved in herbicide metabolism came from the analysis of herbicide residues formed in vivo. Among the major metabolites of most classes of herbi-cides are aryl- or alkyl-hydroxylated, and N-, S- or O-dealkylated products and their glucose conjugates. Dealkylation results from the oxygenation of the a carbon to the heteroatom, followed by spontaneous hydrolytic cleavage, and the elimination of alde-hyde. Most herbicides, for example prosulfuron, diclofop and chlortoluron, can be converted by P450s into several metabolites (Figs 2 and 3). In the case of the phenylurea chlortoluron, detoxi-fication of the herbicide is achieved either via hydroxylation of the ring-methyl, or via di-N-demethylation. The mono-N-demethylated product remains phytotoxic. Chlortoluron pro-vides a good example of metabolism associated with herbi-cide selectivity in weeds and crops. In the tolerant winter wheat, the half-life of chlortoluron is ,24 h, and the main metabolite is ring-methyl hydroxylated. In the susceptible weed
Alopecurus myosuroides, the main metabolite is the
mono-N-demethylated compound. Phytotoxicity of the metabolite is associated with a half-life of .24 h. By contrast, in the tolerant weed Veronica persica, the herbicide has a half-life of only 6 h, and is converted to the non-phytotoxic di-N-demethylated product6
.
Cytochromes P450 for engineering
herbicide tolerance
Danièle Werck-Reichhart, Alain Hehn and Luc Didierjean
P450 inducers and inhibitors behave as herbicide safeners or synergists
Other clues to P450 involvement in vivocan be obtained using P450 inducers (Fig. 4), such as:
• Herbicide safeners (or antidotes). • Ethanol.
• Drugs, such as phenobarbital, aminopyrine or clofibric acid. Or by using inhibitors (Fig. 4), such as:
• Compounds with a methylenedioxo or acetylenic function. • Azoles7
.
The disadvantage of such effectors is that they are usually selective activators or inhibitors of some P450 isozymes. Their advantage is that they can be used to differentiate between P450 isoforms catalysing different reactions8–11
.
Herbicide safeners are widely used to protect monocot crops from herbicide damage, and for more selective weed control12
. Some herbicide safeners bear a structural homology with herbicides. However, their protective effect appears to result, not from competition for the target site, but from a general enhancement of herbicide detoxification processes, including induction of: • P450 oxygenases.
• Glutathione S-transferases. • Glucosyl transferases. • Vacuolar transport. • Synthesis of glutathione. • Glutathione peroxidase. • Sulphate assimilation.
Some other P450 inducers, such as ethanol, phenobarbital, herbi-cides or MgCl2, have partially additive to synergistic effects when
added together with safeners8,9,13,14
. Some of these appear to be active only during specific stages of plant development9
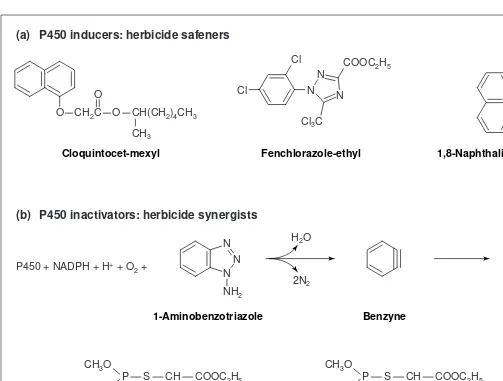
. One of the P450 inhibitors most commonly used as a synergist to characterize P450-dependent reactions in vivois 1-aminobenzo-triazole (ABT). This commercially available molecule is a mecha-nism-dependent irreversible inactivator of P450 enzymes. It shows relatively little selectivity towards P450 isoforms, especially when used at high concentration (>100 mM) because of its small size and the extreme reactivity of the benzyne, which is the product of the oxygenation reaction15
. Synergism between organophosphate insecticides (e.g. malathion or terbufos and its metabolites) and her-bicides is also an indication of the involvement of P450 in herbicide tolerance, because such insecticides behave as mechanism-based inhibitors of some P450 enzymes16–18
.
Weed resistance
Resistance might result from increased metabolism
In the past ten years, an increasing number of herbicide-resistant weed populations have been reported, sometimes showing multiple-or cross-resistance to herbicides with different chemistries and target sites. Weeds with several mechanisms of resistance are usually allog-amous species: resistance mutations accumulate in a single plant as a result of cross-pollination. Resistance mechanisms depend on the weed biotype. They are often related to mutations of target sites, but can also result from increased metabolism, or from both. An increase in P450-dependent metabolism was first demonstrated in resistant Lolium rigidumbiotypes from Australia and Alopecurus myosuroides biotypes that appeared in Europe19,20
. More recently, phenylurea-resistant populations of Phalaris minor in Asia have been described21,22
. Because the isolation of active microsomal fractions from weeds is difficult, evidence for P450 involvement has been pro-vided mainly by in vivoexperiments using inhibitors. In most cases, resistance appears to result from an increase in metabolism that can also be detected in the susceptible biotype. Polar products formed by resistant weeds are the same as those isolated from the tolerant crops:
thus, similar P450s appeared to be involved. From the differential inhibition of the formation of some metabolites, it was assumed that several P450s catalysed different reactions on the same herbicide, or even sometimes the same reaction on a single molecule6,11,21
. Studies performed on resistant Loliumprovided the first evidence that metabolism of herbicides can be induced by light23
.
Fig. 1. Structure and nomenclature of plant cytochromes P450 (P450s). (a) Plant P450s and P450 reductases are anchored to the endoplasmic reticulum (ER) via their N-termini. Recent data suggest that a portion of the globular part of P450 proteins also interacts strongly with the membrane. Domains that are highly conserved in most P450s are shown in black. These include: the loop that cova-lently binds heme via the thiolate of a cysteine (Phe-x-x-Gly-x-Arg-x-Cys-x-Gly); the portion of the I-helix involved in oxygen activation and the transfer of protons to the active site (Ala/Gly-Gly-x-Asp/Glu-Thr-Thr/Ser); a structurally highly conserved region probably impor-tant for heme-binding called the ‘meander’ (Pro-Glu/Asp-Arg/ His-Phe/Trp)5
; and the proline-rich region that forms a hinge between the membrane-anchored N-terminal helix and the globular part of the protein. A few residues interacting with the reductases are also con-served. Most other regions are highly variable, especially those involved in substrate recognition. There are at least two different P450 reductases in higher plants. They channel the electrons from NADPH to the P450 proteins by transferring them from FAD to FMN. (b) The nomenclature of P450 genes is based on the amino acid identity among the proteins they encode. Plant P450 genes have been num-bered in chronological order depending on their date of submission to the P450 nomenclature committee (http://drnelson.utmem.edu/ CytochromeP450.html). Plant P450 families bear numbers from
CYP71to CYP99, and from CYP701and above. In August 1999, 417 plant P450 genes distributed among 48 families were recorded.
Trends in Plant Science P450
Same family Other family
Same subfamily Other subfamily
>40% identity <40% identity Subfamily
How do weeds become rapid metabolizers?
The mechanism of acquiring increased metabolism remains to be determined. Some ultra-rapid metabolizer phenotypes in humans have been shown to result from the duplication of drug-metab-olizing P450 genes24
. A point-mutation in a structural gene is also possible. Another plausible explanation is the altered regulation of a stress-response pathway, in which one or more P450s are induced. In insects, constitutive overexpression regulated in trans is the only mechanism of P450-mediated resistance to an in-secticide reported to date25
. Recent data on the regulation of glu-tathione transferases suggest that a similar mechanism might occur in resistant weed biotypes26
, which would explain some cases of cross-resistance.
In vitro assays
Direct proof of the involvement of plant P450s in herbicide metabolism was obtained as early as 1969 (Ref. 27), when it was shown that the phenylurea monuron was actively dealkylated in cotton microsomes. Together with kaurene hydroxylase, this was one of the first two P450-dependent reactions characterized in higher plants. This work was confirmed with other phenylurea 20 years later6
, and started a more systematic exploration of the importance of P450s in the metabolism of other classes of
herbicides. This work, mostly performed on major crop plants10,28,29
, confirmed the main role of P450s in the oxidation of most classes of herbicides.
In vitroassays allowed more systematic enzymologic studies, confirming the mode of action of some inhibitors16,30
, and showed that P450s with catalytic parameters that were different from those of the constitutive forms (i.e. often with a higher Km) were induced
by chemical treatments8,9
. Several P450 isoforms thus appeared to participate in the metabolism of a single herbicide in the same plant. Assays performed with plant microsomes showed that herbicide-metabolizing activities were almost always induced in much larger proportions than the total P450 content. This indicated that the detoxification pathways involve P450 isozymes that are relatively minor, and hence difficult to purify compared with con-stitutively expressed enzymes. However, the question remained: whether a few specialized P450s were responsible for herbicide metabolism, or if agrochemicals are fortuitous substrates of iso-zymes involved in physiological pathways? Data13
suggesting that inducers and inhibitors have similar effects on diclofop and lauric acid hydroxylases in wheat microsomes, and that both substrates are mutual competitive inhibitors have provided support for the second hypothesis. In this case, the herbicide and the biological substrate appear to be metabolized by the same P450 protein.
Fig. 2. Examples of cytochrome P450-catalysed oxygenations of herbicides. (a) Different reactions can be catalysed by P450 on a single herbi-cide molecule. In wheat, prosulfuron is metabolized via phenyl-ring hydroxylation, alkyl hydroxylation and O-demethylation of the triazine29
. It is not yet known if these different reactions are catalysed by a single P450 or different isoforms, although metabolism in wheat, maize and sorghum appear to involve different P450s. (b) Metabolism of diclofop is an example of what is usually called an ‘NIH shift’. The oxidative attack of halogenated phenyl rings by P450 enzymes often results in a hydroxylation with simultaneous migration of the halogen atom to an adjacent position. One explanation proposed for this migration is the formation of an intermediate arene oxide.
Trends in Plant Science
I II III
CF3
SO2NHCONH
CF3
SO2NHCONH
N N N
OCH3
CF3
SO2NHCONH N
N N
OCH3 CH3
N N N
OCH3
CH3 CH2OH
HO
CF3
SO2NHCONH N
N N
OH
CH3
O O
CH3
OH
O
Cl Cl
O O
CH3
OH
O
HO Cl
Cl O
O CH3
OH
O
HO Cl
P450
P450 P450
P450
Cl
+ +
(a)
(b)
Prosulfuron
Diclofop
O O
CH3
OH
O
Cl Cl
P450 genes for engineering herbicide tolerance Bacterial genes
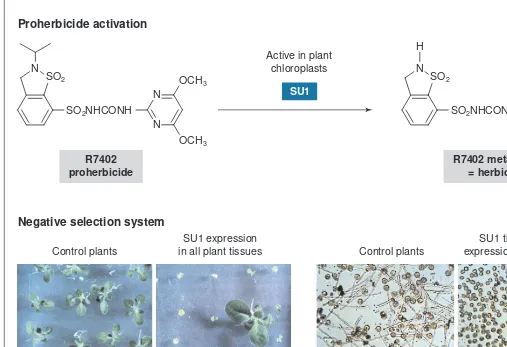
Once the involvement of P450s in herbicide selectivity had been demonstrated, the isolation of their genes became a commercial goal for the control of herbicide tolerance in crops and weeds, and to help the optimization of new active compounds. The purification of minor plant P450s being difficult, the first genes encoding herbicide-metabolizing P450s were not isolated from plants, but from bacteria and mammals. The genes of two inducible sulfonyl-urea metabolizing P450s, SU1and SU2, were isolated from the soil bacterium Streptomyces griseolus. These bacterial P450s are solu-ble enzymes that need two electron transporters, a reductase and a ferredoxin, to obtain reducing equivalent from NAD(P)H. A chloroplast transit sequence was added to the SU1 (CYP105A1) gene to target the protein to the chloroplast stroma, a compartment providing an adequate redox partner (ferredoxin). It was then expressed in tobacco under the control of promoters directing ectopic expression, or a promoter specific for the tapetum31
. When the gene product is targeted to the chloroplast in this way, it acti-vates the harmless sulfonylurea R7402, which becomes a highly phytotoxic herbicide. When SU1is expressed in the whole plant, R7402 treatment results in plant death. When the gene is expressed
from a tapetum-specific promoter, R7402 treatment of immature flowers results in male sterility. Currently, the SU1/R7402 system is widely used as a negative selection marker (Fig. 5).
Another bacterial P450 gene has been isolated from a Rhodococcusstrain that is able to grow on thiocarbamates as sole carbon and nitrogen sources32
. This P450 confers on other bacteria the ability to degrade the herbicide S -ethyl-dipropylcarbamothio-ate (EPTC), and to exert a biosafening activity on inocul-ethyl-dipropylcarbamothio-ated maize seedlings: bacteria accumulated in the crop rhizosphere detoxify EPTC present in the soil, thus protecting maize from herbicide injury.
Mammalian genes
Only a few mammalian P450s have been shown to play a signifi-cant role in the metabolism of drugs and other xenobiotics. Their herbicide-metabolizing ability has been explored to create trans-genic plants. This has been achieved largely by exploiting rat CYP1A1 to generate herbicide-tolerant tobacco and potato plants33–35
. The enzyme, which was reported previously to metab-olize various planar xenobiotics, shows a low substrate specificity, metabolizing herbicides of different chemistries, including atrazine, chlortoluron and pyriminobac-methyl. In addition, CYP1A1
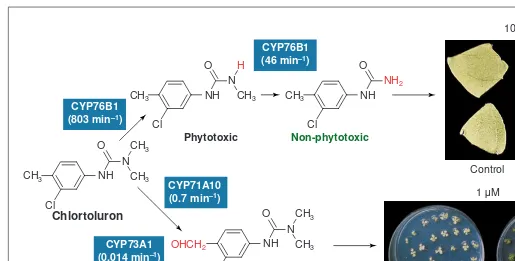
Fig. 3. Plant cytochromes P450 (P450s) metabolizing phenylurea. P450s metabolize the phenylurea chlortoluron either via ring-methyl hydroxyl-ation or via N-dealkylation. The mono-N-dealkylated product remains phytotoxic, but ring-methyl hydroxylation and di-N-demethylation lead to non-phytotoxic compounds. The genes of several plant P450s that catalyse these reactions were isolated recently. Their respective turnover rates are indicated next to each reaction in blue boxes. The CYP73A1and CYP81B1 genes were isolated from Jerusalem artichoke (Helianthus tuberosus) – their main function is phenylpropanoid and oxygenated fatty acid biosynthesis, respectively. Both proteins expressed in yeast catalyse ring-methyl hydroxylation of chlortoluron35,36
, but the efficiency of the reaction is probably too low to confer a significant resistance. CYP71A10was isolated from soybean. The recombinant protein catalyses chlortoluron ring-methyl hydroxylation at a higher rate38
and catalyses the
mono-N-dealkylation of several phenylurea. When over-expressed in tobacco, CYP71A10 increases its tolerance to chlortoluron. The lower part of the figure shows control plants and CYP71A10-transformed plants growing on a medium containing 1 mMchlortoluron. CYP76B1is a gene from Jerusalem artichoke. The protein expressed in yeast catalyses the double dealkylation of phenylurea37
. The second dealkylation is slower than the first, but the turnovers of both reactions are high, and comparable to those measured with physiological substrates. When over-expressed in tobacco, CYP76B1 also confers an increased tolerance to chlortoluron. The upper part of the figure shows excised leaf pieces of control plants and CYP76B1-transformed plants that were aged for ten days under light on a medium containing 10 mMchlortoluron. Leaf bleaching caused by the inhibition of photosynthetic electron transport by the herbicide is suppressed in the recombinant plants. Both CYP71A10 and CYP76B1 also increase resistance to linuron, which can be detoxified by a single N-dealkylation. Figure reproduced, with permission, from Ref. 39.
Control CYP76B1
Control
1 µM 10 µM
CYP71A10
CYP71A10 (0.7 min–1)
CYP73A1 (0.014 min–1)
CYP81A1 (0.28 min–1)
CYP76B1 (803 min–1)
CYP76B1 (46 min–1)
O
NH N CH3
CH3
CH3
OHCH2 Cl
O
NH N CH3
H
CH3
Cl
O
NH NH2
CH3
Cl
O
NH N
CH3
CH3
Cl Chlortoluron
Non-phytotoxic
metabolizes herbicides with low regio-specificity, catalysing both chlortoluron ring-methyl hydroxylation and N-dealkylation, as well as N-dealkylation of atrazine on two different nitrogens. To improve the enzyme efficiency, and input of reducing equivalents, two strategies have been used. The first strategy involves express-ing a fusion of CYP1A1 with the yeast P450-reductase in plants, mimicking bacterial P450s with high catalytic turnovers. The sec-ond strategy targets the fusion proteins to the chloroplast, the major site of NADPH production under light, to improve coupling of P450 reduction with photosynthetic electron transfer. The fusion protein located in the thylakoid membranes shows enhanced activ-ity under light irradiation. However, the transfer of mammalian genes into plants poses an ethical problem, in particular when they encode a protein such as CYP1A1, which has been shown to activate polyaromatic hydrocarbons into procarcinogens.
Plant P450 genes
The first plant P450s available in recombinant systems that were assayed for herbicide metabolism were enzymes involved in phe-nolic or lipid pathways36,37
. They catalyse extremely low, but detectable, chlortoluron hydroxylation. However, the kinetic con-stants of the reaction indicate that these P450s do not play a sig-nificant role in herbicide detoxification in vivo. Two plant P450s that metabolize herbicides more efficiently have been reported in the past year. Both metabolize phenylurea, but none of the other classes of herbicides tested. One gene, CYP76B1, was isolated from Jerusalem artichoke (Helianthus tuberosus) on the basis of its inducibility by Mn21
ions or drugs such as aminopyrine and phenobarbital, which also induce mammalian P450 enzymes. When expressed in yeast, together with an ArabidopsisP450 reductase, the CYP76B1 protein catalyses the dealkylation of several planar
Fig. 4. Effectors of cytochrome P450 enzymes. (a) Herbicide safeners, such as naphthalic anhydride, cloquintocet-mexyl and fenchlorazole-ethyl, are used as seed coatings or for co-treatment with herbicidal molecules to selectively increase the resistance of crops relative to that of weeds. In spite of a structural similarity to herbicides, they have no effect on herbicide target sites, but instead activate herbicide metabolism. Their effect is pleiotropic, and they increase P450s, glutathione transferases and glucosyl transferases, as well as glutathione content in the treated plants. They apparently specifically up-regulate the P450 isoforms involved in detoxification. (b) P450 inhibitors can be used as herbicide synergists. By inhibiting herbicide metabolism, they increase the proportion of toxic molecules reaching the target site. Competitive and tight-binding inhibitors are effective, but mechanism-based irreversible inhibitors are the most efficient. The metabolism of such molecules generates a reactive intermediate that forms a covalent adduct with the P450 heme or apoprotein. They are, therefore, P450-specific (although some might also inactivate some peroxidases), and they are selective for some P450 isoforms. Different classes of insecticides have been shown to behave as herbicide synergists. Some (e.g. carbamates) might be just competitive inhibitors. Others, such as organophosphates, when oxygenated by P450s, release atomic sulphur that covalently binds to the apoprotein, leading to its inactivation. When fields are treated simultaneously with a herbicide and an insecticide that are recognized by the same P450, it could lead to crop destruction.
Trends in Plant Science N
N N
Fe NH2
H2O
2N2
C
C O
O O CH2C
Cloquintocet-mexyl
1-Aminobenzotriazole Benzyne Heme adduct
Protein adduct
Fenchlorazole-ethyl 1,8-Naphthalic anhydride (NA)
Malathion
S CH2COOC2H5 (a) P450 inducers: herbicide safeners
(b) P450 inactivators: herbicide synergists CH(CH2)4CH3
CH3O
P S
CH3O CH3 O
O
Cl
Cl3C
Cl COOC
2H5
N N
N
N P450 + NADPH + H+ + O
2 +
N
N
O
S
N
CH COOC2H5
Malaoxon
O CH3O
P S
CH3O
CH COOC2H5
xenobiotics, including phenylurea, with a high turnover compa-rable to that of physiological substrates38
. Phenylurea are converted to non-phytotoxic di-N-dealkylated products. A second gene, CYP71A10, was isolated from soybean on the basis of P450-conserved sequences39
. The CYP71A10 protein expressed in yeast converts chlortoluron both to ring-methyl hydroxylated and, to a lesser extent, mono-N-demethylated compounds, as such behav-ing like rat CYP1A1. Other phenylurea were exclusively mono-N-demethylated. Catalytic turnover of CYP71A10 with phenyl-urea is more than ten times lower than that of CYP76B1, which is not so surprising considering its looser regio-specificity. Both genes confer increased tolerance to phenylurea when over-expressed in tobacco (Fig. 3).
Only preliminary data are available concerning plant P450s metabolizing other classes of herbicides. Genetic analysis has suggested that, in maize, chlortoluron and bentazon metabolism might be under the control of the same gene. A CYP72, recently
isolated from a tolerant line, appears to encode a protein capable of metabolizing both chlortoluron and bentazon, as well as chlo-rimuron and the insecticide malathion (M. Barrett, unpublished). Data concerning the turnover rates of such a multi-metabolizer P450 are not available.
Concluding remarks
It is clear that P450 differences in substrate specificity, redundance or regulation from plant to plant are responsible for herbicide tol-erance and selectivity. They are also one of the keys to herbicide cross-resistance, which is becoming more and more frequent in weeds. So far, only a few recombinant plant P450s have been examined for their ability to metabolize exogenous molecules, and those assayed have been tested for a few herbicides, and even fewer pesticides and pollutants. Recent data show that several P450 families can metabolize the same class of herbicides, some being poor metabolizers, and others being much more efficient.
Fig. 5. SU1 of Streptomyces griseolus: a cytochrome P450 that metabolizes sulfonylurea. The SU1(CYP105A1) gene was isolated from the soil bacterium Streptomyces griseolus, which metabolizes herbicides of the class of sulfonylurea. Expression of this P450 is induced by sulfonylurea and phenobarbital in the bacteria. SU1 detoxifies some phenylurea herbicides, such as chlorimuron or chlorsulfuron, but with only low effi-ciency. At a higher rate, it activates the proherbicide R7402 via N-dealkylation. This property has been used to develop negative selection sys-tems that are now widely used. To be active in plants, the bacterial protein needs to be targeted to the chloroplast where ferredoxin provides an appropriate electron donor. When the protein is expressed in all plant tissues, proherbicide treatment leads to plant death or severe growth inhi-bition. Control and SU1-transformed plants (left) were grown for 21 days on a medium containing 50 nMR7402. Selection is also possible on soil-grown plants. For example, SU1 is used to select stable insertion events after transposon mutagenesis (http://www.jic.bbsrc.ac.uk/ Sainsbury-lab/jonathan-jones/SINS-database/sins.htm). When the protein is expressed under the control of a promoter that is specifically activated in the tapetum of the anther, the treatment of immature flowers produces pollen that is unable to germinate, resulting in male sterility. Germination of the pollen of control and SU1-transformed plants seven days after R7402 treatment (right). Figure reproduced, with permission, from Ref. 31.
OCH3
Active in plant chloroplasts
OCH3
Control plants
R7402 treatment
SU1 expression in all plant tissues
Inhibition of plant growth
Control plants
SU1 tissue-specific expression in the tapetum N
N N
SO2
SO2NHCONH Proherbicide activation
Negative selection system R7402
proherbicide
SU1
OCH3
OCH3 N
N N
H
SO2
SO2NHCONH
R7402 metabolite = herbicide
R7402 treatment
This is a situation reminiscent of that observed for drug metabolism in humans – P450s showing sometimes broad, sometimes narrower, and overlapping substrate specificities. It is not clear if the poor and non-metabolizers are enzymes with essential physiological func-tions. It is not clear either, if there are P450s specialized in stress response, metabolizing a broad range of exogenous molecules, and if these P450s have a physiological function. The information needed for efficient engineering of herbicide tolerance includes the relative efficiencies of the different isoforms that are able to cata-lyse detoxification of a given molecule, their substrate specificities, and their physiological impacts.
In the case of human P450s, it is usually considered that 20 out of ~50 P450 genes play a role in the metabolism of drugs and other xenobiotics. It is possible that a similar proportion of plant genes participate in herbicide metabolism. Considering the huge number and diversity of P450 genes in the different plant species, the P450 family might provide an amazing source of catalysts for engineering pesticide tolerance by plant transformation, but also for biosafening using soil bacteria. Other possible applications, which have not been exploited to date, include positive markers for plant transformation, soil and waste-water bioremediation and green chemistry. Because P450s are active in the chloroplasts, it is possible to envisage their integration in the chloroplast genome. This would, at the same time, increase the number of copies of relevant genes per plant cell, improve the coupling of electron transfer with photosynthesis, and provide gene containment because plastid genes usually cannot be transmitted by pollen to crop-related weeds.
To date, little is known about the regulation of plant P450 genes or about the mechanisms of acquisition of herbicide tolerance. These mechanisms might be similar in tolerant crops and weeds. It is interesting to note that tolerance appears more frequent in monocot weeds and crops than in dicot plants. This might be related to the higher GC content of their genome, which would favour recombinational events and gene duplication.
Acknowledgements
We are grateful to Barbara Sears for her critical reading of the manuscript. A.H. and L.D. thank Rhône-Poulenc Agro and DuPont Agrochemicals, respectively, for financial support. This paper was written during D.W-R.’s sabbatical at the Carnegie Institution of Washington, Stanford, CA, USA.
References
1Schuler, M.A. (1996) Plant cytochrome P450 monooxygenases. Crit. Rev. Plant. Sci. 15, 235–284
2Chapple, C. (1998) Molecular genetics analysis of plant cytochrome P450-dependent monooxygenases. Annu. Rev. Plant Physiol. Mol. Biol. 49, 311–343
3Halkier, B.A. (1996) Catalytic reactivities and structure/function relationships of cytochrome P450 enzymes. Phytochemistry43, 1–21
4Mansuy, D. (1998) The great diversity of reactions catalysed by cytochromes P450. Comp. Biochem. Physiol. C121, 5–14
5Hasemann, C.A. et al. (1995) Structure and function of cytochromes P450: a comparative analysis of three crystal structures. Structure 3, 41–62
6Mougin, C.P. et al.Plant and fungal cytochromes P450: their role in pesticide transformation. In Pesticide Biotransformation in Plants and Microorganisms: Similarities and Divergences(ACS Series) (Hoagland, R.E. et al., eds), Oxford University Press (in press)
7Werck-Reichhart, D. (1995) Herbicide metabolism and selectivity: role of cytochrome P450. Proc. Brighton Crop Prot. Conf. Weeds7B-1, 813–821
8Frear, D.S. et al. (1991) Induced microsomal oxidation of diclofop, triasulfuron, chlorsulfuron, and linuron in wheat. Pestic. Biochem. Physiol.41, 274–287
9Persan, M.W. and Schuler, M.A. (1995) Differential induction of cytochrome P450-mediated triasulfuron metabolism by naphthalic anhydride and triasulfuron. Plant Physiol.109, 1483–1490
10Moreland, D.E.et al.(1995) Partial characterization of microsomes isolated from mung bean cotyledons. Pestic. Biochem. Physiol. 52, 98–108
11Preston, C. et al.(1996) Multiple resistance to dissimilar herbicide chemistries in a biotype of Lolium rigidumdue to enhanced activity of several herbicide degrading enzymes. Pestic. Biochem. Physiol. 54, 123–134
12Hatzios, K.K. (1997) Regulation of xenobiotic degrading enzymes with herbicide safeners. In Regulation of Enzymatic Systems Detoxifying Xenobiotics in Plants(NATO ASI Series 3. High Technology) (Vol. 37) (Hatzios, K.K., ed.), pp. 275–288, Kluwer
13Zimmerlin, A. and Durst, F. (1992) Aryl hydroxylation of the herbicide diclofop by a wheat cytochrome P450 monooxygenase. Plant Physiol.100, 874–881
14Frear, D.S. and Swanson, H.R. (1996) Cytochrome P450-dependent hydroxylation of prosulfuron (CGA 152005) by wheat seedlings microsomes.
J. Agric. Food Chem. 44, 3658–3664
15Ortiz de Montellano, P.R. and Mattews, J.M. (1981) Autocatalytic alkylation of the cytochrome P450 prosthetic heme group by 1-aminobenzotriazole: isolation of an N,N-bridged benzyne-protoporphyrin IX adduct. Biochem. J. 195, 761–764
16Kreuz, K. and Fonné-Pfister, R. (1992) Herbicide–insecticide interaction in maize: malathion inhibits cytochrome P450-dependent primisulfuron metabolism. Pestic. Biochem. Physiol. 43, 232–240
17Christoffer, J.T. et al.(1994) Malathion antagonizes metabolism-based chlorsulfuron resistance in Lolium rigidum. Pestic. Biochem. Physiol.49, 172–182
18Baerg, R.J. et al.(1996) Insecticide and insecticide metabolite interactions with cytochrome P450 mediated activities in maize. Pestic. Biochem Physiol. 55, 10–20
19Holt, J.S. et al. (1993) Mechanisms and agronomic aspects of herbicide resistance. Annu. Rev. Plant Physiol. Plant Mol. Biol. 44, 203–229
20De Prado, R. and Menendez, J. (1997) Cross-resistance and herbicide metabolism. In Regulation of Enzymatic Systems Detoxifying Xenobiotics in Plants(NATO ASI Series 3. High Technology) (Vol. 37) (Hatzios, K.K., ed.), pp. 351–366, Kluwer
21Singh, S. et al.(1998) Effect of ABT on the activity and rate of degradation of isoproturon in susceptible and resistant biotypes of Phalaris minorand in wheat. Pestic. Sci.53, 123–132
22Singh, S. et al.(1998) Effect of the monooxygenase inhibitor piperonyl butoxide on the herbicidal activity and metabolism of isoproturon in herbicide resistant and susceptible biotypes of Phalaris minorand wheat. Pestic. Biochem. Physiol. 59, 143–153
23Preston, C. and Powles, S.B. (1997) Light-dependent enhanced metabolism of chlortoluron in a substituted urea herbicide-resistant biotype of Lolium rigidum
Gaud. Planta201, 202–208
24Lundqvist, E. et al.(1999) Genetic mechanisms for duplication and multiduplication of the human CYP2D6gene and methods for detection of duplicated CYP2D6genes. Gene226, 327–338
25Feyereisen, R. (1999) Insect P450 enzymes. Annu. Rev. Entomol. 44, 507–533
26Cummins, I. et al.(1999) A role for glutathione transferases functioning as glutathione peroxidases in resistance to multiple herbicides in black-grass.
Plant J.18, 285–292
27Frear, D.S. et al.(1969) N-demethylation of substituted 3-(phenyl)-1-methylureas: isolation and characterization of a microsomal mixed function oxidase from cotton. Phytochemistry8, 2157–2169
28Barrett, M. (1995) Metabolism of herbicides by cytochrome P450 in corn.
Drug Metab. Drug Interact. 12, 299–316
29Frear, S. (1995) Wheat microsomal cytochrome P450 monooxygenases: characterization and importance in the metabolic detoxification and selectivity of wheat herbicides. Drug Metab. Drug Interact. 12, 329–358
30Helvig, C.et al. (1996) Selective inhibition of a cytochrome P450 enzyme in wheat that oxidizes both the natural substrate lauric acid and the synthetic herbicide diclofop. Pestic. Biochem. Physiol.54, 161–171
31O’Keefe, D.P. et al.(1994) Plant expression of a bacterial cytochrome P450 that catalyses activation of a sulfonylurea pro-herbicide. Plant Physiol. 105, 473–482
32Nagy, I. et al. (1995) Degradation of the thiocarbamate EPTC
33Ohkawa, H. et al. (1998) Molecular mechanisms of herbicide resistance. Rev. Toxicol.2, 245–252
34Inui, H. et al.(1999) Herbicide metabolism and cross-tolerance in transgenic potato plants expressing human CYP1A1. Pestic. Biochem. Physiol.64, 33–46
35Lacour, T. and Ohkawa, H. (1999) Engineering and biochemical characterization of the rat microsomal cytochrome P450 to ferredoxin-NADP(1) reductase from plant chloroplasts. Biochim. Biophys. Acta1433, 87–102
36Pierrel, M.A. et al.(1994) Catalytic properties of the plant cytochrome P450 CYP73 expressed in yeast. Substrate specificity of a cinnamate hydroxylase.
Eur. J. Biochem. 224, 835–844
37Cabello-Hurtado, F. et al.(1998) Cloning, expression in yeast and functional characterization of CYP81B1, a plant P450 which catalyses in-chain hydroxylation of fatty acids. J. Biol. Chem.273, 7260–7267
38Robineau, T. et al. (1998) The chemically inducible plant cytochrome P450 CYP76B1 actively metabolizes phenylurea and other xenobiotics. Plant Physiol. 118, 1049–1056
39Siminsky, B. et al.(1999) Expression of a soybean cytochrome P450 monooxygenase cDNA in yeast and tobacco enhances the metabolism of phenylurea herbicides. Proc. Natl. Acad. Sci. U. S. A.96, 1750–1755
R
oots, rhizomes and other plant organs in the soil usually obtain O2 for respiration directly from their immediateenvironment – the soil gaseous atmosphere. But when the soil becomes excessively wet, transfer of O2 from the air into the
soil is effectively blocked because the larger soil pores, which are usually air-filled, are water-filled instead. Without replenishment from the air, any dissolved O2remaining in the soil is quickly
con-sumed by microorganisms and plants, and the soil is then no longer able to supply O2. Aerenchyma – tissue comprising a high
proportion of gas-filled spaces or lacunae – provides the plant with an alternative strategy for obtaining O2. The interconnected
lacunae, extending from below the ground up into the stems and leaves, make up an internal aeration system, enabling parts of the plant to survive or grow for a time in environments that are O2-deficient or even completely devoid of O2.
Spaces form within aerenchymatous organs, either by cell separation at the middle lamella during development (shizogeny), or by cell death and dissolution (lysigeny). Sometimes, as in Sagittaria lancifolia, both processes occur, with lysigenous aerenchyma in the roots and shizogenous aerenchyma in the leaf petiole1
. In lysigenous aerenchyma formation in the root cortex of maize, rice and Sagittaria, cell death is first detected at a distance of a cm or less from the root apical meristem, in the zone where cell elongation has just been completed2–4
. The space created by the death of cells becomes increasingly promi-nent in older zones behind the tip. The system of gas-filled lacunae therefore develops acropetally, extending into the root towards the tip, simultaneous with the root’s extension into the soil.
Aerenchyma function
Lysigenous aerenchyma provides not only an internal pathway for O2transfer, but also simultaneously reduces the number of O2
-consuming cells, a feature that might assist in low O2
environ-ments. It has been suggested that aerenchymatous organs are often water- or fluid-filled5
, a feature that would prevent them from functioning in O2transfer. However, evidence from the
measure-ment of gas-filled porosity, microscopic observation (Fig. 1) and O2transport
6
is overwhelming – the lacunae are indeed usually gas-filled and therefore are able to transfer gases by convection and diffusion.
Aerenchyma formation is inducible by flooding in maize7,8
, in the coastal grass Spartina patens9
and in many other native species, both monocot and dicot, that occupy wetland habitats10
. By contrast, in the maize relatives Tripsacum dactyloides(eastern gamagrass) and Zea luxurians(teosinte)11
, as well as in wetland species, such as rice3,12
and S. lancifolia1,4
, aerenchyma forms con-stitutively, apparently without any requirement for an external stimulus. Eastern gamagrass possesses notable tolerance to drought and flooding, which is thought to be because of its deep rooting pattern and constitutive aerenchyma13
. The roots have been found to exceed 1.8 m in depth, penetrating a resistant clay pan at or below 0.90 m, apparently when the soil was wet and mechanically least resistant. Therefore aerenchyma was likely to have assisted oxygenation of the root at that time. In general, soil compaction can lower the oxygen concentration of the soil (fewer large, gas-filled pores for gas exchange with the atmosphere). Therefore, aerenchyma could be potentially beneficial for growth or function of roots in a densely packed soil layer. These considerations
Programmed cell death and
aerenchyma formation in roots
Malcolm C. Drew, Chuan-Jiu He and Page W. Morgan
Lysigenous aerenchyma contributes to the ability of plants to tolerate low-oxygen soil environments, by providing an internal aeration system for the transfer of oxygen from the shoot. However, aerenchyma formation requires the death of cells in the root cortex. In maize, hypoxia stimulates ethylene production, which in turn activates a signal transduction pathway involving phosphoinositides and Ca21. Death occurs in a predictable pattern, is
regulated by a hormone (ethylene) and provides an example of programmed cell death.
Danièle Werck-Reichhart*, Alain Hehn and Luc Didierjean are at the Dept of Cellular and Molecular Enzymology, Institute of Plant Molecular Biology, Centre National de la Recherche Scientifique UPR 406, 28 rue Goethe, 67083 Strasbourg Cedex, France.
*Author for correspondence (tel 133 3 88 35 83 32;