Directory UMM :Data Elmu:jurnal:T:Tree Physiology:Vol15.1995:
Teks penuh
Gambar
Dokumen terkait
Accordingly, when tree seedlings responded to increased local nutrient supply by increased root growth, concentrations of the growth-limiting nutrients (N and P in Norway spruce
Isotopic branch length coefficients calculated for genetically selected Pinus radiata trees differing in mean branch length and foliage carbon isotope composition.. Average
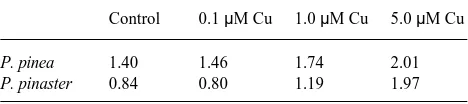
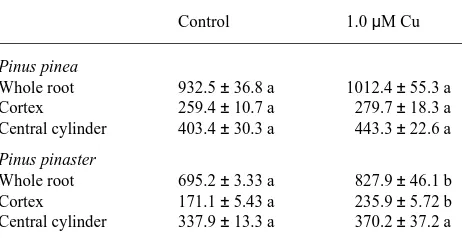
A regression analysis of plant growth was carried out with the data on degree of colonization, total root length, mean length of adventitious roots, intensity of branching of
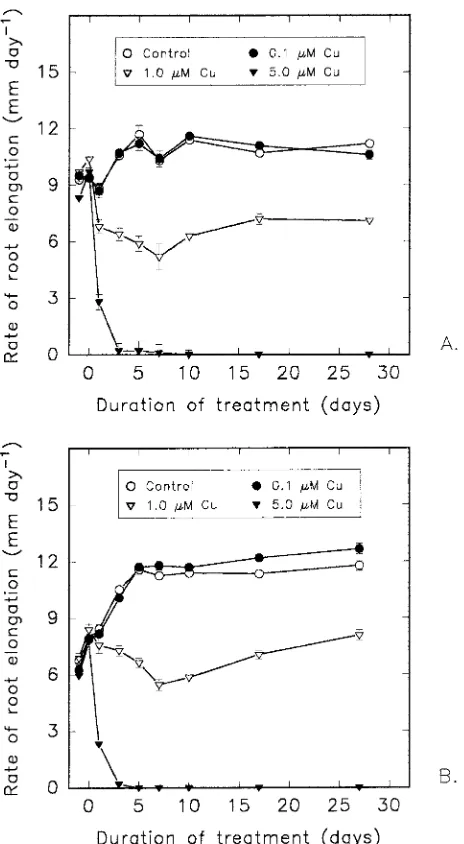
To test effects of fertilization on late-season growth and nutrient content of container-grown red pine (Pinus resinosa Ait.), seedlings were subjected to a factorial combination of
Primary leaf formation could also be induced in etiolated seedlings when the roots were exposed to a 5 min pulse of white light or red light, and the kinetics of leaf formation
(a) Biomass of fine roots (diameter 5 2 mm), and (b) ratios of leaf to fine root biomass of 2-year-old saplings of pedunculate oak, which were grown on nutrient solutions differing
Litter was not in- corporated into deep soil layers (>20 cm) and C inputs from roots were small because only 24% of the total root biomass was located below 20 cm depth.. This
