One-half of global primary production occurs in the oceans1,2, and therefore a fundamental problem for oceanographers is to understand how organisms use this carbon to create spatio-temporal patterns of carbon and energy flux. For example: how much is passed into fish3; how much is respired and returned to the atmos-phere; and how much descends to support the deep-sea biota or to be sequestered on the seafloor4. Interest in the carbon cycle has increased recently owing to global problems, such as climate change, coastal eutrophication and over-fishing5,6. Historically, oceanographers believed that most primary production moved through a chain of small and large animals7–9 and microorganisms were largely ignored10. However, several remarkable discover-ies11–13 (reviewed in REFS 14–16) that have been made during the past 30 years have shown that bacteria domi-nate the abundance, diversity and metabolic activity of the ocean (FIG. 1). A large fraction of primary production
becomes dissolved (dissolved organic matter; DOM)17 by various mechanisms in the food web, and this part of the primary production is almost exclusively accessible to heterotrophic bacteria and archaea (together referred to in this Review as bacteria)9,18,19. As a result, the uptake of organic matter by bacteria is a major carbon-flow path-way, and its variability can change the overall patterns of carbon flux8,20,21. Further, as bacteria use behavioural and biochemical strategies to acquire organic matter — for example, by the expression of enzymes to solubilize particulate organic matter (POM)22 — they interact with sources of organic matter and modify the ecosystem and carbon cycle in different ways23,24. Our aim throughout this Review is to propose a mechanistic and microspatial framework that promotes a better understanding of how bacteria regulate the biogeochemical state of the oceans.
Ecosystem-level coupling
The effects of bacteria on the carbon flux in the sea have been measured over the past 30 years25,26. Although it is impractical to measure the flux of each component of DOM into bacteria, the cumulative carbon flux (bacte-rial carbon demand; BCD) can be estimated as the sum of the carbon that is assimilated (growth) and respired, which can then be compared with primary production as a measure of the coupling strength (BCD ÷ primary production27). This parameter is useful as a global meas-ure of bacterial performance, for example, in concepts that involve the ocean carbon flux or ecosystem-based fisheries21. Measurements of coupling strength have been made on many ocean expeditions, in most oce-anic locations and in regions that differ in primary production, and this has revealed important geo-graphical and seasonal patterns. The following exam-ples show that the coupling of bacteria with primary production is highly variable and that this variability affects ecosystem functioning.
First, in the eastern Mediterranean bacteria take up most of the primary production, which is consistent with the poor fisheries that are present in this area28 (although a high BCD might be an effect rather than a cause of poor fisheries). Second, studies carried out on a north–south oceanic transect (53°N in the Atlantic to 65°S in the Southern Ocean)29 showed latitudinal vari-ation in coupling strength, and, importantly, there were large net-heterotrophic regions. Bacteria took up more DOM than was present as local primary production, which indicates that the spatial or temporal import of organic matter must have occurred. Consequently, these regions have the potential for the net out-gassing of carbon dioxide. More extensive spatial and temporal
Scripps Institution of Oceanography, University of California, San Diego, La Jolla, California 92093, USA. Correspondence to F. A. e-mail: [email protected] doi:10.1038/nrmicro1747
Primary production
The original source of organic material in an ecosystem that is due to carbon dioxide fixation by photosynthetic bacteria, plants or algae, or chemosynthetic microorganisms.
Heterotrophic
The acquisition of carbon and metabolic energy by the consumption of living or dead organic matter.
Microbial structuring of marine
ecosystems
Farooq Azam and Francesca Malfatti
Euphotic zone
Aphotic zone
Sediment
Mesopelagic processes
Aggregation DOM
DMS
Protozoa
Bacteria and archaea Viruses
Benthos POM
Phytoplankton Zooplankton
Aggregates sink Advection
CO2
hv
Benthic flux N, P, Fe, Si C, N, P, Si, Fe
Faecal
pellets Microbial loop
Fish
O2
CO2
Pathogens, pollutants and nutrient run off
Seabed
O2 O2
Predation by birds Fisheries Autotrophic
An organism that synthesizes organic carbon from the fixation of inorganic carbon, for example, by photo- or chemosynthesis.
coverage to account for patchy autotrophic processes might, however,reveal a metabolic imbalance29–31. Finally, during the summer, bacteria–DOM coupling in the Antarctic Ocean was found to be weak. Perhaps, the bacterial hydrolysis of polymeric substrates and monomer uptake was slowed owing to low tempera-ture and low substrate concentrations. This could result in the storage and temporal export of slow-to-degrade DOM in productive summers (when primary production is high) to support the energy needs of the Antarctic food web during the winter. During winter, a particle-based food web might incorporate bacterial biomass that is produced through the use of DOM32. This example illustrates that even when bacteria–DOM coupling is weak, bacteria can still be important for ecosystem functioning. It has been proposed that there is a low-temperature–low-substrate restriction in the Arctic Ocean33,34. Conversely, the excessive external input of organic carbon might have deleterious effects on system functioning. In experimental systems, bacteria outgrew coral-reef communities after the addition of DOM35 or being placed in contact with decaying macroalgae36.
These examples illustrate that the ability or inability of a bacterial assemblage to grow on specific types of organic matter, either owing to the constraints of com-munity composition or environmental gene expres-sion, can be important for the functioning of globally significant ecosystems. This underscores the need to
understand how bacteria function in their natural envi-ronment to influence the flux pathways of fixed carbon. From advances in marine genomics and metagenom-ics37–42 it can be inferred that enormous bacterial gene diversity is available for assemblage-level bacterial interactions with the ocean. Environmental genomics and proteomics are also yielding insights into bacte-rial adaptive strategies to ocean life. The challenge is to determine how these strategies are used by bacteria in natural ecosystems, which will require the explora-tion of the ocean at the micrometre scale — the scale at which the adaptive strategies of bacteria structure marine ecosystems. Bacteria do more than simply cycle carbon; they interact with the whole ocean ecosystem intimately in a multitude of ways43.
Microscale interactions
Spatial distribution of microorganisms. Most oceano-graphic studies assume that bacteria take up homoge-neously distributed DOM. This premise relies on the assumption that the DOM that is released from any source diffuses homogenously into the bulk phase before it is taken up by an organism. This view is changing, however, with the recognition of bacterial
in situ behavioural and physiological responses to DOM-production loci and gradients. The detection of high abundances of decomposer bacteria (106 per ml11) has led to the suggestion that the numbers and activity of primary producers (such as cyanobacteria and algae),
Pelagic
Relating to or occurring in the oceanic water column.
Oligotrophic
An aquatic environment that has low levels of nutrients and primary production (for example, high mountain lakes or the open ocean).
Eutrophic
A marine or lake environment with a high nutrient concentration and high levels of primary production.
Phytoplankton
Composed of microscopic plants and photosynthetic cyanobacteria. These are the main primary producers in marine food webs, ranging in size from 1 µm to approximately 100 µm.
decomposers (such as bacteria) and predators (such as viruses and protists) are similar (Fig.1). Typically, 1 mm3
(or µl) of surface seawater — considered in this Review to be the bacterium’s microenvironment — contains 10,000 viruses; 1,000 bacteria; 100 Prochlorococcus
cells; 10 Synechococcus cells; 10 eukaryotic algae; and 10 protists, although the numbers are highly vari-able16,18,44. The close proximity (0–1 mm) of individual cells suggests that there is potential for many cell–cell interactions.
We note that all microbial trophic groups are in close proximity, so that the adaptive biology of bacteria occurs in a microspatial context that integrates the interactions between all of the trophic groups. For instance, bacteria might respond behaviourally and metabolically to the DOM that is produced by a range of mechanisms, such as algal exudation and cell lysis18, predation by viruses, which releases prey DOM45, and the action of protists, which egest food vacuoles that contain DOM and regen-erated N, P and Fe2+(REFS 46,47). Such DOM hot spots might occur in the microenvironment of a bacterium over short timescales, perhaps even minutes, owing to the high abundance of microorganisms.
Motility. Motility and sensing enable bacteria to adapt in environments that contain DOM gradients. Dark-field microscopy has revealed that motility is common in natural assemblages of marine bacteria48–50, although the fraction of bacteria that are able to swim ranges from 5 to 70%. Motility might enable bacteria to achieve spatial coupling with a DOM source, such as a living or dying alga, or a protist51,52.
Hydrolytic enzymes and hydrolysis-uptake coupling. Marine bacteria hydrolyse polymers and particles using cell-surface-bound hydrolytic enzymes or ecto-hydrolases (protease, glucosidase, lipase, phosphatase, nuclease and chitinase)22,53–59. These enzymes, together with membrane-bound transporters, make the bacte-rial surface reactive for organic matter transformation and uptake. This is the final step that couples bacteria to primary production in the ocean. The bacterial surface is the dominant biotic surface, and it is proposed that 0.1–1 m2 of the bacterial surface per m3 ofseawater interacts intimately with DOM60. A pelagic bacterium that swims through organic matter leaves a stream of monomers and oligomers behind it. This led J. Stern to refer to bacteria as the “perfect swimming stomachs”
(REF. 21).
Bacteria express multiple (multiphasic) transport-ers that have Km values that range from nanomolar to millimolar, consistent with adaptation to environments that contain DOM gradients61–64 (FIG. 2). This raises the question of whether the distinction that is made between oligotrophic and eutrophic bacteria, which is usually based on culturing studies, actually reflects the microspatial adaptations of different strains to nutrient hot spots. Perhaps, bacteria simply move to an appro-priate distance from a DOM gradient, so that they can effectively use the carbon resources that are available. The in situ behaviour of strains — such as the widely distributed oligotrophic Candidatus Pelagibacter ubique (SAR11)65,66 bacteriumor the high-nutrient-loving Roseobacter-clade members — when viewed in such a microspatial context could lead to new predictions about their ecology and distribution67,68.
A biochemical mechanism that tightly couples the transport and hydrolysis of organic matter would be a useful adaptation in ocean environments, where dif-fusion rates are high69(FIG. 2). The existence of such a coupling mechanism has been difficult to demonstrate experimentally, but genomic, metagenomic and pro-teomic data might provide hints as to how bacteria obtain sufficient nutrients in an environment such as the ocean.
A bacterium’s-eye view of organic matter
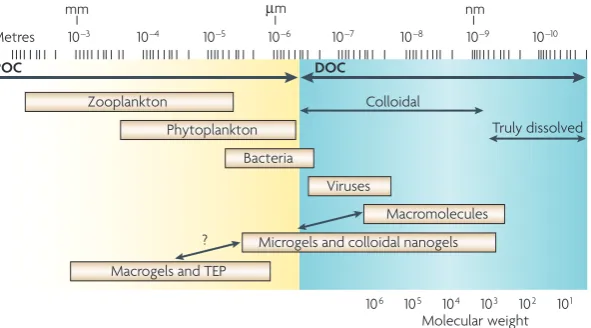
In a sense, microbial oceanography has a long history of studying bacteria at the microscale, including studies on the physiology and growth performance of bacteria that are present in seawater, attached to a particle or clus-tered around algae and detritus. Traditional distinctions between DOM and POM have been based on filtration methods that have used filters with a pore size of 0.45 µm. However, recent research shows that organic matter in seawater is replete with transparent gels that form tan-gled webs of components in the form of colloids that are approximately 10 nm long (108 colloids per ml70,71) and mucus sheets and bundles that are up to 100 µm long (1,000 sheets or bundles per ml72,73). Colloids, sheets and bundles interact to form macromolecular networks that are 100 µm or more long. The polymeric compo-nents of transparent gels are probably derived from microorganisms. Phytoplankton and bacteria produce Zooplankton
Phytoplankton
Bacteria
Viruses
Macromolecules
Microgels and colloidal nanogels Colloidal
Truly dissolved
Macrogels and TEP
Molecular weight
mm µm nm
Metres 10–3 10–4 10–5 10–6
106 105 104 103 102 101
10–7 10–8 10–9 10–10
POC DOC
?
Figure 2 | The size range of organic matter and microbial interactions in the ocean. Organic matter has traditionally been divided into dissolved organic matter and particulate organic matter, based on filtration. Here, the full size range of organic matter is shown, from monomers, polymers, colloids and gel particles to traditional particles (divided into particulate organic carbon (POC) and dissolved organic carbon (DOC)). These organic nutrient pools form an intricate three-dimensional architecture of polymers, colloids, gel particles and large aggregates. Living plankton and dead organisms are thought to be embedded in a cobweb-like structure of organic matter that is dynamic over time and space (not shown). The organic matter architecture provides the spatial context within which microorganisms — bacteria, archaea, microalgae, protists and viruses — interact with each other and with the environment. TEP, transparent
NH4
PO4
Bacterium Polymer
O2 CO2
Chemoreceptor Permease
POM
Direct substrate
Ectoenzyme
Phytoplankton and detritus
cell-surface mucus, which can either be released or, alternatively, solubilized by cell-surface-acting enzymes. Phytoplankton release condensed polysaccharide parti-cles by exocytosis, which form gels in seawater74. Most (90–95%) of the DOM is refractory to degradation by bacteria and it has been proposed that some DOM com-ponents serve as stable scaffolds of a gel architecture that is inaccessible to bacterial ectohydrolases75. Potentially, gels are a nutrient sink and represent a mosaic of nutrient hot spots. This nutrient pool is huge, as approximately 10% of all the DOM (70 × 1015 grams of carbon) that
is found in the ocean is present in the form of gels. This represents a carbon pool that is larger than all of the car-bon that is present as biomass in the ocean72. Depending on their surface properties, gels might adsorb DOM components from seawater, and this could be significant given the large surface area of these gels. Bacteria can attach to gels76,77 and it is possible that they hydrolyse the nutrients that are present on the gel surface using cell-surface hydrolases78. This, in turn, might alter the local architecture of the gel and the nutrient dynamics at the microscale.
The architecture of gels has been detected using stains that target proteoglycan, protein and DNA
(FIG. 3). These methods might not be sensitive enough
to reveal all of the details of gel architecture, and some, or even most, of the gel might be too diffuse to be detected by imaging technology. Techniques that are useful for macromolecular-level imaging, such as atomic force microscopy79–81, could also be useful for studying marine gels. In addition, the architecture of organic matter is based on information obtained using methods that can only detect structures in two dimen-sions, after sample collection on filters. One important, but challenging, goal is to develop methods that can visualize and biochemically characterize the micros-cale architecture of the organic matter gel matrix in
relation to the distribution of bacterial taxa in various physiological states. Confocal laser microscopy might be useful, as gels can be viewed in three dimensions, and this method might be able to more closely pin-point the physical relationship between bacteria and the organic matter of these gels. Fluorescently labelled lectins, and other probes for biochemical composition, might be useful for determining the composition of gels82,83. Microspatial viscosity is probably variable84 (for example, on the algal surface or near a lysing dino-flagellate) and this might influence bacterial adaptive behaviours85,86, such as motility, microspatial distri-bution and in situ physiological states87. Individual bacteria might experience different microspatial nutrient concentrations at different positions within the organic matter matrix. Also, bacteria could export inhibitory molecules that become bound to the gel matrix, so enabling them to compete with other bac-teria by niche modification. Clearly, much remains to be learnt about the structural and chemical dynamics of the microscale architecture of organic matter and its relationship with bacteria.
Research on gel architecture has altered how micro-bial oceanographers think about the function of bacteria in marine ecosystems. Microspatial architecture provides huge surfaces for bacterial attachment and interactions. Indeed, the enormous genetic diversity of marine bac-teria37–39,88,89 might be explained by the ability of gels to provide niche diversity in seemingly homogeneous ocean waters. For example, many, or all, bacteria that are currently considered to be free-living, for example, SAR11, might in fact be attached to the gel matrix. This would mean that oligotrophs, such as SAR11, do not compete directly for dissolved solutes. Acinas and col-leagues90 suggested that the clusters of microdiversity that they detected in pelagic bacteria might be due to the presence of pelagic microniches. As primary produc-tivity and DOC generally decrease offshore91,92, the hot spots of DOM production and bacterial activity might be less abundant. A bacterium’s-eye view of organic matter offers a more elaborate and dynamic spectrum of choices, rather than the traditional dichotomy that is derived from regarding bacteria as either attached or free living.
In situ growth rate variability. A major physiological variable among oceanic bacterial assemblages is the ability to grow over a broad range of growth rates, from nearly zero to more than one doubling per day12,19,25,93,94. These growth rates — in combination with population size and biomass — are reflected by the variation in BCD and bacteria–phytoplankton coupling. Marine isolates typically grow fast in enriched culture media (although notably SAR11 cannot grow on rich media95). We propose that high growth rates occur periodically in nutrient-rich microzones96–98, for example, near to, or on, nutrient-rich particles99,100, plankton surfaces or in the guts of animals. Thus, the slow average-growth rates that are typically observed for pelagic assemblages do not preclude the possibility that a small fraction of the assemblage might be growing rapidly101–103.
Bacterial growth efficiency (BGE). This is a crucial variable for ecosystem function. The BGE compares the fraction of assimilated carbon that is respired with the fraction that is used to increase bacterial bio-mass. The BGE for natural assemblages is usually 10– 30%, but this can vary widely from 1 to 40%; this means that 60–99% of all assimilated carbon is respired27,104–106. The variation in BGE can affect the role of bacteria in carbon cycling, owing to alterations in the partitioning of carbon between carbon dioxideand biomass, which is accessible to animals. BGE varies between oligotrophic and eutrophic ecosystems107. A mixed population has a range of BGE’s that are dependent on the physiological state of individual bacteria and their spatial and tempo-ral interactions with organic matter107. It is important to place BGE in a microspatial context. A goal should be to determine how the environment affects the respira-tion, growth and phylogenetic identity of individual cells and thereby influences BGE on the microscale.
Bacterial coupling to primary production
Bacterial performance (as measured by growth, respi-ration and other metabolic activities) is constrained by the fact that bacteria are obligate osmotrophs, whereas primary production mainly produces particulate and polymeric organic matter. As a result, the in situ physi-ological attributes and adaptations of bacteria must be responsive to the production of DOM in the microenvi-ronment of the bacterium. Another adaptive challenge for the bacterium is to position itself optimally in rela-tion to DOM producrela-tion. For example, is it adapted to growth in DOM-production hot spots or does it exploit the environmental volume-fraction at the tail-end of DOM gradients? Methods that have been developed to determine the growth rate and phylogenetic identity of individual cells within natural assemblages constitute powerful tools to relate in situ physiology and taxonomy in the microenvironment93,94,108–111.
Bacteria, such as SAR11 (REFS 65,66), members of the
Roseobacter clade, including the alphaproteobacteria Roseobacter-clade-affiliated cluster67,68,112, and members of the Bacteroides clades113,114,might be good models for defining the adaptive strategies that are used by bacteria in relation to DOM-production regimes in microenvi-ronments. Available substrates are maintained at pico-molar to nanopico-molar concentrations in the bulk phase — Hedges and colleagues estimated that “1012 diverse organic molecules...[are]...dissolved in every millilitre of seawater…”, which are “dynamically shaped and buffered by microbiological action” — and, therefore, DOM pro-duction forms DOM gradients against the background of extremely low bulk-phase concentrations.
In proposing how bacteria adapt to use low or high DOM concentrations it is important to recognize that the conversion of POM to DOM involves many different phytoplankton organisms that use varied mechanisms of DOM production. Therefore, numerous adaptive strategies might be used by diverse bacterial taxa.
Phytoplankton are organic matter production loci. The main processes that convert up to half of all the primary production into DOM must occur before substantial
amounts are transferred through multiple trophic lev-els and respired. The most likely mechanisms of DOM formation are direct processes, such as phytoplank-ton exudation and lysis (by virus attack or nutrient stress), bacterial interactions with live phytoplankton (for example, commensalism or predation) and the bacterial enzymatic degradation of recently dead phytoplankton and protists. The aggregation of phy-toplankton and phyphy-toplankton detritus could facilitate bacterial access to DOM. Finally, protists that exhibit boom-and-bust growth cycles could enable bacteria to use protist biomass.
Bacteria–phytoplankton interactions. One way for bacte-ria to metabolically couple to primary production would be to swim up to phytoplankton and use cell-surface hydrolases to kill them115
.
Bacteria have various adapta-tions that allow them to attach to dead phytoplankton and cause hydrolysis116. If bacteria are randomly distributed, each bacterium would be a few hundred micrometres away from the nearest phytoplankton cell9 — a dis-tance that most motile marine bacteria could traverse rather quickly (although many bacteria are non-motile and would not use this strategy). Bacteria can cluster around phytoplankton117 to create considerably higher concentrations than the average for seawater, which is 106 bacteria per ml85. Tight spatial coupling between marine bacteria (Pseudoalteromonas haloplanktis and Shewanella putrefaciens) and a motile alga (Pavlova luth-eri) has been filmed and referred to as the ‘pestering’ of algae, because the bacteria closely tracked motile phyto-plankton52. It seems that some phytoplankton–bacteria associations might even be species specific118–120.Phytoplankton produce surface mucus, polysac-charides and proteoglycans, which might serve as a protection from bacteria; this is analogous to corals, which convert substantial photosynthate into mucus for defence against microbial invasions121. Some phyto-plankton also produce inhibitory compounds122. Mucus creates a region around the phytoplankton cell that is rich in organic matter, and is known as the phycosphere117. The nitrogen- and phosphorus-depleted mucus could adsorb nitrogen- and phosphorous-rich materials, such as polymers and colloids, from seawater, resulting in the development of a rich gel medium. Although much research has been done on bacteria that are attached to phytoplankton it has been technically difficult to study the in situ physiology of bacteria (growth, respiration and antibiotic synthesis) in the phycosphere. It is also likely that the community composition of bacteria in the phycosphere will differ from that in the local ocean envi-ronment. Significant diurnal alterations in the bacteria– phytoplankton relationship are to be expected, but these need to be addressed in a microspatial context. As the phycosphere is organically rich it has been proposed that it could support the proliferation of human patho-gens or other bacteria that are adapted to high-nutrient environments97.
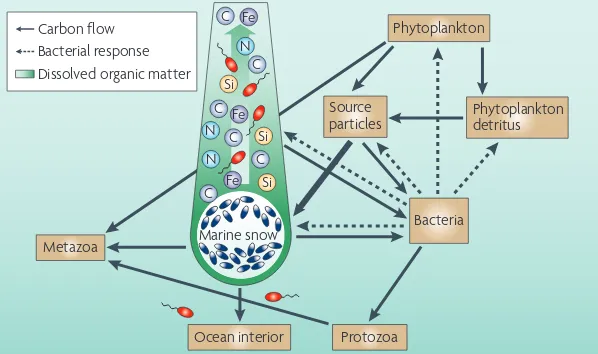
Metazoa
Fe Fe
Fe
C C C C
C C
N N
N
Si Si Si
Marine snow Bacteria Source
particles Phytoplankton detritus Phytoplankton
Protozoa Ocean interior
Bacterial response Carbon flow
Dissolved organic matter
Chemotaxis
The sensing by bacteria of chemical gradients, and movement up or down a gradient towards or away from a chemical source.
Dimethyl sulphide
(DMS). A sulphur-containing organic chemical compound that is a breakdown product of dimethylsulphoniopropionate (DMSP). It is also produced by the metabolism of
methanethiol by marine bacteria that are associated with phytoplankton.
Marine snow
Composed of organic aggregates more than 0.5 mm in diameter. These macroscopic particles are enriched in organic matter and are inhabited by a rich and diverse community of phytoplankton, protozoans and bacteria.
Do bacteria produce either endohydrolases or exohydro-lases, or both, and can bacteria couple hydrolysis and the uptake of DOM in the phycosphere? Endohydrolase action could release polymers into the microenviron-ment that might then be accessible to exohydrolases, which could, in turn, release monomers that bacteria can take up. The released polymers might be detectable as transparent gel particles in seawater124,125.
Microscale processes in the phycosphere. Because the cell-associated phycosphere has fundamental implica-tions for the adaptive biology of bacteria and phyto-plankton, we must address microscale processes within the phycosphere at the mechanistic level. The relation-ship between clustering bacteria and phytoplankton is probably complex and variable over short timeframes. Clustered bacteria could benefit the phytoplankton cell by enhancing nutrient regeneration in the phycosphere at the expense of dissolved organic nitrogen and phos-phorus. The inorganic nutrient hot spot that surrounds the phytoplankton cell could make its microenviron-ment eutrophic in what might otherwise be oligotrophic water according to bulk seawater analysis. This hot spot of nutrients could also receive protist contributions owing to the intense grazing of bacteria by bacterivo-rous protozoa, which would release regenerated nitro-gen, phosphorus and iron into the microenvironment. However, if seawater becomes depleted in dissolved organic nitrogen and dissolved organic phosphorus then the presence of clustering bacteria might also reduce the release of dissolved inorganic nitrogen and dissolved
inorganic phosphorus, thus further limiting primary productivity. Stressed phytoplankton might be unable to defend themselves by the release of mucus or antibiotics122 and clustering or colonizing bacteria could then kill the phytoplankton for use as a growth substrate. The nutrient-hot-spot premise9, if supported, would change our concepts and models of the nutrient regu-lation of primary production. Current models assume there is homogeneity in nutrient distribution. This is another example where further study of a microspatial process promises to yield insights into the regulation of primary production and carbon cycling.
The phycosphere might also be important for the use of dimethylsulphoniopropionate (DMSP) by bacteria. Many marine algae produce this osmolyte and release some of it into the microenvironment. Some marine bacteria, including SAR11 and Roseobacter clades, use DMSP as a source of energy109,126,127. Intracellular pools of DMSP are present in algae at approximately 0.2–0.5 M, although bulk seawater concentrations are approximately 10 nM. DMSP and extracts of Pfiesteria piscicida function as chemoattractants for Silicibacter spp. strainTM1040, an alphaproteobacterium that can grow in close associa-tion with the dinoflagellate P. piscicida. Chemotaxis might enable Silicibacter spp. strainTM1040 to remain close to
P. piscicida128. It would be of interest to know whether community composition and the growth environment in the phycosphere affects the metabolic fate of DMSP129,130. This could affect the rate of bacterial hydrolysis of DMSP to dimethyl sulphide (DMS)131,132. As DMS can affect cli-matic processes, understanding the regulation of the conversion of DMSP to DMS at the microscale could lead to more informed global climate models.
The action of cell-surface-associated bacterial pro-tease and glucosidase on diatom surfaces might reduce diatom ‘stickiness’ and the aggregation potential. This enzymatic ‘pruning’ of the diatom mucus would require microscale interactions between diatoms and bacteria; a high concentration of bacteria in the phycosphere would probably increase the rate of this process133. Diatom aggregation can form large, rapidly sinking aggregates known as marine snow(FIG. 4), which are important for the export of organic matter to the ocean’s depths134 as well as the demise of algal blooms. Understanding the microscale action of bacterial hydrolases on diatom surfaces could help us to predict the timing of bloom termination and the regulation of carbon export in an ecosystem context — this would represent a genome–biogeochemistry connection.
Interaction of bacteria with protists.Protists consume more than half the primary production in many ecosys-tems135. How can we reconcile the large grazing pressure on phytoplankton by protists with an equally high bacte-rial carbon demand? Perhaps when the BCD and protist grazing are both high, a large fraction of the organic mat-ter that is ingested by protists is released through the lysis of these protists by viruses or other factors136. Bacteria can rapidly colonize dead protists and degrade fresh protist detritus. However, it remains to be determined whether heterotrophic bacteria cause significant protist
mortality in an ‘eat them before they eat you’ scenario. In this hypothetical scenario, protists are the proces-sors of primary production, their enzymes convert the POM of prey to DOM and their newly synthesized biomass is consumed by bacteria as predators. Protists are important grazers of small, highly abundant phyto-plankton, such as Synechococcus, Prochlococcus and pico-eukaryotes135. Therefore, protist processing of primary production might be one mechanism that increases the coupling of bacteria to primary production.
Interactions of bacteria with detritus. Dead phytoplank-ton cells are a source of organic matter for bacteria, which readily colonize such material to form a detrito-sphere133,137(FIG. 4). Bacterial colonization and enzymatic hydrolysis could convert detritus to DOM. However, not all bacteria attach to dead phytoplankton. In an experi-mental system, dense clusters of bacteria were recorded swimming around a killed dinoflagellate, and these were modelled to predict bacterial interactions with DOM hot spots51,123,138. Depending on the particle sinking rate, the effects of bacteria on dead phytoplankton cells might result in DOM plumes (as discussed later).
Diatoms attract bacteria, and freshly lysed diatoms comprise organic matter hot spots that support the robust growth of bacteria. Because diatoms require silicon for growth, the mechanisms of silicon recycling are important. Unexpectedly, it was found that bacteria that do not require silicon regulate the regeneration of silicon from dead diatoms116. Although bacteria use their ectohydrolases to solubilize organic matter, the action of these enzymes also removes the protective proteoglycan silaffins fromthe silicon frustules139 and it is the silaffin proteins that normally prevent the dissolution of silicon116. This process requires colonization by bacteria, and those bacteria with high cell-specific protease activity cause more rapid dissolu-tion. In view of the importance of diatom production, and the regeneration of silicon, the regulation of the silicon cycle by bacteria could affect the carbon-export flux. Consequently, an understanding of the biochemi-cal basis of this process should be incorporated into biogeochemical models.
This is an example of a bacteria-mediated process at the microscale that results in the regulation of the silicon cycle and coupled silicon–carbon cycles in the global ocean. Bacterial enzymatic action involves a trivial amount of carbon that is contained in the pro-teoglycan covering and is readily accessible to bacteria. However, the action has a major effect on the biogeo-chemical behaviour of silicon. This might suggest that bacteria have a more general ability that allows them to de-mineralize living or dead silicoflagellates, radiolarians or calcified phytoplankton, most notably Emiliania hux-leyi. However, whether bacteria really do de-mineralize these organisms will depend on the relationship between the mineralized structure and organic matter140.
Marine snow and nutrient hot spots
Aggregation state of the ocean. The aggregation of organic matter is a fundamentally important process in
the functioning of marine ecosystems. Aggregation gener-ally increases the sedimentation rate of organic matter141; the degree of aggregation influences the residence time of component particles in the upper ocean, where they are acted on by bacteria that respire carbon and regenerate nutrients. Sinking aggregates are a dominant conduit for the export flux of organic matter, the variation of which influences carbon storage. The components of the organic matter continuum tend to aggregate to varying degrees forming a dynamic size spectrum. Bacteria can inhibit, as well as enhance, the aggregation state of the system (by reducing the stickiness with hydrolases or increasing it by mucus production)78,133. Aggregates can attract micro-organisms, such as bacteria, protists and viruses, as well as gels, colloids and cell debris. Although most aggregates are microscopic, some are large, such as marine snow, which is visible underwater, and has been extensively studied for its ecosystem significance. Next, we discuss a model that illustrates the significance of microscale activities of bacteria in marine snow, and which has implications for ocean-basin and global-scale processes.
Marine snow. Bacteria colonize marine snow in population densities that reach 108–109 per ml142. The expression of several ectohydrolases, such as protease, lipase, chitinase and phosphatase, is high, but glucosidases are not well expressed. This enzyme complement digests some of the DOM in marine snow78. However, the colonizing bacteria use only a fraction of the hydrolysate (weak coupling)78,143 and, therefore, the sinking marine snow leaves an extended plume of DOM behind them that other bacteria can use. One model predicts that approximately half of all the BCD in the ocean is satisfied by bacterial interaction with these plumes144 (FIG. 4)
.
net efficiency of the biological carbon pump cannot currently be predicted, but we think that better models will be possible through such studies on the biochemical bases of bacteria-organic-matter interactions.
How bacteria influence the aggregation and biogeo-chemical fate of carbon is also relevant to concerns about ocean fertilization and carbon sequestration, in which the goal is to maximize aggregation and export carbon flux. Our discussion in this Review suggests that models that predict the outcome of ocean fertilization should include the roles of bacteria and, specifically, the in situ
expression of selected ectohydrolases148.
Many bacteria are non-motile, notably pelagic Bacteroidetes and SAR11 (REF. 65). Bacteroidetes are specialized for colonizing aggregates such as marine snow113,114,149. How do non-motile bacteria accumulate to form such large populations on marine snow, which has such a short residence time in the upper ocean? Perhaps, non-motile bacteria, being particle special-ists, attach first to the highly abundant small gel par-ticles113,114, which are in the 10-µm length range and are several orders-of-magnitude more abundant than marine snow (typically 1–10 aggregates per litrein the upper ocean)72. Aggregation with larger particles and agglomeration with other materials, such as phyto-plankton or detritus, forms marine snow. Therefore, the source particles150 would contribute diverse bac-teria, including non-motile bacbac-teria, that have been selected for by attaching to the particles that eventually aggregate to form marine snow. Future metagenomic surveys should consider these seascapes of organic mat-ter, which potentially contain an immense diversity of bacteria that are attuned to the nature and dynamics of their miniscule worlds.
Conclusion and future prospects
Microbial oceanography is a field that is caught between scales — microbial processes must be understood at the scale of the individual microorganism, but yet we want to understand the cumulative influence of micro-bial processes on the ocean as a biogeochemical system. We have argued that understanding the biochemical bases of how bacteria interact with the ocean system
at the nanometre (molecular) to millimetre scale can provide insights into globally significant biogeochemi-cal processes. Therefore, an understanding of nanosbiogeochemi-cale biochemistry can be extended hierarchically151 to the global ocean and the Earth’s biogeochemistry. Indeed, some insights are not accessible by large-scale studies alone. We need a robust understanding of microscale biogeochemistry and how it fits with ocean and global biogeochemical studies of all scales. This should result in models of the biochemical bases for the interactions among organisms and the environment.
We need additional methods and instruments that can measure individual bacteria-cell in situ growth and respiration rates in natural seawater in three dimen-sions, without perturbing these assemblages by using filters. It might also be possible to study other basic ecological interactions, such as grazing and phage lysis. Methods and instruments that can be used to study the activities of microorganisms in the context of their ecosystem are on the horizon. They should enable us, for example, to interrogate the chemical and physical characteristics of the environmental architecture in relation to bacterial diversity, distribution and activ-ity. With the current momentum in nanotechnology, nanobiology and advanced imaging we see no reason why microbial ecologists cannot explore the oceans at the nanometre–millimetre scale. Eventually, one goal of microbial oceanographers should be to understand carbon cycling and to visualize microbial interactions that affect the biogeochemical state of the ocean.
Perhaps it is stating the obvious, but we would prob-ably not be concerned for the health of corals or tropical forests if they were invisible. The microscale architecture of the ocean and its relationship with much of the diver-sity in the sea may well be delicate, and could be sensitive to new patterns of enzyme expression or activity that might arise owing to warming and acidification. It is con-ceivable that exploration of the ocean at the microscale will yield novel measures of the ocean’s biogeochemical state, or ocean health. We stress the need for a concerted research effort in microscale biogeochemistry as a disci-pline that is integrated with environmental genomic and ecosystem research and climate science.
1. Field, C. B., Behrenfeld, M. J., Randerson, J. T. & Falkowski, P. Primary production of the biosphere: integrating terrestrial and oceanic components.
Science281, 237–240 (1998).
2. Falkowski, P. G., Barber, R. T. & Smetacek, V. Biogeochemical controls and feedbacks on ocean primary production. Science281, 200–206 (1998).
3. Pauly, D. & Christensen, V. Primary production required to sustain global fisheries. Nature374, 255–257 (1995).
4. Ocean Biogeochemistry: a Synthesis of the Joint Global Ocean Flux Study (JGOFS) (ed. Fashsam, M. J. R.) (Springer, New York, 2003).
This book describes results from a long-term research program on the role of the ocean carbon cycle in global change.
5. Jackson, J. B. C. et al. Historical overfishing and the recent collapse of coastal ecosystems. Science293, 629–637 (2001).
6. Pauly, D. et al. The future for fisheries. Science302, 1359–1361 (2003).
7. Pomeroy, L. R. Oceans food web, a changing paradigm. Bioscience24, 499–504 (1974).
An influential paper that proposed that a major fraction of primary production is used by bacteria and other microorganisms.
8. Williams, P. J. L. Microbial contribution to overall marine plankton metabolism: direct measurements of respiration. Oceanol. Acta4, 359–364 (1981).
9. Azam, F. & Ammerman, J. W. in Flows of Energy and Materials in Marine Ecosystem (ed. Fasham, M. J. R.) 345–360 (1984).
10. Steele, J. The Structure of Marine Ecosystems. (Harvard Univ. Press, Massachusetts, 1974). 11. Hobbie, J. E., Daley, R. J. & Jasper, S. Use of
nucleopore filters for counting bacteria by fluorescence microscopy. Appl. Environ. Microbiol.33, 1225–1228 (1977).
12. Fuhrman, J. A. & Azam, F. Bacterioplankton secondary production estimates for coastal waters of British Columbia, Canada, Antarctica, and California, USA. Appl. Environ. Microbiol.39, 1085–1095 (1980).
13. Hagström, Å., Larsson, U., Horstedt, P. & Normark, S. Frequency of dividing cells: a new approach to the determination of bacterial growth rates in aquatic environments. Appl. Environ. Microbiol.37, 805–812 (1979).
14. Giovannoni, S. J. & Stingl, U. Molecular diversity and ecology of microbial plankton. Nature437, 343–348
(2005).
15. DeLong, E. F. & Karl, D. M. Genomic perspectives in microbial oceanography. Nature437, 336–342 (2005).
An excellent review on the role of microorganisms in marine ecosystems that combined molecular and ecological perspectives.
16. Pomeroy, L. R., Williams, P. J., Azam, F. & Hobbie, E. A. The microbial loop. Oceanography20, 28–33 (2007).
A concise account of the functioning of the microbial loop in the marine ecosystem. 17. Williams, P. J. l. Incorporation of microheterotrophic
processes into the classical paradigm of the planktonic food web. Kiel. Meeresforsch5, 1–28
18. Azam, F. et al. The ecological role of water-column
microbes in the sea. Mar. Ecol. Prog. Ser.10, 257–263 (1983).
19. Ducklow, H. W. & Carlson, C. A. Oceanic bacterial production. Adv. Microb. Ecol.12, 113–181 (1992). 20. Williams, P. J. l. B. The balance of plankton respiration
and photosynthesis in the open oceans. Nature394, 55–57 (1998).
21. Azam, F. Microbial control of oceanic carbon flux: the plot thickens. Science280, 694–696 (1998). 22. Hollibaugh, J. T. & Azam, F. Microbial-degradation of
dissolved proteins in seawater. Limnol. Oceanogr.28, 1104–1116 (1983).
23. Ducklow, H. W. The bacterial component of the oceanic euphotic zone. FEMS Microbiol. Ecol.30, 1–10 (1999). 24. Karl, D. M. Nutrient dynamics in the deep blue sea.
Trends Microbiol.10, 410–418 (2002).
25. Ducklow, H. W. Production and fate of bacteria in the oceans. Bioscience33, 494–501 (1983). 26. Ducklow, H. W. Modeling the microbial food-web.
Microb. Ecol.28, 303–319 (1994). 27. Cole, J. J., Findlay, S. & Pace, M. L. Bacterial
production in fresh and saltwater ecosystems — a cross-system overview. Mar. Ecol. Prog. Ser.43, 1–10
(1988).
28. Turley, C. M. et al. Relationship between primary
producers and bacteria in an oligotrophic sea — the Mediterranean and biogeochemical implications. Mar. Ecol. Prog. Ser.193, 11–18 (2000).
29. Hoppe, H. G., Gocke, K., Koppe, R. & Begler, C. Bacterial growth and primary production along a north–south transect in the Atlantic Ocean. Nature 416, 168–171 (2002).
30. Williams, P. J. l. B. & Bower, D. G. Regional carbon imbalances in the oceans. Science284, 1735 (1999). 31. Karl, D. M., Laws, E. A., Morris, P., Williams, P. J. l. &
Emerson, S. Global carbon cycle (communication arising): metabolic balance of the open sea. Nature 426, 32 (2003).
32. Azam, F., Smith, D. C. & Hollibaugh, J. T. The role of the microbial loop in Antarctic pelagic ecosystems.
Polar Res.10, 239–243 (1991).
33. Pomeroy, L. R., Wiebe, W. J., Deibel, D., Thompson, R. J. & Rowe, G. T. Bacterial responses to temperature and substrate concentration during the Newfoundland spring bloom. Mar. Ecol. Prog. Ser.75, 143–159 (1991).
34. Pomeroy, L. R. & Wiebe, W. J. Temperature and substrates as interactive limiting factors for marine heterotrophic bacteria. Aquat. Microb. Ecol.23, 187–204 (2001).
35. Kline, D., Kuntz, N., Brietbart, M., Knowlton, N. & Rohwer, F. The unexpected and critical role of elevated organic carbon in coral mortality. Mar. Ecol. Prog. Ser. 314, 119–125 (2006).
36. Smith, J. E. et al. Effects of algae on coral:
algal-mediated, microbe-induced coral mortality. Ecol. Lett. 9, 835–845 (2006).
37. Venter, J. C. Environmental genome shotgun sequencing of the Sargasso Sea. Science304, 66–74
(2004).
38. DeLong, E. F. et al. Community genomics among stratified microbial assemblages in the ocean’s interior. Science311, 496–503 (2006). 39. Yooseph, S. et al. The Sorcerer II Global Ocean
Sampling Expedition: expanding the universe of protein families. PLoS Biol.5, e16 (2007). 40. Rusch, D. B. et al. The Sorcerer II Global Ocean
Sampling Expedition: northwest Atlantic through eastern Tropical Pacific. PLoS Biol.5, e77 (2007). 41. Béjà, O. et al. Bacterial rhodopsin: evidence for a new
type of phototrophy in the sea. Science289, 1902–1906 (2000).
42. de la Torre, J. R. et al. Proteorhodopsin genes are distributed among divergent marine bacterial taxa.
Proc. Natl Acad. Sci. USA100, 12830–12835 (2003).
43. Azam, F. & Worden, A. Z. Microbes, molecules, and marine ecosystems. Science303, 1622–1624 (2004).
44. Gray, J. S. et al. in Flows of Energy and Materials in Marine Ecosystems (ed. Fasham, M. R. J.) 706–723 (Plenum, New York,1984).
45. Riemann, L. & Middelboe, M. Viral lysis of marine bacterioplankton: implications for organic matter cycling and bacterial clonal composition. Ophelia56,
57–68 (2002).
46. Barbeau, K., Moffett, J. W., Caron, D. A., Croot, P. L. & Erdner, D. L. Role of protozoan grazing in relieving iron limitation of phytoplankton. Nature380, 61–64
(1996).
47. Barbeau, K., Kujawinski, E. B. & Moffett, J. W. Remineralization and recycling of iron, thorium and organic carbon by heterotrophic marine protists in culture. Aquat. Microb. Ecol.24, 69–81 (2001). 48. Grossart, H. P., Riemann, L. & Azam, F. Bacterial
motility in the sea and its ecological implications.
Aquat. Microb. Ecol.25, 247–258 (2001). 49. Mitchell, J. G., Pearson, L., Dillon, S. & Kantalis, K.
Natural assemblages of marine-bacteria exhibiting high-speed motility and large accelerations. Appl. Environ. Microbiol.61, 4436–4440 (1995). 50. Mitchell, J. G. et al. Long lag times and high velocities in
the motility of natural assemblages of marine-bacteria.
Appl. Environ. Microbiol.61, 877–882 (1995). 51. Blackburn, N., Fenchel, T. & Mitchell, J. Microscale
nutrient patches in planktonic habitats shown by chemotactic bacteria. Science282, 2254–2256 (1998).
An experimental demonstration of the response of marine bacteria to organic matter hot spots and a simulation by numerical modelling.
52. Barbara, G. M. & Mitchell, J. G. Bacterial tracking of motile algae. FEMS Microbiol. Ecol.44, 79–87 (2003). 53. Martinez, J., Smith, D. C., Steward, G. F. & Azam, F.
Variability in ectohydrolytic enzyme activities of pelagic marine bacteria and its significance for substrate processing in the sea. Aquat. Microb. Ecol. 10, 223–230 (1996).
54. Arrieta, J. M. & Herndl, G. J. Assessing the diversity of marine bacterial β-glucosidases by capillary electrophoresis zymography. Appl. Environ. Microbiol. 67, 4896–4900 (2001).
55. Kirchman, D. L. & White, J. Hydrolysis and mineralization of chitin in the Delaware Estuary.
Aquat. Microb. Ecol.18, 187–196 (1999). 56. Nagata, T., Meon, B. & Kirchman, D. L. Microbial
degradation of peptidoglycan in seawater. Limnol. Oceanogr.48, 745–754 (2003).
57. Arnosti, C., Durkin, A. S. & Jeffrey, W. H. Patterns of extracellular enzyme activities among pelagic marine microbial communities: implication for cycling of dissolved organic carbon. Aquat. Microb. Ecol.38, 135–145 (2005).
58. Obayashi, Y. & Suzuki, S. Proteolytic enzymes in coastal surface seawater: significant activity of endopeptidases and exopeptidases. Limnol. Oceangr. 50, 722–726 (2005).
59. Cottrell, M. T., Yu, L. Y. & Kirchman, D. L. Sequence and expression analyses of Cytophaga-like hydrolases in a western Arctic metagenomic library and the Sargasso Sea. Appl. Environ. Microbiol.71, 8506–8513 (2005).
60. Williams, P. J. in Microbial Production and the Decomposition of Organic Material Ch. 3 (eds.Kaiser, M., Attrill, M., Jennings, S., Thomas, D. N. & Williams, P. J. le B) (Oxford Univ. Press, 2005). 61. Azam, F. & Hodson, R. E. Multiphasic kinetics for
D-glucose uptake by assemblages of natural marine-bacteria. Mar. Ecol. Prog. Ser.6, 213–222 (1981). 62. Nissen, H., Nissen, P. & Azam, F. Multiphasic uptake
of D-glucose by an oligotrophic marine bacterium.
Mar. Ecol. Prog. Ser.16, 155–160 (1984). 63. Riemann, L. & Azam, F. Widespread N
-acetyl-D-glucosamine uptake among pelagic marine bacteria and its ecological implications. Appl. Environ. Microbiol.68, 5554–5562 (2002).
64. Alonso, C. & Pernthaler, J. Concentration-dependent patterns of leucine incorporation by coastal picoplankton. Appl. Environ. Microbiol.72,
2141–2147 (2006).
65. Giovannoni, S. J. et al. Genome streamlining in a cosmopolitan oceanic bacterium. Science309, 1242–1245 (2005).
66. Morris, R. M. et al. SAR11 clade dominates ocean surface bacterioplankton communities. Nature420, 806–810 (2002).
67. Moran, M. A. Genome sequence of Silicibacter pomeroyi reveals adaptations to the marine environment. Nature432, 910–913 (2004). 68. Moran, M. A. et al. Ecological genomics of marine
Roseobacters. Appl. Environ. Microbiol.73, 4559–4569 (2007).
69. Blackburn, N., Azam, F. & Hagstrom, A. Spatially explicit simulations of a microbial food web. Limnol. Oceanogr.42, 613–622 (1997).
70. Koike, I., Hara, S. I., Terauchi, K. & Kogure, K. Role of sub-micrometre particles in the ocean. Nature345, 242–244 (1990).
A fundamental discovery that showed the existence of highly abundant sub-micrometre organic particles in the ocean.
71. Wells, M. L. & Goldberg, E. Occurrence of small colloids in seawater. Nature353, 342–344 (1992). 72. Verdugo, P. et al. The oceanic gel phase: a bridge in the DOM–POM continuum. Mar. Chem.92, 67–85 (2004).
An excellent synthesis that showed that organic matter in the sea consists of a gel phase that forms a size continuum. This framework is crucial for understanding the ecology of bacteria and their biogeochemical activities.
73. Chin, W. C., Orellana, M. V. & Verdugo, P. Spontaneous assembly of marine dissolved organic matter into polymer gels. Nature391, 568–572 (1988). 74. Chin, W. C., Orellana, M. V., Quesada, I. & Verdugo, P.
Secretion in unicellular marine phytoplankton: demonstration of regulated exocytosis in Phaeocystis globosa. Plant Cell Physiol.45, 535–542 (2004).
This paper describes how phytoplankton might
contribute to the gel phase of seawaterby
exocytosis.
75. Ogawa, H., Amagai, Y., Koike, I., Kaiser, K. & Benner, R. Production of refractory dissolved organic matter by bacteria. Science292, 917–920 (2001). 76. Long, R. A. & Azam, F. Abundant protein-containing
particles in the sea. Aquat. Microb. Ecol.10, 213–221 (1996).
77. Alldredge, A. L., Passow, U. & Haddock, S. H. D. The characteristics and transparent exopolymer particle (TEP) content of marine snow formed from thecate dinoflagellates. J. Plankton Res.20, 393–406 (1998). 78. Smith, D. C., Simon, M., Alldredge, A. L. & Azam, F.
Intense hydrolytic enzyme activity on marine aggregates and implications for rapid particle dissolution. Nature359, 139–142 (1992). 79. Santschi, P. H. et al. Fibrillar polysaccharides in
marine macromolecular organic matter, as imaged by atomic force microscopy and transmission electron microscopy. Limnol. Oceanogr.43, 896–908 (1998). 80. Dupres, V. et al. Nanoscale mapping and functional
analysis of individual adhesins on living bacteria.
Nature Methods2, 515–520 (2005). 81. Dufrêne, Y. F. Nanoscale exploration of microbial
surfaces using the atomic force microscope. Future Microbiol.1, 387–396 (2006).
82. Neu, T. R., Walczysko, P. & Lawrence, J. R. Two-photon imaging for studying the microbial ecology of biofilm systems. Microb. Environ.19, 1–6 (2004). 83. Decho, A. W. & Kawaguchi, T. Confocal imaging of
in situ natural microbial communities and their extracellular polymeric secretions (EPS) using nanoplast resin. BioTechniques27, 1246–1251 (1999).
84. Belas, R., Simon, M. & Silverman, M. Regulation of lateral flagella gene transcription in Vibrio parahaemolyticus. J. Bacteriol.167, 210–218 (1986).
85. Bowen, J. D., Stolzenbach, K. D. & Chisholm, S. W. Simulating bacterial clustering around phytoplankton cells in a turbulent ocean. Limnol. Oceangr.38, 36–51 (1993).
86. Fenchel, T. & Blackburn, N. Motile chemosensory behaviour of phagotrophic protists: mechanisms for and efficiency in congregating at food patches. Protist 150, 325–336 (1999).
87. Seymour, J. R., Mitchell, J. G. & Seuront, L. Microscale heterogeneity in the activity of coastal
bacterioplankton communities. Aquat. Microb. Ecol. 35, 1–16 (2004).
88. Rocap, G., Distel, D. L., Waterbury, J. B. & Chisholm, S. W. Resolution of Prochlorococcus and
Synechococcus ecotypes by using 16s–23s ribosomal DNA internal transcribed spacer sequences. Appl. Environ. Microbiol.68, 1180–1191 (2002). 89. Johnson, Z. I. et al. Niche partitioning among
Prochlorococcus ecotypes along ocean-scale environmental gradients. Science311, 1737–1740 (2006).
90. Acinas, S. G. Fine-scale phylogenetic architecture of a complex bacterial community. Nature430, 551–554 (2004).
This study shows the existence of microscale phylogenetic clusters among marine bacteria assemblages, which has significance for gene diversity and the interaction with ocean systems. 91. Kolber, Z. S., Van Dover, C. L., Niederman, R. A. &
Falkowski, P. G. Bacterial photosynthesis in surface waters of the open ocean. Nature407, 177–179 (2000).
93. Teira, E., Reinthaler, T., Pernthaler, A., Pernthaler, J. & Herndl, G. J. Combining catalyzed reporter deposition-fluorescence in situ hybridization and
microautoradiography to detect substrate utilization by bacteria and archaea in the deep ocean. Appl. Environ. Microbiol.70, 4411–4414 (2004). 94. Cottrell, M. T. & Kirchman, D. L. Single-cell analysis
of bacterial growth, cell size, and community structure in the Delaware estuary. Aquat. Microb. Ecol.34, 139–149 (2004).
Presents a method for the simultaneous phylogenetical and physiological interrogation of individual cells in natural marine assemblages. 95. Rappe, M. S., Connon, S. A., Vergin, K. L. &
Giovannoni, S. J. Cultivation of the ubiquitous SAR11 marine bacterioplankton clade. Nature418, 630–633 (2002).
96. Mourino-Perez, R. R., Worden, A. Z. & Azam, F. Growth of Vibrio cholerae O1 in red tide waters off California. Appl. Environ. Microbiol.69, 6923–6931 (2003).
97. Worden, A. Z. et al. Trophic regulation of Vibrio cholerae in coastal marine waters. Environ. Microbiol. 8, 21–29 (2006).
98. Hamasaki, K., Long, R. A. & Azam, F. Individual cell growth rates of marine bacteria, measured by bromodeoxyuridine incorporation. Aquat. Microb. Ecol.35, 217–227 (2004).
99. Fandino, L. B., Riemann, L., Steward, G. F., Long, R. A. & Azam, F. Variations in bacterial community structure during a dinoflagellate bloom analyzed by DGGE and 16s rDNA sequencing. Aquat. Microb. Ecol.23,
119–130 (2001).
100. Riemann, L., Steward, G. F. & Azam, F. Dynamics of bacterial community composition and activity during a mesocosm diatom bloom. Appl. Environ. Microbiol. 66, 578–587 (2000).
101. Rodriguez, G. G., Phipps, D., Ishiguro, K. & Ridgway, H. F. Use of a fluorescent redox probe for direct visualization of actively respiring bacteria.
Appl. Environ. Microbiol.58, 1801–1808 (1992). 102. Lebaron, P., Servais, P., Agogue, H., Courties, C. &
Joux, F. Does the high nucleic acid content of individual bacterial cells allow us to discriminate between active cells and inactive cells in aquatic systems? Appl. Environ. Microbiol.67, 1775–1782 (2001).
103. Gasol, J. M., Zweifel, U. L., Peters, F., Fuhrman, J. A. & Hagstrom, A. Significance of size and nucleic acid content heterogeneity as measured by flow cytometry in natural planktonic bacteria. Appl. Environ. Microbiol.65, 4475–4483 (1999).
104. Reinthaler, T. & Herndl, G. J. Seasonal dynamics of bacterial growth efficiencies in relation to phytoplankton in the southern North Sea. Aquat. Microb. Ecol.39, 7–16 (2005).
105. Reinthaler, T., Winter, C. & Herndl, G. J. Relationship between bacterioplankton richness, respiration, and production in the southern North Sea. Appl. Environ. Microbiol.71, 2260–2266
(2005).
106. Alonso-Saez, L. et al. Large-scale variability in
surface bacterial carbon demand and growth efficiency in the subtropical northeast Atlantic Ocean. Limnol. Oceanog.52, 533–546 (2007). 107. Del Giorgio, P. A. & Cole, J. J. Bacterial growth
efficiency in natural aquatic systems. Ann. Rev. Ecol. Syst. 29, 503–541 (1998).
108. Ouverney, C. C. & Fuhrman, J. A. Combined microautoradiography —16s rRNA probe technique for determination of radioisotope uptake by specific microbial cell types in situ.Appl. Environ. Microbiol. 65, 1746–1752 (1999).
109. Malmstrom, R. R., Cottrell, M. T., Elifantz, H. & Kirchman, D. L. Biomass production and assimilation of dissolved organic matter by SAR11 bacteria in the Northwest Atlantic Ocean. Appl. Environ. Microbiol. 71, 2979–2986 (2005).
110. Cottrell, M. T. & Kirchman, D. L. Natural
assemblages of marine proteobacteria and members of the Cytophaga–Flavobacter cluster consuming low- and high-molecular-weight dissolved organic matter. Appl. Environ. Microbiol.66, 1692–1697 (2000).
111. Alonso, C. & Pernthaler, J. Incorporation of glucose under anoxic conditions by Bacterioplankton from coastal North Sea surface waters. Appl. Environ. Microbiol.71, 1709–1716 (2005). 112. Selje, N., Simon, M. & Brinkhoff, T. A newly discovered
Roseobacter cluster in temperate and polar oceans.
Nature427, 445–448 (2004).
113. Kirchman, D. L. The ecology of Cytophaga– Flavobacteria in aquatic environments. FEMS Microbiol. Ecol.39, 91–100 (2002).
114. Alonso, C., Warnecke, F., Amann, R. & Pernthaler, J. High local and global diversity of Flavobacteria in marine plankton. Environ. Microbiol.9, 1253–1266 (2007).
115. Mayali, X. & Azam, F. Algicidal bacteria in the sea and their impact on algal blooms. J. Eukaryot. Microbiol. 51, 139–144 (2004).
116. Bidle, K. D. & Azam, F. Accelerated dissolution of diatom silica by marine bacterial assemblages. Nature 397, 508–512 (1999).
Reports the surprising finding that marine assemblages that do not require silicon mediate and regulate the dissolution of diatom frustules, by proteolytically removing the proteoglycan that can protect the frustule from dissolving. This has implications for carbon and silicon cycles in the ocean. 117. Bell, W. H., Lang, J. M. & Mitchell, R. Selective
stimulation of marine bacteria by algal extracellular products. Limnol. Oceanogr.19, 833–839 (1974). 118. Rooney-Varga, J. N. et al. Links between
phytoplankton and bacterial community dynamics in a coastal marine environment. Microb. Ecol.49,
163–175 (2005).
119. Grossart, H. P., Levold, F., Allgaier, M., Simon, M. & Brinkhoff, T. Marine diatom species harbour distinct bacterial communities. Environ. Microbiol. 7, 860–873 (2005).
120. Sapp, M. et al. Species-specific bacterial communities in the phycosphere of microalgae? Microb. Ecol.53, 683–699 (2007).
121. Wild, C. et al. Coral mucus functions as an energy carrier and particle trap in the reef ecosystem. Nature 428, 66–70 (2004).
122. Trick, C. G., Harrison, P. & Anderson, R. J. Extracellular secondary metabolite production by the marine dinoflagellate Prorocentrum minimum in culture. Can. J. Fish. Aquat. Sci. 38, 864–867 (1981). 123. Mitchell, J. G., Pearson, L. & Dillon, S. Clustering of
marine bacteria in seawater enrichments. Appl. Environ. Microbiol.62, 3716–3721 (1996). 124. Smith, D. C., Steward, G. F., Long, R. A. & Azam, F.
Bacterial mediation of carbon fluxes during a diatom bloom in a mesocosm. Deep-Sea Res. II 42, 75–97
(1995).
125. Alldredge, A. L., Passow, U. & Logan, B. E. The abundance and significance of a class of large, transparent organic particles in the Ocean. Deep-Sea Res. I40, 1131–1140 (1993).
Reports the finding of abundant
mucopolysaccharide particles from 2 to 200 µm in length. These particles provide large surface areas for bacterial interactions and activities.
126. Malmstrom, R. R., Kiene, R. P. & Kirchman, D. L. Identification and enumeration of bacteria assimilating dimethylsulfoniopropionate (DMSP) in the North Atlantic and Gulf of Mexico. Limnol. Oceanogr.49, 597–606 (2004).
127. Gonzalez, J. M. et al.Silicibacter pomeroyi sp nov and
Roseovarius nubinhibens sp nov.,
dimethylsulfoniopropionate-demethylating bacteria from marine environments. Int. J. Syst. Evol. Microbiol.53, 1261–1269 (2003).
128. Miller, T. R., Hnilicka, K., Dziedzic, A., Desplats, P. & Belas, R. Chemotaxis of Silicibacter sp. strain TM1040 toward dinoflagellate products. Appl. Environ. Microbiol.70, 4692–4701 (2004). 129. Yoch, D. C., Ansede, J. H. & Rabinowitz, K. S. Evidence
for intracellular and extracellular
dimethylsulfoniopropionate (DMSP) lyase and DMSP uptake sites in two species of marine bacteria. Appl. Environ. Microbiol.63, 3182–3188 (1997). 130. Howard, E. C. et al. Bacterial taxa that limit sulfur flux
from the ocean. Science 314, 649–652 (2006). 131. Moran, M. A., González, J. M. & Kiene, R. P. Linking a
bacterial taxon to organic sulfur cycling in the sea: studies of the marine Roseobacter group.
Geomicrobiol. J.20, 375–388 (2003). 132. Lovelock, J. E., Maggs, R. J. & Rasmussen, R. A.
Atmospheric dimethyl sulphide and the natural sulphur cycle. Nature237, 452–453 (1972). 133. Azam, F. & Smith, D. C. in Particle Analysis in
Oceanography (ed. Demers, S.) 213–235
(Springer-Verlag, Berlin, 1991).
134. Richardson, T. L. & Jackson, G. A. Small phytoplankton and carbon export from the surface ocean. Science315, 838–840 (2007).
135. Landry, M. R. & Calbet, A. Microzooplankton production in the oceans. ICES J. Mar. Sci.61, 501–507 (2004).
136. Hagstrom, A., Azam, F., Andersson, A., Wikner, J. & Rassoulzadegan, F. Microbial loop in an oligotrophic pelagic marine ecosystem — possible roles of cyanobacteria and nanoflagellates in the organic fluxes. Mar. Ecol. Prog. Ser.49, 171–178 (1988). 137. Biddanda, B. A. & Pomeroy, L. R. Microbial
aggregation and degradation of phytoplankton-derived detritus in seawater.1. Microbial succession.
Mar. Ecol. Prog. Ser. 42, 79–88 (1988). 138. Mueller, R. S. et al.Vibrio cholerae strains
possess multiple strategies for abiotic and biotic surface colonization. J. Bacteriol.189, 5348–5360 (2007).
139. Kroger, N., Lorenz, S., Brunner, E. & Sumper, M. Self-assembly of highly phosphorylated silaffins and their function in biosilica morphogenesis. Science 298, 584–586 (2002).
140. Hedges, J. I. et al. Evidence for non-selective preservation of organic matter in sinking marine particles. Nature409, 801–804 (2001). 141. Turley, C. M. & Stutt, E. D. Depth-related cell-specific
bacterial leucine incorporation rates on particles and its biogeochemical significance in the Northwest Mediterranean. Limnol. Oceanogr.45, 419–425 (2000).
142. Alldredge, A. L., Cole, J. J. & Caron, D. A. Production of heterotrophic bacteria inhabiting macroscopic organic aggregates (marine snow) from surface waters.
Limnol. Oceanogr.31, 68–78 (1986). 143. Cho, B. C. & Azam, F. Major role of bacteria in
biogeochemical fluxes in the ocean’s interior. Nature 332, 441–443 (1988).
144. Kiorboe, T. & Jackson, G. A. Marine snow, organic solute plumes, and optimal chemosensory behavior of bacteria. Limnol. Oceanogr.46, 1309–1318 (2001).
This paper presents a model of marine snow colonized by bacteria that solubilize organic matter, and shows that an extended plume of DOM persists behind the sinking marine snow, which attracts bacteria from surrounding seawater. It predicts that half of the organic matter that is used in the sea by bacteria is from these microenvironments.
145. Azam, F. & Long, R. A. Oceanography — sea snow microcosms. Nature414, 495–498 (2001). 146. Helmstetter, C. E. & Cummings, D. J. An improved
method for the selection of bacterial cells at division. Biochim. Biophys. Acta82, 608–610
(1964).
147. Lochte, K. & Turley, C. Bacteria and cyanobacteria associated with phytodetritus in the deep-sea. Nature 333, 67–69 (1988).
148. Oliver, J. L., Barber, R. T., Smith, W. O. & Ducklow, H. W. The heterotrophic bacterial response during the Southern Ocean iron experiment (SOFeX). Limnol. Oceanogr.49, 2129–2140 (2004).
149. Bauer, M. et al. Whole genome analysis of the marine
Bacteroidetes ‘Gramellaforseti’ reveals adaptations to degradation of polymeric organic matter. Environ. Microbiol.8, 2201–2213 (2006).
This paper presents whole-genome analyses
of a marine Bacteroidetes spp. and makes a
prediction about its adaptive biology that is important for the solubilization and degradation of particulate organic matter in the ocean. 150. Azam, F., Smith, D. C., Steward, G. F. & Hagström, Å.
Bacteria-organic matter coupling and its significance for oceanic carbon cycling. Microb. Ecol. 28, 167–179 (1994).
151. Allen, T. Scale in microscopic algal ecology: a neglected dimension. Phycologia16, 253–257 (1977).
Acknowledgements
We thank T. Hollibaugh, G. Steward, D. Smith and J. Fuhrman for insightful comments on the manuscript. This work was supported by the Gordon and Betty Moore Foundation and grants to F.A. from the National Science Foundation.
Competing interests statement
The authors declare no competing financial interests.
DATABASES
Entrez Genome Project:http://www.ncbi.nlm.nih.gov/ entrez/query.fcgi?db=genomeprj
Candidatus Pelagibacter ubique|Emiliania huxleyi|Pavlova lutheri|Pseudoalteromonas haloplanktis | Shewanella putrefaciens | TM1040