i
METODE NESTED- POLYMERASE CHAIN REACTION
Skripsi
disusun sebagai salah satu syarat untuk memperoleh gelar Sarjana Sain Biologi
Oleh Ngaliyatun 4450408025
JURUSAN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM UNIVERSITAS NEGERI SEMARANG
ii
Daya Infektivitas Plasmodium berghei Iradiasi Pada Hati dan Limpa Mencit Menggunakan Metode Nested-Polymerase Chain Reaction” disusun berdasarkan hasil penelitian saya dengan arahan dosen pembimbing. Sumber informasi atau kutipan yang berasal atau dikutip dari karya yang diterbitkan telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini. Skripsi ini belum pernah diajukan untuk memperoleh gelar dalam program sejenis di perguruan tinggi manapun.
Semarang, 19 September 2013
Ngaliyatun 4450408025
iii Skripsi yang berjudul
Uji daya infektivitas Plasmodium berghei Iradiasi Pada Hati dan Limpa Mencit (Mus musculus) Menggunakan Metode Nested-Polymerase Chain Reaction Disusun oleh
Nama : Ngaliyatun NIM : 4450408025
telah dipertahankan dihadapan sidang panitia ujian skripsi FMIPA UNNES pada Tanggal 05 September 2013.
Panitia Sekretaris
Prof Dr. Wiyanto, M.Si. Andin Irsadi, S.Pd, M.Si. NIP. 196310121988031001 NIP.197403102000031001
Ketua Penguji
Dr. Drh. R. Susanti, MP. NIP. 197111071998022001
Anggota penguji/ Anggota penguji/ Pembimbing Utama Pembimbing Pendamping
Ir. Tuti Widianti, M.Biomed Dr. Mukh Syaifudin NIP. 195102071979032001 NIP.196506011989011001
iv
dan Limpa Mencit Menggunakan Metode Nested-Polymerase Chain Reaction. Skripsi, Jurusan Biologi FMIPA Universitas Negeri Semarang. Ir.Tuti Widianti, M. Biomed dan Dr. Mukh Syaifudin
Plasmodium berghei adalah parasit jenis protozoa penyebab malaria pada rodensia yang ditularkan melalui gigitan nyamuk Anopheles betina yang terinfeksi parasit tersebut. Iradiasi dapat menyebabkan perubahan struktur protein, degradasi protein maupun perubahan konformasi DNA. Dosis iradiasi 150-175 Gy dapat menurunkan daya infeksi P.berghei pada mencit dengan ditunjukkan oleh periode prepaten yang panjang serta jumlah kematian mencit yang rendah. Tujuan dari penelitian ini adalah untuk mendeteksi keberadaaan Plasmodium iradiasi pada hati dan limpa mencit (Mus musculus) menggunakan metode Nested Polymerase Chain Reaction (PCR).
Penelitian ini merupakan penelitian eksperimental yang dilakukan di Laboratorium Biologi Molekuler Bidang Biomedika PTKMR BATAN. Populasi penelitian adalah mencit strain Swiss Webster jantan. Sampel yang digunakan yaitu adalah mencit strain Swiss Webster jantan berumur ±2 bulan dengan berat badan sekitar 35 gram, diperoleh dari Pusat Penyakit Tropis, Badan Litbang Kesehatan, Kementerian Kesehatan Jakarta. Penelitian ini menggunakan P. berghei yang diiradiasi dosis 175 Gy tanpa booster, 175 Gy dengan booster, dan 0 Gy dan diinfeksikan ke dalam tubuh mencit. Setelah 2 bulan hati dan limpa mencit diambil dan dilakukan ekstraksi DNA. Hasil ekstraksi diamplifikasi menggunakan nested-PCR dengan dua pasang primer yaitu rPLU1 dan rPLU5 untuk amplifikasi pertama dan rPLU3 dan rPLU4 untuk amplifikasi kedua. Elektroforesis menggunakan gel agaros dilakukan untuk melihat ada tidaknya pita DNA spesifik yang berukuran sesuai dengan DNA target.
Hasil pemeriksaan DNA genom dari sampel hati dan limpa mencit menunjukkan kualitas cukup baik. Proses amplifikasi nested-1 tidak menunjukkan adanya pita berukuran 1640 bp baik pada sampel maupun pada kontrol positif. Amplifikasi nested-2 tidak menunjukkan adanya pita berukuran 240 bp pada sampel tetapi pada kontrol positif menunjukkan ukuran pita berukuran 240 bp.
Simpulan penelitian adalah P. berghei iradiasi tidak terdeteksi pada hati dan limpa mencit (Mus musculus) menggunakan metode nested-PCR. Hal ini berarti bahwa penggunakan iradiasi efektif melemahkan atau mencegah daya infektif parasit.
v
Puji syukur penulis panjatkan kehadirat Allah SWT atas segala rahmat dan hidayah-Nya sehingga penulis dapat menyelesaikan skripsi dengan judul “Uji Daya Infektifitas Plasmodium berghei Iradiasi Pada Hati dan Limpa Mencit (Mus
musculus) Pasca Booster Menggunakan Metode Nested-Polymerase Chain Reaction
.
” ini.Penyelesaian skripsi ini tidak lepas dari bantuan dan dukungan semua pihak yang terkait. Untuk itu, penulis mengucapkan terima kasih kepada :
1. Dekan Fakultas Matematika dan Ilmu Pengetahuan Alam (FMIPA). 2. Ketua Jurusan Biologi FMIPA Universitas Negeri Semarang.
3. Ibu Ir.Tuti Widianti, M.Biomed selaku dosen pembimbing utama dan Bapak Dr. Mukh Syaifudin selaku dosen pembimbing pendamping yang selalu sabar membimbing dan mengarahkan penulis dalam menyusun rancangan penelitian, pelaksanaan penelitian dan penyusunan laporan hasil penelitian (skripsi) ini. 4. Ibu Dr. drh.R.Susanti, MP. selaku dosen penguji utama yang memberikan
masukan dan saran dalam penyusunan skripsi ini.
5. Bapak dan ibu tercinta, Martawintana dan Samini, yang selalu memberi semangat, dukungan moral dan material tanpa mengenal lelah serta pamrih. 6. Teman-teman BIPANNES ’08 (Ferdi, Vicky, Nidaul, Yulia) yang mewarnai
hidup, menjadi penyemangat dan sumber inspirasi penulis.
7. Semua pihak yang tidak dapat penulis sebutkan satu persatu yang telah membantu penyusunan skripsi ini.
Tiada gading yang tak retak, skripsi ini masih jauh dari sempurna, oleh karena itu saran dan kritik yang membangun penulis harapkan dari pembaca sekalian. Semoga skripsi ini bermanfaat. Amin.
Semarang, September 2013
vi
Halaman
HALAMAN JUDUL ... i
PERNYATAAN KEASLIAN SKRIPSI ... ii
PENGESAHAN ... iii
ABSTRAK ... iv
KATA PENGANTAR ... v
DAFTAR ISI ... vi
DAFTAR TABEL ... viii
DAFTAR GAMBAR ... ix DAFTAR LAMPIRAN ... x BAB I PENDAHULUAN A. Latar Belakang ... 1 B. Rumusan Masalah ... 3 C. Penegasan Istilah ... 4 D. Tujuan Penelitian... 4 E. Manfaat Penelitian... 4
BAB II TINJAUAN PUSTAKA dan HIPOTESIS A. Tinjauan Pustaka ... 5
1. Siklus Hidup Plasmodium berghei... 5
a. Siklus aseksual ... ... 6
b. Siklus seksual ... 7
2. Iradiasi Sinar Gamma ... 8
3. Analisa DNA dengan Teknik Nested-PCR ... 10
a. Molekul DNA ... 10
b. Teknik Nested-Polymerase Chain Reaction ... 12
B. Hipotesis ... 13
BAB III METODE PENELITIAN A. Lokasi dan Waktu Penelitian ... 14
B. Subyek Penelitian ... 14
vii
G. Data dan Metode Pengumpulan Data ... 20
H. Analisa Data ... 20
BAB IV HASIL dan PEMBAHASAN A. Hasil... 21 B. Pembahasan... 24 BAB V PENUTUP A. Kesimpulan ... 31 B. Saran ... 31 DAFTAR PUSTAKA ... 32 LAMPIRAN ... 36
viii
1. Alat penelitian isolasi dan amplifikasi DNA ... 16
2. Bahan Penelitian ... 16
3. Hal amplifikasi DNA pada hati dan limpa mencit ... 20
ix
1. Siklus hidup Plasmodium sp ... 8
2. Gambaran skematis nested-PCR ... 11
3. Produk amplifikasi PCR nested-2 Plasmodium sp ... 12
4. Rancangan penelitian ... 15
5. Elektroforegram genom DNA pada gel agaros 2% ... 21
6. Elektroforegram nested-1 pada gel agaros 2%... 22
x
1. Alat penelitian ... 36
2. Dokumentasi penelitian... 38
3. Surat ijin penelitian ... 39
4. Surat persetujuan penelitian tugas akhir ... 40
1
A. Latar belakang
Malaria merupakan penyakit yang disebabkan oleh parasit jenis protozoa dari genus Plasmodium. Salah satunya adalah Plasmodium berghei yang ditularkan melalui gigitan nyamuk Anopheles betina yang terinfeksi parasit tersebut (Shi et al. 2007). P berghei mempunyai siklus hidup dan morfologi yang sama dengan jenis Plasmodium falciparum yang menyebabkan malaria pada manusia.
Sejauh ini, belum ada vaksin yang efektif mencegah malaria. Penyebabnya antara lain adalah resistensi parasit terhadap obat anti malaria serta siklus perkembangbiakan parasit yang rumit. Selain itu juga dipengaruhi oleh vektor nyamuk Anopheles sp. yang resisten terhadap insektisida. Resistensi Plasmodium terhadap klorokuin terjadi karena ada perubahan membran sel parasit sehingga klorokuin akan dikeluarkan dari sel (Abeku 2007).
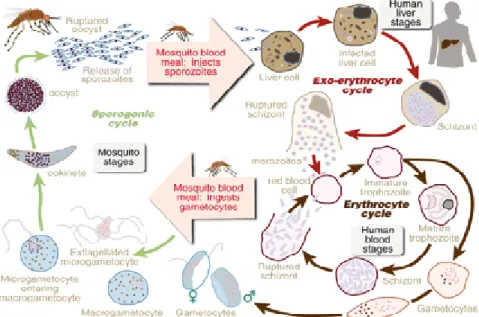
Plasmodium mempunyai 2 siklus hidup yaitu siklus seksual dan aseksual. Plasmodium termasuk parasit obligat karena selalu membutuhkan hospes selama siklus hidupnya. Siklus seksual membutuhkan tubuh nyamuk sebagai hospes
definitif sedangkan mamalia digunakan sebagai hospes intermediate selama
siklus aseksual. Siklus seksual dimulai pada saat nyamuk menelan darah yang terinfeksi Plasmodium (Capurro et al. 2000). Volume darah sekitar 1-2 µl yang ditelan nyamuk betina berisi 1-10 gametosit, 5-6 diantaranya akan menjadi ookinet. Hanya 2 ookinet yang akan berhasil menjadi ookista dalam waktu 2-7 hari dan menghasilkan sekitar 16.000 sporozoit (Sinden 1999). Sporozoit Plasmodium yang terbentuk akan diinfeksikan ke dalam tubuh hospes pada saat nyamuk betina terinfeksi menggigit manusia.
Sporozoit yang diinfeksikan oleh nyamuk akan memasuki aliran darah. Sebagian sporozoit akan difagosit oleh sel darah putih dan sebagian yang lain menginfeksi sel hati. Sporozoit tersebut selanjutnya memperbanyak diri
membentuk skizon yang berisi merozoit. Skizon yang matang pecah kemudian menginfeksi sel darah merah. Akibat aktivitas ini dapat menimbulkan anemia bahkan menyebabkan kematian. Anemia disebabkan oleh merozoit yang mengingesti sitoplasma eritrosit hospes dan mengubah hemoglobin menjadi asam amino dan pigmen hemozoin. Pigmen yang dihasilkan oleh Plasmodium menyebabkan perubahan pada hati dan limpa menjadi coklat kehitaman (Brown 1979).
Limpa merupakan salah satu organ yang penting dalam produksi limfosit. Limpa pada penderita malaria berfungsi sebagai filter untuk menghancurkan eritrosit yang terinfeksi Plasmodium. Plasmodium dan pigmen pada eritrosit difagositosis secara aktif oleh makrofag limpa (Djanah 2007).
Diagnosa Plasmodium secara mikroskopis pada darah penderita malaria di Pakistan, Iran, dan Afghanistan mempunyai tingkat infeksi 0-2,5% sedangkan menggunakan nested-PCR tingkat infeksinya berturut-turut sebesar 6,5%, 22%, dan 35% (Zakeri et al. 2010). Riset yang dilakukan oleh Ndao et al. (2004) menunjukkan bahwa metode nested-PCR lebih akurat dibandingkan dengan metode apusan darah dan deteksi antigen.
Penggunaan sinar gamma untuk melemahkan Plasmodium sebagai bahan dasar vaksin telah banyak diteliti. Keefektifan sinar gamma dalam melemahkan parasit telah dibuktikan dengan adanya imunitas protektif pada hewan coba setelah diimunisasi sporozoit P.falciparum dan P.berghei iradiasi. Dosis radiasi yang optimal dapat menghambat perkembangan parasit. Dosis radiasi optimal untuk melemahkan P. falciparum stadium sporozoit adalah antara 150–200 Gy (Hoffman et al. 2002). Dosis iradiasi 150-175 Gy dapat menurunkan daya infeksi
P.berghei pada mencit dengan ditunjukkan oleh periode prepaten yang panjang
serta jumlah kematian mencit yang rendah (Darlina & Tetriana 2008).
Berdasarkan respon imun yang diteliti yakni pengamatan makrofag pada dosis 150 Gy belum menunjukkan efektivitas bahan vaksin. Berdasarkan pemeriksaan menggunakan elektroforesis gel poliakrilamid menunjukkan bahwa kandungan protein P.berghei semakin menurun dengan kenaikan dosis radiasi
(Tetriana et al. 2008). Perubahan protein dapat diakibatkan oleh denaturasi protein, degradasi protein maupun perubahan konformasi asam deoksiribonukleat (DNA).
Genom P. berghei berukuran 2,3-2,4 x107 pasang basa (pb) (Gardner et
al. 2002). Perubahan DNA dapat dideteksi menggunakan teknik polymerase chain reaction (PCR). PCR merupakan suatu metode invitro untuk
mengamplifikasi segmen DNA dari suatu kompleks DNA melalui suatu reaksi enzimatik yang sederhana dengan menggunakan pasangan primer yang spesifik melalui mekanisme perubahan suhu (Sulandari & Zein 2003).
Nested-PCR merupakan modifikasi dari PCR yang bertujuan mengurangi
kontaminasi produk PCR yang disebabkan oleh kesalahan amplifikasi primer.
Nested-PCR melibatkan dua pasang primer yang digunakan dalam dua proses
PCR yang berurutan. Set primer kedua digunakan untuk memperkuat DNA target PCR nested-1 dan menghasilkan DNA target yang lebih pendek dari produk PCR pertama (Neumaier et al. 1998). Teknik PCR merupakan metode yang umum untuk mendeteksi resistensi Plasmodium secara lebih sensitif. Sejumlah 13 sampel yang diteliti oleh Saiwichai et al. (2009) menggunakan mikroskop menunjukkan hasil negatif, sedangkan 9 sampel diantaranya positif terdeteksi Plasmodium menggunakan nested-PCR.
Selain keakuratan yang tinggi, sensitivitas dari metode nested- PCR juga tinggi seperti penelitian Saiwichai et al. (2009) yang menggunakan pemeriksaan darah segar untuk mendeteksi P. gallinaceum. Tingkat sensitivitas yang didapatkan adalah sebesar 0.0000085% parasitemia atau 0,2 sel darah merah terinfeksi/µl. Penelitian Iqbal et al. (1999) juga menunjukkan bahwa metode PCR lebih sensitif dibandingkan pemeriksaan mikroskopis.
B. Rumusan masalah
Permasalahan dalam penelitian ini adalah sejauh mana daya infektivitas
Plasmodium berghei iradiasi pada hati dan limpa mencit yang dideteksi
C. Penegasan istilah
Untuk menghindari salah pengertian dalam memahami isi skripsi ini, perlu ada batasan-batasan terhadap beberapa istilah sebagai berikut :
1. Plasmodium berghei adalah protozoa penyebab malaria pada rodensia yang digunakan sebagai model dalam penelitian.
2. Iradiasi merupakan teknik radiasi buatan dengan meradiasi bahan dengan sinar radiasi dan yang digunakan adalah sinar gamma dosis 175 Gy dengan laju dosis 380 Gy/jam. Sumber radiasi gamma yang digunakan adalah 60Co. 3. Nested-PCR merupakan modifikasi dari PCR yang bertujuan mengurangi
kontaminasi produk PCR yang disebabkan oleh kesalahan amplifikasi primer.
Nested-PCR melibatkan dua pasang primer yang digunakan dalam dua proses
PCR yang berurutan. Set primer kedua digunakan untuk memperkuat DNA target PCR nested-1 dan menghasilkan DNA target yang lebih pendek dari produk PCR pertama (Neumaier et al. 1998).
D. Tujuan penelitian
Tujuan penelitian ini adalah mendeteksi daya infektivitas Plasmodium
berghei iradiasi pada hati dan limpa mencit menggunakan metode Nested-Polymerase Chain Reaction (PCR).
E. Manfaat penelitian
Manfaat yang dapat diperoleh dari penelitian ini adalah :
1. Sebagai langkah untuk penelitian lebih lanjut tentang pembuatan vaksin malaria melalui iradiasi parasit.
2. Menambah wawasan tentang deteksi parasit P. berghei yang diiradiasi gamma.
5
BAB II
TINJAUAN PUSTAKA & HIPOTESIS
A. Tinjauan Pustaka
Malaria merupakan penyakit yang disebabkan oleh parasit jenis protozoa dari genus Plasmodium. Terdapat 400 spesies Plasmodium, 12 diantaranya bersifat patogen pada manusia. Siklus hidup Plasmodium melibatkan interaksi antara parasit, vektor, dan mamalia. Plasmodium falciparum merupakan salah satu parasit dari genus Plasmodium yang menginfeksi manusia sebagai hospes intermediate. P.
falciparum sering menyebabkan malaria otak dan menyebabkan kematian. Selain
menggunakan manusia sebagai hospes, Plasmodium menggunakan tubuh nyamuk sebagai hospes definitive (vektor) salah satunya adalah nyamuk Anopheles gambiae (Sandoz 1973).
Sejak urutan genom P. falciparum dan A. gambiae diketahui, upaya mengendalikan malaria dari sisi parasit dan vektor secara molekuler sudah mulai dilakukan (Kanzo &Zheng 2003). Sulitnya memahami interaksi antara parasit dengan vektor dan mamalia dikarenakan keragaman spesies parasit dan vektor tersebut (Sinden 2002).
1. Siklus Hidup Plasmodium berghei
P. berghei adalah hemoprotozoa yang menyebabkan penyakit malaria pada
rodensia, terutama rodensia kecil seperti mencit. P. berghei banyak digunakan dalam penelitian malaria pada manusia. Hal ini disebabkan teknologi pembiakan secara
invitro dan pemurnian pada tahapan siklus hidup, dan pengetahuan genom telah
diketahui. P. berghei mempunyai ukuran genom yang paling mirip dengan genom P.
falciparum dibandingkan dengan jenis Plasmodium yang lain (Gardner et al. 2002).
Selain genom, kemiripan sifat biokimiawi dan siklus hidup P. berghei juga digunakan sebagai pertimbangan yang kuat tentang penggunaan parasit tersebut dalam penelitian malaria pada manusia. P. falciparum tidak digunakan dalam penelitian karena parasit tersebut hidup di dalam tubuh manusia sedangkan model
yang digunakan dalam penelitian adalah mencit. Manusia tidak dapat digunakan sebagai model dalam penelitian malaria dikarenakan alasan etika.
Secara umum siklus hidup Plasmodium mempunyai dua hospes yakni manusia dan nyamuk Anopheles. Siklus aseksual berlangsung pada tubuh manusia disebut fase skizogoni dan siklus seksual terjadi di dalam tubuh nyamuk disebut fase
sporogoni (Garnham 1965).
a. Siklus aseksual
Sporozoit merupakan tahapan parasit yang berasal dari kelenjar ludah
nyamuk Anopheles betina masuk ke dalam tubuh manusia melalui gigitan nyamuk tersebut. Sporozoit selanjutnya memasuki sel-sel hati dan dimulailah stadium
eksoeritrositik dalam waktu tiga puluh menit. Sporozoit yang berada di sel hati
tumbuh menjadi skizon dan berkembang menjadi merozoit (10.000-30.000 merozoit, tergantung spesiesnya). Sel hati akan pecah dan merozoit akan masuk aliran darah (Kappe et al. 2003).
Penelitian Frevert et al. (2005) menunjukkan bahwa sporozoit P.berghei secara aktif menyerang sel makrofag. Hal ini diperankan oleh tethers yaitu sporozoit yang aktif menginvasi makrofag. Proses terbentuknya tethers terjadi saat invasi sporozoit ke dalam sel-sel yang melapisi lumen sinusoid hati (Vandenberg et al. 1990).
Motilitas sporozoit berperan penting saat invasi sporozoit ke dalam sel-sel hati inang (Vandenberg et al. 1990). Sporozoit masuk ke hati secara tiba-tiba mengikuti lapisan sel sinusoidal. Parasit masuk melalui sel sinusoidal searah atau melawan aliran darah menuju sel Kupffer dan melintasi ruang Disse. Sel Kupffer dapat dideteksi menggunakan lisosom autoflourescensi berwarna orange. Sporozoit akan mencapai hepatosit dalam waktu beberapa menit( Frevert et al. 2005).
Tahap perkembangan parasit di dalam hati menghasilkan bentuk parasit baru. Parasit tersebut hidup di dalam sel darah merah dengan lingkungan seluler dan molekuler yang berbeda (Mikolajczak et al. 2006). Parasit yang lemah atau mati berbentuk sabit dan tidak menunjukkan gerakan sel. Tiga jam setelah infeksi sekitar 5x106 sporozoit, sekelompok hepatosit mengalami nekrosis yang akan diinfiltrasi
oleh sel inflamatori. Infeksi sporozoit pada hepatosit menyebabkan kerusakan hepatosit dan ditandai oleh tingginya jumlah alanine aminotransferase di dalam serum.
Plasmodium tidak hanya menyebabkan kerusakan pada hati tetapi juga menyebabkan pembesaran limpa (Brown 1979). Limpa merupakan kelenjar tanpa saluran yang berhubungan erat dengan sistem sirkulasi. Limpa mempunyai dua fungsi yaitu membentuk respon imun dan melawan antigen yang berada di dalam darah. Limpa akan membuang bahan partikel dan sel darah yang sudah tua atau rusak, terutama eritrosit dari sirkulasi. Sistem sirkulasi darah pada limpa memiliki fungsi penting terhadap rangsangan antigen dan ekstraksi hemoglobin serta zat besi.
Limpa berperan penting dalam mengatasi infeksi malaria (clearance).
Cytoadherence merupakan salah satu cara Plasmodium untuk menghindari clearance
limpa dan digunakan sebagai mekanisme pertahanan Plasmodium untuk menghindari sirkulasi darah. Cytoadherence menyebabkan oklusi pembuluh darah limpa dan kerusakan organ limpa (Mohanty et al. 2006).
b. Siklus seksual
Siklus seksual terjadi di dalam tubuh nyamuk nyamuk Anopheles betina menghisap darah yang mengandung gametosit. Pada mikrogamet (jantan), sejumlah 6-8 inti sel akan bergerak ke tepi sel mikrogamet dan membentuk flagel atau filamen. Pembuahan terjadi bila mikrogamet masuk ke dalam makrogamet sehingga terbentuk
zigot. Zigot berubah bentuk seperti cacing pendek disebut ookinet. Ookinet dapat
menembus lapisan epitel dan membran basal dinding lambung. Di tempat ini ookinet akan berkembang menjadi ookista (Garnham 1965).
Riset yang dilakukan oleh Wijayanti et al. (1997) menunjukkan bahwa infeksi
P. berghei pada mencit Swiss dapat menyebabkan kematian pada hospes. Imunitas
hospes muncul karena adanya sistem pada mencit. Imunitas mencit yang lebih tinggi menyebabkan penurunan angka mortalitas mencit dan jumlah parasitemia yang rendah. Patogen pada manusia dideteksi oleh sel-sel dari sistem imun innate seperti sel dendritik, sel Natural Killer( NK), basofil, eosinofil, dan sel Mast melalui pattern
receptors (NLRs) (Takeuchi & Akira 2010). Interaksi antara hospes dan parasit ini dapat mengurangi jumlah parasit dan merangsang sistem immune adaptive( sel B dan sel T) yang akan mengenali dan mengikat antigen asing melalui ekspresi reseptor pada permukaan sel ( Palm & Medzhitov 2009). Siklus hidup Plasmodium dapat dilihat pada Gambar 1 berikut.
Gambar 1 Siklus hidup Plasmodium sp
Kematian sel terprogam (apoptosis) pada awal perkembangan Plasmodium di dalam tubuh vektor digunakan untuk regulasi infeksi parasit. Hal ini akan mempengaruhi kelangsungan hidup parasit. Zigot dan ookinet P.berghei yang sudah mati ditunjukkan dengan peningkatan apoptosis sel di dalam lumen midgut. Infeksi Plasmodium menginduksi terjadinya apoptosis pada sel-sel dari jaringan midgut dan epitel folikuler (Hurd &Carter 2004).
2. Iradiasi sinar gamma
Radiasi adalah energi yang dipancarkan dalam bentuk partikel atau gelombang. Iradiasi merupakan radiasi yang sengaja dilakukan oleh manusia untuk tujuan tertentu. Radiasi pengion merupakan salah satu bentuk radiasi yang menyebabkan munculnya partikel bermuatan listrik. Radiasi sinar-X, sinar gamma,
alfa, dan beta termasuk jenis radiasi pengion. Radiasi pengion berinteraksi dengan sistem seluler melalui pengubahan struktur, fungsi, dan respon sel terhadap produk seluler (Nikjoo 2003).
Radiasi sinar gamma pada sel mempunyai efek secara tidak langsung (stokastik) maupun langsung (deterministik). Efek langsung dapat ditandai dengan kematian sel ataupun pemutusan ikatan senyawa penyusun sel. Efek tidak langsung terjadi karena sebagian besar penyusun sel adalah air. Radiasi gamma menyebabkan air terhidrolisis menghasilkan radikal bebas yang akan merusak sel termasuk DNA pada sel. Interaksi radiasi dengan DNA menyebabkan kerusakan DNA seperti single
strand break (SSB), double strand break (DSB), base damage (BD) dan lain-lain.
Kerusakan tingkat molekul disebabkan oleh deposisi energi, produksi ionisasi, eksitasi oleh molekul yang berasal dari reaksi fisika dan spesies radikal, produk molekul yang lain yang berasal dari reaksi kimia (Nikjoo 2003).
Sinar gamma sudah banyak dimanfaatkan dalam berbagai bidang penelitian. Iradiasi sinar gamma dimanfaatkan dalam bidang pangan seperti menginaktivasi bakteri patogen pada produk makanan (Gunes et al. 2011). Selain itu iradiasi sinar gamma juga dimanfaatkan dalam bidang vaksin. Penelitian Sanakkalaya et al (2005) menunjukkan bahwa iradiasi gamma dapat menurunkan kemampuan replikatif bakteri yang bisa digunakan sebagai alternatif vaksin untuk menginduksi imunitas protektif dari infeksi Brucella abortus. Sinar gamma juga dimanfaatkan dalam penelitian bahan vaksin mastitis dengan cara diinaktivasi tanpa merusak protein secara keseluruhan (Hermanto et al. 2008 ). Teknik radiasi pengion digunakan untuk pembuatan vaksin malaria untuk mencegah infeksi P. berghei pada stadium eritrositik (Tetriana et al. 2008). Hal ini disebabkan teknik iradiasi lebih menguntungkan karena respon imun lebih kuat dan luas dalam inang setelah pemberian vaksin.
Pemanfaatan radiasi pengion dalam pembuatan vaksin malaria telah dilakukan. Tujuan utamanya adalah menentuan dosis yang tepat untuk melemahkan Plasmodium. Dosis radiasi gamma 150 Gy menyebabkan perubahan profil protein yaitu hilangnya protein pada kisaran 15 kDa (Tetriana et al. 2008). Iradiasi dengan dosis 150-200 Gy merupakan stadium paling efektif untuk melemahkan Plasmodium
pada stadium sporozoit. Hal ini dibuktikan oleh berkurangnya kemampuan infeksi Plasmodium sampai ke sel-sel hati.
3. Analisis DNA dengan Teknik Nested-PCR
a. Molekul DNA
DNA adalah asam nukleat yang mengandung materi genetik dan berfungsi untuk mengatur perkembangan biologis seluruh bentuk kehidupan secara seluler. DNA di dalam sel berupa DNA mitokondria, DNA kloroplas dan DNA inti. Keseluruhan DNA yang menyusun masing- masing komponen disebut DNA genom (Muladno 2002).
P. berghei memiliki 14 kromosom. Secara analisis molekuler P. berghei sama
seperti Plasmodium yang menginfeksi manusia. P. berghei mempunyai genom berukuran 2,3-2,4 x107 pb. DNA inti P. berghei mengandung (A+T) yang tinggi sekitar 82% yang tersebar pada DNA koding dan DNA non-koding. Genom P.
berghei mengandung (G+C) sekitar 25-30 %. Plasmodium mempunyai sekitar 5300
gen yang mengkode berbagai protein yang berperan dalam metabolisme, transpor materi organik, replikasi-perbaikan-rekombinasi DNA dan lain-lain. Selain itu parasit mempunyai gen pengkode enzim sebagai biokatalis dan transport protein (Gardner et
al. 2002).
b. Teknik Nested-Polymerase Chain Reaction
Teknik PCR telah digunakan untuk mendeteksi berbagai penyakit infeksi (Sulistyaningsih 2007). Nested-PCR merupakan metode amplifikasi sekuens DNA yang spesifik secara invitro. Teknik ini menggunakan dua kali proses amplifikasi dengan 2 pasang primer yang spesifik. Pasangan primer pertama berfungsi menggandakan fragmen seperti PCR standar, sedangkan pasangan primer kedua berfungsi untuk memperkuat suatu fragmen DNA produk PCR pertama. Keuntungan dari nested PCR adalah jika terdapat kesalahan amplifikasi fragmen, maka akan diamplifikasi untuk kedua kalinya oleh sepasang primer pada nested-2.
Amplifikasi metode PCR memerlukan beberapa macam bahan yaitu: a) primer, suatu oligonukleotida tunggal yang sekuensnya berkomplemen dengan
cetakan DNA dan panjangnya antara 18-30 basa, b) enzim Taq DNA polymerase, c) dNTPs (dATP, dCTP, dGTP, dTTP), DNA target, akuabides, dan buffer PCR yang di dalamnya sudah terdapat garam magnesium (Sulandari & Zein 2003). Prinsip PCR ada tiga, yaitu denaturasi yang berarti pemutusan untai ganda menjadi untai tunggal, annealing yakni penempelan primer pada tempat yang spesifik dan elongasi yaitu pemanjangan primer dengan bantuan enzim DNA polymerase membentuk untaian DNA. Ilustrasi PCR dapat dilihat pada gambar 2.
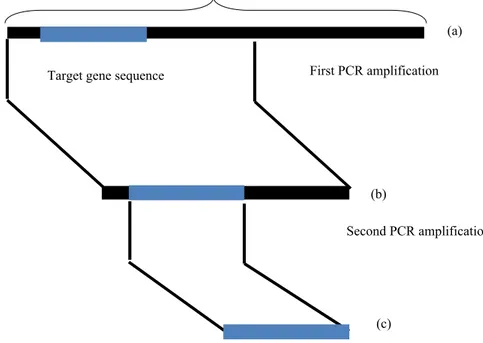
Gambar 2 Gambaran skematis nested-PCR( Pooe 2011)
Nested-PCR adalah teknik yang lebih akurat dibandingkan pemeriksaan
mikroskopis. Nurhayati et al. (2009) mengkaji tentang penggunakan mikroskop yang selama ini menjadi standar emas (gold standar) dalam pemeriksaan Plasmodium yang dinilai sekarang kurang akurat. Diagnosis parasit tidak cukup hanya mengandalkan teknik mikroskopis saja dikarenakan perubahan morfologi dan munculnya berbagai strain baru yang disebabkan obat anti-malaria yang digunakan secara tidak tepat sehingga parasit menjadi resisten terhadap obat.
(a) First PCR amplification Target gene sequence
(b)
(c)
Second PCR amplification Whole genom
Berbagai jenis Plasmodium dapat dibedakan dalam beberapa tahap perkembangannya. Plasmodium dapat dibedakan melalui bentuk skizon, trofozoit, dan bentuk gametosit. Setiap Plasmodium mempunyai bentuk spesifik tetapi pemeriksaan mikroskopis masih cukup sulit dan membutuhkan ketelitian yang tinggi. Selain pemeriksaan mikroskopis juga digunakan metode immunokromatografi yang lebih mudah, cepat dan ekonomis dibandingkan secara mikroskopis tetapi keakuratannya masih kurang jika dibandingkan dengan PCR (Arum et al. 2006). Contoh elektroforegram nested-2 dapat dilihat pada Gambar 3.
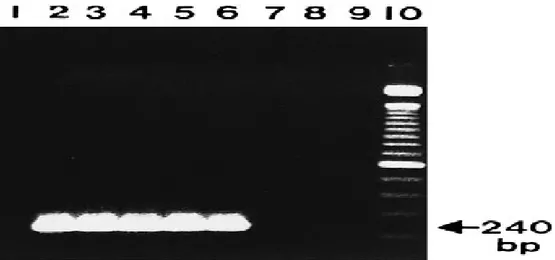
Gambar 3 Produk amplifikasi nested-2 Plasmodium sp dari sampel darah (Singh et al. 1999)
Metode lain untuk mendeteksi protein atau asam nukleat telah dikembangkan. Teknik dip-stick digunakan untuk mendeteksi secara immunoenzimatik protein yang kaya histidin II yang spesifik pada P. falciparum. Selain itu deteksi berdasarkan asam nukleat yakni hibridisasi DNA atau RNA berlabel yang sensivitasnya ditingkatkan dengan PCR juga dikembangkan. Kelebihan menggunakan PCR adalah dapat mendeteksi Plasmodium dalam tingkat infeksi ringan dengan hasil yang lebih akurat (Saiwichai et al. 2009).
B. Hipotesis
Berdasarkan tinjauan pustaka dan permasalahan tersebut di atas, maka hipotesisnya adalah bahwa iradiasi sinar gamma dapat menurunkan daya infektivitas
P. berghei pada tubuh mencit sehingga tidak terdeteksi saat diuji menggunakan nested-Polymerase chain Reaction.
14
A. Lokasi dan Waktu Penelitian
Penelitian dilakukan di Laboratorium Biologi Molekuler, Bidang Biomedika, Pusat Teknologi Keselamatan dan Metrologi Radiasi (PTKMR), Badan Tenaga Nuklir Nasional (BATAN), Jl. Lebakbulus Raya No. 49 Pasar Jum’at ,Jakarta Selatan, mulai bulan Mei sampai Agustus 2012.
B. Subyek Penelitian
Mencit yang digunakan dalam penelitian ini adalah mencit strain Swiss
Webster jantan. Sampel yang digunakan yaitu mencit strain Swiss Webster jantan
berumur ±2 bulan dengan berat badan sekitar 35 gram, diperoleh dari Pusat Penyakit Tropis, Badan Litbang Kesehatan, Kementerian Kesehatan Jakarta.
C. Variabel Penelitian
1. Variabel bebas: dosis radiasi dan nested-PCR. 2. Variabel Tergantung: daya infektivitas mencit 3. Variabel kendali: umur dan berat badan mencit.
D. Rancangan Penelitian
Penelitian ini adalah penelitian eksperimental untuk menguji daya infektivitas P. berghei iradiasi. Penelitian dilakukan dengan menggunakan dosis radiasi 0 Gy dan 175 Gy. Dosis 175 Gy dapat menurunkan daya infeksi P.
berghei pada mencit. Hati dan limpa diambil dari mencit yang telah diinfeksi P. berghei 0 Gy satu kali suntikan, dosis 175 Gy satu kali suntikan dan dosis 175 Gy
dua kali suntikan (booster). Suntikan kedua dilakukan untuk meningkatkan daya imun mencit sehingga diharapkan P. berghei dapat dinetralisir oleh sistem imun yang dipicu parasit iradiasi. Bagian organ yang diuji diambil dari 3 titik yang berbeda dan dianggap sebagai 3 kali ulangan. Kontrol yang digunakan adalah ekstrak DNA P. berghei yang didapatkan dari Laboratorium Malaria, Lembaga Biologi Molekuler Eijkman Jakarta. Rancangan penelitian disajikan pada Gambar 4.
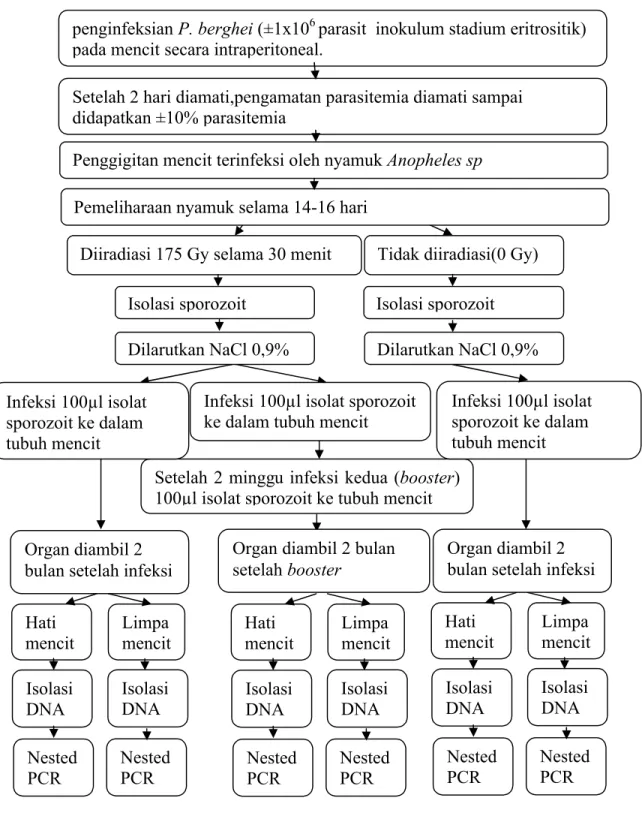
Gambar 4 Rancangan penelitian.
Tidak diiradiasi(0 Gy) Diiradiasi 175 Gy selama 30 menit
Setelah 2 minggu infeksi kedua (booster) 100µl isolat sporozoit ke tubuh mencit
Hati mencit Limpa mencit Isolasi DNA Organ diambil 2 bulan setelah infeksi Infeksi 100µl isolat sporozoit ke dalam tubuh mencit
Organ diambil 2 bulan setelah booster
Organ diambil 2 bulan setelah infeksi
Nested PCR Isolasi DNA Nested PCR
penginfeksian P. berghei (±1x106 parasit inokulum stadium eritrositik) pada mencit secara intraperitoneal.
Setelah 2 hari diamati,pengamatan parasitemia diamati sampai didapatkan ±10% parasitemia
Penggigitan mencit terinfeksi oleh nyamuk Anopheles sp Pemeliharaan nyamuk selama 14-16 hari
Isolasi sporozoit Isolasi sporozoit Dilarutkan NaCl 0,9% Dilarutkan NaCl 0,9%
Infeksi 100µl isolat sporozoit ke dalam tubuh mencit
Infeksi 100µl isolat sporozoit ke dalam tubuh mencit
Hati
mencit Limpamencit Isolasi DNA Nested PCR Isolasi DNA Nested PCR Hati mencit Limpa mencit Isolasi DNA Nested PCR Isolasi DNA Nested PCR
E. Alat dan Bahan Penelitian
Alat dan bahan yang digunakan dalam penelitian berturut-turut disajikan pada Tabel 1:
Tabel 1 Alat penelitian
No Uraian Alat
1. Pengambilan sampel organ Alat bedah, pinset, plastik steril 2. Penyimpanan sampel organ Freezer (-20°C)
3. Isolasi DNA Latex glove, pisau silet, pinset, neraca Ohaus Pioneer TM, microtube 1,5 ml, rak
microtube, dry bath, microcentrifuge survall Legend MICRO 17 R, kotak sampel, tip kuning & biru, lemari pendingin Sharp(-20°C), inkubator dan vortex Labnet®, mikropipet Eppendorf, tabung QIAamp Mini Spin.
4. Elektroforesis hasil isolasi dan amplifikasi DNA
Rak microtube, well forming combs, horizontal elektroforesis Mupid ®-eXu, Gel docTM XR+ with Image LabTM soft
ware, Hot plate Cimarec®, tip kuning & putih, mikropipet Eppendorf .
5. Amplifikasi DNA GeneAmp® PCR System 9700 Applied Biosystem, microtube 0,2 µl, mikropipet Eppendorf, tip kuning& putih, microcentrifuge survall Legend MICRO 17 R, vortex.
Tabel 2 Bahan Penelitian
No Uraian Bahan
1. Isolasi DNA Tissue Lysis Buffer, Proteinase K, Lysis Buffer, Wash Buffer (1), Wash Buffer (2), Elution Buffer, Etanol Absolut, Etanol 80%, RNase, Tris-EDTA.
2. Elektroforesis hasil isolasi dan amplifikasi DNA
UltrapureTM Agarose InvitrogenTM , EtBr
aMResco®, Bufer TBE 0,5X, loading buffer Invitrogen®, DNA ladder aMResco®, parafilm M®, shaker DRS-12
3. Amplifikasi DNA Primer (rPLU1, rPLU5, Primer rPLU3, rPLU4) Invitrogen®, dNTPs Applied Biosystem®, Enzim taq polymerase Applied Biosystem®, ddH2O, MgCl2 Applied
F. Prosedur Penelitian
a. Perolehan sporozoit dan isolasinya.
Sporozoit diperoleh dengan terlebih dahulu melakukan penginfeksian secara intraperitoneal P. berghei (±1x106 parasit inokulum stadium eritrositik) pada mencit dan 2 hari kemudian diamati parasitemia dalam darah mencit setiap hari dengan mengambil darah perifer dari ujung ekor. Setelah diperoleh parasitemia ±10%, mencit terinfeksi diletakkan dalam kandang nyamuk
Anopheles sp. dan dibiarkan nyamuk mengigit mencit. Nyamuk yang
mengkonsumsi darah (terinfeksi) dipelihara selama 14-16 hari dalam kandang khusus untuk memperoleh sporozoit. Nyamuk tersebut diiradiasi sinar gamma dosis 0 dan 175 Gy selama 30 menit. Isolasi kelenjar ludah nyamuk yang mengandung sporozoit dilakukan dengan membedah nyamuk menurut prosedur standar. Isolat dilarutkan dalam NaCl 0,9%. Isolat kelenjar ludah kemudian disuntikkan secara intravena pada 3 mencit sehat melalui ekornya sebanyak 100µl setiap kali penyuntikan. Untuk beberapa perlakukan, penyuntikan kelenjar ludah mengandung sporozoit ini diulangi 2 minggu kemudian (booster).
b. Isolasi DNA hati dan limpa mencit.
Isolasi DNA dilakukan menggunakan kit QIAGEN. Sampel hati 25 mg dan limpa 10 mg dipotong-potong menjadi bagian yang lebih kecil menggunakan pinset. Sampel dimasukkan ke dalam tabung 1,5 ml dan ditambahkan 180 µl
tissue lysis buffer. Selanjutnya sampel ditambahkan 20 µl Proteinase-K, divorteks,
dan diinkubasi pada suhu 56°C selama 1 jam sampai jaringan mengalami lisis. Selama waktu inkubasi dilakukan 2-3 kali vorteks . Pengendapan pelet dilakukan dengan cara disentrifus sebentar. Kemudian ditambahkan 4 µl RNA-se (100 mg/ml), divorteks 15 detik dan diinkubasi 2 menit pada suhu kamar. Kemudian pada tabung ditambahkan 200 µl lysis buffer, lalu divorteks lagi selama 15 detik, diinkubasi pada suhu 70°C selama 10 menit. Selanjutnya tabung ditambahkan 200 µl etanol absolut, divorteks lagi selama 15 detik, disentrifus sebentar kemudian
dipindahkan dengan hati-hati ke dalam tabung QIAamp Mini Spin dan disentrifus pada kecepatan 8000 rpm selama 1 menit.
Supernatan yang dihasilkan dipindahkan ke QIAamp Mini spin kolom baru. Selanjutnya supernatan ditambahkan 500 µl buffer Wash buffer(1) tanpa membasahi dinding, disentrifus pada kecepatan 8000 rpm selama 1 menit. Hasilnya dipindahkan ke dalam pada tabung QIAamp Mini spin yang bersih. Kemudian supernatan ditambahkan 500 µl wash buffer(2) tanpa membasahi dinding, disentrifus pada kecepatan 14000 rpm selama 3 menit. Supernatan yang dihasilkan dipindahkan pada tabung QIAamp Mini spin yang bersih dan disentrifus lagi pada kecepatan 14000 rpm selama 1 menit. Pada tahap ini sudah didapatkan ekstrak DNA. Tabung QIAamp Mini spin ditempatkan pada tabung 1,5 ml kemudian ditambahkan 80 µl elution buffer dan diinkubasi pada suhu ruang selama 5 menit, disentrifus pada kecepatan 8000 rpm selama 1 menit. Ekstrak DNA yang sudah didapatkan kemudian disimpan pada suhu -20°C.
c. Amplifikasi DNA
Tahap PCR untuk deteksi Plasmodium menggunakan 2 kali PCR yakni
nested-1 dan nested-2. Nested-2 dilakukan setelah diketahui hasil PCR pada nested-1. Primer yang digunakan untuk nested-1 adalah rPLU1 & rPLU5 dengan
ukuran panjang DNA target sebesar 1640 bp (Michael 2005) sedangkan primer pada nested-2 adalah rPLU3 & rPLU4 dengan ukuran panjang DNA target sebesar 240 bp (Singh et al.1999). Produk PCR adalah gen 50S ribosomal sub unit L21.
PCR nested-1 menggunakan primer rPLU1 sebagai primer F dengan panjang 24 basa dan urutan basa 5’-TCA AAG ATT AAG CCA TGC AAG TGA-3’, sedangkan rPLU 5 sebagai primer R dengan panjang 21 basa dan urutan basanya adalah 5’-CCT GTT GTT GCC TTA AAC TCC-3’. Pada Nested-2, rPLU3 sebagai primer F mempunyai panjang 30 basa dengan urutan 5’-TTT TTA TAA GGA TAA CTA CGG AAA AGC TGT-3’ sedangkan rPLU4 sebagai
primer R mempunyai panjang 30 basa dengan urutan 5’-TAC CCG TCA TAG CCA TGT TAG GCC ATT ACC-3’.
Langkah awal adalah membuat larutan mix PCR dengan komposisi sebagai berikut. Perbandingan distillated water adalah 17,75 µl; Buffer A sebanyak 2,5µl ; MgCl21µl; dNTPs 0,5µl; primer F 0,25µl; primer R 0,25µl; Taq
polymerase 0,25µl.
Semua komponen yang sudah dicampur di dalam tabung kemudian divorteks dan disentrifus sebentar. Setiap tabung PCR diisi campuran tersebut sebanyak 22,5 µl. Sebanyak 3 µl ekstrak DNA dimasukkan ke dalam masing-masing tabung PCR dan diberi label. Tabung divorteks sebentar agar tidak ada komponen yang menempel pada dinding tabung. Kemudian semua tabung dimasukkan ke dalam mesin PCR (GeneAmp® PCR System 9700 Applied Biosystem) yang sudah dihidupkan terlebih dahulu.
Kondisi PCR yang digunakan adalah sebagai berikut. Kondisi PCR untuk nested-1:
a) Pre denaturasi pada suhu 94°C selama 4 menit.
b) Siklus sebanyak 29 kali terdiri denaturasi pada suhu 94°C selama 30 detik, annealing pada suhu 55°C selama 1 menit, dan elongasi pada suhu 72°C selama 1 menit.
c) Post elongasi pada suhu 72°C selama 4 menit.
Kondisi untuk nested-2:
a) Pre denaturasi pada suhu 94°C selama 4 menit
b) Siklus sebanyak 30 kali terdiri dari denaturasi pada suhu 94°C selama 30 detik, annealing pada suhu 62°C selama 1 menit, dan elongasi pada suhu 72°C selama 1 menit
d. Elektroforesis gel agaros hasil isolasi dan amplifikasi DNA
Elektroforesis dilakukan pada ekstrak DNA dan sampel produk PCR. menggunakan gel agaros 2%. Produk ekstraksi DNA dielektroforesis dengan perbandingan sampel : loading buffer adalah 4: 2. Produk PCR dielektroforesis dengan perbandingan sampel : loading buffer adalah 8: 3. Sampel yang sudah dicampur dengan loading buffer dimasukkan ke dalam sumur gel agaros yang direndam dalam TBE 0,5 X. Gel kemudian di-running selama 30 menit pada 50 Volt. Selanjutnya gel direndam dengan larutan yang mengandung ethidium
bromide (EtBr) selama 15 menit sambil digoyang menggunakan shaker dan
direndam dengan aquades sambil digoyang selama 10 menit. Visualisasi pita dilakukan DNA menggunakan Gel Doc yang sudah dilengkapi software Image Lab.
G. Data dan Metode Pengumpulan Data
Data dalam penelitian ini diperoleh berdasarkan hasil pengamatan panjang fragmen pita-pita DNA produk PCR. Jika terdapat pita DNA spesifik dari sampel yang teramplifikasi dan DNA target berukuran sesuai dengan primer yang digunakan, maka sampel tersebut mengandung parasit P. berghei. Data pada hati dan limpa yang didapatkan dimasukkan ke dalam Tabel 3.
Tabel 3 Hal amplifikasi DNA pada hati dan limpa mencit.
No Dosis Ulangan I Ulangan II Ulangan III
Kontrol positif
Kontrol negatif Hati Limpa Hati Limpa Hati Limpa Hati Limpa Hati Limpa 1. 0 Gy
2. 175 Gy 3. 175Gy booster
H. Analisa Data
Pita-pita DNA yang tampak pada gel hasil elektroforesis dibandingkan dengan pita DNA pada kontrol positif dan dianalisis secara deskriptif kualitatif untuk menentukan ada tidaknya P. berghei pada organ hati dan limpa.
21
A. Hasil Penelitian
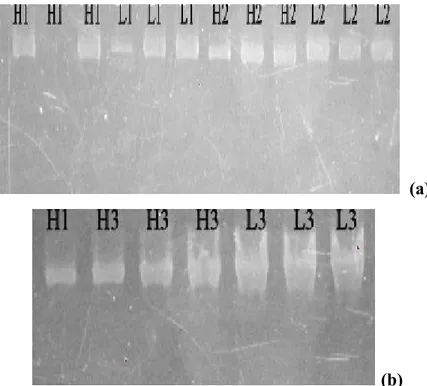
Pengambilan sampel dilakukan pada 3 titik yang berbeda pada hati dan limpa secara acak. Hal tersebut dianggap sebagai 3 kali ulangan isolasi DNA pada masing-masing sampel. Sebelum dilakukan amplifikasi DNA dengan nested-PCR, dilakukan pemeriksaan terhadap kandungan DNA genom pada semua sampel hasil isolasi. Keberadaan/kandungan isolat DNA kemudian dicek menggunakan elektroforesis gel agaros 2%. Elektroferogram hasil isolasi DNA disajikan pada Gambar 5. Tampak bahwa isolasi DNA menggunakan kit dari hati dan limpa berhasil dengan baik, kecuali H1 (Hati 0 Gy) (Gambar 5a).
(a)
(b)
Gambar 5. Elektroforegram genom DNA pada gel agarose 2%. H1, Hati 0 Gy; L1, Limpa 0 Gy; H2, Hati 175 Gy dengan booster; L2, Limpa 175 Gy dengan booster; H3, Hati 175 Gy tanpa booster; dan L3, 175 Gy tanpa booster.
Hasil isolasi DNA dari sampel hati dan limpa kemudian diamplifikasi menggunakan nested-PCR. Metode ini telah banyak digunakan pada penelitian
penyebab penyakit infeksi seperti Plasmodium (Sulistyaningsih 2007). Nested-PCR adalah jenis Nested-PCR yang menggunakan dua kali proses Nested-PCR. Hasil Nested-PCR
nested-1 dicek menggunakan elektroforesis gel agaros 2%. Hasil amplifikasi pada nested-1 mempunyai ukuran DNA target yang lebih besar dari nested-2 yaitu
sebesar 1640 bp (Michael 2005). Gen targetnya adalah 50S ribosomal protein L21. Amplifikasi PCR menggunakan ekstrak DNA P. berghei sebagai kontrol positif. Salah satu sumur pada gel agaros hanya diisi dengan mix PCR sebagai kontrol negatif yang bertujuan untuk mengetahui ada tidaknya kontaminan selama proses amplifikasi berlangsung. Elektroforegram hasil PCR nested-1 disajikan pada Gambar 6.
(a)
(b)
Gambar 6 Elektroforegram nested-1 pada gel agarose 2%. M. Marker, H1. Hati 0Gy, L1. Limpa 0 Gy, H2. Hati 175 Gy dengan booster, L2. Limpa 175 Gy dengan booster, H3. Hati 175 Gy tanpa booster, L3. 175 Gy tanpa
booster, (K+) kontrol positif, (K-) kontrol negatif.
Data hasil PCR nested-1(Gambar 6) diketahui bahwa tidak ada DNA yang muncul dan terdeteksi pada gel agarose. Hal ini disebabkan karena terlalu rendah/sedikitnya hasil DNA yang diamplifikasi. Ukuran DNA ini sangat besar yakni 1640 bp (base pair).
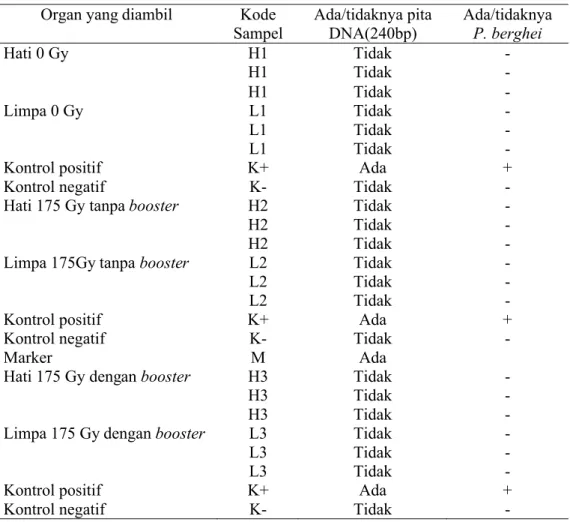
Hasil amplifikasi DNA yang diperoleh dari nested-1 selanjutnya diamplifikasi dengan PCR nested-2. PCR nested-2 digunakan untuk memfokuskan wilayah amplifikasi pada nested-1. DNA target pada PCR nested-2 mempunyai ukuran yang lebih kecil daripada nested-1 yaitu sebesar 240 bp (Singh et al.1999). Gen targetnya adalah 50S ribosomal protein L21. PCR nested-1 dan nested-2 mempunyai gen target sama yaitu 50S ribosomal protein L21 karena nested-PCR adalah PCR bersarang yaitu daerah produk PCR nested-2 berada di dalam produk PCR nested-1. Produk PCR nested-1 merupakan daerah genus Plasmodium sedangkan produk PCR nested-2 adalah daerah spesies spesifik. Elektroforegram hasil PCR nested-2 menggunakan gel agaros 2% disajikan pada Gambar 7 & Tabel 4. (a) (b) (c)
Gambar 7 Elektroforegram nested-2 pada gel agaros 2%. M. Marker, (a) H1. Hati 0Gy, L1. Limpa 0 Gy , (b) H2. Hati 175 Gy dengan booster, L2. Limpa 175 Gy dengan booster, (c) H3. Hati 175 Gy tanpa booster, L3. 175 Gy tanpa booster, (K+) kontrol positif, (K-) kontrol negatif.
240 bp 240
Tabel 4. Data elektroforegram hasil PCR nested-2
Organ yang diambil Kode Sampel Ada/tidaknya pita DNA(240bp) Ada/tidaknya P. berghei Hati 0 Gy H1 Tidak -H1 Tidak -H1 Tidak -Limpa 0 Gy L1 Tidak -L1 Tidak -L1 Tidak
-Kontrol positif K+ Ada +
Kontrol negatif K- Tidak
-Hati 175 Gy tanpa booster H2 Tidak
-H2 Tidak
-H2 Tidak
-Limpa 175Gy tanpa booster L2 Tidak
-L2 Tidak
-L2 Tidak
-Kontrol positif K+ Ada +
Kontrol negatif K- Tidak
-Marker M Ada
Hati 175 Gy dengan booster H3 Tidak
-H3 Tidak
-H3 Tidak
-Limpa 175 Gy dengan booster L3 Tidak
-L3 Tidak
-L3 Tidak
-Kontrol positif K+ Ada +
Kontrol negatif K- Tidak
-Keterangan:(+) = ada pendaran pita DNA ; (-) = tidak ada pendaran pita DNA
B. Pembahasan
1. Pemeriksaan DNA genom
DNA diperiksa menggunakan teknik elektroforesis yang akan menunjukkan pita DNA pada gel dalam larutan penyangga. Migrasi pita DNA pada pH netral bergerak dari kutub negatif menuju kutub positif. Pita DNA berukuran besar berjalan lebih lambat dibanding DNA berukuran kecil. Hasil ekstrak DNA yang didapatkan mempunyai ukuran DNA yang besar karena merupakan DNA genom sampel hati dan limpa mencit. Ukuran DNA genom yang didapatkan tidak diukur sehingga tidak diketahui besarnya ukuran genom hati dan limpa mencit. Selain ukuran DNA, kecepatan migrasi DNA dipengaruhi oleh
konsentrasi agarose, voltase, ethidium bromide dan komposisi larutan bufer (Muladno 2002).
Pemeriksaan ini dilakukan untuk mengetahui kualitas DNA yang telah disimpan sebelum dilakukan proses PCR. DNA berkualitas baik ditunjukkan oleh pita yang kompak dan tidak terdapat smear. Smear merupakan DNA yang terpotong-potong dan berukuran kecil. Smear juga dapat disebabkan oleh kemurnian DNA hasil isolasi yang rendah. Hasil pemeriksaan DNA dalam penelitian ini menunjukkan bahwa sebagian besar sampel terdapat smear. Salah satu sampel hati 0 Gy tidak terdapat DNA sehingga isolasi DNA pada sampel tersebut diulang kembali (Gambar 5). Kualitas DNA akan mempengaruhi DNA target yang diinginkan.
Kualitas koleksi DNA yang baik dapat disebabkan adanya beberapa faktor, antara lain penyimpanan DNA menggunakan buffer dan pada suhu dibawah -20oC. Penyimpanan ekstrak DNA biasanya menggunakan elution buffer. Penyimpanan DNA dalam jangka waktu yang lama dapat mempengaruhi kualitas DNA. Penyimpanan pada suhu dingin atau kondisi beku lebih efektif untuk mempertahankan DNA, seperti dinyatakan oleh Zetzsche dan Gemeinholzer (2009) bahwa penyimpanan suhu -20oC lebih baik dibandingkan suhu -4oC.
2. Amplifikasi P. berghei iradiasi pada hati dan limpa mencit tahap nested-1
Nested-PCR merupakan proses amplifikasi ganda menggunakan dua
pasang primer. Tahapan nested-PCR terdiri dari nested-1 dan nested-2. Nested-1 menggunakan primer rPLU1 dan rPLU5 sedangkan nested-2 menggunakan pasangan primer rPLU3 dan rPLU4. Primer ini merupakan titik awal dimulainya penggandaan DNA dalam proses PCR hingga didapatkan segmen DNA dengan ukuran 1640 bp untuk nested-1 dan 240 bp untuk nested-2.
Hasil amplifikasi nested-1 menunjukkan bahwa tidak ada pita yang muncul pada sampel setelah dielektroforesis. Pita hanya tampak pada marker sedangkan pada sampel dan kontrol positif tidak terdapat pendaran pita DNA target. Hal ini disebabkan komposisi mix PCR yang kurang tepat. Konsentrasi primer yang
terlalu tinggi menyebabkan kesalahan penempelan sekuens DNA sehingga hasil amplifikasi tidak sesuai dengan yang diharapkan. Sebaliknya apabila konsentrasi primer yang digunakan terlalu rendah akan menyebabkan hasil amplifikasi yang didapatkan sangat sedikit (Muladno 2002).
Komposisi pre-mix PCR yang digunakan antara nested-1 dan nested-2 mempunyai perbandingan yang sama, sedangkan DNA target antara keduanya berbeda. Nested-1 mempunyai DNA target sebesar 1640bp sedangkan nested-2 sebesar 240bp. Kurangnya komponen (pereaksi PCR, khususnya enzim Taq) untuk menyusun fragmen DNA berukuran 1640 bp menyebabkan tidak munculnya pita pada kontrol positif. Kontrol negatif menunjukkan bahwa hasil PCR tidak diperoleh pita sehingga dapat dipastikan tidak terjadi kontaminasi.
3. Amplifikasi DNA P. berghei iradiasi pada hati dan limpa tahap nested-2
Meskipun pada nested-1 tidak menunjukkan hasil amplifikasi baik untuk sampel maupun kontrol positif, tetapi belum tentu pada nested-2 tidak muncul produk gen target. Amplifikasi nested-2 tetap dilanjutkan menggunakan hasil PCR
nested-1 sebagai DNA cetakan.
Analisa data nested-2 (Tabel 4) menunjukkan bahwa sampel hati maupun limpa tidak ada yang mengandung P. berghei. Dosis 0 Gy menunjukkan bahwa pada hati dan limpa mencit tersebut tidak mengandung P. berghei. Hal ini dimungkinkan karena isolat yang diinfeksikan ke dalam tubuh mencit tidak mengandung P. berghei. Tidak semua tubuh nyamuk yang dibiakkan dilakukan pemeriksaan ada tidaknya P. berghei di dalam tubuhnya tetapi hanya sebagian nyamuk saja yang diperiksa. Selain itu, tidak diketahui secara pasti jumlah P.
berghei iradiasi yang diinfeksikan ke dalam tubuh nyamuk karena penginfeksian
dalam bentuk isolat tubuh nyamuk.
Dosis iradiasi 175 Gy diharapkan sifat infektif parasit sudah melemah sehingga tidak dapat menembus hati. Iradiasi sinar gamma pada parasit menyebabkan kerusakan materi genetik parasit. Hal ini menyebabkan daya infektifnya berkurang walaupun masih dapat melakukan proses biokimia atau
biologi lainnya, demikian juga dengan dosis 175 Gy booster. Booster bertujuan untuk meningkatkan daya imun mencit sehingga imunitasnya lebih tinggi daripada tanpa booster. Hasil PCR nested-2 dosis 175 Gy baik hati maupun limpa ternyata sesuai dengan yang diharapkan.
Sirkulasi darah merupakan tempat pertama P. berghei masuk ke dalam tubuh hospes yaitu dalam bentuk sporozoit. Sel-sel dari sistem imun innate adalah pertahanan tubuh pertama yang melawan mikroorganisme. Setelah terjadi fagositosis, antigen akan diproses dan muncul peptida major histocompability
complex( MHC) (Schmidt 2011). Sebagian sporozoit yang lolos akan menuju ke
organ hati. Sporozoit masuk ke dalam lumen sinusoidal searah atau melawan aliran darah. Sporozoit ini dapat menyerang sel Kupffer. Sel Kupffer adalah makrofag yang ada di dalam hati. Jika berhasil melewati sel Kupffer sporozoit akan melewati ruang Disse untuk menginfeksi hepatosit. Iradiasi yang diberikan membuat sporozoit lemah sehingga sulit untuk melawan sel Kupffer (Frevert et al. 2005).
Selain hati, limpa digunakan untuk identifikasi adanya infeksi Plasmodium. Limpa berfungsi melawan antigen yang berada di darah. Limpa akan membuang bahan partikel asing dan sel darah yang tua atau rusak. Sel darah yang terinfeksi akan dihancurkan oleh sistem imun di limpa. Plasmodium mempunyai sistem pertahanan untuk menghindari sistem imun limpa. Plasmodium yang lolos dari sistem imun limpa menyebabkan oklusi pembuluh darah di dalam limpa sehingga limpa akan membesar (Mohanty et al. 2005).
Meskipun nested-1 pada kontrol positif tidak muncul pita DNA tetapi setelah dilanjutkan nested-2 terdapat produk amplifikasi dengan ukuran sebesar 240 bp. Tidak terdeteksinya DNA ini dapat disebabkan oleh tidak adanya DNA parasit dalam sampel. Proses lisis sel secara mekanik kurang maksimal karena penghancuran jaringan dengan cara dijepit menggunakan pinset. Kemungkinan tersebut dapat terjadi karena sampel dalam penelitian ini berupa organ yaitu hati dan limpa yang mempunyai jumlah parasit lebih sedikit dibandingkan dengan sampel darah, sedangkan pada kontrol positif sebagai DNA pada parasit P.
berghei murni dan bukan di dalam organ. Singh et al. (1999) menyatakan bahwa
kemampuan PCR untuk mendeteksi tingkat parasitemia yang sedikit lebih dapat dipercaya daripada secara mikroskopis.
Kemampuan infeksi parasit juga dipengaruhi oleh cara infeksinya. Vaughan et al. (1999) menyatakan bahwa tingkat infeksi gigitan alami nyamuk lebih besar daripada melalui inokulasi intravena. Penelitian ini menggunakan infeksi intravena sehingga kemampuan infeksinya lebih rendah dibandingkan secara alami. Darah perifer merupakan tempat pertama masuknya P. berghei ke dalam tubuh manusia. Parasit yang masuk ke peredaran darah sebagian akan difagositosis oleh sel neutrofil. Jumlah sel parasit yang berhasil lolos dari peredaran darah lebih sedikit karena dipengaruhi oleh kemampuan infeksius parasit tersebut serta reaksi individual terhadap infeksi P. berghei tidak sama. Ada berbagai faktor ang mempengaruhi, seperti variasi genetik, metabolisme, dan sistem imun masing-masing hospes (Miller et al. 2002). Parasit yang berhasil lolos akan memasuki organ hati untuk melanjutkan siklus hidupnya.
DNA P. berghei tidak diiradiasi ditemukan di dalam limpa 5 jam setelah infeksi, tetapi dalam waktu 25 jam intensitasnya berkurang karena parasit ini telah dihancurkan oleh sel-sel makrofag limpa (Ferreira et al. 1986). Limpa merupakan tempat berkumpulnya limfosit-limfosit aktif yang masuk ke dalam darah. Limpa memberikan reaksi yang cepat terhadap antigen yang dibawa oleh APC (Antigen
Presenting Cell) dalam darah. Limpa berfungsi sebagai organ aktivasi sistem
imun adaptif oleh sebab itu limpa merupakan filter imunogenik dari sistem sirkulasi( Iskandar et al. 2006).
Pembesaran limpa merupakan petunjuk adanya infeksi Plasmodium. Selain itu infeksi menyebabkan perubahan warna hati dan limpa menjadi coklat kehitaman karena parasit mengeluarkan pigmen hemozoin. Hemozoin merupakan produk detoksifikasi parasit yang dilepaskan ke dalam peredaran darah ketika eritrosit terinfeksi sudah matang. Hemozoin digunakan oleh parasit untuk menghambat fungsi monosit dan tidak bisa berdiferensiasi menjadi sel dendritik (Schmidt 2011).Sampel yang digunakan baik hati maupun limpa dalam penelitian
ini tidak mengalami pembesaran ataupun perubahan warna menjadi gelap tetapi masih berwarna merah seperti hati dan limpa normal. Sampel hati dengan tanpa radiasi seharusnya mengalami hapatomegali dan splenomegali karena kemampuan infeksi parasit lebih tinggi dibandingkan dengan parasit yang diiradiasi (Darlina & D Tetriana, 2008).
Setiap penghancuran sel darah merah yang mengandung merozoit akan merangsang reaksi humoral dan seluler. Ini menyebabkan fagositosis
Plasmodium, sel yang diinfeksi, pigmen, dan sisa sel-sel histiosit bebas dan
makrofag dari sistem retikulo-endoteal khususnya limpa akan semakin membesar. Penimbunan pigmen oleh parasit selama pertumbuhan di eritrosit memberi warna kelabu pada organ hati dan ginjal (Brown 1979), sedangkan pada sampel tidak menunjukkan adanya perubahan warna pada hati dan limpa. Hal ini disebabkan karena adanya sistem imun yang dimediasi oleh sel T yang dipicu oleh pemberian parasit iradiasi.
Sel T adalah sel di dalam salah satu grup sel darah putih yang diketahui sebagai limfosit dan memainkan peran utama pada kekebalan selular. Sel T mampu membedakan jenis patogen dengan kemampuan berevolusi sepanjang waktu demi peningkatan kekebalan setiap kali tubuh terpapar patogen. Sel T yang telah disintesis dari kelenjar timus disebut sel T CD4+, akan terbawa oleh sirkulasi darah hingga masuk ke dalam limpa dan bermigrasi ke dalam jaringan limfatik, kemudian bermigrasi kembali ke dalam sirkulasi darah, hingga suatu saat terjadi terstimulasi oleh antigen tertentu (Janeway et al. 2001). Sel ini menyerang sel tubuh yang terinfeksi dan sel pathogen yang relatif besar secara langsung.
Penggunaan sinar gamma untuk iradiasi parasit digunakan untuk melemahkan daya infektivitas P. berghei di dalam siklus hidupnya dalam tubuh mamalia (Syaifudin et al. 2008; Tetriana 2007). Iradiasi sinar gamma akan mengurangi kemampuan Plasmodium untuk melanjutkan siklus hidupnya. Tidak terdeteksinya Plasmodium pada hati dan limpa ini dapat disebabkan sel limfosit T yang berperan dalam respon imun tubuh terhadap infeksi malaria. Setelah injeksi
parasit pasca iradiasi gamma, Plasmodium akan mengalir bersama darah dan akan difagositosis oleh neutrofil. Neutrofil dapat memfagosit dan mempunyai lisosom yang mengandung asam hidroksilase dan peroksidase untuk membunuh mikroorganisme (Eales 1999). Kemampuan infeksi yang sudah dilemahkan atau jumlah parasit yang sedikit menyebabkan parasit tidak mampu masuk ke dalam sel-sel hati. Pada tahap ini siklus hidupnya terhenti sehingga tidak bisa terdeteksi pada hati dan limpa.
31
BAB V
SIMPULAN DAN SARAN
A. Simpulan
Berdasarkan hasil penelitian dan pembahasan dapat disimpulkan bahwa
P. berghei iradiasi tidak ditemukan pada hati dan limpa mencit (Mus musculus)
menggunakan metode nested-PCR. Hal ini berarti bahwa iradiasi gamma efektif untuk menurunkan daya infektif P. berghei .
B. Saran
Penelitian tentang uji daya infektivitas P. berghei iradiasi menggunakan
nested-PCR perlu dilengkapi dengan pemeriksaan P. berghei iradiasi di dalam
tubuh nyamuk dan pengukuran jumlah sporozoit iradiasi yang diinfeksikan ke dalam tubuh mencit. Cara lisis sel perlu menggunakan cara yang lebih maksimal seperti penggerusan. Selain itu juga perlu dilengkapi dengan pengamatan mikroskopis organ, apus darah untuk memastikan bahwa di dalam peredaran darah terdapat parasit iradiasi yang diinginkan dan pengamatan serum untuk deteksi kerusakan hati seperti serum alanine aminoransferase.
32
Diseases 13(5): 681-686.
Abraham EG & M Jacobs – Lorena. 2004. Mosquito midgut barries to malaria parasite development. Insect Biochem Mol Biol 34 (7): 667-671.
Anonim.2011.Definition of Nested PCR. on line at http://www.pcrstation.com/nested-pcr/[diakses tanggal 20 April 2012].
Arum IL, AP Purwanto, S Arfi, H Tetrawindu, M Octora, Mulyanto, K Surayah & Amanukarti. 2006. Uji diagnostik Plasmodium malaria menggunakan metode imunkromatografi diperbandingkan dengan pemeriksaan mikroskopis.
Indonesian Journal of clinical Pathology and Medical Laboratory 12(3):
118-122.
Brown HW. 1979. Dasar Patologi Klinis. Penterjemah: Bintari Rukmono, Hoedojo, Nani S. Djakaria, Siti Doemilah Soeprihatin, Sri S. Margono, Sri Oemijati, Srisasi Ganda husada & Wita Pribadi. Jakarta: Gramedia.
Capurro M, de LJ Coleman, BT Beerntsen, KM Myles, KE Olson, E Rocha, AU Krettli & AA James. 2000. Virus-expressed, recombinant single-chain antibody blocks sporozoite infection of salivary glands in Plasmodium
gallinaceum-infected Aedes aegypti. Am J Trop Med Hyg. 62: 427-433.
Darlina & D Tetriana. 2008. Daya infeksi Plasmodium berghei stadium eritrositik yang diirradiasi sinar gamma . on line at: www.batan.go.id[ diakses tanggal 20 April 2012].
Djanah SN. 2007. Jumlah dan aktivitas proliferasi limfosit lien mencit swiss jantan yang diinfeksi Plasmodium berghei akibat pemberian 5 dan 100 mg/kgbb/hari ekstrak etanol Phyllantus niruri. Kes Mas 1(1): 1-50.
Eales L. 1999. Immunology for lifescientist. Newyork: John Wiley & sons.
Ferreira A, Enea V, Morimoto T & Nussenzweig V. 1986. Infectivity of Plasmodium
berghei sporozoites measured with a DNA probe. Mol Biochem Parasitol
19(2): 103-109.
Frevert U, E Sabine, Z Sergine, S Jorg, Ng Bruce, M Kai , L Leonard & H Yee. 2005. Intravital observation of Plasmodium berghei infection of the liver.
PLoS Biology 3(6): 1034-1046.
Gardner MJ, N Hall, E Fung, O White, M Berriman, RW Hyman, JM Carlton, A Pain , KE Nelson, S Bowman, IT Paulsen, K James, JA Eisen, K Rutherford, SL Salzberg, A Craig, S Kyes, MS Chan, V Nene, SJ Shallom, B Suh, J Peterson, S Angiuoli, M Pertea, J Allen, J Selengut, D Haft, MW Mather, AB Vaidya, DM Martin, AH Fairlamb, MJ Fraunholz, DS Roos, SA Ralph, GI McFadden, LM Cummings, GM Subramanian, C Mungall, JC Venter, DJ Carucci, SL Hoffman, C Newbold, RW Davis, CM Fraser & B Barrell. 2002. Genome sequence of the human malaria parasite Plasmodium falciparum. Natur
e 419(6906):498-511.Garnham PCC. 1965. The structure of early sporogenic stages
of Plasmodium berghei. Ann Soc belge Med trop 45(3): 259-266.
Ghosh A, P Srinivasan, EG Abraham, H Fujioka & M Jacobs-Lorena. 2003. Molecular strategies to study Plasmodium- mosquito interactions. Trends
Parasitol 19 (2): 94-101.
Gunes G, N Yilmaz & A Ozturk. 2011. Effects of irradiation dose and O2 and CO2 concentrations in packages on foodborne pathogenic bacteria and quality of ready-to-cook seasoned ground beef product (meatball) during refrigerated storage. The ScientificWorld Journal 20(12): 1-7.
Hermanto S, I Sugoro & Akmalia. 2008. Profil protein Escherichia coli hasil inaktivasi iradiasi gamma sebagai bahan vaksin mastitis. Depok: Prosiding Seminar Nasional Biokimia UI.
Hoffman SL, LML Goh, TC Luke, I Schneider, TP Le, DL Doolan, J Sachi, P de la Vega, M Dowler, C Paul, DM Gordon, JA Stoute, LWP Church, M Sedegah, DG Heppener, WR Ballou & TL Richie. 2002. Protection of humans against malaria by immunization with radiation-attenuated Plasmodium falciparum Sporozoites.
J Infect Diseases 185(8): 1155-1164.
Hurd H & V Carter. 2004. The role of programed cell death in Plasmodium-mosquito interactions. Int J Parasitol 34 (13-14): 1459-1472.
Iqbal J, S Ali, R H Parsotam & R Al-Owaish. 1999. Comparison of the optimal test with PCR for diagnosis of malaria in immigrants. Journal of Clinical
Microbiology 37(11): 3644-3646.
Iskandar T, DT Subekti & EF Diani. 2006. Gambaran splenosit, limpa, dan kekebalan pada mencit galur BALB/C yang diberi allantoin dan diinfeksi Toxoplasma gondii. Dalam: Seminar Nasional Teknologi Peternakan dan Veteriner, Balai Penelitian Veteriner. Bogor.
Janeway CA, Jr P Travers, M Walport & MJ Shlomchik. 2001. Immunobiology: The Immune System in Health and Disease. 5th edition, New York: Garland Science.
Kanzo K SM & L Zheng. 2003. The mosquito genome-a turning point?. Trends
Parasitol 19 (8): 329-331.
Kappe SHI, K Kaiser & K Matuschewski. 2003. The Plasmodium sporozoite journey: a rite of passage. TRENDS in Parassitology 19(3): 135-143.
Leids Universitair Medisch Centrum. The genome of P. berghei. On line at:
https://www.lumc.nl/con[ diakses tanggal 25 Januari 2013].
Michael JCR. 2005. Plasmodium sp infection in ex-captive Bornean orangutans (Pongo pygmaeus) housed at the orangutans care center and quarantine, Padang Panjang, Kalimantan Tengah, Indonesia. (Thesis). Departement of Archaeology, Simon Fraser University.
Mikolajczak SA & SH Kappe. 2006. A clash to conquer: The malaria parasite liver infection. Mol Microbiol 62(6): 1499-1506.
Miller LH, DI Baruch, K Marsh & OK Duombo. 2002. The pathogenic basis of malaria. Nature (415): 673-667.
Mohanty S, DK Patel, SS Pati & SK Mishra. 2006. Adjuvant therapy in cerebral malaria. Indian J Med Res 124(3): 245.
Muladno. 2002. Seputar Teknologi Rekayasa Genetika. Bogor: Pustaka Wirausaha Muda.
Neumaier M, A Braun & C Wagener. 1998. Fundamentals of quality assessment of molecular amplification methods in clinical diagnostics. Clinical Chemistry 44(1):12-26.
Nikjoo H. 2003. Radiation track and DNA damage. Iran J Radiat Res 1(1): 3 – 16. Nurhayati S, D Tetriana, Darlina, T Rahardjo & M Syaifudin. 2009. Pemeriksaan
Mikroskopis Plasmodium sp Sebagai Penunjang Pengembangan Vaksin Malaria Iradiasi. Jakarta. On line at http://www.batan.go.id/ptkmr [diakses tanggal 20 Januari 2012].
Palm NW & R Medzhitov. 2009. Pattern recognition receptors and control of adaptive immunity. Immunol (227): 221–233
Pooe OJ. 2011. The detection of Plasmodium falciparum in human saliva samples (Disertasi). South Africa: Zululand University.
Saiwichai T, M Maneepak, P Songprakhon, P Harnyuttanakorn & S nithiuthai. 2009. Species – specific nested PCR for detecting Plasmodium gallinaceum infresh chicken blood. Top Mad Parasitol 32:75-81.
Sanakkayala N, A Sokolovska, J Gulani, HH Esch, N Sriranganathan, SM Boyle, GG Schurig & R Vemulapalli. 2005. Induction of antigen-specific Th1-Type Immune Responses by gamma-irradiated recombinant Brucella abortus RB51.
Clinical and Diagnostic Laboratory Immunology. 12( 12): 1429-1436.
Sandoz.1973. Atlas of haematology. Switzerland: Sandoz.
Schmidt KE. 2011. Analysis of parasite-specific T cells and cellular interactions in the spleen during Plasmodium berghei induced experimental cerebral malaria. (Disertasi). Germany: University of Bonn.
Shahabuddin M. 1998. Plasmodium ookinete development in the mosquito midgut: A case of reciprocal manipulation . Parasitology 116: 83-93.
Shi Q, MM Lynch, M Romero & JM Burns. 2007. Enhanced protection against malaria by chimeric merozoite surface protein vaccine. Infection and
Immunity 75(3): 1349-1358.
Sinden RE. 1999. Plasmodium differentiation in the mosquito. Parassitologia 41 (1-3): 139-148.
. 2002. Molecular interactions between plasmodium and its insect vectors.