Evaluasi Galur Kedelai Transgenik Toleran Aluminium
pada Fasilitas Uji Terbatas
Evaluation for Tolerance of Transgenic Soybean Lines to
Aluminum in Biosafety Containtement
Saptowo J. Pardal1 dan Suharsono2
1Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian Jl. Tentara Pelajar 3A, Bogor 16111, Indonesia
E-mail: s_j_pardal@yahoo.com
2Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor Jl. Dramaga, Bogor 16680, Indonesia
Naskah diterima 24 November 2015, direvisi 22 Juni 2016, disetujui diterbitkan 27 Juni 2016
ABSTRACT
Some acid soil is potential for the agricultural development. Constraints for soybean production in the acid soils are Aluminum toxicity and macro nutrient deficiencies. Breeding for soybean varieties tolerant to acid soil is needed. This could be made through genetic engineering, by inserting acid tolerance genes into a soybean genome. Thirty one soybean lines (T0) had been obtained by insertion of Al tolerance genes (MaMt2) through an Agrobacterium mediated transformation, which nine of them contained MaMt2 gene based on PCR test. Further evaluation of those lines was carried out in the Biosafety Containment, where four T1 soybean lines were carrying MaMt2 gene, namely line GM2, GM5, GM10 and GM14. The study was aimed to evaluate the degree of tolerance of T2 generation of GM2, GM5, GM10 and GM14 lines to Al toxicity. Results showed that T2 line were able to grow in hygromicin media, indicating that those T2 lines were containing hygromicin resistant gene (hptII). Phenotypic analysis of T2 lines in four acid soil media treatments indicated that all lines could survive and grow on acid soil without liming and adding compost. GM2 line grew best on the acid medium than did other lines.
Keywords: Transgenic soybean, Al tolerance, phenotypic and molecular evaluation.
ABSTRAK
Sebagian lahan masam memiliki potensial untuk perluasan lahan pertanian. Kendala utama pengembangan kedelai di lahan masam antara lain keracunan Al dan defisiensi hara. Pengapuran kurang ekonomis dan pengaruhnya mudah hilang sehingga diperlukan varietas kedelai toleran lahan masam. Perakitan kedelai toleran lahan masam dapat dilakukan melalui rekayasa genetik dengan menyisipkan gen toleran aluminium (Al), gen MaMt2 dengan bakteri
Agrobacterium tumefaciens ke dalam genom kedelai. Sebanyak 31 tanaman kedelai T0 telah dihasilkan melalui insersi gen toleran Al menggunakan perantaraan Agrobacterium tumefaciens dan sembilan galur diantaranya positif mengandung gen MaMt2 berdasarkan hasil uji PCR. Selanjutnya, pada pengujian lebih lanjut
galur tersebut di fasilitas uji terbatas diperoleh empat galur T1 yang masih membawa gen MaMt2, yaitu GM2, GM5, GM10 dan GM14. Keempat galur ini digunakan dalam penelitian ini. Penelitian ini bertujuan untuk mengetahui toleransi terhadap Al dari empat galur kedelai T2 (GM2, GM5, GM10 dan GM14) yang berasal dari galur T1 secara fenotipik dan genetik (molekuler). Tahapan pengujian meliputi uji kepekaan benih T1 terhadap higromisin, uji toleransi tanaman T2 terhadap cekaman Al pada fasilitas uji terbatas, dan konfirmasi gen
MaMt2 pada tanaman T2. Hasil penelitian menunjukkan keempat galur T2 yang diuji berkecambah pada medium higromisin, artinya semua galur tersebut membawa gen ketahanan higromisin (hptII). Kemudian hasil uji fenotipik tanaman T2 menggunakan empat media tumbuh dengan pH dan kandungan Al berbeda menunjukkan keempat galur kedelai T2 hanya dapat tumbuh pada media tanah dengan perlakuan Al (tanpa kapur dan kompos). Pertumbuhan galur GM2 lebih baik daripada GM5, GM10 dan GM14.
Kata kunci: Kedelai transgenik, toleransi Al, karakter fenotipik, molekuler.
PENDAHULUAN
Di Indonesia, kedelai merupakan pangan utama setelah padi. Kebutuhan kedelai terus meningkat setiap tahun, sedangkan produksi nasional masih rendah, yang tidak mencukupi semua kebutuhan (Tasma et al. 2015). Salah satu kendala yang dihadapi adalah rendahnya produktivitas kedelai dan semakin berkurangnya areal tanam kedelai akibat alih fungsi lahan, terutama di pulau Jawa (Anggraito 2012).
Laju alih fungsi lahan pertanian yang tinggi di Pulau Jawa, sebagai daerah penghasil utama produk pertanian, termasuk kedelai, menjadi salah satu ancaman bagi keberlanjutan penyediaan bahan pangan. Pada sisi lain, Indonesia masih memiliki lahan marjinal (suboptimal) yang potensial dijadikan lahan pertanian (Prasetyo dan Suriadikarta 2006). Namun tanah ini
umumnya masam, kejenuhan basa rendah, kandungan Al berpotensi meracuni tanaman, kandungan bahan organik rendah, kahat hara tertentu seperti P, Ca, Mg, N dan K (Sopandie 2006, Prasetyo dan Suriadikarta 2006, Mulyani et al. 2009).
Aluminium (Al) merupakan unsur ketiga terbanyak yang terdapat di bumi setelah oksigen dan silikon. Pada taraf tertentu, Al yang ada di tanah menjadi faktor pembatas pertumbuhan tanaman pada lahan masam (Pilon-Smits et al. 2009). Pemberian kapur pada tanaman kedelai kurang ekonomis karena unsur ini cepat hilang pengaruhnya. Oleh karena itu diperlukan varietas kedelai toleran lahan masam dengan kelarutan logam tinggi untuk dikembangkan pada lahan-lahan masam, terutama di luar Jawa (Anggraito 2012).
Perakitan varietas kedelai toleran Al dapat melalui proses penapisan, seleksi, dan persilangan. Namun hingga saat ini belum ada varietas kedelai toleran Al atau yang dapat beradaptasi dan berproduksi tinggi pada lahan masam. Alternatif yang dapat digunakan untuk merakit varietas kedelai toleran lahan masam adalah melalui bioteknologi atau rekayasa genetik, dengan cara menyisipkan gen toleran lahan masam ke tanaman kedelai budi daya (Suharsono dan Yusuf 2006).
Harendong (Melastoma malabatrichum)
merupakan tanaman yang mampu tumbuh pada lahan dengan pH rendah dan kelarutan Al tinggi, sehingga potensial digunakan sebagai sumber gen toleransi terhadap cekaman pH rendah dan Al tinggi (Muhaemin 2008). Tanaman ini memiliki kemampuan tinggi untuk menyerap dan mengakumulasi Al pada daun, terutama pada sel-sel epidermis atas dan mesofil, dan pada seluruh jaringan akar, sehingga tanaman ini disebut akumulator Al (Watanabe et al. 2008). Mutiasari (2008) melaporkan tanaman M. affine D. Don. atau M. malabathricum yang ditumbuhkan pada media cair dengan pH 4,0 dan 3,2 mM Al dapat mengakumulasi 8,81 g Al/kg yang terdapat dalam daun tua selama dua bulan.
Salah satu gen toleransi terhadap pH rendah dan Al tinggi adalah MaMt2. Gen penghasil senyawa metalothionein ini diisolasi dari M. malabatrichum. Gen berasal dari transkripsi balik mRNA M. malabatrichum
yang telah diisolasi, dikarakterisasi, dan diklon ke dalam plasmid pGEMT-easy (Suharsono et al. 2009a), kemudian disisipkan ke dalam vektor ekspresi pIG6 dalam bakteri A. tumefaciens. Konstruksi ekspresi berlebih untuk gen MaMt2 dilakukan dengan menggabungkan pada promoter kuat Ubiquitin jagung dan ditambahkan terminator NOS di bagian hilir, sehingga pola ekspresi gen akan berubah dari gen
inducible menjadi konstitutif (ekspresi terus menerus/ berlebih). Konstruksi ini menguntungkan tanaman,
karena dapat memproduksi protein target yang tidak bergantung pada senyawa-senyawa penginduksi atau kondisi lingkungan yang ada (Hannum 2012).
Pada penelitian terdahulu (Anggraito 2012) telah dilakukan transformasi gen toleransi terhadap Al (gen
MaMt2) dari tanaman harendong ke tanaman kedelai varietas Lumut dan menghasilkan 31 galur kedelai hasil regenerasi (T0). Sembilan di antara galur tersebut mengandung gen MaMt2 dan setelah ditanam di rumah kaca fasilitas uji terbatas (FUT) diperoleh empat galur (event) T1 yang positif mengandung gen MaMt2, yaitu GM2, GM5, GM10, dan GM14 (Anggraito 2012). Selanjutnya, galur T1 tersebut diselfing dan menghasilkan benih T2 (Pardal et al. 2014) yang perlu dievaluasi lebih lanjut untuk mengetahui pengaruh integrasi gen MaMt2 terhadap morfologi tanaman dan toleransinya terhadap cekaman Al pada tanah masam (pH rendah).
Penelitian ini bertujuan untuk mengetahui toleransi empat galur kedelai T2 (GM2, GM5, GM10 dan GM14) yang berasal dari galur T1 secara fenotipik dan genetik (molekuler) terhadap Al.
BAHAN DAN METODE
Penelitian dilakukan di rumah kaca pada Fasilitas Uji Terbatas (FUT), dan Laboratorium Biologi Molekuler, Balai Besar Penelitian Pengembangan Bioteknologi dan Sumber Daya Genetik Pertanian (BB Biogen), Bogor, pada tahun 2015. Bahan yang digunakan adalah benih empat galur kedelai transgenik T2 yang merupakan hasil penyerbukan sendiri (selfing) tanaman kedelai transgenik T1 yang positif membawa gen MaMt2, yaitu GM2, GM5, GM10, dan GM14. Varietas Lumut, Sindoro, dan Tanggamus digunakan sebagai kontrol. Lumut merupakan varietas kedelai yang peka terhadap Al (kontrol negatif) yang merupakan tetua dari keempat galur kedelai transgenik tersebut sedangkan Sindoro dan Tanggamus merupakan varietas toleran terhadap Al (kontrol positif). Penelitian terdiri atas tiga tahapan, yaitu: (1) uji higromisin, (2) uji toleransi galur kedelai T2 terhadap Al, dan (3) deteksi gen MaMt2.
1. Uji Higromisin
Pengujian dilakukan pada ruang gelap di Laboratorium Biologi Molekuler, BB Biogen, Bogor, pada tahun 2015, untuk mengetahui galur kedelai T2 yang diuji bersifat homosigot. Bahan yang diuji adalah benih empat galur kedelai transgenik T1 (GM2, GM5, GM10 dan GM14), masing-masing 20 biji. Bahan lain yang digunakan adalah antibiotik higromisin, cawan petri diameter 10 cm, ember plastik hitam ukuran 5 kg (diameter 30 cm), kertas saring diameter 10 cm, dan akuades.
Pengujian mengikuti metode Pardal et al. (2014) menggunakan rancangan acak lengkap dengan dua ulangan. Konstruksi gen MaMt2 yang disisipkan ke dalam kedelai membawa gen hptII, yaitu gen tahan antibiotik higromisin (Suharsono et al. 2009b). Biji kedelai T2 dari masing-masing galur diambil 20 biji, kemudian diuji daya kecambahnya pada dua petridish yang berisi larutan higromisin 25 mg/l (10 biji/petridish). Sebagai kontrol, benih kedelai T2 juga dikecambahkan pada petridish
yang berisi akuades (tanpa higromisin). Petridish yang berisi benih kedelai selanjutnya diletakkan di dalam bak plastik dan ditutup dengan aluminium foil atau kain hitam, lalu disimpan di ruang gelap.
Parameter yang diamati adalah jumlah benih kedelai yang berkecambah. Pengamatan dilakukan dua kali, yaitu pada dua dan empat hari setelah inkubasi (perkecambahan). Kriteria pengamatan dan ketahanan terhadap higromisin adalah jika semua biji dapat berkecambah dengan baik (normal), sehingga kelompok biji tersebut dinyatakan homosigot. Sebaliknya, jika terdapat satu saja biji yang tidak berkecambah, maka benih tersebut bersifat heterosigot. Kelompok benih kedelai T2 yang homosigot selanjutnya digunakan untuk uji toleransi terhadap Al dan pH rendah menggunakan tanah masam di rumah kaca.
2. Uji Toleransi terhadap Al
Pengujian dilakukan di rumah kaca pada Fasilitas Uji Terbatas (FUT) BB Biogen, Bogor, pada tahun 2015, untuk mengetahui toleransi galur kedelai transgenik terhadap Al. Bahan yang diuji adalah empat kecambah dari empat galur kedelai GM2, GM5, GM10 dan GM14 yang dapat berkecambah pada media Higromisin. Uji toleransi dilakukan dengan cara menanam kecambah dari keempat galur transgenik pada empat macam media tumbuh perlakuan Al, yaitu media A, B, C dan D menggunakan rancangan acak kelompok dengan empat ulangan. Media tanah ditempatkan dalam polibag (4 kg tanah/polibag).
Komposisi media tanah adalah sebagai berikut: (1) media A berisi 4 kg tanah masam dari Jasinga, Bogor,
ditambah 3 g pupuk NPK; (2) media B berisi 4 kg tanah masam Jasinga, ditambah 3 g NPK dan 25 g kompos; (3) media C berisi 4 kg tanah masam Jasinga, ditambah 3 g NPK, 25 g kompos, dan 6 gram kapur, dan (4) Media D berisi 4 kg tanah biasa ditambah 3 gram NPK dan 25 gram kompos. Sebelum digunakan, keempat media tersebut dianalisis kandungan tanahnya di Balai Penelitian Tanah (Balittanah) Bogor. Hasil analisis disajikan pada Tabel 1.
Parameter yang diamati adalah tinggi tanaman, jumlah daun kuning, jumlah bunga, jumlah polong, dan jumlah biji. Tinggi tanaman diamati setiap minggu selama satu bulan (4 minggu) sejak tanam untuk mengetahui laju pertumbuhan tanaman. Sebagai kontrol varietas peka Al digunakan kedelai nontransgenik varietas Lumut, sedangkan sebagai kontrol toleran digunakan varietas Sindoro. Kecambah kedelai T2 yang dapat tumbuh pada tanah masam (mengandung Al dan pH rendah) dirawat hingga dewasa dan dibiarkan menyerbuk sendiri (selfing) untuk menghasilkan benih kedelai T3.
3. Deteksi Gen MaMt2
Analisis molekuler dilakukan di laboratorium Biologi Molekuler, BB Biogen, Bogor, pada tahun 2015, untuk mengetahui keberadaan gen MaMt2 pada tanaman kedelai T2 toleran Al. Bahan yang digunakan adalah tanaman dari empat galur kedelai transgenik T2 (GM2, GM5, GM10, GM14) yag toleran Al, yaitu tanaman kedelai yang dapat tumbuh pada media A. Bahan lain yang digunakan adalah perangkat untuk ekstraksi DNA (Invitrogen, USA) dan uji PCR, sepasang primer gen
MaMt2, marka DNA (1 Kb leader), agarosa, dan akuades steril.
Analisis molekuler menggunakan metode Pardal et al. (2004a). Tahapan analisis meliputi isolasi/ekstraksi DNA, running PCR, gel elektroforesis, visualisasi hasil elektroforesis, dan analisis data pita DNA. Pada tahap ekstraksi DNA, daun kedelai disampling pada saat tanaman berumur 2 minggu setelah tanam. DNA kedelai diekstraksi dari sampel daun menggunakan miniprep extraction kit (Invitrogen-USA). DNA hasil ekstraksi
Tabel 1. Karakteristik tanah yang digunakan sebagai media tumbuh untuk uji tolertansi tanaman kedelai T2 terhadap Al.
pH Bahan organik KCl 1N Media tanah Asal tanah
H2O KCl C N C/N Al3+ H+
A Jasinga 4,8 3,8 0,28 0,03 9 5,02 0,68
B Jasinga 5,0 4,4 3,18 0,37 9 0,62 0,33
C Jasinga 8,1 7,6 2,64 0,25 11 0,00 0,11
D Cimaggu 7,0 6,2 1,89 0,24 8 0,00 0,02
dianalisis untuk mengetahui keberadaan gen MaMt2
dengan teknik Polymerase Chain Reaction (PCR) menggunakan sepasang primer spesifik gen MaMt2. Prosedur PCR mengikuti protokol dari Amplification Kit PCR (Invitrogen-USA). Hasil PCR diseparasi pada gel elektroforesis, kemudian divisualisasikan dengan alat
GelDoc. Sampel tanaman kedelai disebut positif mengandung gen MaMt2 jika diperoleh pita DNA berukuran 160 pb, setara dengan gen MaMt2.
HASIL DAN PEMBAHASAN
1. Uji Higromisin
Hasil uji kepekaan terhadap highromisin berdasarkan pengamatan pada hari keempat setelah tanam menunjukkan seluruh benih dari keempat galur kedelai transgenik (GM2, GM5, GM10, GM14) berkecambah dengan baik pada media higromisin maupun akuades (kontrol), walaupun terdapat perbedaan pada panjang akar dan warna kecambah (Tabel 2). Akar kecambah pada media higromisin rata-rata lebih pendek dibandingkan dengan kecambah pada media air (1:3) dan warna kecambah lebih pucat (Gambar 1). Hal ini disebabkan karena kecambah kedelai transgenik mengandung gen ketahanan antibiotik higromisin, hptII, sehingga dapat berkecambah pada media higromisin 25 mg/l. Akar kecambah yang lebih pendek dan warna agak pucat dibandingkan dengan kecambah kontrol menunjukkan adanya seleksi dari higromisin. Sebaliknya, benih kedelai kontrol toleran dan peka Al tidak dapat berkecambah pada media higromisin, tetapi dapat berkecambah 100% pada media air (Tabel 2). Hal ini disebabkan karena benih kedelai nontransgenik tidak memiliki gen ketahanan terhadap higromisin.
2. Uji Toleransi Al
Secara umum semua galur kedelai transgenik dapat tumbuh pada empat media perlakuan Al, namun kurang baik akibat kemasaman tanah, terutama pada media A. Dari 10 kecambah kedelai T2 hasil uji higromisin yang ditanam pada media perlakuan, tidak semuanya tumbuh menjadi tanaman. Hal ini disebabkan oleh adanya pengaruh senyawa antibiotik higromisisn pada kecambah dan adanya pengaruh tanah masam, sehingga jumlah sampel atau ulangan tanaman untuk setiap galur pada pengujian Al hanya empat. Pertumbuhan tanaman kedelai pada media A kurang bagus dibandingkan dengan media lainnya, yang
Gambar 1. Pertumbuhan benih kedelai GM2 yang dikecambahkan pada cawan petri berisi media larutan higromisin 25 mg/l (H) dan media akuades (A). FUT BB Biogen, Bogor. 2015.
I = kecambah kedelai GM2 umur 2 hari setelah dikecambahkan pada H, akar masih pendek,
II = kecambah kedelai GM2 umur 4 hari setelah perkecambahan akar memanjang, namun lebih pendek dibanding media kontrol (air). Tabel 2. Perkecambahan benih galur T2 dan varietas kedelai pada larutan higromisin dan air (4 HSP). FUT BB Biogen, Bogor. 2015.
Jumlah benih berkecambah (%) pada media: Galur T2/
varietas kontrol Higromisin Akuades (25 mg/l) (tanpa higromisin) Rata-rata % Rata-rata % GM 2 10 100 10 100 GM 5 10 100 10 100 GM 10 10 100 10 100 GM 14 10 100 10 100 Lumut 1 10 10 100 Sindoro 0 0 10 100 Tanggamus 1 10 10 100
Benih galur kedelai transgenik GM 2, GM 5, GM 10, dan GM 14 transgenik berkecambah 100% pada media larutan higromisin 25 mg/l, namun panjang akar lebih pendek (1:3) dan warna kecambah lebih pucat dibandingkan akar kecambah pada media akuades (kontrol).
II
A
HA
I
H
ditandai oleh jumlah daun lebih sedikit dan berwarna kekuningan.
Hasil analisis terhadap empat jenis tanah perlakuan Al (media A,B, C, dan D) sebelum ditanami menunjukkan media A memiliki pH 4,8 dengan kandungan Al3+ terlarut
5,02 cmol/kg, C 0,28%, dan N 0,03%. Media B memiliki pH 5,0 dengan kandungan Al3+ terlarut 0,62 cmol/kg, C 3,18%
dan N 0,37%. Media C memiliki pH 8,1, Al3+ terlarut 0,0
cmol/kg, C 2,64%, dan N 0,25%. Media D (kontrol) memiliki pH 7,0, Al3+ terlarut 0,0 cmol/kg, C 1,89% dan N 0,24%.
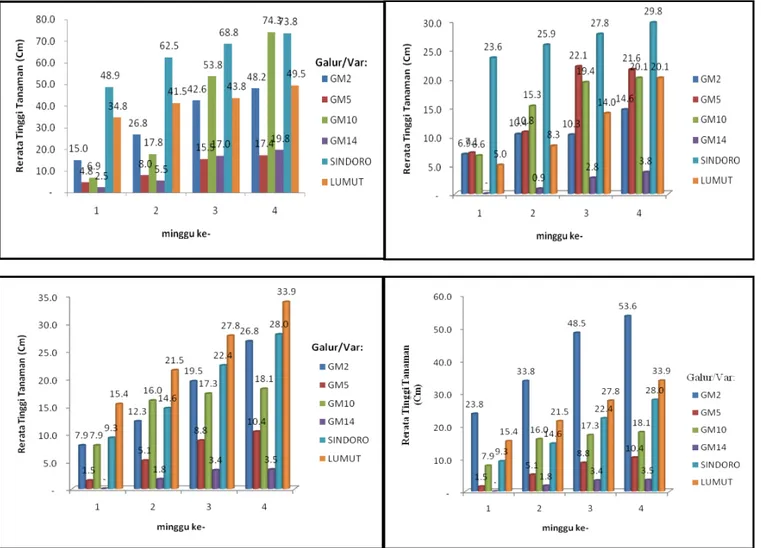
Hasil pengamatan pada media A yang berisi tanah masam Jasinga dengan pH 4,8 dan kandungan Al3+
terlarut 5,02 cmol/kg, menunjukkan tanaman dari keempat galur kedelai transgenik T2 dapat tumbuh. Pada empat minggu setelah tanam, galur GM 10, GM 2, dan varietas Sindoro menunjukkan pertumbuhan yang lebih baik. Pertumbuhan terbaik ditunjukkan oleh galur GM 10 dengan rata-rata tinggi tanaman 74,3 cm, kemudian Sindoro 73,8 cm, dan GM 2 48,2 cm. Gen MaMt2 pada galur kedelai T2 menyebabkan tanaman dapat mengeksresikan senyawa metalothionein (MT) dan mengakumulasi ion Al yang terlarut di tanah, sehingga tanaman dapat tumbuh baik meskipun lahan ber-pH rendah dan kelarutan Al tinggi. Hal ini sesuai dengan pendapat Muhaemin (2009), dimana tanaman yang memiliki gen MaMt2 memiliki kemampuan yang tinggi dalam menyerap dan mengakumulasi Al pada daun, terutama pada sel-sel epidermis atas dan mesofil, serta pada seluruh jaringan akar.
Ekspresi gen MaMt2 pada tanaman diregulasi oleh berbagai faktor, di antaranya oleh cekaman logam berat (Usha et al. 2007, Huang and Wang 2009, 2010). Hal ini menunjukkan senyawa metallothionein diekspresikan pada akar sebagai tanggap toleransi terhadap cekaman Al. Hal serupa juga dilaporkan oleh Trisnaningrum (2009), bahwa cekaman Al terhadap Melastoma affine
pada konsentrasi 3,2 mM meningkatkan ekspresi gen
MaMt2 pada akar, tetapi tidak pada daun. Varietas Sindoro yang merupakan varietas toleran Al juga dapat tumbuh baik pada media tanah masam. Fungsi metallothionein produk protein dari gen MaMt2 dihubungkan dengan kemampuannya untuk mendetoksifikasi logam berat atau fungsi homeostasis logam bivalen. Hal ini terkait dengan kandungan asam amino sistein yang tinggi. Gugus sulfhidril (S-H) dari residu sistein akan mengkelat logam, sehingga tidak reaktif bagi sel atau jaringan (Ryan et al. 2009). Meski sampai saat ini belum ada laporan pengkelatan logam Al oleh metallothionein, tetapi melihat kemampuan gugus sulfhidril mengikat logam-logam bivalen (Cu, Zn, Mn, Cd, dsb.) maka kemungkinan protein metallothionein juga mampu mengkelat Al, meskipun dalam jumlah molekul yang lebih rendah. Untuk itu perlu
dilanjutkan uji tantang dengan berbagai konsentrasi Al untuk mengetahui kerja protein MT dalam mengkelat logam Al.
Pada media B yang berisi tanah masam ditambah dengan kompos, galur-galur kedelai transgenik tidak dapat tumbuh dengan baik. Hal ini dapat dilihat dari tinggi tanaman. Kompos yang ditambahkan ternyata belum mampu menaikkan pH tanah (hasil analisis tanah B menunjukkan pH 5 dan kandungan Al3+ terlarut0,62
cmol/kg), sehingga tanah masih bersifat masam dan mengandung Al. Kondisi ini tidak sesuai untuk pertumbuhan tanaman kedelai. Selain itu, pada media ini tanaman kedelai terinfeksi penyakit, sehingga pertumbuhannya tidak optimal. Galur yang tumbuh lebih baik adalah Sindoro dan GM 5. Tanaman tertinggi dilanjutkan oleh galur/varietas Sindoro (29,8 cm), GM 5 (21,6 cm), dan GM 10 (20,1 cm) (Gambar 2).
Pada media C, galur-galur kedelai transgenik juga tidak dapat tumbuh dengan baik, kecuali galur GM 2, varietas Lumut, dan Sindoro (Gambar 2). Pada media ini, tanah masam ditambah dengan kompos dan kapur. Kapur yang ditambahkan meningkatkan pH tanah (hasil analisis tanah C menunjukkan pH 8,1 dan kandungan Al3+ terlarut 0,0
cmol/kg), namun kapur mudah mengalami leaching
(pencucian) oleh air pada saat penyiraman sehingga hilang pengaruhnya (Anggraito 2012).
Pada media D yang berisi tanah biasa (nonmasam) ditambah kompos dan NPK memiliki pH 7 (normal) dan kandungan Al3+ terlarut 0,0 cmol/kg, sehingga semua
galur tumbuh dengan baik. Hal ini ditunjukkan oleh tinggi tanaman selama 4 minggu pengamatan. Galur yang tumbuh lebih baik adalah GM 2 dan Lumut. Galur GM 2 memiliki pertumbuhan terbaik dengan rata-rata tanaman tertinggi (53,6 cm) pada minggu keempat setelah tanam (Gambar 2).
Kedelai transgenik galur GM5 menunjukkan pertumbuhan yang kurang baik pada media C dan D. Hal ini disebabkan oleh kondisi tanaman yang terinfeksi penyakit (virus). Namun galur ini masih lebih baik dibandingkan dengan galur GM14. Galur kedelai transgenik GM14 menunjukkan pertumbuhan yang paling merana (tinggi tanaman terendah) pada media C dan D. Seluruh galur GM14 terinfeksi penyakit (virus) paling parah, sehingga tanaman tidak dapat tumbuh normal (Gambar 2).
3. Deteksi Gen MaMt2
Pada uji toleransi kedelai transgenik T2 galur GM 2, GM 5, GM 10, dan GM 14 terhadap Al diperoleh beberapa tanaman yang dapat bertahan hidup (tumbuh) pada media yang mengandung Al, khususnya media A yang berisi tanah masam Jasinga (pH 4,8) dan 3 g NPK.
Analisis PCR terhadap 13 sampel DNA kedelai T2 galur GM2, GM5, GM10 dan GM14 menghasilkan pita DNA berukuran 160 pb, sama dengan ukuran DNA pada sampel plasmid yang mengandung gen MaMt2 (kontrol). Hal ini berarti tanaman tersebut positif mengandung gen
MaMt2 (Gambar 3).
Tanaman kedelai yang menunjukkan toleransi terhadap Al dan mengandung gen MaMt2, khususnya yang dapat tumbuh pada media perlakuan A, dipelihara terus hingga dewasa untuk menghasilkan benih kedelai T3 MaMt2. Tanaman kedelai dari galur GM 2, GM 5, dan GM 10 dapat bertahan hidup hingga dewasa dan menghasilkan polong serta biji T3 masing-masing sebanyak 50 biji (GM 2), delapan biji (GM 5), dan 30 biji (GM 10). Tanaman dari galur GM 14 tidak dapat bertahan hidup dan tidak menghasilkan biji T3. Biji kedelai T3 akan digunakan sebagai materi pengujian lebih lanjut.
Gambar 2. Histogram pertumbuhan kedelai transgenik dan kontrol pada empat macam media tumbuh (A – D) hingga empat minggu setelah tanam. FUT BB Biogen, Bogor, 2015.
Gambar 3. Foto gel elektroforesis sampel DNA hasil PCR tanaman DNA galur kedelai T2. M = 1 kb DNA ladder (Marka DNA); 1 = Kontrol positif (plasmid); 2 = kontrol negatif (air); 3 = GM2 (2); 4 = GM2 (3); 5 = GM2 (4); 6 = GM5 (2); 7 = GM5(3); 8 = GM5(4); 9 = GM10 (1); 10 = GM10 (2); 11 = GM10 (3); 12 = GM10 (4); 13 = GM14 (1); 14. GM14 (2), and 15 = GM14 (4).
KESIMPULAN
Benih galur kedelai transgenik T2 (GM2, GM5, GM10 dan GM14) dapat berkecambah dan tumbuh pada media yang mengandung higromisin 25 mg/l. Artinya, keempat galur membawa gen ketahanan terhadap higromisin,
hptII.
Semua galur T2 yang diuji toleran terhadap media tumbuh yang mengandung Al3+ terlarut 5,02 cmol/kg
dengan pH 4,8. Semua galur kedelai yang toleran terhadap Al mengandung gen MaMt2.
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Badan Penelitian dan Pengembangan Pertanian yang telah membiayai penelitian ini melalui program KKP3N. Ucapan terima kasih juga disampaikan kepada Dr. Utut Widyastuti dari IPB dan Dr. Yustinus Ulung Anggraito dari Universitas Negeri Semarang yang telah memberikan materi penelitian. Penulis juga mengucapkan terima kasih kepada Pratiwi, Endang Ibrahim, dan Riri Sundasari yang telah membantu pelaksanaan penelitian ini.
DAFTAR PUSTAKA
Anggraito, Y.U. 2012. Transformasi genetik Nicotiana benthamiana
L., dan kedele dengan gen MaMt2 penyandi metallothionein tipe II dari Melastoma malabathricum L. Disertasi S3, Sekolah Pascasarjana IPB, Bogor.
Hannum, S. 2012. Isolasi dan analisis ekspresi gen toleran aluminium pada Melastoma malabathricum L. Disertasi S3. Sekolah Pascasarjana IPB, Bogor.
Huang, G.Y. and Y.S. Wang, 2009. Expression analysis of type 2
metallothionein gene in mangrove species (Bruguiera
gymnorhiza) under heavy metal stress. Chemosphere 77: 1026-1029.
Huang, G.Y. and Y.S. Wang, 2010. Expression and characterization analysis of type 2 metallothionein from grey mangrove species (Avicenia marina) in response to metal stress. Aquat. Toxicology 99: 86-92.
Muhaemin, 2008. Analisis pertumbuhan Melastoma (Melastoma
malabathricum auct. Non L dan M. affine D. Don) yang mendapat cekaman pH rendah dan aluminium. Tesis. Program Pascasarjana, IPB.
Mulyani, A., A. Rachman, dan A. Dairah, 2009. Penyebaran lahan masam, potensi dan ketersediaannya untuk pengembangan pertanian. Buku Fosfat Alam. Balittanah, Badan Litbang Pertanian. www.balittanah.litbang.pertanian.go.id
Mutiasari, A. 2008. Akumulasi aluminium pada Melastoma affine
dan M. malabathricum. Tesis. Program Pascasarjana, IPB.
Pardal, S.J., Suharsono, Y.U. Anggraito, dan U. Widyastuti. 2014. Perakitan kedelai transgenik toleran terhadap lahan masam. Laporan Akhir KKP3N Balitbangtan. 20 halaman.
Pilon-Smits, E.A.H, C.F. Quinn, W. Tapken, M. Malagoli, and M. Schiavon, 2009. Physiological functions of beneficial elements. Curr. Opinion in Plant Biology 12:267-274. Prasetyo, B.H., D.A. Suriadikarta, 2006. Karakteristik,potensi dan
teknologi pengelolaan tanah ultisol untuk pengembangan pertanian lahan kering di Indonesia. J. Litbang Pertanian 25(2): 39-46.
Ryan, P.R., H. Raman, S. Gupta, W.J. Horst, and E. Delhaize. 2009. A second mechanism for aluminum resistance in wheat relies on the constitutive efflux of citrate from roots. Plant Physiol. 149:340-351.
Sopandie, D. 2006. Perspektif fisiologi dalam pengembangan tanaman pangan di lahan marjinal. Orasi ilmiah Guru Besar. Fakultas Pertanian Institut Pertanian Bogor. 121p.
Suharsono dan M. Yusuf. 2006. Isolasi dan karakterisasi gen-gen yang berhubungan dengan toleransi tanaman terhadap pH rendah dan aluminium tinggi. Laporan Penelitian HPTP. LPPM-Institut Pertanian Bogor.
Suharsono. 2007. Pembuatan pustaka genom kedelai (Glycine max
(L.) Merrill) kultivar lumut di dalam fage lambda. Biosfera 24(2):83-90.
Suharsono, S. Firdaus, dan U. Widastuti. 2008. Isolasi dan pengklonan fragmen cDNA dari gen penyandi multidrug resistance associated protein dari Melastoma affine. Makara Sains 12(2):102-107.
Suharsono, N. Trisnaningrum, L.D. Sulistyaningsih, and U. Widyastuti. 2009a.Isolation and cloning of fragment of cDNA of gene encoding for multidrug resistance associated protein from Melastoma affine. Biotropia 9(1):27-36.
Suharsono, Y. Anwar, and U. Widyastuti. 2009b. Isolation and cloning of cDNA of gene encoding for metallothionein type 2 from soybean [Glycine max (L.) (Merrill)] cv Slamet. Biodiversitas 10 (3): 109-114.
Tistama, R, U. Widyastuti, D. Sopandie, A. Yokota, K. Akashi, and Suharsono. 2012. Physiological and biochemical responses to aluminum stress in the root of a biodiesel plant Jatropha curcas L. Hayati. J. Bioscience 19(1):37-43.
Trisnaningrum, N. 2009. Analisis ekspresi gen penyandi
metallothionein tipe II pada Melastoma affine L. yang mendapat cekaman pH rendah dan aluminium. Tesis. Program Pascasarjana, IPB.
Trisnaningrum, N., M. Yunus, and Suharsono. 2009. Expression of gene encoding for metallothionein from Melastoma affine. Postgraduate GP Education Workshop, Ibaraki University, Japan, January 12-13.
Usha, B., S.R. Prashanth, and A. Parida, 2007. Differential expression of two metallothionein encoding gene during heavy metal stress in the mangrove species, Avicenia marina
(Forsk.) Vierh. Curr. Science 93(9):1215-1219.
Watanabe, T., S. Misawa, S. Hiradate, and M. Osaki, 2008. Root
mucilage enhances aluminum accumulation in Melastoma
malabathricum, an aluminum accumulator. Plant Signal & Behav 3(8):603-605.