PEMANFAATAN CENDAWAN UNTUK MENINGKATKAN
PRODUKTIVITAS DAN KESEHATAN TERNAK
Riza Zainuddin Ahmad
Balai Besar Penelitian Peteriner, Jalan R.E. Martadinata No. 30, Bogor 16114
ABSTRAK
Cendawan merupakan mikroorganisme eukariotik, memproduksi spora, tidak berklorofil, mcmperolch nutrisi dengan cara absorbsi, bereproduksi secara seksual dan aseksual, mempunyai struktur somatik dalam bentuk hifa, dan berdinding sel yang terdiri alas kitin dan selulosa. Cendawan-cendawan tertentu dapat dimanfaatkan untuk ternak, seperti kapang Beauveria bassiana, Duddingtoniaflagrans, dan Metarhizium anisopliae yang dapat digunakan untuk meningkatkan keschatan ternak dan pengendali hayati. Khamir Saccharomyces cerevisiae dapat berperan sebagai probiotik dan imunostimulan untuk meningkatkan produktivitas dan keschatan ternak. Mikroba isolat lokal Indonesia ini merupakan hasil penelitian dalam kurun waktu 20 tahun, dimulai pada tahun 1980-an sampai dengan 2000-an. Sebelum terpilih sebagai cendawan yang bermanfaat bagi ternak, cendawan melalui tahapan scleksi yang panjang. Dari isolat-isolat yang berhasil diisolasi dan diidentifikasi, hanya sedikit yang terpilih sebagai pengendali hayati, probiotik atau imunostimulan.
Kata kunci: Cendawan, pengendali hayati, probiotik, imunostimulan, ternak
ABSTRACT
The use of fungi for increasing productivity and healthy of livestock
Fungi are eukaryotic microorganisms, produce spores, have no chlorophyll, obtain nutrition by absorbing, reproduce sexually and asexually, have somatic structure in hifa shape, and have cell wall made of chitin and cellulose. Certain fungi can be used for livestock. These mould and yeast are Beauveria bassiana, Duddingtonia flagrans, and
Metarhizium anisopliae that can be used for increasing healthy as biological control. Saccharomyces cerevisiae
yeasts are used for increasing productivity and healthy as probiotic and immunostimulant. These original isolates from Indonesia were the results for the last 20 years beginning in 1980s to 2000s. Before being chosen as fungi used for the livestock, the fungi went through some stages of a long selection. However, from the isolates successfully isolated and identified only a few were chosen as biological control, probiotic or immunostimulant.
Keywords: Fungi, biological control, probiotic, immunostimulant, livestock
Cendawan dapat digolongkan menjadi
jamur, kapang, dan khamir (Alexopou-Ins et al. 1996; Dube 1996). Cendawan dapat dimanfaatkan dalam budi daya ternak antara lain sebagai pengendali hayati, probiotik, dan imunostimulan. Pemanfaatan beberapa cendawan golongan kapang dan khamir seperti
Duddingtonia flagrans, Beauveria bassiana, Metarhizium anisopliae, dan Saccharomyces cerevisiae sebagai agen
hayati pengendalian penyakit dapat meningkatkan produktivitas ternak.
Selama pemeliharaan, temak dapat terserang berbagai penyakit sehingga tidak mampu mencapai produktivitas yang optimal, bahkan dapat menyebabkan kematian. Penyakit yang menyerang ternak dapat disebabkan oleh virus,
bakteri atau parasit (cacing, cendawan, protozoa, serangga, dan akarid).
Gejala klinis yang muncul pada temak yang terserang cacing adalah kurus, konversi pakan buruk, mencret, anemia, edema, dan bila infestasi cacing terlalu ba-nyak dapat menyebabkan kematian. Pada domba, bila infestasi cacing Haemonchus
contortus mencapai 5.000 larva maka
ternak dapat menderita haemonchosis. Pada kasus akut, infestasi cacing mencapai
2.000-20.000 larva, dan pada kasus
hiperakut, infestasi cacing sekitar 30.000 larva (Soulsby 1986; Urqurhart et al. 1987). Pada ternak yang terserang caplak, gejala klinis yang muncul adalah temak menjadi kurus, caplak pada kulit dapat terlihat dengan karat mata, gelisah, kulit dan bulu rusak, kulit menebal, kegatalan, serta dapat pula
menimbulkan kematian. Infestasi tungau dan caplak sering diikuti oleh infestasi virus, bakteri, dan mikroorganisme lain sehingga akan meningkatkan morbiditas dan mortalitas ternak.
Pengamatan di beberapa daerah di Jawa Barat menunjukkan, umumnya seekor ternak dapat terinfeksi 3-4 spesies yang didominasi oleh cacing H. contortus dan
Trichostrongylus spp. dengan rata-rata
prevalensi 67% (Kusumamihardja dan Zalizar 1992; Ridwan et al. 1996). Kerugian akibat infeksi nematoda, termasuk H.
contortus pada kambing, diperkirakan
mencapai Rp7 miliar/tabun (Rachmat et al.
1998). Kerugian akan bertambah besar bila
masalah haemonchosis tidak ditanggu-langi dengan sungguh-sungguh. Hal ini berkaitan dengan kendala yang harus
diatasi, seperti sistem betemak dengan cara cligembalakan serta pemberian antel-mintik secara terus-menerus dengan dosis rendah clan cara pemberian yang kurang tepat.
Kerugian akibat gangguan caplak di Indonesia, meski belum dilaporkan secara pasti, diperkirakan cukup besar, karena pada petemakan sapi di Amerika Serikat kerugian diperkirakan mencapai USS60 juta/tahun (Steelman 1976). Di Indonesia, gangguan caplak menjadi masalah di beberapa daerah di Sumatera, Jawa, Sulawesi Utara, Sulawesi Selatan, clan Sumbawa (Sigit et al. 1983). Demikian pula dengan tungau penyebab skabies meru-pakan masalah penting di Amerika Utara, Asia, Afrika Selatan, dan Eropa. Di Inggris tercatat lebih dari 3.500 kasus pada abad ke-19 (Kirkwood 1986). Di Indonesia, tungau dilaporkan menyerang ternak kambing dan menimbulkan kematian dengan prevalensi 4-11 % (Balai Penelitian Veteriner 1992; Budiantoro 2004).
Penanggulangan penyakit cacingan serta serangan tungau clan serangga lain pada ternak umumnya dilakukan dengan menggunakan antelmintik, pestisida atau akarisida secara berkala. Namun, pemakai-an obat-obatpemakai-an dari bahpemakai-an kimiawi secara terus-menerus akan menimbulkan resis-tensi dan residu pada ternak dan lingkung-an (Waller 1997; Larsen 2000). Selain dengan bahan kimiawi, penanggulangan penyakit cacingan serta serangan tungau, caplak, dan serangga dapat memanfaatkan cendawan sebagai agens pengendali hayati, terutama cendawan dari golongan kapang. Kapang pengendali parasit cacing nematoda disebut kapang nematofagus, dan kapang pengendali parasit serangga, tungau, caplak, dan kutu disebut kapang entomofagus. Perbaikan dan peningkatan produktivitas ternak dapat memanfaatkan khamir sebagai probiotik clan imunosti-mulan.
Hasil eksplorasi plasma nutfah cendawan Indonesia cukup banyak clan perlu diteliti untuk mengetahui mikroba, khususnya cendawan isolat lokal, yang dapat climanfaatkan untuk meningkatkan produktivitas ternak. Umumnya cendawan melalui beberapa tahapan seleksi setelah diisolasi clan diidentifikasi, seperti uji in
vitro clan in vivo, serta aplikasi pada skala laboratorium dan skala besar. Dari ber-bagai tahap pengujian tersebut, hanya sedikit cendawan yang terpilih sebagai pengendali hayati, probiotik atau imuno-stimulan. Hasil penelitian menunjukkan,
dari Sembilan isolat yang diuji, hanya satu yang dapat digunakan sebagai probiotik, dan dari sejumlah isolat D. flagrans yang ditemukan hanya sedikit yang dapat digu-nakan sebagai nematofagus (Agarwal et al. 2000; Ahmad 2005a). Tulisan ini mengulas beberapa cendawan isolat lokal yang bermanfaat bagi ternak, baik sebagai pengendali hayati, probiotik maupun imunostimulan.
CENDAWAN SEBAGAI PENGENDALI HAYATI
cendawan yang termasuk golongan ini, berclasarkan target utamanya, dapat dibagi menjadi dua, yaitu sebagai pengendali cacing nematoda (nematofagus) clan sebagai pengendali serangga, tungau, caplak, clan kutu (entomofagus). Meski-pun beberapa khamir dapat mematikan nematoda dan serangga, efektivitasnya rendah sehingga tidak digunakan lagi. Dari hasil penelitian, kapang nematofagus yang terpilih yaitu D. flagrans (Mendoza-De gives et al. 1999; Ahmad 2008), dan sebagai entomofagus adalah B. bassiana dan M anisopliae (Holder clan Keyhani 2005; Maranga et al. 2005).
Kapang Nematofagus
D. flagrans
Kapang D. flagrans mempunyai ciri khas, yang juga merupakan keunggulannya dibandingkan dengan cendawan nemato-fagus lain, yaitu mampu membentuk klamidospora oleh hifanya sendiri. Kapang davat tumbuh pada suhu 20-30°C.
Penambahan 20 larva nematoda/cM2 media agar akan menginduksi cendawan untuk membentuk perangkap, optimal pada suhu 30°C yang akan menghasilkan 700-800/ cm2 media agar/hari. Kapang
menghasil-kan dua macam spora, yaitu yang memiliki dinding tipis, akan menghasilkan konidia dengan jumlah terbatas pada saat cenda-wan berumur muda, dan yang memiliki dinding tebal, berupa klamidospora yang dihasilkan dari hifa yang matang. Selama proses penuaan, jumlah klamidospora meningkat sampai batas tertentu (Gronvoldet al. 1996).
Isolat lokal D. flagrans dari tanah asal
Bogor dapat tumbuh pada berbagai media, yaitu Corn Meal Agar (CMA), Potato
Dextrose Agar (PDA), dan Saboroud
Dextrose Agar (SDA). Tumbuh baik pada
suhu 22-30°C, dapat membuat jerat untuk larva infektif, mampu menurunkan larva instar 3 H. contortus, memerlukan cahaya, dan tahan disimpan selama 4 bulan pada kulkas (suhu 4-10°C) (Ahmad 2003) (Gambar 1). Kapang ini dapat digunakan dalam pengendalian parasit nematoda
Osepohagostomum dentatus dan Hyostro-ngylus rabidus pada babi, Cvsthostome,
Strongylus vulgaris dan Strongylus eden-tatum pada kuda, serta H. contortus dan Trichostrongylus colubriformis pada
domba dan sapi (Larsen et al. 1995; 1996; Nansen et al. 1996; Faedo et al. 1998; Larsen et al. 1998; Larsen 2000). Hasil penelitian tersebut menunjukkan bahwa kapang dapat mengendalikan larva nema-toda infektif secara optimal. Pemanfaatan
D. flagrans dapat dikombinasikan dengan
cendawan nematofagus lain seperti A.
oligospora (Larsen 2000).
Di Indonesia, pemanfaatan cendawan nematofagus mempunyai prospek
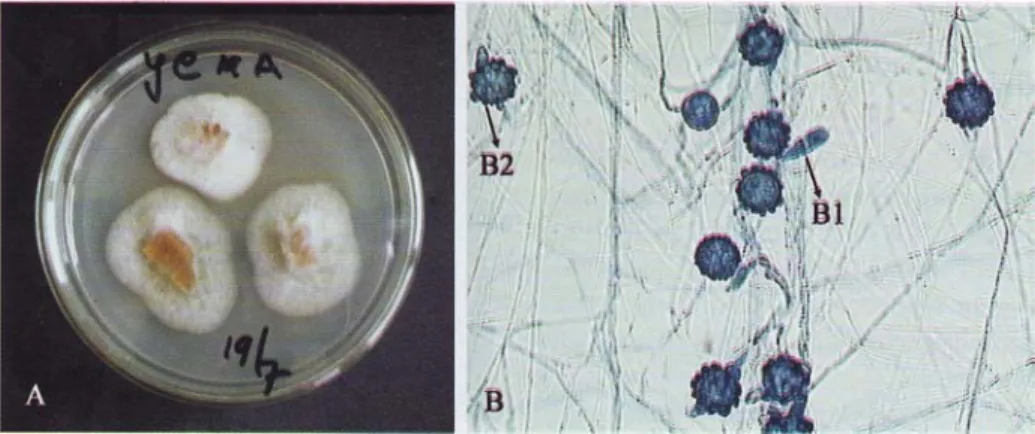
Gambar 1. Duddingtonia flagrans pada media potato dextrose agar (A); pengamatan
mikroskopik perbesaran 400 x dan pewarnaan laktofenol biru (B); konidia (BI); dan klamidospora (B2).
yang cerah karena didukung ketersediaan plasma nutfah dan kondisi temak rumi-nansia kecil. Penggunaan D. flagrans dengan closis 500.000 konidia mampu menurunkan larva H. contortus secara signifikan. Bila clikombinasikan dengan S.
cerevisiae secara signifikan mampu
menu-runkan jumlah larva H. contortus (Ahmad 2003;2008).
Mekanisme D. flagrans dalam, me-mangsa nematoda adalah dengan cara membuat perangkap bagi larva infektif Pada saat larva instar 3 bergerak mengenai hifa, kapang akan mengeluarkan zat kemo-atraktan yang menganclung sekresi. Larva yang mendekat kemudian akan melekat pada hifa clan dijerat oleh hifa vegetatif. Selanjutnya, kapang mensekresikan clan memproduksi enzim hidrolisis (kitinase dan protease) yang akan menguraikan kutikula untuk memudahkan hifa melakukan penetrasi (Gronvold et al. 1993; Ahman 2000; Meyer dan Wiebe 2003). Hifa vegetatif yang masuk ke dalam tubuh larva akan tumbuh dan berkembang hingga akhimya larva mati. Dalam proses mem-bunuh larva, kapang dibantu oleh nemato-toksin yang diduga sebagai lektin (Rosen 1996).
D. flagrans merupakan kapang
nematofagus pilihan untuk mengendalikan larva instar 3 cacing nematoda pada temak. Hal tersebut diclasarkan pada penelitian yang telah dilakukan sebelumnya (Gron-vold et al. 1993; Larsen 2000; Patra 2007). Kapang D. flagrans lebih efektif dan efisien mengendalikan larva nematoda di-banding Arthrobotrys spp., meski
Arthrobotrys oligospora dapat
memproduksi lebih banyak konidia dengan perlakuan yang sama. Hal ini karena D. flagrans mampu membentuk klamidospora (Mendoza-De Gives et al.
1999).
KapangEntomofagus
Beauveria bassiana
Koloni B. bassiana pada medium PDA yang diinkubasi pada suhu 25°C clan berumur 14 hari, membentuk lapisan seperti tepung (Gambar 2). Koloni pada bagian tepi mula-mula berwarna putih kemudian menjadi kuning pucat atau kemerahan.
B. bassiana mempunyai perangkat
penghasil spora (konidia) berukuran kecil dan lembut. Konidia yang berupa spora aseksual menghasilkan rangkaian akripetal pada sel-sel konidiogenus yang panjang
(sel penghasil konidia). Bentuk konidia bervariasi, yaitu globos, elips, silindris, clan koma. Konidia berbentuk elips ber-ukuran 2,90-4,20 µm x 1,80-2,50 µm, bentuk silindris 3,30-4,80 µm x 2,10-2,50 µm, clan bentuk koma 1,90-2,50 gm, hidup kosmopolitan, haploid. Konidia dengan jumlah yang banyak dihasilkan oleh hifa.
Hifa berukuran lebar 1-2 µm dan berkelompok dalam sekelompok sel-sel konidiogen berukuran 3-6 pm x 3µm. Selanjutnya, hifa bercabang-cabang clan menghasilkan sel-sel konidiogen kembali dengan bentuk seperti botol, leher kecil, dan panjang ranting dapat mencapai lebih dari 20 µm dan lebar 1µm. Cendawan ini tidak membentuk klamidospora, namun dapat membentuk blastospora.
Konidia dihasilkan dalam bentuk simpodial dari sel-sel induk yang terhenti pada ujungnya. Pertumbuhan konidia diinisiasi oleh sekumpulan konidia. Setelah itu, spora tumbuh dengan ukuran yang lebih tinggi karena akan berfungsi sebagai titik tumbuh. Pertumbuhan selanjutnya dimulai di bawah konidia berikutnya, setiap saat konidia dihasilkan pada ujung hifa dan dipakai terus, selanjutnya ujung-nya akan terus tumbuh. Dengan cara seperti ini, rangkaian konidia dihasilkan oleh konidia-konidia muda (rangkaian akropetal), dengan kepala konidia menjadi lebih panjang. Ketika seluruh konidia dihasilkan, ujung konidia penghubung dari sel-sel konidiogenus mempunyai pertumbuhan zig-zag dan mengikuti pertumbuhan asal seperti kepala (Brady 1979; Barron 2005).
B. bassiana merupakan parasit
agresif clan memiliki berbagai spesies inang serangga, meski ticlak menyerang semua jenis serangga. Namun karena
menginfeksi serangga pada berbagai umur dan stadium (telur, larva, dewasa), serang-an dapat menimbulkserang-an epizotik alam. Konidia dan miselia dalam jumlah banyak, bermassa padat dapat menyebar pada seluruh tubuh serangga yang terinfeksi (Feron 1981; Nankingan et al. 1994; Barron 2005). Caplak dan tungau dapat pula diserang (Brooks clan Wall 2002; Holder clan Keyhani 2005). Umumnya kadaver serangga secara optimum dapat meng-hasilkan konidia dan bersporulasi pada kelembapan lebih dari 95% (basah) dengan suhu 26-30°C (hangat). Bila kelembapan rendah maka proses sporulasi akan menurun (Fernandez 1994).
Umumnya kapang patogen mengin-feksi serangga melalui integumen. Hifa masuk ke dalam jaringan kemudian terus masuk ke hemosel (Barson 1977). B.
bassiana dapat menginfeksi serangga
melalui integumen, saluran pencernaan, trakhea, clan luka. Kapang ini dapat di-tularkan dari satu inang ke inang lain melalui konidia. Setelah masuk ke hemosel, cendawan terus tumbuh hingga seluruh bagian tubuh serangga tertutupi oleh miselim cendawan clan akhimya serangga mati. Pada kondisi yang sesuai, cendawan dapat tumbuh dan menembus kutikula di antara rugs tubuh serangga kemudian
membentuk konidia yang barn (Broome et
al. 1976; Maddox 1982).
serangga yang terinfeksi ditandai dengan gejala lemah, kurang aktif, dan pada kutikula ditemui bercak hitam yang menunjukkan tempat penetrasi cendawan, selanjutnya miselia tumbuh pada seluruh tubuh serangga dan berwarna putih. sporulasi pada larva mati terjadi Setelah 10 hari inkubasi (Barson 1977). Mekanisme infeksi didukung oleh enzim seperti
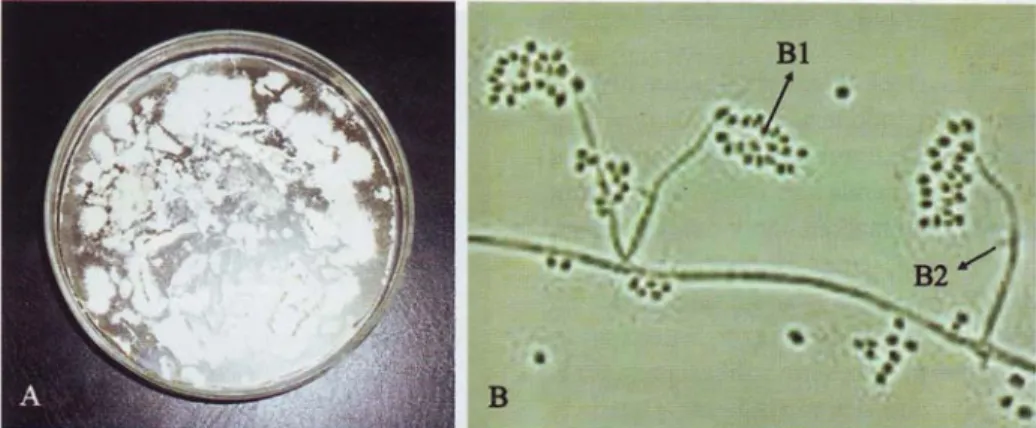
Gambar 2. Kapang Beauveria bassiana pada media potato dextrose agar (A); pengamatan mikroskopik perbesaran 400 x dan pewarnaan laktofenol biru (B); konidia (BI); dan hifa (B2).
protease, lipolitik, amilase, clan kitinase. Enzim-enzim tersebut mampu menghidro-lisis kompleks protein di dalam integumen (Brady 1979). Hal yang sama dapat terjadi pada tungau clan caplak.
Kapang Entomofagus
Metarhizium anisopliae
Kapang ini banyak ditemukan di dalam tanah, bersifat saprofit, dan umumnya dijumpai pada berbagai stadia serangga yang terinfeksi, tumbuh pada suhu 65-85°F dan kelembapan 30-90%. Kapang ini mempunyai koloni berwarna hijau zaitun, konidiofor dapat mencapai panjang 75 µm, bertumpuk-tumpuk diselubungi oleh konidia yang berbentuk apikal berukuran 6-9,50 jim x 1,50-3,90 rim, bercabang-cabang, berkelompok membentuk massa yang padat dan longgar (Gambar 3). Kapang ini dapat membunuh serangga, tungau, dan caplak seperti halnya B.
bassiana (Barnet 1969; Wikardi 2000;
Cloyd 2003; Geuther et al. 2004; Holder dan Keyhani 2005; Maranga et al. 2005).
Dalam menginfeksi serangga dan akarida, konidia berkecambah pada kutikula inang dan melakukan penetrasi dengan enzim hidrolisis (peptidase dan kitinase), lalu dengan bantuan tekanan mekanis enzim tersebut menghancurkan kulit dengan cara lisis. Setelah kapang masuk, koniclianya dengan cepat memper-banyak diri sehingga blastospora segera menyelaputi tubuh inang. Ketnatian inang disebabkan oleh kolonisasi miselia yang ekstensif sehingga menyebabkan starvasi, atau melalui racun yang dilepaskan pada saat penyerangan. Desikasi kadaver digu-nakan sebagai nutrisi dan air oleh hifa. Hifa memecah kutikula setelah serangga coati. Konidia bebas berkembang secara pasif atau aktif untak meneruskan siklus infeksi (Wikardi 2000; Onofre et al. 2001). Meka-nisme serupajuga ditemui pada caplak dan tungau yang terserang kapang ini.
CENDAWAN SEBAGAI PROBIOTIK DAN IMUNOSTIMULAN
Salah satu cendawan yang berpotensi sebagai probiotik dan imunostimulan adalah S. cerevisiae. cendawan ini ter-masuk dalam kelompok khamir. Pada saat berkembang biak, khamir akan membelah
diri dan menghasilkan tunas yang ber-kecambah multipolar. Spora berdiameter 5-10 µ. S. cerevisiae merupakan khamir bersel tunggal, yang berkembang biak dengan cepat secara seksual dan aseksual. Per-kembangbiakan melalui tunas kecambah multipolar dan tunas dapat terjadi pada seluruh permukaan Binding sel (Gambar 4).
Reproduksi seksual membentuk askospora di dalam askus. Dalam satu askus umumnya terdapat empat asko-spora dalam berbagai bentuk. Khamir ini mempunyai cin morfologi mikroskopis, membentuk blastospora (spora) bulat lonjong, silindris, oval atau bulat telur pendek dan panjangnya dipengaruhi oleh strain (Elliot 1994; Dube 1996).
Menurut besarnya sel, khamir dike-lompokkan menjadi tiga, yaitu yang selnya berukuran 3,50-10µx 5-19 [t,yang selnya benikuran 3-8µx 4-18 Vt, dan yang berukuran 2,50-7µx 4,50-18 p. Selain itu, terdapat sel berfilamen pada spora yang
berukuran lebih dari 30 µ dan berpseudo-miselium. Morfologi makroskopik me-nunjukkan koloni berbentuk bulat, berwarna putih, krem, abu-abu hingga kecoklatan, permukaan koloni berkilau sampai kusam, licin, dengan tekstur lunak (Lodder 1970; Barnet et al. 2000).
Peran
S. cerevisiae
sebagai ProbiotikProbiotik adalah makanan pelengkap berupa mikroba hidup yang memberikan keuntungan pada saluran pencernaan inang. Probiotik adalah bahan makanan yang tidak tercerna clan memberikan keuntungan pada inang melalui stimulasi selektif terhadap aktivitas satu atau sejumlah bakteri yang terdapat di dalam kolon (Roberfoid 2000). Lebih lanjut Fuller (1992) clan Karpinska et al. (2001) menjelaskan bahwa probiotik adalah imbuhan pakan berbentuk mikroba hidup
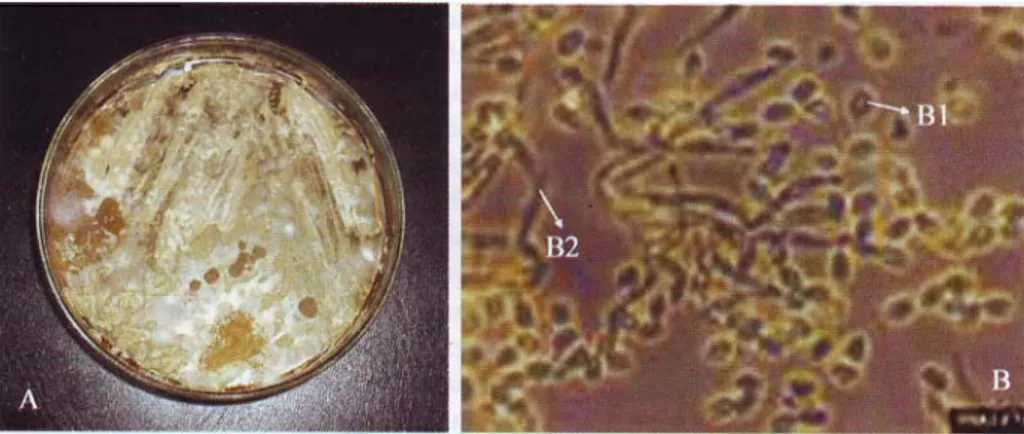
Gambar 3. Kapang Metarhizium anisopliae pada media potato dextrose agar (A); pengamatan mikroskopik perbesaran 400 x dan pewarnaan laktofenol biri (B); konidia (BI); dan hifa (B2).
Gambar 4. Kapang Saccharomyces cerevisiae pada media CAM (A); pengamatan mikroskopik perbesaran 400 x dan pewarnaan laktofenol biru (B); dan spora (BI).
yang menguntungkan dan mempengaruhi induk semang melalui perbaikan keseim-bangan mikroorganisme dalam saluran pencernaan. Di bidang peternakan, probiotik bermanfaat bagi kesehatan, produksi, dan pencegahan penyakit pada ternak. Salah satu probiotik adalah khamir S. cerevisiae.
Pemberian S. cerevisiae sebagai imbuhan mikroba hidup ke dalam tubuh akan mempengaruhi induk semang (unggas, ruminansia) melalui perbaikan keseimbangan mikroorganisme dalam saluran pencernaan. Pada ternak rumi-nansia, pemberian probiotik akan mening-katkan bakteri selulolitik dan asam laktat pada saluran pencernaan. Pada unggas, probiotik akan menambah jumlah mikroba yang menguntungkan dan menekan mikroba yang merugikan dengan cara berkompetisi untuk hidup di dalam saluran pencernaan.
Peran S. cerevisiae sebagai Imunostimulan
Secara sederhana imunostimulan adalah suatu bahan (material) yang bila diberikan pada hewan dan manusia dapat mening-katkan sistem pertahanan tubuh untuk menghadapi serangan penyakit. Imunosti-mulan meningkatkan "limfosit T" sebagai imunitas seluler yang penting untuk memproteksi tubuh dari bakteri dan virus intraseluler. "Limfosit B" juga meningkat untuk menambah imunitas humoral dan serum antibodi. Serum ini berfungsi untuk menetralkan endotoksin, sehingga pada akhirnya imunostimulan digunakan pada ternak untuk meningkatkan kemampuan melawan bakteri dan menurunkan waktu yang diperlukan untuk memperbanyak antibodi (Beta Glucan Research 2004).
Imunostimulan dapat digolongkan menjadi dua, yaitu yang bekerja secara spesifik dan nonspesifik. Beberapa materi atau substansi yang terlibat dalam proses yang spesifik adalah imunisasi aktif dan pasif, baik oleh virus, bakteri maupun cendawan, sedangkan yang nonspesifik berupa stimulasi limfosit dan makrofag (Tizard 1987).
Sistem pertahanan tubuh dapat di-bagi menjadi dua, yaitu kekebalan humoral dan seluler. Humoral terdapat pada darah, yang dikenal dengan antibodi, juga di-temukan sebagai molekul protein serum. Respons seluler berupa mekanisme fagositosis (membunuh kuman penyakit
dengan cara memakannya) dengan cara meningkatkan sensitivitas Reticulo Endo-thelial System (RES), yaitu sistem endo-telial berupa organ tubuh seperti ginjal, hati, limpa, dan timus.
Limfosit dapat digolongkan menjadi set B dan set T. Set T ditemukan dalam peredaran darah hewan tingkat tinggi dan memegang peranan penting dalam imu-nitas seluler. Set B pada mamalia dihasilkan oleh sumsum tulang dan pada unggas oleh bursa fabricius dan set T oleh timus. Selan-jutnya set B menghasilkan antibodi, yang merupakan salah satu elemen humoral untuk beradaptasi secara imunitas, sedangkan set T membantu set B dengan mengaktivasi makrofag untuk memper-tahankan tubuh terhadap infeksi mikroba.
Set makrofag merupakan salah satu set yang termasuk dalam kekebalan alami dengan cara menghasilkan substansi kimia. Selanjutnya substansi ini menjadi pertahanan set imun yang penting. Set-set tersebut akan membentuk kekebalan tubuh dengan cara menyerang Bel-Bel asing. Makrofag mempunyai reseptor pada set membran untuk tujuh macam residu gula. Saat reseptor berikatan dengan residu, makrofag diaktifkan dan kemudian menghasilkan sitokin. Sitokin inilah yang berfungsi sebagai pengatur respons imun tubuh (Tizard 1987). Sistem tersebut secara bersama-sama membentuk sistem pertahanan dalam tubuh melawan infeksi penyakit. Dengan demikian, imunostimu-lan sangat bermanfaat dalam meningkat-kan sistem kesehatan tubuh. Salah satu imunostimulan adalah beta-D glukan.
Beta-D glukan merupakan salah satu komponen dinding khamir yang esensial sebagai imunostimulan. Komponen ter-sebut mempunyai campuran yang unik yang efektivitas dan intensitasnya dalam sistem pertahanan tubuh melalui aktivasi set darah putih yang spesifik seperti makrofag dan set Natural Killer (NK). Beta-D glukan akan berikatan dengan permukaan set makrofag dan set NK, dan berfungsi sebagai pemicu proses aktivasi makrofag. Proses ini akan meningkatkan sirkulasi makrofag dalam tubuh untuk mencari benda-benda asing yang masuk ke dalam tubuh, selain menambah jumlah Bel-Bel makrofag (Life Source Basics 2002).
Beta-D glukan meningkatkan fungsi imun, termasuk fagositosis (kemampuan menangkap benda asing, partikel yang di-lepaskan sitokin; hormon interseluler IL-1, IL-6, GM-CSF, interferon) dan pernbuatan antigen. Beta-D glukan juga menstimulasi
RES dalam proses peningkatan jumlah makrofag, dan mengaktivasi sel-sel darah putih selain makrofag. Sel-sel tersebut ialah granulosit dan monosit. Beta-D glukan dapat berperan sebagai imuno-modulator (pengatur sistem kekebalan tubuh) untuk meningkatkan kemampuan set T, set B, dan makrofag dalam melawan infeksi penyakit, juga membantu mem-perbaiki jaringan yang rusak melalui proses regenerasi dan penyembuhan (Beta Glucan Research 2004 ).
PEMANFAATAN CENDAWAN UNTUK KESEHATAN TERNAK
Penelitian pengendalian biologic dengan cendawan nematofagus dan entomofagus, Berta peningkatan kesehatan dengan probiotik dan imunostimulan sudah lama dirintis (Larsen 2000; Brooks dan Wall 2002; Fox 2002; Ahmad 2005a; 2005b). Penelitian dimulai dari isolasi, identifikasi, dilanjutkan dengan pengujian secara in vitro dan in vivo dan diakhiri dengan aplikasi atau komersialisasi (Tabel 1).
Di Indonesia, penelitian cendawan nematofagus barn dimulai pada tahun 1990-an dan memasuki tahap aplikasi pada. tahun 2000-an, sedangkan di dunia sudah dimulai pada awal abad ke-19 (Larsen 2000). Penelitian dimulai dengan meng-isolasi dan mengidentifikasi isolat (Ahmad 1999). Setelah isolat diperoleh, dilakukan pengujian ketahanan hidup pada suasana asam, basa, enzim, dan suhu tertentu se-cara in vitro (Beriajaya dan Ahmad 1999). Selanjutnya dilakukan pengujian kemam-puan isolat dalam mereduksi H. contortus secara in vitro. Isolat yang lolos lalu diuji secara in vivo pada ternak (Ahmad 2001; Beriajaya et al. 2001). Proses pengujian terakhir dilakukan dengan aplikasi secara peroral atau langsung di lapangan. Hasil penelitian menunjukkan bahwa pemberian peroral memberikan hasil yang lebih baik. Bila dikombinasikan dengan S. cerevisiae dapat mereduksi larva H. contortus secara signifikan (Beriajaya et al. 2001; Ahmad 2008).
Penelitian pengendalian hayati dengan cendawan entomofagus berjalan lebih lambat dibandingkan dengan pe-nelitian cendawan nematofagus pada bidang peternakan. Umumnya kapang entomofagus digunakan untuk keperluan pertaman, untuk mengatasi hama serang-ga dan sejenisnya, hanya sedikit yang
diteliti untuk hewan dan itu pun terbatas pada caplak dan tungau (Brooks dan Wall 2002; Holder dan Keyhani 2005). Penelitian lanjutan yang berkembang adalah meng-ambil bagian gen yang menghasilkan toksin untuk caplak, tungau, dan serangga lain. Gen tersebut lalu dimanipulasi dan diperbanyak melalui rekayasa genetik sehingga dihasilkan produk antiserangga tanpa bentuk utuh dari kapangnya sendiri.
Penelitian S. cerevisiae sebagai probiotik dan imunostimulan berkembang lebih cepat dibanding untuk pengendali hayati. Shins et al. (1989) menyimpulkan bahwa S. cerevisiae termasuk salah satu mikroba yang banyak dipakai pada ternak sebagai probiotik, bersama-sama dengan bakteri dan cendawan lain seperti
Asper-gillus niger, A. oryzae, Bacillus punilus, B. centuss, Lactobacillus acidophilus, Saccharomyces crimers, Streptococcus lactic, dan S. termophilus.
Tedesco et al. (1994) menunjukkan adanya korelasi antara pemberian S.
cerevisiae dan populasi bakteri pada
kelinci, yaitu jumlah bakteri patogen menurun dan jumlah bakteri aerob dan anaerob yang menguntungkan dalam usus meningkat. Kumprecht et al. (1994) melaporkan pemberian campuran S.
cerevisiae dan Streptococcus faecum pada
ayam broiler menurunkan jumlah bakteri
Escheria coli dalam sekum hingga 50%.
Selanjutnya Kompiang (2002) meneliti penggunaan "khamir (ragi) laut" dengan
S. cerevisiae dalam pakan ayam dan
mendapatkan hasil yang positif, yaitu bobot badan meningkat setelah pem-berian S. cerevisiae. Selanjutnya Kum-prechtova et al. (2000) memberikan S.
cerevisiae 47 dengan dosis 200 g/100 kg
pakan untuk meningkatkan penampilan daging dan mengurangi bau amonia nitrogen pada feces ayam. Pemberian S.
cerevisiae juga meningkatkan penampilan
bobot ayam dan secara in vitro mampu menekan pertumbuhan S. typhimurium meski secara in vivo tidak memberikan hasil yang signifikan (Istiana et al. 2002). Meski tidak semuanya memberikan respons positif terhadap pemberian pakan imbuhan, pada sapi dapat meningkatkan produksi susu rata-rata 4,30% dan per-tambahan bobot badan rata-rata 8,70%. Pada domba, pemberian campuran S.
cerevisiae dan Bioplas pada ransum,
masing-masing dengan dosis 4 g Bioplas/ kg bobot badan dan 1 g S. cerevisiae yang mengandung 14 x 1011 koloni menghasilkan
konversi pakan sebesar 6 kg/kg pertam-bahan bobot badan (Ratnaningsih 2000).
Penelitian imunostimulan berkem-bang pesat. Penemuan substansi beta-D glukan berawal dari hasil Pillemer (1940)
dalam Life Source Basics (2002), yang
meneliti substansi yang memiliki kemam-puan menghasilkan aktivator mekanisme pertahanan tubuh yang disebut zymosan. Meski substansi tersebut mampu mensti-mulasi secara nonspesifik respons imun, zat aktifnya belum diketahui. Selanjutnya Diluzio (1970) dalam Life Source Basics
(2002) berhasil menemukan substansi tersebut dan komponen aktifnya adalah beta-D glukan. Salah satu bahan ini ter-dapat pada barley (Schizophyllum spp.) dan dinding khamir S. cerevisiae. Hasil penelitian laboratorium dan komersial secara umum membuktikan manfaat S.
cerevisiae pada ayam broiler dan petelur,
babi, ikan lele, sapi, dan udang sehingga dapat mengurangi biaya obat-obatan dan vaksinasi.
Beberapa contoh penggunaan S.
cerevisiae sebagai imunostimulan pada
hewan adalah sebagai berikut. Transfer gen beta-glukan pada udang dan mikro-organisme laut seperti mikroalga dan bakteri nonpatogenik lain dapat meningkatkan kekebalan tubuh. Pada lele dumbo, beta-glukan dengan dosis 750 mg/kg pakan dapat berperan sebagai imunostimulan yang positif terhadap respons kebal nonspesifik melalui uji tantang terhadap infeksi bakteri
Aeromonas hydrophila (Rukyani et al.
1987). Pada udang hitam (Penaeus
monodon), pemberian beta-glukan 1
g/kg pakan memperlihatkan peran imun yang positif terhadap kenaikan hematosit (Sitthipun et al. 2000). Pada udang dan ikan, penggunaan S. cerevisiae sebagai imunostimulan dengan dosis 50 mg/kg bobot badan dapat mengatasi serangan bakteri dan kuman lain seperti
Aeromonas salmonicida dan vibriosis
(Fox 2002).
PERMASALAHAN DALAM APLIKASI DI LAPANGAN
Penggunaan kapang entomofagus (B.
bassiana dan M. anisopliae) untuk
me-ngendalikan serangga hama pada tanaman perkebunan sudah banyak dilakukan, namun untuk pengendalian caplak dan tungau pada ternak barn pada tahap awal penelitian (Ahmad 2004). Identifikasi cendawan sangat penting dalam men-dapatkan cendawan entomofagus, karena bila salah memilih akan merusak tatanan fauna dalam lingkungan seperti kematian serangga yang menguntungkan atau berguna (Wikardi 2000). Karena itu, perlu dilakukan penelitian lanjutan. Untuk aplikasi kapang nematofagus (D.
flagrans) hanya perlu memasyarakatkannya sehingga penggunaan cendawan nematofagus dalam pengendalian penyakit pada ternak lebih lugs dibandingkan dengan
antel-mintik.
Tabel 1. cendawan dan kegunaannya untuk kesehatan ternak.
Spesies cendawan Kegunaan Jenis ternak Pustaka
Saccharomyces Probiotik, Ayam, domba, kelinci Rukyani et al. (1987); cerevisiae imunostimulan sapi, ikan, dan udang Shins et al. (1989);
Tedesco et al. (1994); Sitthipun et al. (2000); Wina 2000; Fox (2002); Ratnaningsih (2000); Ahmad (2005b). Duddingtonia Nematofagus Babi, kambing, domba, Larsen (2000);
flagrans kambing, kuda, dan sapi Ahmad (2005a); (2008) Metarhizium Entomofagus Kambing dan sapi Frazzon et al. (2000);
anisopliae Smith et al. (2000);
Onofre et al. (2001); Brooks dan Wall (2002); Cloyd (2003).
Beauveria Entomofagus Kambing dan sapi Holder dan Keyhani
bassiana (2005);
Memperhatikan hasil-hasil pene-litian terdahulu maka S. cerevisiae dapat
digunakan sebagai probiotik, namun be-berapa faktor harus dipertimbangkan seperti biaya produksi dalam skala besar, pengaruh buruk terhadap ternak, dan zat khasiat yang terkandung di dalamnya. Penggunaan S. cerevisiae sebagai imu-nostimulan hendaknya tidak melalui suntikan, tetapi secara oral bersama-sama pakan.
Peningkatan atau penurunan meka-nisme pertahanan tubuh bergantung pada jumlah glukan yang dikonsumsi ternak. Oleh karena itu, responsnya pada ternak
sangat bervariasi, bergantung pada ada atau'tidaknya receptor yang dikenal oleh komponen gula dari beta-D glukan. Hal ini berhubungan erat dengan struktur darah, di mana darah terdiri atas hemo-siania (molekul yang membawa oksigen) dan lektin (molekul yang beraksi terhadap, imun) (Life Source Basics 2002). Selain itu penggunaan imunostimulan beta-D glukan relatif lebih aman daripada antibiotik yang mempunyai efek resistensi. Perlakuan pada ruminansia berbeda dengan pada unggas atau ikan, sehingga dalam meng-aplikasikannya perlu mengetahui fisiologi
dan tingkah laku hewan dengan baik agar diperoleh hasil yang optimal.
KESIMPULAN
Hasil uji produktivitas dan aplikasi isolat lokal Indonesia kapang B. bassiana, D. flagrans, M anisopliae, dan khamir S.
cerevisiae sebagai pengendali hayati,
probiotik dan imunostimulan memberikan hasil yang baik sehingga dapat diman-faatkan untuk meningkatkan produktivitas dan kesehatan ternak.
DAFTAR PUSTAKA
Agarwal, N., D.N. Kamra, L.C. Chaudhary, A. Sahoo, and Pathak. 2000. Selection of Saccharomyces cerevisiae strains for use as a microbial feed additive. (http://www. B I ackwell. synergy. com/links/doi/ 10.1046/ J.1472-765X.2000.00826.X/Full/). [15 Oc-tober 2003].
Ahmad, R.Z. 1999. Pemakaian dua metode isolasi cendawan nematofagus dari tanah di daerah Bogor. him. 1.027-1.029. Prosiding Seminar Nasional Peternakan dan Veteriner, Jilid II. Pusat Penclitian dan Pengembangan Peter-nakan, Bogor.
Ahmad, R.Z. 2001. Isolasi dan Seleksi Cendawan Nematofagus untuk Pengendalian Haemon-chosis pada Domba. Tesis. Program Pasca-sarjana Institut Pertanian Bogor.
Ahmad, R.Z. 2003. Potensi Duddingtonia flagrans sebagai cendawan nematofagus. Jurnal Mikologi Kedokteran Indonesia 2003; (4): 14-20, 2004; 5(1-2): 14-20.
Ahmad, R.Z. 2004. Cendawan Metarhizium anisopliae sebagai pengendali hayati ekto-parasit caplak dan tungau pada ternak. Wartazoa 14(2): 73-78.
Ahmad, R.Z. 2005a. Pemanfaatan cendawan Arthrobotrys oligospora dan Duddingtonia flagrans untuk pengendalian Haemonchosis pada ruminansia kecil di Indonesia. Jurnal Penelitian dan Pengembangan Pertanian 24(4): 143-148.
Ahmad, R.Z. 2005b. Pemanfaatan khamir Saccharomyces cerevisiae untuk ternak. Wartazoa 15(1): 49-55.
Ahmad, R.Z. 2008. Efektivitas cendawan Duddingtonia flagrans dan Saccharomyces cerevisiae dalam pengendalian racing Haemonchus contortus pada domba. Diser-tasi. Sekolah Pascasarjana, Institut Pertanian Bogor.
Ahman, J. 2000. Extracellular serine proteases as virulence factors in nematophagous fungi: Molecular characterization and functional analysis of the PII protease in Arthrobotrys
oligospora. Dissertation, Lund University. Swedia. (http/www.lub.1u.se/cgi-bin/show-diss.pl?db- global & fname =sci -476 html. I - 3). [30 April 2006].
Alcxopoulus. C.J., C.W. Mims, and M. Blackwell. 1996. Introductory to Mycology. 4' ed. John Wiley and Sons. Inc., Newyork-Chichester-Brisbane-Toronto Singapore. p. 869. Balai Penclitian Veteriner. 1992. Kudis. Infor-
masi Teknis Penyakit Hewan: 22-24. Barnet, H.L. 1969. Illustrated Genera of
Im-perfect Fungi. 2"1 Ed. Burgess Publishing
Company, Mincapolis.
Barnet, J.A., R.W. Payne, and D. Yarrow. 2000. Yeasts. Characteristics and Identification. 3'" Ed. Cambridge Univ Press.
Barron. 2005. Beauveria bassiana. (http://www. Uoguelph.ca/-g barron/Miscelincous) [20 April 2005].
Batson, G. 1977. Laboratory evaluation of Beauveria bassiana as pathogen of the larvae stage of the large elm beetle, Soco-lytus. J. Vertebrata Pathol. 29(3): 361-366. Beriajaya dan R.Z. Ahmad, 1999. Cendawan
Arthrobotrys oligospora untuk pengendalian ncmatoda Haemonchus contortus pada domba. him. 980-985. Prosiding Seminar Nasional Peternakan dan Veteriner Jilid II. Pusat Penclitian dan Pengembangan Pe-ternakan, Bogor.
Beriajaya, R.Z. Ahmad, dan E. Kusumaningtyas. 2001. Efikasi nematofagus pada ternak domba dan kambing. him. 364-370. Prosi-ding Hasil Penclitian bagian Proyek. Reka-yasa Teknologi Petemakan. Pusat Penelitian dan.Pengembangan Peternakan, Bogor. Beta Glucan Research. 2004. Saccharomyces
cerevisiae. (http://www.betaglucan.orgi). [20 December 2004].
Brady, B.L.K. 1979. Pathogenic Fungi and Bacteria. Commonwealth Agricultural Bureaux, England.
Brooks, A.J. and R. Wall. 2002. Infection of Psoroptes mites the fungus Metarhizium anisopliae. Exp. Appl. Acarol. 25: 869-880. Broome, J.R., P.P. Sikorowski, and B.R.
Nore-marit. 1976. A mechanism of pathogenecity of Beauveria bassiana on larva of the imported fire and Solenopsis richteri. J. Vertebrata Pathol. 28: 87-91.
Budiantoro. 2004. Kerugian ckonomi akibat skabies dan kesulitan dalam pemberantasan-nya. him. 51-63. Prosiding Seminar Parasi-tologi dan Toksikologi Veteriner, 20-21 April 2004. Pusat Penclitian dan Pengem-bangan Peternakan, Bogor dan DFID-UK. Campos, R.A., W. Arruda, J.T. Boldo, and Siva.
2005. Boophilus microplus infection by B e au v e r i a a m o r p h a a n d B e au v e r i a bassiana: SEM analysis and regulation of subtilisin-like protcases and chitinases. Current Microbiol. 5: 257-261.
Cloyd, R.A. 2003. The Entomophatogenic Fungus Metarhizium anisopliae. University of Illinois. (http://www. Entomology.Wisc. Edu/mben/kyf607.html: 1-2). [10 December 2003].
Dube, H.C. 1996. An Introduction to Fungi. 2"' Ed. Vikas House PVT, Delhi.
Elliot, G.C. 1994. Reproduction in Fungi Genetical and Physiological Aspects. Botany Depar-tment Univ of Glasgow Chapman & Hall. London.
Faedo, M., E.H. Barnes, R.J. Dobson, and P.J. Waller. 1998. The potential of nemato-phagous fungi to control the free-living stages of nematode parasites of sheep: Pasture plot study with Duddingtonia flagrans. Vet. Parasitol. 76: 129-135. Fernandez, S. 1994. Study of Conidia Production
and Transmission of B. bassiana (balsamo) Vuill, in Colorado Potato Bettie (Leptintarsa decemlineata) PhD thesis, University of Maine.
Feron. 1981. Biological control of insect pest by entomogenous fungi. Annu. Rev. Entomol.: 409-442.
Fox, J.M. 2002. Immunology of fish and shrimp. (http://www.Sci.tamucc.edu/pals/maric/index /webpage/dlcc2.html). [15 October 2003]. Frazzon, A.P.M., I.D.S. Vas Junior, A. Masuda,
A. Scrank, and M.H. Vainstcin. 2000. In vitro
assessment of Metarhizium anisopliae
isolates to control the cattle tick Boophilus microplus. Abstract. Vet. Parasitol. 94(1-2): 117-125.
Fuller, R. 1992. Probiotics the Scientific Basis. Chapman & Hall. The University Press Cambridge.
Geuther, F.J., S.F. Steven, and S.G. Patricia. 2004. Virulence of Metarhizium anisopliae to embryos of the grass shrimp Palaemo-netespugio. (http://www. Isb.vt.edu/brarg/ brasym95/gcnthner95.htm:1-8) [15 March 2004].
Gronvold, J., J. Wolstrup, P. Nansen, S.A. Henriksen, M. Larsen, and J. Bresciani. 1993. Biological control of nematode parasites in cattle with nematode-trapping fungi: a survey of Danish studies. Vet. Parasitol. 48: 311-325.
Gronvold, J., P. Nansen, S.A. Henriksen, M. Larsen, J. Wolstrup, J. Bresciani, H. Rawat, and L. Fribert. 1996. Induction of traps by
Ostertagia ostertagi larvae, chlamydosporc production and growth rate in the nematode-trapping fungus Duddingtonia flagrans. J. Helminthol. 70: 291-297.
Holder, D.J. and N.O. Keyhani. 2005. Adhesion of the Entomopathogenic fungus Beauveria (cordyceps) bassiana to substrata. Appl. Environ. Microbiol. 71(9): 5.260-5.266, Istiana, E. Kusumaningtyas, D. Gholib, dan S.
Hastiono. 2002. Isolasi dan identifikasi
Saccharomyces cerevisiae beserta in vitro
terhadap (Salmonella typhimurium). hlm. 459-462. Prosiding Seminar Nasional Tck-nologi Petemakan dan Veteriner, Ciawi Bogor, 30 September-1 Oktober 2002. Pusat Pene-litian dan Pcngembangan Petemakan, Bogor. Karpinska, E., B. Blaszcak, G. Kosowska, A.
Degrski, M. Binek, and W.B. Borzemska. 2001. Growth of the intestinal anaerobes in the newly hatched chicks according to the feeding and providing with normal gut flora. Bull. Vet. Pulawary. 45: 105-109.
Kirkwood, A.C. 1986. History, biology and control of sheep scab. Parasitol. Today 11: 302-307.
Kompiang, I.P. 2002. Pengaruh ragi Saccharo-myces cerevisiae dan ragi laut sebagai pakan imbuhan probiotik terhadap kinerja unggas. Jurnal Ilmu Ternak dan Veteriner 7(1): 18-21.
Kumprecht, I., P. Zobac, Z. Gasnarek, and E. Robosova. 1994. The effect of continuous applications of probiotics preparations based on
S. cerevisiae var. elipsoideus and Strepto-coccus faecium C-68 (SF-68) on chicken
broiler yield. Zivocisma-yroba 39(6): 491-503.
Kumprechtova, D., P. Zobac, and I. Kumprecht. 2000. The effect of Saccharomyces cerevi-siae Sc 47 on chiken broiler performance an nitrogen output. Czech. J. Anim. Sci. 45: 169-177. (http://www. Buypro-biotics.Com/index 3.cfm? Book chapter id= 33). [20 December 2000].
Kusumamihardja, S. dan L. Zalizar. 1992. Pe-ngaruh musim pada hipobiose Haemonchus contortus dan fluktuasi populasi nematoda saluran pencernaan domba di Indramayu, Jawa Barat. hlm. 171-192. Prosiding Seminar Nasional Hasil Penelitian Perguruan Tinggi. Larsen, M., P. Nansen, J. Wolstrup, J. Gronvold,
S.A. Henrikesn, and A. Zorn. 1995. Bio-logical control of trichostrongyles in calves by fungus Duddingtonia flagrans fed to animals under natural grazing conditions. Vet. Parasitol. 60: 321-330.
Larsen, M., P. Nansen, C. Grondhal, S.M. Thamsborg, 1. Gronvold, 1. Wolstrup, S.A. Henriksen, and J. Monrad. 1996. The capacity of fungus Duddingtonia flagrans
to prevent strongyle infections in foals on pasture. Parasitology 113: 1-6.
Larsen. M., M. Faedo, P.J. Waller, and D.R. Hannessy. 1998. The potential of nemato-phagous fungi to control the free-living stages of nematodes parasites of sheep: Studies with Duddingtonia flagrans. Vet, Parasitol. 76: 121-128.
Larsen, M. 2000. Prospects for controlling animal parasitic nematodes by predacious microfungi. Parasitology 120: S121-SI31. Life Source Basics. 2002. WGP. Beta glucan.
(http: www. Life Source basics. Com/ beta_glucan.htm) [10 December 2002].
Lodder, J. 1970. The Yeast, A taxonomic study. 2" Ed. The Netherland, North Holland Co. Amsterdam.
Maddox, J.V. 1982. Use of Insects Pathogens in Pest Management. John Willey and Sons, New York, Chester, Brisbane, Toronto, Singapore.
Maranga, R.O., G.R. Kaaya, J.M. Mucke, and A. Haimnall. 2005. Effects of combining the fungi Beauveria bassiana and Metarhizium anisopliae on the mortality of the tick
Amblyomma variegatum (ixodidae) in relation to seasonal changes. Mycopathologia 154(4): 527-532.
Mendoza-De Gives, P., K.G. Davies, S.J. Clark, and J.M. Behnke. 1999. Predatory beha-viour of trapping fungi againts srf mutants of Caenorhabditis elegans and different plant and animal parasite nematodes. Para-sitology 119: 95-164.
Meyer, W.J. and M.G. Wiebe. 2003. Enzyme production by nematode-trapping fungus,
Duddingtonia flagrans. Abstract. J. Bio-technol. Letters 25: 791-795.
Nankingan, C.M., W.M. Ongenga-Latigo, G.B. Allard, and J. Ogwang. 1994. Studies on the
potential of Beauveria bassiana for the control the banana weevil cosmopolites sordidus Geermarin Uganda. J. African Crop Sci. (1): 300-302.
Nansen, P., M. Larsen, A. Roepstorff, J. Gronvold, J. Wostrup, and S.A.A. Henriksen. 1996. Control of Oe soph ag o s to mu m dentatum and Hystongylus rubidus in outdoor-reard pigs by daily feeding with the microfungus Dudding to n ia f lagron s.
Parasitol. Res. 82: 580-584.
Onofre, S.B., M.M. Cindia, M. Neiveia, and L.A. Joao. 2001. Pathogenicity of four strains entomopathogenic fungi against the bovine tick Boophilus microplus. Am. J. Vet. Res. 62: 1.478-1.480.
Patra, A.K. 2007. Nutritional management in organic livestock farming for improved ruminant health and production, an overview. Livestock Res. Rural Dev. 19: 1-21.
Rachmat, R., A. Rauf, dan M.Z. Kanto. 1998. Kontribusi getah pepaya dalam pengendalian penyakit casing pada kambing. hlm. 432-434. Prosiding Seminar Hortikultura. Kcrja Sama Fakultas Pertanian dan Kehutanan Universitas Hasanudin dengan Instalasi Pe-nelitian dan Pengkajian Pertanian Jencponto. Balai Pengkajian Tcknologi Pertanian Kendari.
Ratnaningsih, A. 2000. Pengaruh Pemberian Probiotik S. cerevisiae dan Bioplus pada Ransum Ternak Domba terhadap Konsumsi Bahan Kcring, Keccrnaan dan Konversi Ransum (in vivo). Skripsi Fakultas Peternakan Universitas Padjajaran, Bandung.
Ridwan, Y., S. Kusumamihardja, P. Dorny, and J. Vercruysse. 1996. The epidemiology of gastro-intestinal nematodes of sheep in West Java, Indonesia. Hemerazoa 78: 8-18.
Roberfoid, M.B. 2000. Probiotics and probiotics: are they functional foods 1-3 Am. J. Clinical New. 71(Suppl): 16.828-16.878.
Rosen, S. 1996. Fungal Lcctins. Molecular structure and function of a member of a novel lectin family. PhD Dissertation Lund University. (http://www. Lub.1u.se/cgi-bin.show-diss.pl?db= global& fname =sci-467.html). [4 Juni 2006].
Rukyani, A., E. Silvia, A. Sunarto, dan Taukhid. 1987. Pcningkatan respons kebal non-spesifik pada ikan lele Jumbo (Clarias spp.)
dengan pemberian imunostimulan (beta-glucan). Jumal Penelitian Perikanan Indone-sia 111(1): (Abstrak).
Shins, T., S. Hyung, K. Kyun, and A. Choong. 1989. Effects of CYC on the performance of dairy, beef cattle and swine. Seoul. Korea. Sigit, H.S., S. Parosoedjono, dan M.S. Akib. 1983.
Inventarisasi dan pemetaan parasit Indonesia tahap pertama. Ektoparasit. Laporan Pe-nelitian Proyek Pcningkatan dan Pengem-bangan Perguruan Tinggi Institut Pertanian Bogor.
Sitthipun, M., A.H. Kittikun, and K. Supa-mattayta. 2000. Immunostimulant and
vaccination in black tiger shrimp, Penaeus monodon Fabricius: Extraction of beta-glucan from yeast and its application in black tiger shrimp (Penaeus monodon Fabricius). (http://www.clib.psu.ac.th/acad-43/smol I.htm). [27 December 2000].
Smith, K.E., R. Wall, and N.P. French. 2000. The control of sheep scabmite Psoroptes ovis with entom op atho genic fungi. Vet. Piras4tol. 92: 97-105.
Soulsby, E.J.L. 1986. Helmiths, Protozoa and Arthropods of Domesticated Animal. 711 ed. Bailliere Tindall, London.
Steelman, C.P. 1976. Effects of external and internal arthropod parasites on domestic
livestock production. Ann. Rev. Entomol. 21: 55-178,
Tedesco, D., C. Castrovilli, G. Coni, D. Bartoli, V. Volltro, and F. Polidori. 1994. Use of probiotics in the feeding of neat rabbits: Effects on performance and intestinal microorganism. Rivista dj. Coniglicoltura 31(10): 41-46.
Tizard, I. 1987. Veterinary Immunology, an Introduction. W.B. Saunders Company, Philadelphia, USA.
Urqurhart, G.M., J. Armour, J.L. Duncan, A.M. Dunn, and F.W. Jennings. 1987. Veterinary Parasitology, Department of Veterinary Parasitology. Faculty of Veterinary Medicine,
The Univ of Glasgow, Scotland, Longman Scientific & Technical. Churchill Livingstone Inc, New York.
Waller, P.J. 1997. Sustainable helminth control of ruminants in developing countries. Vet. Parasitol. 71: 195-207.
Wikardi, E.A. 2000. Cendawan patogen serangga sebagai bahan baku insektisida. Pemanfaatan mikroba dan parasitoid dalam agroindustri tanaman rempah dan obat. Perkembangan Teknologi Tanaman Rempah dan Obat (XII): 21-28.
Wina, E. 2000. Pemanfaatan ragi (yeast) sebagai pakan imbuhan untuk meningkatkan produk-tivitas ternak ruminansia. Wartazoa 9(2): 50-56.