TINJAUAN PUSTAKA
Pengaruh Cekaman Aluminium Terhadap Tanaman
Pada kondisi asam atau pH 4 aluminium di dalam tanah dalam keadaan terlarut dalam bentuk Al3+ yaitu Al(H2O2)63+. Ketika pH meningkat, Al dalam bentuk Al(OH)2+ dan Al(OH)2+, dan ketika mendekati pH netral Al dalam bentuk Al(OH)3. Pada kondisi basa Al dalam bentuk Al(OH)4- (Marschner 1995).
Aluminium menyebabkan penghambatan dalam penyerapan nutrisi dan mineral sehingga menghambat pertumbuhan akar yaitu menghambat perpanjangan axis akar utama dan lateral sehingga akar menjadi pendek dan menebal (stubby). Ujung akar (tudung akar, meristem, dan zona perpanjangan) mengakumulasi Al lebih banyak, dan mengalami kerusakan fisik lebih parah dibandingkan jaringan akar lainnya (Delhaize & Ryan 1995). Aluminium juga menghambat pembelahan dan pemanjangan sel akibat gangguan sintesis DNA/RNA (Matsumoto 1991).
Tanaman mempunyai beberapa mekanisme toleransi terhadap cekaman aluminium (Al) sehingga mampu hidup dalam keadaan mendapat cekaman Al atau lingkungan yang asam (Kochian 1995). Mekanisme toleransi tanaman terhadap aluminium dapat dibagi dua yaitu mekanisme eksternal dan mekanisme internal. Mekanisme eksternal (exclusion mechanism) melalui imobilisasi Al pada dinding sel, induksi pH di daerah rhizosfer atau apoplas akar, permeabilitas selektif membran plasma terhadap Al, eksudasi fosfat dan efflux Al. Mekanisme internal mencakup pengkelatan Al di sitosol oleh asam organik, protein, atau ligan organik lain, mengurung Al di vakuola dan sintesis protein/ enzim yang toleran Al.
Pada tanaman tembakau (Nicotiana tobacum L.) ekspresi gen parA dan parB diinduksi oleh cekaman Al (Ezaki et al. 1997). Gen pAL111 (identik dengan parA yang menyandi auksin) dan gen pAL142 (identik dengan parB yang menyandi GST) pada tembakau ekspresinya diinduksi oleh aluminium dan defisiensi Pi (Ezaki et al. 1995). Pada Arabidopsis, cekaman Al menginduksi ekspresi beberapa gen yang juga berhubungan dengan sistem pertahanan terhadap patogen (defense-response) seperti gen GST, peroxidase dan blue copper binding protein (Richards et al. 1998). Gen yang diinduksi Al terlihat sama dengan gen
yang diinduksi oleh cekaman akibat defisiensi fosfat (Ezaki et al. 1995), keracunan metal (Snowden et al. 1995), infeksi patogen (Cruz-Ortega et al. 1997), dan cekaman oksidatif (Richard et al. 1998). Menurut Richard et al. (1998) ekspresi dari gen GST, peroxidase dan blue copper binding protein dipengaruhi oleh aktifitas spesies oksigen aktif (AOS) H2O2 saat tanaman mengalami cekaman.
Protein Heterotrimerik-G subunit α
Protein heterotrimerik-G adalah protein peripheral membran plasma yang menghadap ke permukaan dalam sel (menghadap ke sitosol). Protein ini merupakan reseptor membran sel dan berfungsi sebagai mediator penyampai pesan/signal dari luar sel (eksternal) ke molekul efektor sehingga menghasilkan respon intraseluler (Fujisawa et al. 2001). Protein heterotrimerik-G terdiri dari subunit α, β, γ (Fujisawa et al. 2001).
Protein heterotrimerik-G disebut protein G karena mengikat mononukleotida GDP dan GTP. Subunit α merupakan subunit yang mengatur pertukaran GTP-GDP pada mamalia (Fujisawa et al. 2001). Gα pada tanaman memiliki homologi yang sama dengan mamalia. Ada kemungkinan βγ mempunyai peranan secara langsung dalam meregulasi efektor dan interaksinya dengan reseptor (Ma 1994).
Protein G subunit α atau Gα terdapat pada plasma membran (Weiss et al. 1997; Iwasaki et al. 1997). Protein Gα mengaktifkan kanal Ca2+ pada plasma membran tomat (Aharon et al. 1998), meningkatkan level IP3 kedelai (Legendre et al. 1993) dan meningkatkan spesies oksigen aktif (AOS) H2O2 pada kultur sel kedelai (Legendre et al. 1992). Berdasarkan analisis mutasi pada gen Gα (dwarf1), maka Gα terlibat pada perpanjangan batang dan pembentukan benih padi (Fujisawa et al. 2001) dan ketahanan terhadap patogen (Suharsono et al. 2002). Protein heterotrimerik-G berperan dalam meregulasi ketahanan terhadap patogen (Aharon et al. 1998; Beffa et al. 1995; Legendre et al. 1993), regulasi lintasan biosintesis benzo phenathridine alkaloid (Mahady et al 1998), dan regulasi kanal K+ pada sel mesofil (Fairley-Grenot dan Asmann 1991; Li & Asmann 1993).
Pada kedelai ada dua kopi gen yang menyandikan protein heterotrimerik-G subunit α yaitu Sheterotrimerik-GA1 (Kim et al. 1995) dan Sheterotrimerik-GA2 (heterotrimerik-Gotor et al. 1996). Sheterotrimerik-GA1 diisolasi dari akar kedelai kultivar Williams. SGA1 juga telah diisolasi dari kedelai kultivar Lumut dan Slamet, dan memiliki kemiripan 91% dengan SGA1 dari kultivar Williams berdasarkan urutan nukleotidanya (Suharsono & Suharsono 2004). SGA2 diekspresikan pada semua organ vegetatif tanaman. Transkripsi terjadi pada semua jenis sel akar, daun dan batang (Gotor et al. 1996).
Gen Gα pada kultivar Slamet terinduksi pada 8 jam setelah perlakuan cekaman aluminium. Kemungkinan setelah 72 jam perlakuan cekaman Al beberapa sel sudah tidak aktif mengekspresikan gen Gα (Mashuda 2007).
Saat inaktif subunit α berikatan dengan GDP dan berasosiasi dengan βγ membentuk kompleks. Ketika ligan terikat pada permukaan sel reseptor, reseptor menjadi aktif dan mengkatalisis perubahan ikatan GDP pada subunit α menjadi GTP. Hal tersebut menyebabkan terjadinya perubahan konformasi subunit α sehingga akhirnya berpisah dengan βγ (disosiasi). Subunit α akan meregulasi efektor dengan cara berikatan pada efektor dan mengaktifkan signal transduksi seperti pada adenilat siklase. Protein heterotrimerik-G kembali tidak aktif ketika GTP diubah menjadi GDP dan subunit α kembali berasosiasi dengan βγ (Becker et al. 2000; Ma 1994).
Protein heterotrimerik-G meregulasi banyak efektor yang berada di bawahnya (downstream) seperti adenilat siklase, phosphalipaseC, dan efektor transducin (Ma 1994). Phosphoinositide spesifik phospholipase C (PLC) menghidrolisis phosphatidyl inositol 4,5-biphosphate (P1P2) menjadi 2 second messengers yaitu inositol 1,4,5-triphosphate (IP3) dan diacylglycerol (DAG). IP3 dapat mengikat reseptor membran seperti kanal Ca2+, dan melepas Ca2+ dari retikulum endoplasma ke dalam sitosol sehingga level Ca2+ meningkat. Protein kinase C akan teraktifkan oleh level Ca2+ yang meningkat dan DAG. Ion Ca2+ mengaktifkan protein kalmodulin yang akan mengaktifkan protein kinase dan fosfatase dalam satu atau lebih jalur persinyalan (Becker et al. 2000).
Akhir dari transduksi sinyal mengarah ke pengaturan satu atau lebih aktifitas seluler, melalui pengaktifan enzim spesifik maupun sintesis enzim atau protein lain dengan mengaktfikan atau menon-aktifkankan gen-gen spesifik untuk
memberikan respon terhadap sinyal yang diterima. Gluthathione S-Transferase (GST) merupakan gen yang banyak terlibat dalam menanggapi respon yang disebabkan oleh berbagai cekaman, diantaranya adalah cekaman patogen, cekaman aluminium. GST juga termasuk golongan gen yang berhubungan dengan sistem antioksidasi (Ezaki et al. 2004).
Glutathione S-Transferase
Tanaman memiliki mekanisme ketahanan yang sangat efektif untuk menghadapi cekaman akibat kerusakan oksidatif (induced oxidative damages). Salah satu protein yang terlibat dalam ketahanan sel terhadap cekaman oksidatif adalah GST (Glutathione S-Transferase). GST adalah protein dengan berat molekul sekitar 50 kDa, terdiri dari 2 subunit polipeptida (Dixon et al. 2002). GST mengkatalis pemindahan tripeptida glutathione (γ-glutamyl-cysteinyl-glycine; GSH) menjadi substrat (R-X) yang mengandung eletropilik reaktif untuk membentuk hasil reaksi S-glutathionylated polar (Dixon et al. 2002). Molekul-molekul yang telah berkonjugasi dengan GSH selanjutnya dikirim ke vakuola melalui ATP-binding transporter. Pengiriman ke vakuola bertujuan untuk membatasi efek dari penghambatan produk akhir GST dan melindung sel dari bahaya lebih lanjut akibat senyawa-senyawa yang berkonjugasi dengan GSH (Rea 1999).
GST dikelompokkan berdasarkan identitas sekuennya dan dibagi menjadi phi, tau, tetha, zeta dan lambda. Kelas tetha dan zeta banyak terdapat pada mamalia sedangkan selebihnya terdapat pada tanaman (Dixon et al. 2002). Pada Arabidopsis terdiri atas 48 gen GST dengan kelompok tau dan phi GST paling banyak. Masing-masing terdiri dari 28 kelompok tau, 13 phi, 3 dari kelompok theta, 2 zeta dan 2 lamda (Dixon et al. 2002). Pada jagung terdapat 12 kelompok phi, 28 tau dan 2 zeta sedangkan pada kedelai terdapat 20 gen dari kelompok tau, 1 zeta dan 4 phi (McGonigle et al. 2000).
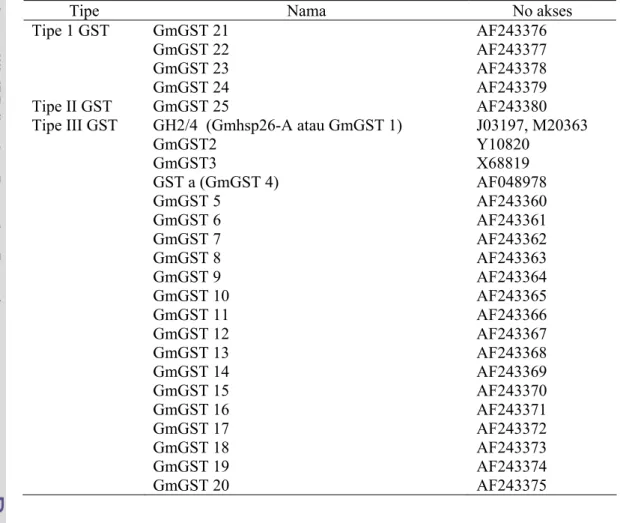
Berdasarkan kesamaan sekuennya, GST tanaman dibagi menjadi 3 tipe yaitu GSTI, GSTII dan GSTIII (Droog et al. 1995). Menurut McGonigle et al. (2000) pada kedelai terdapat 25 sekuen GST (Tabel 1). Pada kedelai ada 4 tipe GSTI, 1 tipe GSTII dan 20 tipe GSTIII. Individual cDNA pada kedelai sebanyak
6% tipe GSTI, 2% cDNA tipe GSTII, dan 92% adalah tipe GSTIII. Khusus sekuen GST tipe III yaitu GmGST8 terdapat sebanyak 33%.
Struktur GST tanaman memiliki kesamaan yang tinggi dengan struktur GST mamalia. GST ditemukan pada setiap tahap perkembangan tanaman dari awal perkecambahan sampai tua dan terdapat di setiap jaringan tanaman (McGonigle et al. 2000).
Tabel 1. GST yang terdapat pada kedelai (McGonigle et al. 2000)
Tipe Nama No akses
Tipe 1 GST Tipe II GST Tipe III GST GmGST 21 GmGST 22 GmGST 23 GmGST 24 GmGST 25 GH2/4 (Gmhsp26-A atau GmGST 1) GmGST2 GmGST3 GST a (GmGST 4) GmGST 5 GmGST 6 GmGST 7 GmGST 8 GmGST 9 GmGST 10 GmGST 11 GmGST 12 GmGST 13 GmGST 14 GmGST 15 GmGST 16 GmGST 17 GmGST 18 GmGST 19 GmGST 20 AF243376 AF243377 AF243378 AF243379 AF243380 J03197, M20363 Y10820 X68819 AF048978 AF243360 AF243361 AF243362 AF243363 AF243364 AF243365 AF243366 AF243367 AF243368 AF243369 AF243370 AF243371 AF243372 AF243373 AF243374 AF243375
Ekspresi gen GST8 pada akar kultivar Slamet diinduksi oleh cekaman Al yaitu pada 8 jam dan 24 jam setelah perlakuan cekaman Al. GST8 tidak terekspresi setelah 48 jam perlakuan cekaman Al. Lamanya perlakuan tidak menyebabkan meningkatnya ekspresi GST8 tetapi menurunkan ekspresi GST8. Ini menunjukkan bahwa pada kultivar Slamet gen GST8 memberi respon terhadap cekaman Al di awal (Mashuda 2007). Sedangkan ekspresi gen GST12 pada akar kultivar Slamet terinduksi oleh cekaman pH rendah dan cekaman Al. Gen GST12
memperlihatkan ekspresi tertinggi pada 8 jam setelah perlakuan cekaman Al. Kemudian seiring lamanya waktu cekaman menyebabkan penurunan ekspresi gen GST12 (Mashuda 2007). Pola ekspresi ini menunjukkan bahwa ada keterlibatan gen GST12 terhadap cekaman Al.
Aktivitas gen GST akan meningkat sebagai respon rangsangan adanya kerusakan oksidatif (Marrs 1996; Droog 1997). Gen GST diinduksi oleh berbagai rangsangan dari lingkungan meliputi serangan jamur, cekaman dehidrasi, etilen, pelukaan (wounding) (Marrs 1996). Aktifitas gen GST meningkat akibat pemanasan dan kondisi cekaman garam pada benih transgenik tembakau (Roxas et al. 1997). Aktivitas GST di akar meningkat lebih tajam dibanding di batang pada Triticum aestivum akibat cekaman osmotik (Galle et al. 2005). Auksin, hormon-hormon, logam berat, H2O2, garam, suhu (heat shock), cekaman lingkungan telah menginduksi promotor GH2/4 yang menyandi GST pada tembakau transgenik (Ulmasov et al. 1995). Gen GST berperan dalam detoksifikasi dan proteksi sel dari cekaman oksidatif, melindungi dari cekaman biotik dan abiotik meliputi serangan patogen, xenobiotik dan racun logam berat yang merupakan respon tanaman terhadap perubahan kondisi lingkungan (Ulmasov et al. 1995; Droog et al. 1993; Marrs 1996).