TINJAUAN PUSTAKA
1. Penyakit Pustul Bakteri
Penyakit pustul bakteri disebabkan oleh Xanthomonas uxonopod~.~ pv. glycmes (Nakano) Dye Vauterin (Xag). Bakteri Xag berukuran 0,5-0,9 x 1,4-2,3 pm, berbentuk batang, memiliki satu flagel polar dan merupakan bakteri Gram negatif. Bakteri Xag dapat mencairkan gelatin dalam waktu 6 hari, membentuk asam sitrat dari arabinosa, glukosa, manosa, selobiosa, trehalosa, dan sukrosa dalam 2 hari, menguraikan protein susu dalam 13 hari, menghidrolisis pati dalam 2 hari, serta masih dapat tumbuh pada suhu 36" C. Sifat fisik yang mudah diamati ialah bentuk permukaan koloni mucold, sirkuler dengan tepi lembut yang cembung dan benvarna kuning pada agar yeast dextrose carbonate (YDC) (Moffett dan Croft 1983, Schaad dan Stall 1988).
Bakteri masuk ke dalam jaringan tanaman kedelai melalui stomata, hidatoda atau luka, kemudian berkembang di dalam sel daun kedelai. Pemencaran bakteri antar tanaman dibantu oleh percikan air hujan yang disertai dengan angin. Epidemi penyakit pustul bakteri terjadi pada kedelai yang masa pembungaannya bertepatan dengan cuaca basah (Hartman et a/. 1999). Gejala awal penyakit pustul berupa bercak hijau pucat sebesar mata jarum. Bercak dapat meluas hingga mencapai diameter 3 mm dengan bagian tengah bercak menonjol benvarna kecoklatan. Pustul ini terjadi karena hipertrofi dan hiperplasia. Pada gejala lanjut, daun tampak mengering dipenuhi bercak pustul, mudah sobek serta gugur lebih awal. Daun
kedelai yang gugur prematur menyebabkan produktivitas tanaman kedelai menurun berkaitan dengan jumlah polong hampa yang tinggi (Sinclair dan Backman 1989).
Gejala pustul bakteri berbeda dengan pustul karat yaitu pustul bakteri terdapat pada daun-daun muda pada bagian atas tanaman, sedangkan pustul karat terdapat mula-mula pada daun-daun tua di bagian bawah lalu berkembang ke daun yang lebih muda di bagian atas. Pada permukaan bawah helai daun kedelai yang bergejala pustul karat yang disebabkan oleh cendawan Phakopsora pachyrlzzz Syd. terdapat urediosorus yang tampak seperti gumpalan tanah benvarna coklat yang terasa kasar jika diraba (Hartman et a/. 1999).
Stadium kritis tanaman kedelai terhadap bakteri pustul yaitu 40 hari setelah tanam. Gejala pustul muncul pada daun muda dan gejala semakin parah pada kedelai berumur 60 hari setelah tanam terutama pada saat pengisian polong (Semangun 1990). Inokulasi dengan cara penyemprotan suspensi Xag berkerapatan 10' CFUIml pada daun kedelai berumur 25 hari setelah tanam menunjukkan bahwa periode inkubasi bervariasi antara 8 hingga 27 hari bergantung kepada ketahanan varietas kedelai. Suhu dan kelembaban udara relatif yang optimum selama masa inkubasi adalah 29' C dan 89 % (Dirmawati 1996).
Bakteri pustul kedelai dapat menyerang kacang buncis, kacang panjang, bahkan di India menyerang kecipir (lJo11chos hzflorus) (Hartman et a/. 1999). Hal ini sesuai dengan laporan Aini (1992) bahwa polong kacang buncis (Phaseolus vulgurw) dan kacang panjang (Vzgna smensu) dapat diinfeksi oleh bakteri Xag melalui inokulasi buatan. Namun secara alami tidak pernah diperoleh gejala pustul pada polong kacang buncis dan kacang panjang. Bakteri penyebab pustul dapat bertahan dalam serasah kedelai di permukaan tanah, tetapi tidak hidup dalam serasah
kedelai yang dibenamkan minimum 15 cm di dalam tanah selama satu bulan. Menurut Sinclair dan Backman (1992), di Amerika Serikat bakteri pustul kedelai dilaporkan menyerang gulma Brunnichza cirrhosa Gaertn.
2. Pseudomonasfluorescens Migula sebagai Agens Biokontrol
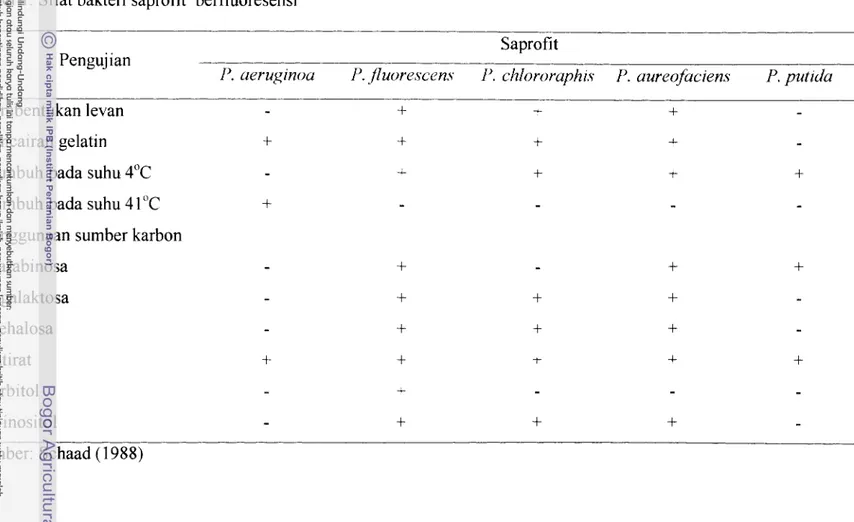
P.seudomona.s fluorescens Migula. merupakan bakteri saprofit seperti halnya /-'. uerugmosa P. putzda, P. chlororuphw, dan P. aureofaczens. Sifat fisik dan kimia bakteri saprofit berfluoresensi disajikan pada Tabel I . Beberapa peneliti melaporkan bahwa P. JEuorescens dapat digunakan sebagai agens biokontrol berbagai penyakit tanaman. Menurut Fravel et a/. (1998), I-'. ,fluorescens A506 yang diformulasikan dalam bentuk tepung dan secara komersial dikenal dengan merek dagang Blight Ban A506 efektif mengendalikan penyakit Erwmza amylovora pada buah apel, pir, kentang, tomat, dan stroberi. Formulasi suspensi P. fluorescens A506 dengan merek dagang Victus dinyatakan efektif mengendalikan I-'. tolu,~zl pada jamur merang.
Mekanisme P. fluorescens mengendalikan penyakit tumbuhan diantaranya dengan memproduksi siderofor, hidrogen sianida, dan antibiotik pirolnitrin, pyoluteorin serta 2,4-diasetilfluoroglusinol. Antibiotik 2,4-diasetilfluoroglusinol diproduksi P. ,fluore.scen.s P f 5 pada medium yang mengandung glukosa. Pada medium yang mengandung gliserol, bakteri ini tidak membentuk antibiotik 2,4- diasetilfluoroglusino1 ( Rodriguez dan Pfender 1997). Bakteri P. ,fluore.scens memproduksi senyawa berfluoresensi benvarna kuning kehijauan. Senyawa fluoresein atau pioverdin berpendar di bawah cahaya ultraviolet (panjang gelombang 266 nm).
Tabel 1 . Sifat bakteri saprofit berfluoresensi
Saprofit Pengujian
1'. aeruginou P. f1uorescen.s P. cl~lororaphrs P. aureofaciens P, putzda -
Pembentukan levan
-
+
+
+
-Pencairan gelatin Tulnbuh pada suhu 4°C Tumbuh pada suhu 4 1 "C
Penggunaan sumber karbon
D-galaktosa Trehalosa Butirat Sorbitol
Pigrnen pioverdin terbentuk pada medium King's B (Schaad 1988). Pigrnen pioverdin merupakan molekul dengan berat 400-2000 dalton yang tidak larut dalam kloroform, tetapi larut dalam air. Senyawa pioverdin mengikat ion ~ e ) ' (Abdallah
199 1, Weisbeek dan Gerrits 2000).
Siderofor pada mikroorganisme atau tumbuhan berfungsi untuk pengambilan unsur besi dalam tanah yang pada umumnya terikat pada batu-batuan. Kelarutan ion besi di tanah yang dapat dimanfaatkan oleh organisme hanya 10-l8 M (Weisbeek dan Gerrits 2000). Menurut Hemming (l990), konsentrasi ion besi yang dapat larut dari mineral Fe(OH)3 dalam tanah 2 x 1
o-)'
M.Di lain pihak, semua makhluk hidup memerlukan ion ~ e ) + untuk pertumbuhan sel. Unsur besi berfungsi sebagai kofaktor enzim metabolisme dalam makhluk hidup. Tanaman membutuhkan ion besi 1 0 ' ~ hingga M, sedangkan mikroorganisme memerlukan lo-' hingga lo-' M. Tukey (1 990) melaporkan bahwa ion besi yang terkandung dalam eksudat tanaman sebanyak M.
Bakteri P. putida WCS358 membentuk senyawa siderofor yang disebut pseudobaktin. Pseudobaktin adalah senyawa kuinol yang dirangkaikan oleh rantai oligopeptida. Panjang rangkaian serta komposisi oligopeptida bervariasi dan menjadi karakteristik pembeda pseudobaktin. Pengikatan ion besi dijembatani oleh senyawa katekolat, hidroksamat, atau gabungan antara hidroksamat dan asam hidroksi aspartik. Selanjutnya setelah bakteri mengikat ion Fe3+, konsentrasi Fe3+di dalam sel selalu dijaga.
Kekurangan ion besi mengurangi pertumbuhan bakteri sedangkan kelebihan zat besi akan meracuni sel bakteri. Sistem pengkelatan ion besi diatur oleh konsentrasi ion besi. Pembentukan senyawa pengkelat zat besi meningkat jika
bakteri ditumbuhkan pada medium dengan kandungan ion ~ e " terbatas. Pengaturan ion besi yang dibutuhkan bakteri dikendalikan oleh gen Fur yang terdapat dalam sitoplasma. Menurut Litwin et al. (1993) dalam Weisbeek dan Gerrits (2000), gen Fur juga berperan dalam virulensi patogen.
Neilands dan Nakamura (1991) menyatakan bahwa molekul FeC13 sebanyak 0,l hingga 5,O pM per ml medium pertumbuhan dapat menghambat pembentukan siderofor. Namun unsur sebanyak 0,03 pM per ml medium tumbuh memicu pembentukan siderofor (dapat diamati pada kisaran panjang gelombang 400 hingga 600 nm). Menurut Scher dan Baker (1982), siderofor yang berupa katekol hidroksamat dalam filtrat bakteri dapat dideteksi dengan panjang gelombang 4 10 nm.
Menurut Mount dan Lacy (1982), pioverdin yang diproduksi oleh P. jluorescens B 10 bersifat antibiotik terhadap Fusarzum spp. Neilands dan Nakamura ( 199 1 ) menyatakan bahwa siderofor pioverdin atau pseudobaktin yang diproduksi oleh P. jluorescens WCS374, P. Jluorescens ATCC 13525, dan P.Jluorescens 17400 adalah kromoforpeptida. Kromoforpeptida siderofor adalah struktur seperti membran yang mengandung pigrnen dan mengikat mineral besi. Menurut Matzanke (1991), kromoforpeptida siderofor yang diproduksi oleh I'.~eudomonas spp. adalah gabungan katekolat dan hidroksamat yaitu Tris N metiltioformohidroksamat yang dapat mengikat dua molekul Fe3+ Di lain pihak, senyawa Tris N metilformohidroksamat diketahui dapat membunuh bakteri maupun cendawan. Weisbeek dan Gerrits (2000) menyatakan bahwa sistem pengkelatan besi berkaitan dengan produksi metabolit sekunder yang bersifat antagonis terhadap mikroorganisme lain.
Abdallah (199 1) menyatakan bahwa siderofor yang disebut pioverdin atau pseudobaktin diekskresikan ke luar sel oleh Pseudomonas spp. untuk mengikat ion ~e"dan mentranspornya ke dalam sel. Pioverdin dibentuk setelah inkubasi selama 40 jam yaitu pada fase stasioner.
Selain siderofor, Alstrom dan Burns (1989) menyatakan bahwa P. fluorescens dapat memproduksi metabolit volatil diantaranya hidrogen sianida. Jumlah hidrogen sianida yang diproduksi P. fluorescens CHAO dalam kultur cair selama 36 jam kultivasi adalah 37 nmollml. Voisard et a/. (1989) melaporkan bahwa hidrogen sianida yang diproduksi oleh P. fluorescens CHAO dapat menekan infeksi 7'hreluvropsis baszcola pada akar tembakau, tetapi tidak meracuni akar tembakau.
3. Bacillus subtilis (Ehrenberg) Cohn sebagai Agens Biokontrol
Bacillus subtilis merupakan bakteri Gram positif yang membentuk spora. Spora berfungsi untuk bertahan hidup antara lain pada suhu lingkungan 70' hingga 100' C. Pelczar dan Chan (1988) menyatakan bahwa resistensi B. subtilis terhadap panas hilang jika bakteri disterilisasi dengan panas lembab maupun kering. Spora tidak tahan terhadap sterilisasi dengan panas lembab bersuhu 100" C selama 2 jam, sedangkan jika disterilisasi dengan panas kering bersuhu 11 5" C memerlukan waktu 40 menit.
Pada umumnya B. subtilis dapat digunakan sebagai agens biokontrol terhadap patogen tanaman walaupun diketahui terdapat strain yang dapat membusukkan biji kedelai. Sinclair dan Backman (1989) melaporkan bahwa biji kedelai yang
diinokulasi B. suhtilis strain virulen (isolat VS) pada suhu 30-35" Can kelembaban udara relatif 98 % akan menunjukkan busuk berlendir 5 hari setelah inokulasi.
Sel H. suhtilis berbentuk batang, berukuran 0,3-2,2 x 1,2-7,O pm . dan mempunyai flagel peritrikus (Pelczar dan Chan 1988). Bakteri ini dapat membentuk endospora yang berbentuk oval di bagian sentral sel. Bakteri ini dapat tumbuh pada suhu 4 5 ' ~ ~ pH 5-7, NaCl 7 %, menghidrolisis pati, serta membentuk asam sitrat dari karbohidrat glukosa, arabinosa, manitol, dan silosa (Leary dan Chun 1988).
Bakteri B. subtili,~ yang bersifat antagonis mampu menekan pertumbuhan mikroorganisme lain karena memproduksi antibiotik berupa lipopeptida yang disebut basitrasin dengan mekanisme merusak membran sel bakteri (Leary dan Chan 1988). Jenis metabolit sekunder lain yang diproduksi B. suhtilis adalah biosurfaktan yang disebut surfaktin atau subtilisin. Surfaktin merupakan lipopeptida siklik yang berfungsi menurunkan tegangan permukaan air dan juga bersifat antibiotik (Hommel dan Ratledge 1993, Desai dan Desai 1993).
Menurut Mulligan dan Gibbs (1993), produksi optimum surfaktin B.subtilis dipengaruhi oleh komposisi medium tumbuh. Setiap liter medium pertumbuhan dengan komposisi glukosa 4 %, amonium nitrat 0,4 %, dan Fe SO4 0,32 mM meningkatkan produksi surfaktin dari 0,O g menjadi 8,O g selama 20 hingga 40 jam inkubasi. Produksi surfaktin dipengaruhi oleh jumlah minimum zat besi di dalam setiap liter medium pertumbuhan yaitu 0,2 ppm.
4. Komponen Bioaktif Tanaman sebagai Bakterisida Botani
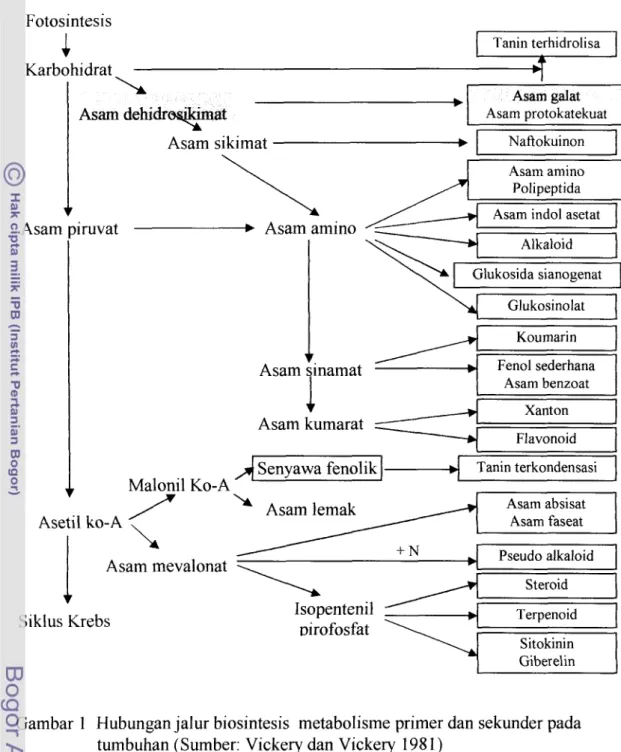
Fotosintesis atau fiksasi karbon dioksida merupakan proses biosintesis karbohidrat dalam tumbuhan berhijau daun. Karbohidrat merupakan salah satu bahan awal atau prekursor metabolisme sekunder. Metabolit sekunder adalah senyawa intermediet yang diproduksi dalam jumlah sedikit dan tidak diperlukan untuk fungsi pertumbuhan normal tanaman. Metabolit sekunder berfungsi sebagai antimikroba atau penolak serangga. Selama kondisi tanaman tidak menguntungkan karena terdapat stimuli lingkungan atau mikroba, maka jumlah metabolit sekunder meningkat dan mungkin tetap diproduksi setelah kondisi membaik. Metabolit yang berupa senyawa berbobot molekul rendah ini terakumulasi di dalam tanarnan (Sastrohamidjojo 1996, Vickery dan Vickery 198 1). Gambar 1 menunjukkan hubungan antara jalur metabolisme primer dan sekunder sehingga mengarah pada produksi komponen bioaktif tanaman, seperti halnya asam galat.
Asam galat merupakan penyusun senyawa tanin tumbuhan. Di dalam tumbuhan, asam galat berbentuk ester dengan alkohol. Asam galat tergolong dalam senyawa fenolik. Asam galat diproduksi tumbuhan normal dengan konsentrasi rendah. Konsentrasi asam galat meningkat pada reaksi ketahanan tumbuhan terhadap serangga maupun patogen (Mann 1987, Harborne 1987).
Fotosintesis
4
[ Tanin terhidrolisa1
Karbohidrat -
\
Asam galatAsam dehidrw&pat Asam protokatekuat
Asam sikimat
.
1
[
Naflokuinon\ I 1
I
Asam aminoAsam indol asetat Asam piruvat
Alkaloid Glukosida sianogenat
Koumarin
Asam sinamat Fenol sederhana
I Asam benzoat
I 1
A
senvawafenoi;I;1 Tanin terkondensasi1
4
AAsam kumarat --
4
7 1KO-A'
I I JAsarn lemak
4
Asam absisatAsetil ko-A , Asam faseat
Xanton
J
FlavonoidL
+ N Asam mevalonat Siklus Krebs Steroid Isopentenil Terpenoid oirofosfat1
-Sitokinin7
Giberelin1
Gambar 1 Hubungan jalur biosintesis metabolisme primer dan sekunder pada tumbuhan (Sumber: Vickery dan Vickery 198 1)
Keterangan: Pada kondisi tanaman normal reaksi lebih mengarah kepada perubahan glukosa (karbohidrat) menjadi Asetil ko-A yang masuk ke dalam Siklus Krebs. Pada kondisi abnormal tanalnan karena kondisi lingkungan yang tidak menguntungkan maka reaksi mengarah kepada perubahan glukosa (karbohidrat) menjadi molekul metabolit sekunder melalui siklus asam dehidrosikimat dan asam sikimat.
Asam galat dibentuk dari aromatisasi asam dehidrosikimat (Manitto 1991, Vickery dan Vickery 1981). Menurut Prindle dan Wright (1991), asam galat mempunyai bobot molekul500-3000 dalton. Asam galat merupakan senyawa dengan struktur cincin aromatik yang mengandung satu atau dua gugus hidroksil dan dapat membentuk ikatan silang yang stabil dengan protein, selulosa, dan pektin. Asam galat terdapat dalam vakuola sel dan mudah larut dalam air. Menurut Harborne (1987), asam galat terdapat dalam daun sebagai fraksi yang larut dalam etanol, terutama pada daun yang membentuk lignin. Menurut Manitto (1991), asam galat dibentuk dalam daun maupun jaringan tumbuhan yang membentuk puru (gall) atau hipertrofi sel.
Dalam peranannya sebagai zat antibakteri, asam galat merusak perrneabilitas membran bakteri sehingga membocorkan metabolit dalam sel bakteri (Manitto 199 1 ).
Pelczar dan Chan (1987) menyatakan bahwa asam galat membentuk ikatan yang stabil dengan membran sitoplasma bakteri sehingga merusak integritas membran sitoplasma dan mengakibatkan kebocoran substansi dalam sel bakteri. Oleh karena membran sel merupakan lokasi beberapa jenis enzim, diantaranya enzim ATP-ase maka kerusakan membran sel mengganggu fungsi enzim ATP-ase dalam produksi energi. Hambatan dalam produksi energi akan mengganggu pembentukan komponen sel bakteri.
Bakterisida botani yang sering digunakan untuk pengendalian penyakit tumbuhan adalah mimba (Azadzrachta zndzca Juss) dengan memanfaatkan bagian daun, daging buah, biji, kulit batang, dan akar. Tanaman mimba mengandung 70 senyawa terpenoid diantaranya azadirachtin, nimbin, dan salanin. Maharishi (1993)
inenggunakan ekstrak daun mimba 2 % dalam pelarut air untuk mengendalikan penyakit bercak daun cabai yang disebabkan oleh bakteri X a. pv. ve.szcutorzu. Daun inimba yang diemulsikan dalain etanol telah dikomersialkan dengan inerek dagang Margoside OK-80EC 0,05 % atau Margoside CK-20EC 0,I % untuk pengendalian penyakit bercak daun cabai.
Keefektifan ekstrak daun miinba segar konsentrasi 2 % dalam pelarut klorofonn saina dengan streptomisin 100 ppm dalam pengendalian penyakit bercak daun cabai. Pengendalian penyakit bercak daun dengan penggunaan biji mimba lebih efektif dibandingkan dengan daun karena bahan aktif yang terkandung dalain setiap g biji (setara 3,5 mg azadirachtin murni) lebih banyak dibandingkan dengan daun.
Heyne (1987) mengkoinpilasi berbagai jenis tanaman di Indonesia yang digunakan sebagai bakterisida penyakit manusia. Deskripsi beberapa bahan tanainan yang digunakan sebagai bakterisida pustul bakteri kedelai disajikan pada Tabel 2.
Walaupun penelitian lanjut belum dilakukan, namun senyawa fenolik yang terkandung dalain kulit ranting maupun daun matoa, daun kipahit, dan daun paku papila dilaporkan dapat menyembuhkan penyakit pada manusia yang disebabkan oleh bakteri, misalnya inata bernanah, kulit bernanah, bahkan gonorrlzoeu. Minyak atsiri yang terkandung dalain riinpang lengkuas merah maupun helai daun sereh wangi dilaporkan dapat inenyembuhkan penyakit kulit maupun diare pada manusia. Bahan- bahan tanaman tersebut telah digunakan secara turun temurun di Indonesia.
Tabel 2 Deskripsi enam bahan tanaman yang dideterminasi sebagai bakterisida
Nama tanaman Bagian tanaman
(Famili) yang digunakan Ekologi Senyawa aktif
untuk bakterisida A lpinia galanga S W (lengkuas merah) (Zingiberaceae) A ndropogon nardus var.genulnusHACK (sereh wangi) (Gramineae) Elaeis guineensis
J ACO (kelapa sawit)
(Palmae)
I,ygodium scandens
SWARTZ (paku ka- wat) (Schizaeceae) I'ierasma , javanica BL (kipait) (Simarubaceae) Ponletia pinnata FORST (matoa) (Sapindaceae)
Rimpang berwarna Rumpun memerlu- 250 g rimpang me-
merah kan cahaya matahari ngandung 0,15- 1,5%
minyak atsiri
Pelepah daun, helai Rumpun hidup di la- Daun mengandung
daun. han kritis, di bawah 77-83%minyak atsi-
naungan ri
Buah berisi biji Ke- Pohon di pantai Buah mengandung
ras 40% lemak kuning
Daun menyirip me- Rumpun hidup di ta- Senyawa fenolik
manjang nah lembab, di po-
hon
Daun dan kulit ba- Rumpun hidup di Senyawa fenolik
tang yang tua pinggir sungai
Daun dan ranting Pohon di daerah Senyawa fenolik
100-1400 m dpl