YUSMA YENNIE
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis Isolasi dan Identifikasi V.
parahaemolyticus patogenik pada Udang Tambak adalah karya saya dengan
arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam daftar pustaka di bagian akhir tesis ini.
Bogor, Agustus 2011
Yusma Yennie
NIM F251080201
ABSTRACT
YUSMA YENNIE. Isolation and Identification of Pathogenic Vibrio
parahaemolyticus in Shrimp Culture Under the direction of RATIH
DEWANTI-HARIYADI and ACHMAD POERNOMO.
Vibrio parahaemolyticus (Vp) is a halophilic bacterium found in brackish
water and is the leading cause of gastroenteritis due to seafood consumption. In Indonesia presence of this pathogen in seafood has caused several export rejection. This research aimed to identify presence of pathogenic Vibrio
parahaemolyticus from shrimp cultured in traditional and intensive ponds.
Bacterial isolation was carried out using FDA BAM (2004), phenotypic characterization was done using API 20E biochemical test kit and genetic characterization was conducted with Polymerase Chain Reaction (PCR) using a pair of specific primer for each virulent factor gene (tdh and trh genes). Biochemical identification with API 20E biochemical test showed that 16/32(50%) and 6/32 (18.8%) shrimp samples from traditional and intensive ponds contained Vibrio parahaemolyticus, respectively. Eighty one percent (13/16) of Vibrio parahaemolyticus isolated from traditional pond and 50% (3/6) of those obtained from intensive pond were pathogenic due to their possesion of
tdh gene. When gen encoding trh was used as the basis for classification, 15/16
(93.8%) and 4/6 ( 66.7%) of Vibrio parahaemolyticus obtained from traditional and intensive ponds, respectively, were pathogenic . Out of the 22 Vibrio
parahaemolyticus isolates, 16 (72.7%) were pathogenic based on the possesion of
gene encoding for tdh and 19 (86.4%) can be classified as pathogen based on the
trh gene. Overall, pathogenic Vibrio parahaemolyticus was found at a frequency
of 13-15/32 (43%) of the shrimp samples from traditional pond while 3-4/32 (11%) was found in shrimps from intensive pond, respectively.
RINGKASAN
YUSMA YENNIE. Isolasi dan Identifikasi Vibrio parahaemolyticus Patogenik
pada Udang Tambak. Dibimbing oleh RATIH DEWANTI-HARIYADI dan
ACHMAD POERNOMO.
Vibrio parahaemolyticus merupakan flora normal di lingkungan perairan
payau dan salah satu spesies Vibrio spp yang bersifat patogen terhadap komoditas udang maupun pada manusia. Vibrio parahaemolyticus merupakan bakteri Gram negatif, bersifat halofilik dan dapat menyebabkan gastroenteritis pada manusia melalui konsumsi pangan hasil perikanan. Udang merupakan salah satu komoditas hasil perikanan yang menduduki peringkat pertama dalam pangsa ekspor produk perikanan Indonesia serta merupakan salah satu komoditas unggulan revitalisasi KKP selain tuna dan rumput laut sejak 2005. Berbagai permasalahan yang terkait dengan ekspor udang mengakibatkan ditolaknya produk udang ke luar negeri dan hal ini mengakibatkan kerugian yang cukup besar bagi unit-unit pengolahan udang. Salah satu penyebab penolakan ekspor udang karena kandungan mikroba patogen seperti Salmonella, Vibrio cholerae dan Vibrio parahemolyticus.
Kasus penolakan ekspor udang karena kontaminasi V. parahemolyticus pernah dilaporkan yaitu pada tahun 2005 dan 2007, sebanyak 26 ton udang beku dan 4.8 ton produk sushi ebi dari Indonesia ditolak di Uni Eropa. Kasus terakhir, berdasarkan data Kementerian Kelautan dan Perikanan, tahun 2009 dan 2010 sebanyak 27 ton dan 13 ton ekspor ikan Indonesia ditolak oleh Cina karena terkontaminasi V. parahemolyticus. Dalam perdagangan internasional, beberapa negara seperti Uni Eropa, USA, dan Jepang menetapkan persyaratan mutu dan keamanan pangan terkait dengan V. parahaemolyticus pada produk perikanan termasuk udang baik beku maupun olahan. Indonesia juga di dalam SNI mensyaratkan V. parahaemolyticus sebagai parameter mutu pada produk perikanan.
Kasus penyakit bawaan pangan (foodborne diseases) karena V.
parahaemolyticus patogenik telah banyak dilaporkan dalam berbagai kasus KLB.
Di Indonesia ditemukan sebesar 3.7% (19/514 pasien) dengan gastroenteristis akut dan diketahui positif V. parahaemolyticus sepanjang tahun 1974 (Bonang et al. 1974). Selain itu ditemukan juga V. parahaemolyticus patogenik sebesar 7.3% dari sampel klinis pasien diare beberapa rumah sakit dalam kurun waktu 1995-2001 (Tjaniadi et al. 2003). Sementara itu di Jepang, dalam kurun waktu 1996-1998 terjadi 20-30% kasus KLB dan pada tahun 1996-1998 dilaporkan kasus ini melebihi jumlah kasus yang disebabkan oleh Salmonella (IDSC, 1999 dalam US-FDA, 2005).
Kemampuan V. parahaemolyticus menyebabkan penyakit pada manusia
umumnya dihubungkan dengan kemapuannya memproduksi hemolisis. Berdasarkan kemampuannya tersebut dikenal 3 jenis hemolisin pada V.
parahaemolyticus yaitu thermolabile hemolysin (TLH), thermostable direct hemolysin (TDH) dan thermostable direct hemolysin-related hemolysin (TRH). Thermolabile hemolysin (TLH) adalah protein yang memiliki aktivitas phospholipase A2/lysophospholipase akan tetapi peranan hemolisin ini terkait
dengan kemampuannya menyebabkan penyakit tidak diketahui secara pasti. Sementara itu TDH dan TRH diduga merupakan faktor virulen pada V.
parahaemolyticus yang dikaitkan sebagai penyebab penyakit pada manusia. Thermostable direct hemolysin (TDH) dikenal sebagai faktor virulen karena
aktivitas β hemolisisnya yang dapat melisis membran sel darah merah sehingga mengakibatkan gastroenteritis. Keberadaan TDH ditandai dengan adanya zona bening pada koloni Vibrio parahaemolyticus yang ditumbuhkan pada media agar Wagatsuma dan dikenal dengan istilah Fenomena Kanagawa (KP+).
Thermostable direct hemolysin-related hemolysin (TRH) disebut sebagai faktor
virulen lain dari V. parahaemolyticus, dimana keberadaannya dikaitkan dengan hasil Fenomena Kanagawa negatif (KP-) tetapi dapat menyebabkan gastroenteritis.
Identifikasi V. parahaemolyticus patogenik pada awalnya dilakukan
berdasarkan reaksi biokimiawi akan tetapi metode ini memiliki kelemahan diantaranya waktu analisis yang panjang, akurasi dan sensitivitas yang rendah serta belum tersedianya metode analisis untuk mengidentifikasi faktor virulen TRH. Berlandaskan alasan tersebut, dikembangkan metode analisis untuk mengidentifikasi V. parahaemolyticus patogenik. Salah satu metode analisis yang adalah metode analisis berdasarkan pendekatan molekuler yaitu teknik
Polymerase Chain Reaction (PCR).
Penelitian ini secara umum bertujuan untuk mengetahui frekuensi
V. parahaemolyticus patogenik pada komoditas udang di tambak tradisional dan
intensif yaitu dengan cara mengisolasi dan mengidentifikasi V. parahaemolyticus berdasarkan sifat-sifat biokimiawinya serta mengidentifikasi faktor virulen V.
parahaemolyticus berdasarkan amplifikasi gen penyandi tdh dan trh pada
komoditas udang tambak tersebut. Isolasi bakteri dilakukan dengan mengacu pada metode BAM-FDA (2004), konfirmasi V. parahaemolyticus berdasarkan reaksi biokimiawi menggunakan API 20E biochemical test kit, danidentifikasi V.
parahaemolyticus patogenik dilakukan berdasarkan amplifikasi gen penyandi tdh
dan trh menggunakan sepasang primer spesifik dari dua gen tersebut dengan teknik PCR
Isolasi V. parahaemolyticus pada sampel udang di tambak tradisional dan
intensif berturut-turut menunjukkan hasil sebesar 32 dan 28 isolat (n=32). Sementara itu konfirmasi V. parahaemolyticus menggunakan API 20E
biochemical test kit menunjukkan hasil sebanyak 16/32 (50%) dan 6/32 (18.8%)
sampel udang di tambak tradisional dan intensif positif sebagai V.
parahaemolyticus. Hasil ini menunjukkan bahwa frekuensi isolasi V. parahaemolyticus sampel udang di tambak tradisional lebih tinggi dibandingkan
tambak intensif. Jika dibedakan berdasarkan lokasi pengambilan sampel pada tambak tradisional, frekuensi isolasi V. parahaemolyticus berkisar antara 30-69%. Identifikasi V. parahaemolyticus patogenik berdasarkan amplifikasi gen
penyandi tdh pada sampel udang tambak tradisional dan intensif berturut-turut menunjukkan hasil sebesar 13/16 isolat dan 3/6 isolat positif gen tdh. Amplifikasi berdasarkan gen penyandi trh pada sampel udang di tambak tradisional dan intensif diperoleh hasil sebanyak 15/16 isolat dan 4/6 isolat adalah positif gen trh. Secara keseluruhan, hasil penelitian ini menunjukkan bahwa V. parahaemolyticus dari sampel udang tambak tradisional dan intensif ternyata berifat patogen sebesar 43% dan 11%.
Hasil penelitian menunjukkan bahwa terdapat V. parahaemolyticus patogenik pada komoditas udang baik di tambak tradisional maupun intensif. Hal ini diduga terkait dengan penerapan cara berbudidaya yang baik (Good
Aquaculture Practices) yang belum optimal terutama di tambak tradisional.
© Hak cipta milik IPB, tahun 2011
Hak cipta dilindungi undang-undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
Dilarang mengumumkan dan memperbanyak sebagian atau seluruh Karya tulis dalam bentuk apa pun tanpa izin IPB
ISOLASI DAN IDENTIFIKASI
Vibrio parahaemolyticus PATOGENIK PADA
UDANG TAMBAK
YUSMA YENNIE
Tesissebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Ilmu Pangan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2011
Judul Tesis : Isolasi dan Identifikasi V. parahaemolyticus patogenik pada Udang Tambak
Nama : Yusma Yennie
NRP : F251080201
Menyetujui, Komisi Pembimbing
Dr. Ratih Dewanti-Hariyadi, M.Sc Dr. Achmad Poernomo Ketua Anggota
Mengetahui,
Ketua Program Studi Ilmu Pangan Dekan Sekolah Pascasarjana IPB
Dr.Ratih Dewanti-Hariyadi, M.Sc Dr. Ir. Dahrul Syah, M.Sc.Agr
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT karena atas rahmat dan ridho-Nya, karya ilmiah sebagai hasil tugas akhir dapat diselesaikan. Penulis mengangkat tema keamanan pangan dengan judul penelitian Isolasi dan Identifikasi
V. parahaemolyticus patogenik pada Udang Tambak.
Pada kesempatan ini penulis mengucapkan terima kasih yang sebesar-besarnya kepada :
1. Dr. Ratih Dewanti-Hariyadi, M.Sc dan Dr. Achmad Poernomo, M.AppSc selaku pembimbing yang telah memberikan arahan, masukan, saran, dan semangat selama penelitian dan penulisan karya ilmiah ini.
2. Dr. Ir. Sri Hendrastuti Hidayat, M.Sc selaku dosen penguji atas saran dan masukan yang diberikan sebagai bentuk lain dari pembimbingan menuju kesempurnaan karya ilmiah ini.
3. Balai Besar Riset Pengolahan Produk dan Bioteknologi Kelautan dan Perikanan-Balitbang KP-KKP, yang telah mendanai penelitian ini.
4. Rekan-rekan di Kelti Keamanan Pangan, Lab. Mikrobiologi (Ayu dan Radest) dan Lab. Bioteknologi (Gintung dan mbak Dewi) BBRP2B atas bantuan, dan dorongan semangat selama penelitian.
5. Teman-teman lingkup BBRP2B (Ema, Ida, Devi, Diah, Yanti) dan teman-teman IPN 2008, atas persahabatan dan dorongan semangatnya selama penelitian dan penulisan karya ilmiah ini.
6. Suami dan anak-anak tercinta, abah, ibu, dan seluruh keluarga besar atas doa, kasih sayang, toleransi, pengertian, dorongan semangat, dan bantuannya. Semua pihak yang tidak dapat disebutkan satu persatu.
Penulis berharap semoga karya ilmiah ini dapat memberikan manfaat.
Bogor, Agustus 2011
RIWAYAT HIDUP
Penulis dilahirkan di Medan pada tanggal 24 Pebruari 1974 dari Ayah Ahmad Yusmadi Yusuf dan Ibu Ainal Mardiah yang merupakan anak ketiga dari lima bersaudara.
Tahun 1998 penulis menyelesaikan pendidikan S-1 pada Program Studi Teknologi Hasil Perikanan Institut Pertanian Bogor. Penulis melanjutkan studi ke program pasca sarjana pada tahun 2008 di program studi ilmu pangan Fakultas Teknologi Pertanian Institut Pertanian Bogor melalui program beasiswa Kementerian Kelautan dan Perikanan.
Penulis bekerja sebagai Pegawai Negeri Sipil sejak tahun 1999 dan ditempatkan di UPT-LPPMHP, Dinas Kelautan dan Perikanan Propinsi Sumatera Utara. Tahun 2001 sampai sekarang, penulis bekerja sebagai peneliti di Balai Besar Riset Pengolahan Produk dan Bioteknologi Kelautan dan Perikanan-Balitbang KP-KKP dan bergabung dengan kelompok peneliti bidang keamanan pangan dan lingkungan Kelautan dan Perikanan. Peneliti telah melakukan beberapa penelitian di bidang keamanan pangan antara lain identifikasi logam berat pada produk perikanan, identifikasi residu bahan berbahaya pada produk perikanan baik segar dan olahan, dan saat ini sedang melakukan penelitian dengan topik identifikasi V. parahaemolyticus patogenik pada komoditas udang vaname di unit pengolahan udang, pengumpul, dan retail.
DAFTAR ISI
Halaman
DAFTAR TABEL...xv
DAFTAR GAMBAR ...xvi
DAFTAR LAMPIRAN ... xvii
PENDAHULUAN Latar Belakang ...1 Perumusan Masalah ...6 Tujuan Penelitian ...6 Hipotesis ...7 TINJAUAN PUSTAKA Karakteristik Vibrio parahaemolyticus...8
Faktor Lingkungan yang Mempengarui Keberadaan Vibrio parahaemolyticus ...10
Penyakit Bawaan Pangan (food borne diseases) oleh Vibrio parahaemolyticus ...11
Faktor Virulen Vibrio parahaemolyticus ...16
Identifikasi Vibrio parahaemolyticus Patogenik ...19
METODE PENELITIAN Tempat dan Waktu Penelitian ...23
Bahan dan Alat...23
Pelaksanaan Penelitian ...24
Survei Lapang ...25
Pengambilan dan Preparasi Sampel Udang Tambak ...26
Isolasi Vibrio parahaemolyticus dari Udang Tambak ...26
Identifikasi Vibrio parahaemolyticus pada Udang Tambak ...27
Uji biokimia pendahuluan (Presumtif)...27
1. Pewarnaan Gram...27
2. Uji motilitas ...27
3. Uji oksidase...28
Uji biokimia lanjutan (Konfirmasi) ... 28
Identifikasi V. parahaemolyticus Patogenik dari Isolat yang berasal dari Udang Tambak ... 29
1. Isolasi DNA genom bakteri ... 29
2. Amplifikasi gen tdh dan trh... 31
HASIL DAN PEMBAHASAN Survei Lapang ... 33
Isolasi V. parahaemolyticus dari Udang Tambak ... 38
Identifikasi V. parahaemolyticus pada Udang Tambak ... 40
Identifikasi V. parahaemolyticus Patogenik dari Isolat yang Berasal dari Udang Tambak ... 51
1. Isolasi DNA genom bakteri ... 52
2. Amplifikasi Gen tdh dan trh V. parahaemolyticus ... 52
Rekomendasi untuk Perbaikan Usaha Budidaya Udang ... 59
SIMPULAN DAN SARAN ... 62
Simpulan... 62
Saran ... 63
DAFTAR PUSTAKA ... 64
DAFTAR TABEL
Halaman
1. Volume ekspor hasil perikanan menurut komoditas utama
(2005-2009)... 1
2. Impor produk perikanan yang terkontaminasi V. parahaemolyticus di Taiwan... 2
3. Persyaratan V. parahaemolyticus pada produk perikanan di berbagai negara... 3
4. Hubungan antara antigen tipe O dan K pada V. parahaemolyticus ... 9
5. KasusKLB karena infeksi V. parahaemolyticus di Asia... 13
6. KasusKLB karena infeksi V. parahaemolyticus di Eropa dan Amerika... 14
7. Primer oligonukleotida yang digunakan untuk deteksi gen tdh dan trh ... 24
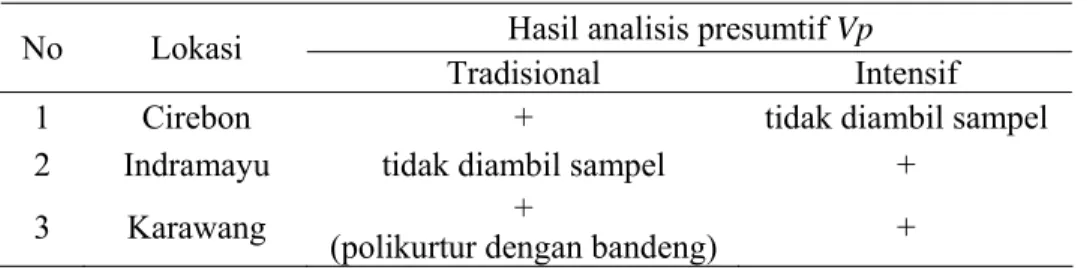
8. Hasil analisis presumtif V. parahaemolyticus dari udang vaname yang diambil dari 3 lokasi survei ... 34
9. Data pengamatan dan pengukuran parameter lingkungan di tambak tradisional dan intensif ... 35
10. Hasil isolasi V. parahaemolyticus pada sampel udang tambak ... 39
11. Identifikasi V. parahaemolyticus presumtif pada sampel udang tambak tradisional di Kecamatan Cantigi ... 41
12. Identifikasi V. parahaemolyticus presumtif pada sampel udang tambak intensif di Kecamatan Patrol ... 42
13. Hasil uji reaksi biokimiawi dari isolat-isolat V. parahaemolyticus presumtif sampel udang tambak dengan API 20E ... 44
14. Hasil identifikasi dan tingkat kemiripan isolat V. parahaemolyticus sampel udang di tambak tradisional dengan API 20E ... 45
15. Hasil identifikasi Vp pada sampel udang berdasarkan lokasi di tambak tradisional... 46
16. Hasil identifikasi dan tingkat kemiripan isolat V. parahaemolyticus sampel udang di tambak intensif dengan API 20E ... 48
17. Distribusi gen penyadi tdh dan trh pada isolat V. parahaemolyticus dari sampel udang di tambak tradisional dan intensif... 56
DAFTAR GAMBAR
Halaman
1. Sel Vibrio parahaemolyticus menggunakan scanning
electron micrograph (SEM)... 8
2. Diagram alir pelaksanaan penelitian ... 25
3. Tambak udang sistim tradisional... 36
4. Tambak udang sistim intensif... 37
5. V. parahaemolyticus pada TCBS ... 39
6. Sel bakteri V. parahaemolyticus yang diamati dengan mikroskop perbesaran 1000X... 43
7. Visualisasi hasil amplifikasi DNA dengan target gen penyandi tdh bakteri V. parahaemolyticus di tambak tradisional pada gel agarosa2% (TBE1X). M: marker DNA ladder 100bp, 1 : kontrol positif (AQ4037), 2: kontrol negatif, (a) dan (b) : 16 isolat V. parahaemolyticus sampel udang (lajur 3-10) ... 53
8. Visualisasi hasil amplifikasi DNA dengan target gen penyandi tdh bakteri V. parahaemolyticus di tambak intensif pada gel agarosa 2% (TBE1X). M : marker DNA ladder 100bp, 1 : kontrol positif (AQ4037), 2 : kontrol negatif, lajur 3-8 : isolat V. parahaemolyticus sampel udang... 54
9. Visualisasi hasil amplifikasi DNA dengan target gen penyandi trh bakteri V. parahaemolyticus di tambak tradisional pada gel agarosa2% (TBE1X). M: marker DNA ladder 100bp, 1 : kontrol positif (AQ4037), 2: kontrol negatif, (a) dan (b) : 16 isolat V. parahaemolyticus sampel udang (lajur 3-10) ... 54
10. Visualisasi hasil amplifikasi DNA dengan target gen penyandi trh bakteri V. parahaemolyticus di tambak intensif pada gel agarosa 2% (TBE1X). M : marker DNA ladder 100bp, 1 : kontrol positif (AQ4037), 2 : kontrol negatif, lajur 3-8 : isolat V. parahaemolyticus sampel udang... 55
DAFTAR LAMPIRAN
Halaman
1. Hasil isolasi V. parahaemolyticus dari udang tambak tradisional ... 76
2. Hasil isolasi V. parahaemolyticus dari udang tambak intensif ... 77
3. Hasil identifikasi isolat presumtif V. parahemolyticus dengan API 20E yang dianalisis menggunakan apiwebTM ... 78
PENDAHULUAN
Latar Belakang
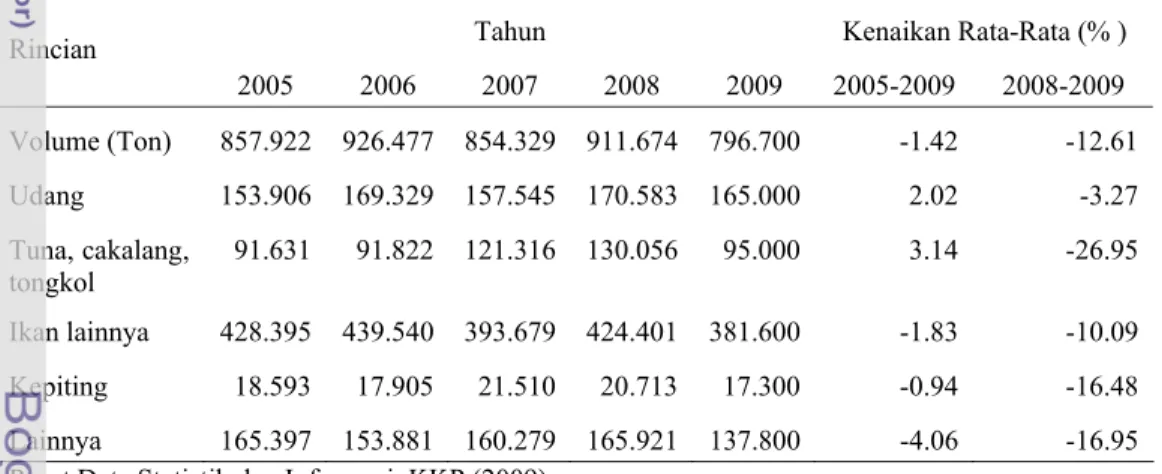
Udang merupakan salah satu komoditas unggulan program revitalisasi Kementerian Kelautan dan Perikanan (KKP) selain tuna dan rumput laut sejak tahun 2005. Disamping itu udang menempati urutan ke-5 terbesar dalam deretan ekspor non-migas dan memberikan kontribusi sebesar 50% dari total nilai ekspor perikanan Indonesia. Berdasarkan data yang dikeluarkan oleh Pusat Data Statistik dan Informasi, KKP (2009), tentang volume ekspor hasil perikanan menurut komoditas utama tahun 2005-2009, terlihat udang menduduki urutan pertama (Tabel 1).
Tabel 1. Volume ekspor hasil perikanan menurut komoditas utama (2005-2009)
Tahun Kenaikan Rata-Rata (% ) Rincian 2005 2006 2007 2008 2009 2005-2009 2008-2009 Volume (Ton) 857.922 926.477 854.329 911.674 796.700 -1.42 -12.61 Udang 153.906 169.329 157.545 170.583 165.000 2.02 -3.27 Tuna, cakalang, tongkol 91.631 91.822 121.316 130.056 95.000 3.14 -26.95 Ikan lainnya 428.395 439.540 393.679 424.401 381.600 -1.83 -10.09 Kepiting 18.593 17.905 21.510 20.713 17.300 -0.94 -16.48 Lainnya 165.397 153.881 160.279 165.921 137.800 -4.06 -16.95 Pusat Data Statistik dan Informasi, KKP (2009)
Ekspor komoditas udang Indonesia mengalami masalah beberapa tahun ini. Permasalahan ekspor udang Indonesia mengakibatkan volume dan nilai ekspor menurun dan beberapa permasalahan yang dihadapi terkait dengan standar mutu dan sanitasi. Permasalahan yang terkait dengan sanitasi pada komoditas udang umumnya karena adanya kontaminasi bakteri patogen seperti Salmonella,
Vibrio parahaemolyticus, dan Vibrio cholera (DKP, 2003). Pada tahun 2005
sebanyak 26 ton ekspor udang Indonesia ditolak Uni Eropa karena kontaminasi V.
parahaemolyticus, sedangkan pada tahun 2007 ekspor produk sushi ebi sebanyak
4.8 ton ditolak oleh Uni Eropa karena alasan yang sama (Ditjen P2HP-KKP, 2010). Kasus terakhir, berdasarkan data Kementerian Kelautan dan Perikanan, pada tahun 2009 dan 2010 sebanyak 27 ton dan 13 ton ekspor ikan Indonesia ditolak oleh Cina karena terkontaminasi V. parahemolyticus. Wong et al. (1999) melaporkan bahwa produk perikanan yang diekspor ke Taiwan dari beberapa negara di Asia termasuk Indonesia pernah terdeteksi mengandung V.
parahaemolyticus (Tabel 2) walaupun pada seluruh sampel tidak ditemukan V. parahaemolyticus patogenik yang diidentifikasi dengan metode PCR.
Tabel 2. Impor produk perikanan yang terkontaminasi V. parahaemolyticus di Taiwan
Negara asal Jenis produk Jumlah sampel Jumlah sampel positif % Vp Kepiting 126 41 32.5 Lobster 59 26 44.1 Bekicot 95 20 21.1 Vietnam Ikan 106 47 44.3 Hongkong Kepiting 114 81 71.1 Thailand Kepiting 32 26 81.3 Udang 62 47 75.8 Indonesia Ikan 92 27 29.3 Jumlah 686 315 45.9 Wong et al. (1999)
Dalam perdagangan internasional, beberapa negara seperti Uni Eropa, USA, dan Jepang menetapkan persyaratan mutu dan keamanan pangan terkait dengan V. parahaemolyticus pada produk perikanan termasuk udang baik beku maupun olahan. Indonesia juga di dalam Standar Nasional Indonesia (SNI) mensyaratkan V. parahaemolyticus sebagai parameter mutu pada produk perikanan. Tabel 3 menunjukkan peraturan dari beberapa negara pengimpor
untuk persyaratan mutu dan keamanan pangan produk udang serta persyaratan udang beku berdasarkan SNI 01-2705.1-2006.
Tabel 3. Persyaratan V. parahaemolyticus pada produk perikanan di
berbagai negara
Negara Jenis pangan Persyaratan Referensi
Uni Eropa Krustasea, moluska,
dan kerang olahan MPN ≤100 cfu/g
ISO 8914 USA Produk perikanan
siap saji
MPN >104/g (KP + / -) FDA –Compliance
Programme Guidance Manual 7303.844 Jepang Produk perikanan untuk
konsumsi mentah
MPN < 100/g Food Sanitation & Quarantine Law
Article 11 Indonesia Udang beku APM <3/g atau KP + *
(* jika diperlukan)
SNI 01-2705.1-2006
Ababouch et al. (2005); Badan Standardisasi Nasional (2006)
Vibrio spp. merupakan flora normal pada lingkungan perairan payau
seperti pantai atau muara sungai serta umum terdapat selama kegiatan budidaya udang (Vandenberghe et al. 2003). Keberadaan Vibrio spp. terutama berkaitan dengan bahan organik dan fluktuasi oksigen terlarut pada lahan budidaya. Selain itu dalam kondisi normal peningkatan suhu akan menimbulkan keragaman spesies
Vibrio (Barbieri et al. 1999; Pfeffer et al. 2003). Beberapa spesies patogen Vibrio
seperti V. harveyi dan V. parahaemolyticus merupakan bakteri yang menginfeksi udang (Jiravanichpaisal dan Miyazaki, 1995; Lavilla-Pitogo, 1995; Lightner, 1993) dan umumnya disebut dengan patogen oportunistik yang dapat menyebabkan penyakit pada udang (Goarant et al. 1999). Infeksi V.
parahaemolyticus pada udang terjadi pada fase juvenil sampai dewasa. Penyakit
pada udang ini disebut dengan red disease syndrome yaitu berubahnya warna tubuh udang menjadi merah dan mengakibatkan kematian. Kematian udang karena penyakit ini berkisar 1-20% (Alapide-Tendencia dan Dureza, 1997)
Sistim pemeliharaan udang di tambak umumnya dibedakan atas sistim tradisional dan intensif. Kedua sistim tambak ini memiliki beberapa perbedaan antara lain sumber air, pengelolaan kualitas air, padat tebar benur udang, konstruksi lahan tambak, sistim pengairan, dan jenis pakan. Tambak tradisional
umumnya tidak menerapkan manajemen pengelolaan kualitas air tambak, dimana sumber air untuk pemeliharaan umumnya berasal dari aliran sungai yang berada di sekitar tambak. Hal ini memberi peluang besarnya kandungan kontaminan dari sumber air seperti logam berat dan bakteri patogen yang dapat menyebabkan tingkat kematian pada udang cukup tinggi. Selain itu tambak tradisional tidak dilengkapi oleh sistim aerasi yang berfungsi mengatur ketersediaan oksigen (Komarawidjaja dan Garno, 2003).
Disamping merupakan patogen pada udang, beberapa spesies Vibrio juga bersifat patogen pada manusia. Lebih dari 12 spesies Vibrio diketahui terkait dengan penyakit pada manusia, dan spesies V. cholerae dan V. parahaemolyticus merupakan patogen yang dominan penyebab penyakit pada manusia (Kaysner dan DePaola, 2004). V. cholerae merupakan spesies patogen Vibrio yang memiliki lebih dari 200 serotipe, akan tetapi hanya serotipe O1 dan O139 yang bersifat patogen dan menyebabkan penyakit pada manusia. Sementara itu serotipe lainnya disebut dengan non O1/O139 dan jarang menginfeksi manusia (Kaper et al. 1995; Anderson et al. 2004). V. cholerae memproduksi enterotoksin kolera yang menyebabkan penyakit kolera pada manusia. Penyakit kolera ditularkan melalui jalur fekal-oral, melalui air yang terkontaminasi saat pencucian bahan pangan ataupun bahan pangan yang terkontaminasi feses manusia yang biasa digunakan untuk pupuk (Dobosh et al. 1995; Kaysner, 2000; Popovic et al. 1993). Galur O1 umumnya diisolasi dari sampel klinis sedangkan galur non O1/O139 diisolasi dari lingkungan perairan dan produk perikanan.
Spesies Vibrio lain yang patogen terhadap manusia adalah V.
parahaemolyicus, merupakan bakteri Gram negatif yang umumnya terdeteksi
pada air, sedimen, plankton, produk perikanan (krustasea, ikan dan moluska). Hal ini karena bahan-bahan tersebut memiliki kondisi optimum bagi pertumbuhan bakteri ini seperti ketersediaan nutrien, kandungan garam, pH dan Aw.
Di Indonesia, penelitian keberadaan V. parahaemolyticus pada produk perikanan termasuk udang masih jarang dilakukan. Dewanti-Hariyadi et al. (2002) melaporkan bahwa V. parahaemolyticus yang diisolasi dari sampel udang yang berasal dari tambak di Jawa Barat dan Jawa Tengah, pasar grosir dan unit pengolahan berturut-turut sebesar 21.8%, 3.1%, 11.1%, dan 70%. Namun
demikian tidak diketahui apakah V. parahaemolyticus pada sampel udang tersebut bersifat patogenik. Penelitian lain juga melaporkan bahwa ditemukan V.
parahaemolyticus pada seluruh sampel kerang mentah dan olahan (n=47) yang
berasal dari perairan dan pasar lokal di Padang-Sumatera Barat, dimana 36% dari isolat tersebut merupakan V. parahaemolyticus patogenik yang diidentifikasi
berdasarkan gen penyandi tdh (thermostable direct hemolysin) (Marlina et al. 2007).
Jika dikaitkan dengan kesehatan masyarakat, V. parahaemolyticus juga pernah diisolasi dari sampel klinis pasien diare di beberapa rumah sakit di Indonesia dan diketahui sebesar 7.3% (n=2812) merupakan V. parahaemolyticus dengan Fenomena Kanagawa (KP) positif (Tjaniadi et al. 2003). Persentase ini lebih kecil dibandingkan dengan kejadian keracunan oleh Salmonella akan tetapi lebih tinggi dari persentase kejadian yang disebabkan oleh Enterohemorrhagic
E. coli (EHEC).
Untuk mengetahui bahwa V. parahaemolyticus bersifat patogenik,
umumnya dilakukan pengujian Kanagawa yakni dengan mengamati pembentukan daerah bening pada agar Wagatsuma yang menandakan adanya hemolisin. V.
parahaemolyticus yang menghasilkan Kanagawa positif (KP+) adalah galur yang
menghasilkan faktor virulen thermostable direct hemolysin (TDH) yang disebut dengan gen tdh. Meskipun demikian tidak semua V. parahaemolyticus patogenik ditandai dengan hasil uji Kanagawa positif. Hal ini ditemukan pada pasien penderita diare yang tidak menunjukkan hasil KP+. Faktor virulen ini kemudian dikenal dengan thermostable direct hemolysin related hemolysin (TRH) dan disebut gen trh. Analisis berdasarkan reaksi biokimiawi ini ternyata memiliki beberapa kelemahan sehingga dikembangkan metode analisis untuk mengidentifikasi V. parahaemolyticus patogenik menggunakan pendekatan molekuler seperti metode polymerase chain reaction (PCR). Metode ini merupakan metode enzimatis untuk melipatgandakan secara eksponensial suatu sekuen nukleotida tertentu dengan cara in vitro. Metode PCR banyak dikembangkan untuk pengujian mikrobiologi karena memiliki keunggulan diantaranya sensitifitas tinggi, ketepatan hasil uji tinggi, waktu pengujian relatif cepat dan dapat digunakan untuk pengujian komponen yang jumlahnya sangat
sedikit (Yuwono, 2006). Identifikasi V. parahaemolyticus patogenik menggunakan metode PCR dilakukan dengan cara mengamplifikasi sekuen nukleotida berdasarkan keberadaan gen penyandi tdh dan trh.
Perumusan Masalah
Salah satu permasalahan penolakan ekspor udang Indonesia adalah
kontaminasi bakteri patogen diantaranya Vibrio parahaemolyticus. V. parahaemolyticus merupakan patogen oportunistik terhadap udang dan juga
merupakan bakteri patogen penyebab penyakit pada manusia. Akan tetapi ketersediaan data keberadaan V. parahaemolyticus patogenik pada komoditas udang tambak masih sangat terbatas.
Identifikasi V. parahaemolyticus patogenik komoditas udang tambak di
Indonesia masih jarang dilakukan. Metode identifikasi V. parahaemolyticus patogenik dengan metode konvensional berdasarkan reaksi biokimia telah banyak dilakukan akan tetapi memiliki kelemahan seperti waktu analisis yang lama, ketepatan hasil uji dan sensitifitas yang rendah. Selain itu metode konvensional tidak dapat mengidentifikasi keberadaan TRH pada sampel.
Metode PCR merupakan salah satu pengembangan metode identifkasi V.
parahaemolyticus patogenik dengan pendekatan molekular yang telah banyak
dikembangkan karena dapat menghasilkan akurasi dan ketepatan hasil uji yang lebih tinggi. Identifikasi V. parahaemolyticus patogenik pada udang tambak dengan metode PCR diharapkan dapat memperoleh hasil uji yang akurat.
Tujuan penelitian
Tujuan umum penelitian ini adalah mengevaluasi frekuensi isolasi V.
parahaemolyticus patogenik pada komoditas udang tambak tradisional dan
intensif. Sedangkan tujuan khusus penelitian ini adalah :
a. Mengisolasi dan mengidentifikasi V. parahaemolyticus secara biokimiawi pada komoditas udang yang berasal dari tambak tradisional dan intensif serta manganalisis faktor yang berkontribusi terhadap keberadaan bakteri ini. b. Mengidentifikasi V. parahaemolyticus patogenik pada komoditas udang
Hipotesis
Hipotesis pada penelitian ini adalah :
a. V. parahaemolyticus berpotensi ditemukan pada komoditsas udang tambak baik tambak tradisional maupun intensif, dengan frekuensi isolasi yang lebih tinggi terdapat pada tambak tradisional.
b. V. parahaemolyticus patogenik pada sampel lingkungan dan produk perikanan umumnya berkisar 1-2%.
c.
TINJAUAN PUSTAKA
Karakteristik Vibrio parahaemolyticus
Vibrio parahaemolyticus adalah salah satu spesies bakteri dari famili Vibrionaceae yang merupakan bakteri Gram negatif berbentuk batang (curved
atau straight ), anaerob fakultatif, tidak membentuk spora, pleomorfik, bersifat motil dengan single polar flagellum. Bakteri ini merupakan bakteri halofilik (tumbuh optimum pada media yang berkadar garam 3%), tidak memfermentasi sukrosa dan laktosa, dapat tumbuh pada suhu 10-44oC (optimum suhu 37oC), dimana waktu generasi bakteri pada fase eksponensial adalah 9-13 menit di kondisi optimum pertumbuhannya. Sementara itu kisaran pH dan Aw pertumbuhannya berturut-turut adalah 4.8 – 11 (optimum 7.8 – 8.6) dan 0.94 –
0.99 (optimum 0.981) (Baumann dan Schubert, 1984; Jay et al. 2005; Lake et al. 2003). Beberapa karakter Vibrio parahaemolyticus yang membedakannya dengan spesies Vibrio lainnya diantaranya tidak memfermentasi sukrosa seperti Vibrio cholerae dan Vibrio alginolyticus. Selain itu pada media padat bakteri ini tumbuh dengan menggunakan lateral flagella serta sifatnya yang halofilik dengan kisaran garam 0.5-8%, sedangkan bakteri Vibrio cholerae mampu tumbuh pada media tanpa garam (Holt dan Krieg, 1984).
Gambar 1. Sel V. parahaemolyticus menggunakan scanning electron micrograph (SEM), bar = 1µm
Berdasarkan antigennya, Vibrio parahaemolyticus terdiri atas kelompok antigen yaitu : tipe H (flagellar), tipe O (somatic) dan tipe K (capsular). Antigen tipe H merupakan antigen paling umum untuk seluruh galur Vibrio
parahaemolyticus, sedangkan antigen tipe O bersifat thermolabile dan antigen
tipe K bersifat thermostable. Saat ini, terdapat 12 grup antigen O dan lebih dari 70 antigen K yang telah menghasilkan 76 serotipe (Tabel 4), dimana 5 dari antigen K dengan 2 antigen O membentuk serotipe O:K, yang kemudian digunakan untuk investigasi penyebaran penyakit yang disebabkan oleh Vibrio
parahaemolyticus (Kaysner dan DePaola, 2004). Skema antigen ini pertama kali
dipublikasikan oleh Sakazaki et al. (1963), selanjutnya beberapa serotipe ditambahkan oleh Komite Serotipe Vibrio parahaemolyticus Jepang dimana antigen K : 2, 14, 16, 29, 35 dan 62 bukan merupakan hasil dari Komite. Antigen K : 4, 5, 6, 7, 8, 9, dan 19 bisa ditemukan lebih dari satu antigen O.
Tabel 4. Hubungan antara antigen tipe O dan K pada V. parahaemolyticus
Antigen O Antigen K 1 2 3 4 5 6 7 8 9 10 11 12 1,25,26,32,38,41,56,58,64,69 3,28 4,5,6,7,27,30,31,33,37,43,45,48,54,57,58,59,65 4,8,9,10,11,12,13,34,42,49,53,55,63,67 5,15,17,30,47,60,61,68 6,18,46 7,19 8,20,21,22,39,70 9,23,44 19,24,52,66,71 36,40,50,51,61 52
Kaysner dan De Paola (2004)
Vibrio parahaemolyticus merupakan flora normal di lingkungan
perairan payau seperti pantai, muara sungai atau tambak yang tersebar di seluruh dunia. Keberadaan bakteri ini umumnya lebih sering ditemui pada wilayah
beriklim sedang dan tropis atau pada musim panas di negara-negara empat musim.
V. parahaemolyticus umumnya terdeteksi pada air laut, sedimen, plankton, ikan,
krustasea, kekerangan dan moluska. Produk perikanan memberikan semua kondisi yang dibutuhkan oleh V. parahaemolyticus untuk tumbuh dan
berkembang biak seperti keberadaan garam, kandungan nutrien, pH dan Aw yang optimum dan faktor lainnya sehingga bakteri ini sering disebut flora normal pada produk perikanan.
Faktor Lingkungan yang Mempengaruhi Keberadaan V. parahaemolyticus
Bakteri patogen yang terkait dengan produk perikanan secara umum dikelompokkan atas 3 yaitu : bakteri yang merupakan flora normal perairan laut (Vibrio spp., Listeria monocytogenes, Clostridium botulinum dan Aeromonas
hydrophila), bakteri yang berasal dari kontaminasi feses (Salmonella spp., E. coli
patogenik, Shigella spp., Campylobacter spp., dan Yersinia enterocolitica), dan bakteri yang berasal dari kontaminasi selama pengolahan (Bacillus cereus,
Staphylococcus aureus, Listeria monocytogenes, dan Clostridium perfringens). V. parahaemolyticus merupakan flora normal yang hidup di perairan payau dan
sebagian jenisnya bersifat patogen pada produk perikanan seperti udang, ikan, kepiting, tiram, kerang, dan jenis cephalopoda. (Feldhusen, 2000; Liston, 1990; Liu dan Chen, 2004; Su dan Liu, 2007).
Keberadaan V. parahaemolyticus di lingkungan perairan dan produk
perikanan dipengaruhi oleh musim, lokasi, polutan, jenis sampel dan metode analisis (Cook et al. 2002; DePaola et al. 1990; DePaola et al. 2000; Kaneko dan Colwell, 1973; Kaysner et al. 1990; Watkins dan Cabelli, 1985) . Suhu perairan merupakan faktor penting yang mengontrol tingkat V. parahaemolyticus pada lingkungan, dimana terjadi peningkatan jumlah V. parahaemolyticus pada kisaran suhu 10-30°C (De Paola et al. 1990; Kaneko dan Colwell, 1973; Watkins dan Cabelli, 1985). Penelitian menunjukkan V. parahaemolyticus jarang ditemukan saat suhu perairan di bawah 10°C akan tetapi keberadaannya akan meningkat sejalan dengan meningkatnya suhu perairan. Studi ekologi lainnya menyebutkan bahwa V. parahaemolyticus dapat bertahan hidup pada biota perairan (plankton, kekerangan, kustasea, ikan) dan sedimen selama musim dingin dan akan terlepas ke perairan saat suhu meningkat pada awal musim panas (Kaneko dan Colwell, 1973). DePaola et al. (1990) melaporkan, hasil survei 9 pantai di USA dalam kurun waktu 1984-1985 menunjukkan densitas V. parahaemolyticus yang cukup rendah di perairan (4 koloni/ml) ketika terjadi penurunan suhu di bawah 16°C.
Namun demikian, densitas bakteri ini meningkat menjadi 103 koloni/ml saat suhu perairan mencapai 25°C. Pada budidaya tiram di Oregon juga menunjukkan adanya korelasi positif antara jumlah V. parahaemolyticus dengan peningkatan suhu dan pupolasi tertinggi terjadi pada musim panas (Duan dan Su, 2005).
Sementara itu keberadaan V. parahaemolyticus di produk perikanan diketahui lebih banyak teridentifikasi pada saat terjadi peningkatan suhu lingkungan. Keysner dan DePaola (2000) melaporkan jumlah V.
parahaemolyticus pada tiram yang dipanen pada musim semi dan panas lebih
banyak dibandingkan musim dingin yaitu di atas 103cfu/g dan dapat berkembang biak dengan cepat pada tiram yang terpapar suhu tinggi. Hasil penelitian Gooch
et al. (2002) menunjukkan bahwa populasi V. parahaemolyticus pada tiram
meningkat 50-790 kali lipat dari jumlah awal selama 24 jam setelah panen jika disimpan pada suhu 26°C. Sementara itu, hasil survei sampel tiram yang diambil dari restoran dan seluruh pasar produk hasil perikanan tingkat eceran sampai grosir di USA selama Juni 1998 - Juli 1999 menyimpulkan bahwa V.
parahaemolyticus memiliki kepadatan tertinggi (> 103MPN/g) pada musim panas
(Cook et al. 2002).
V. parahaemolyticus dapat dideteksi pada rentang salinitas yang cukup
besar (5-35 ppt) dengan salinitas optimal berkisar 22 ppt (DePaola et al. 1990). Beberapa penelitian menyebutkan bahwa terdapat korelasi yang tidak langsung antara polusi fekal dengan keberadaan V. parahaemolyticus karena diduga merupakan biostimulasi dari mikrofauna yang berasosiasi dengan V.
parahaemolyticus (Watkins dan Cabelli, 1985). Selain itu V. parahaemolyticus
diduga terkait erat dengan keberadaan zooplankton terutama copepoda yang dikaitkan dengan aktivitas dan afititas kitinase dari kitin (Fratamico et al. 2005; Kaneko dan Colwell, 1973; Watkins dan Cabelli, 1985).
Penyakit Bawaan Pangan (foodborne diseases) oleh Vibrio parahaemolyticus
Keberadaan V. parahaemolyticus patogenik pada produk perikanan dapat
menyebabkan penyakit pada manusia melalui konsumsi pangan (foodborne
diseases) terutama melalui pangan mentah atau yang tidak dimasak sempurna.
pangan olahan dan mentah atau melalui pencucian pangan dengan air yang mengandung V. parahaemolyticus. Penyakit karena V. parahaemolyticus adalah gastroenteristis seperti diare (98%), kejang bagian perut (82%), mual (71%), muntah (52%), dan demam (27%) dengan masa inkubasi 4-96 jam dengan rata-rata 15 jam (Barker dan Gangarosa, 1974; Levine et al. 1993) . Sebagian kecil kasus, bakteri ini menyebabkan kerusakan (luka) pada mukosa usus sehingga terdapat darah pada feses penderita bahkan dapat menyebabkan septisemia.
Penyakit bawaan pangan oleh V. parahaemolyticus umumnya lebih sering terjadi di negara beriklim tropis karena merupakan kondisi optimum pertumbuhan bakteri ini. Awalnya kasus V. parahaemolyticus terjadi secara sporadis akan tetapi menyebar dengan cepat ke seluruh dunia dan menjadi kasus kejadian luar biasa (KLB). Kasus infeksi karena V. parahaemolyticus melalui konsumsi pangan pertama sekali terjadi di Osaka-Jepang pada tahun 1951 akibat mengkonsumsi ikan sardine mentah. Kasus ini memakan korban sebanyak 272 orang menderita sakit dan 20 orang meninggal (Daniels et al. 2000). Tahun 1998 kasus infeksi karena V. parahaemolyticus meningkat dua kali lipat dibandingkan tahun 1997 dan melebihi jumlah kasus yang disebabkan oleh Salmonella. Pada kurun waktu 1996-1998 terjadi 20-30% kasus KLB (IDSC, 1999 dalam US-FDA, 2005). Negara Asia lainnya adalah Taiwan, sebanyak 57% (42/74 kasus KLB) adalah yang disebabkan oleh V. parahaemolyticus seperti yang dilaporkan Kementerian Kesehatan Taiwan tahun 1994. Kurun waktu 1986-1995 sebanyak 35% (197/555 kasus) merupakan kasus yang disebabkan oleh V. parahaemolyticus (Pan et al. 1996; Pan et al.1997). Sementara itu terjadi 5 kasus KLB di di Thailand, dimana 7-93 orang terinfeksi setelah mengkonsumsi kepiting dan ikan makarel olahan pada tahun 1971. Pada November 1970-Juni 1973, sebanyak 7930 sampel klinis penderita diare teridentifikasi V. parahaemolyticus (Phayakvichien et al. 1990). Di Vietnam, kasus KLB terdeteksi sebanyak 548 kasus pada tahun 1997-1999, dimana 90% terjadi pada usia di atas 5 tahun. Dalam kasus ini sebanyak 77% menderita muntah, diare (53%), dan diare berdarah (6%) (Tuyet et al. 2002). Indonesia sendiri pernah terjadi kasus sebesar 3.7% (19/514 pasien) dengan gastroenteristis akut dan diketahui positif V. parahaemolyticus sepanjang tahun
1974 (Bonang et al. 1974). Kasus KLB karena infeksi V. parahaemolyticus yang terjadi di negara-negara di Asia dapat dilihat pada Tabel 5.
Tabel 5. Kasus KLB karena infeksi V. parahaemolyticus di Asia
No Negara Insiden V. parahaemolyticus Referensi
1 Jepang - Kasus I tahun 1951 (272 orang sakit; 20 orang meningal
- 20-30% kasus KLB (1996-1998)
Daniels et al. (2000) US-FDA (2005)
2 Taiwan - 57% (42/72 kasus KLB) terjadi pada tahun 1994
- Tahun 1986-1995 sebesar 35% (197/ 555 kasus) adalah kasus Vp
Pan et al.(1996) Pan et al.(1997) 3 Thailand - Terjadi 5 kasus KLB karena konsumsi
kepiting dan makarel pada tahun 1971 - Tahun 1970-1973 terdapat Vp pada
7390 sampel klinis pasien diare
Phayakvichien et al. (1990)
4 Vietnam - Tahun 1997-1999, terjadi 548 kasus Tuyet et al. (2002)
5 Indonesia - Tahun 1974 ditemukan 3.7% (19/154 pasien) dengan gastroenteris akut karena Vp
Bonang et al. (1974)
Kasus KLB V. parahaemolyticus juga dilaporkan terjadi di negara Eropa
dan Amerika (Tabel 6) walaupun lebih jarang dibandingkan negara-negara di Asia. Robert-Pillot et al. (2004) menyebutkan bahwa kasus KLB serius pernah terjadi di Perancis pada tahun 1997 karena mengkonsumsi udang yang diimpor dari Asia dan memakan korban 44 orang. Amerika Serikat melaporkan terjadi 40 kasus KLB yaitu sepanjang 1973 – 1998 di 15 negara bagian dan wilayah Guam dengan 1064 penderita dan median tingkat serangan 56% (3 - 100%) dimana sebagian besar kasus terjadi di bulan Juli. Penyebabnya adalah tiram dan kerang mentah (38% kasus) atau yang tidak dimasak sempurna. Pada periode ini, 30% KLB terjadi pada 1997 – 1998 dan tiga diantaranya cukup besar yaitu Juli– Agustus 1997, keracunan disebabkan oleh konsumsi tiram mentah dari Puget Sound, Washington selanjutnya dua kasus KLB gastroenteritis karena V.
parahaemolyticus terjadi pada Mei–Juni 1998 akibat mengkonsumsi tiram mentah
yang berasal dari Galveston Bay, Texas dan di akhir Juli 1998, KLB V.
parahaemolyitcus terkait dengan konsumsi tiram dan kerang mentah yang berasal
Kasus KLB V. parahaemolyitcus di New York, Oregon dan Washington,
kembali terjadi pada 20 Mei – 31 Juli 2006 setelah mengkonsumsi tiram dan remis mentah dan olahan di restoran. Tiram dan remis berasal dari daerah pantai Washington dan British Columbia-Canada yang didistribusikan secara nasional ke pasar ikan dan restoran. Luasnya daerah pemasaran berdampak pada meluasnya daerah sebaran penyakit. Pada tahun 2006, 122 kasus berasal dari 17 sumber produk hasil perikanan yang sama dan berimplikasi pada penutupan perusahaan pemanenan tiram yang merupakan pemasok utama tiram penyebab KLB (Balter et
al, 2006). Selain itu kasus KLB juga dilaporkan di Chile pada November
1997-April 1998, dimana kasus ini terkait dengan konsumsi kerang. Hal ini diduga adanya pengaruh El Nino selain suhu perairan yang kemungkinan dapat membantu blooming bakteri. Spanyol juga menghadapi kasus KLB karena V.
parahaemolyticus antara Agustus-September 1999, dimana 64 orang menderita
sakit setelah mengkonsumsi tiram mentah yang berasal dari pasar lokal (Cordova
et al. 2002; Lozano-Leon et al. 2003).
Tabel 6. Kasus KLB karena infeksi V. parahaemolyticus di Eropa dan Amerika
No Negara Insiden V. parahaemolyticus Referensi
1 Perancis - Tahun 1997 karena konsumsi udang yang diimpor dari Asia
Robert-Pillot et al. (2004)
2 Amerika Serikat
- Tahun 1973 – 1998, 40 kasus KLB di 15 negara bagian dan wilayah Guam dengan 1064 penderita
- Mei-Juli 2006, terjadi kasusKLB setelah mengkonsumsi tiram dan remis mentah dan olahan
Daniels et al. (2000)
Balter et al. (2006)
3 Chile - November 1997-April 1998, terjadi kasus KLB terkait dengan konsumsi kerang
Cordova et al.
(2002)
4 Spanyol - Bulan Agustus-September 1999, sebanyak 64 orang terinfeksi Vp
Lozano-Leon et al. (2003)
Penyakit bawaan pangan karena V. parahaemolyitcus sangat terkait dengan cara mengkonsumsi bahan pangan tersebut. Kasus V. parahaemolyitcus yang terjadi di Taiwan disebabkan oleh kebiasaan masyarakatnya mengkonsumsi produk perikanan dalam kondisi mentah. Kondisi yang sama tampaknya juga
terjadi di beberapa negara Asia lainnya yang mempunyai kebiasaan mengkonsumsi produk perikanan dalam kondisi mentah seperti Jepang, Cina, Vietnam, dan Thailand. Selain itu infeksi karena V. parahaemolyitcus juga terjadi setelah mengkonsumsi produk perikanan olahan. Hal ini terutama disebabkan oleh pemasakan yang tidak sempurna sehingga tidak membunuh semua V.
parahaemolyitcus yang ada, atau proses penanganan yang buruk seperti kondisi
higiene dan sanitasi tidak terjaga, penyimpanan produk pada suhu ruang selama beberapa jam sebelum diolah/dikonsumsi, atau terjadinya kontaminasi silang antara produk yang telah dimasak dengan produk mentah. Di Thailand, tingkat kontaminasi V. parahaemolyticus pada produk perikanan sebesar 77.5% pada kurun waktu 1971-1972. Penelitian lain menyebutkan terdapat 27% dan 49% produk perikanan beku dan mentah mengandung V. parahaemolyticus. Sementara itu sebanyak 78% seafood mentah di Bangkok pada Mei-Oktober 1994 terkontaminasi V. parahaemolyticus, dimana kontaminasi tertinggi terdapat pada remis (100%), kerang (96%), udang (68%) dan kepiting bakau (51%). Produk udang beku yang siap diekspor juga ditemukan V. parahaemolyticus sebesar 64% pada Mei 1995-Juli 1996. Sampel udang beku di unit pengolahan di Propinsi Chachoengsao-Thailand juga terkontaminasi V. parahaemolyticus sebesar 80% (April-Mei 1999) (Limuthaitip, 1995; Kowcachaporn, 1997; Phayakvichien et al. 1990; Phumiprapat, 1992; Pungchitton, 1999 dalam Jaesawang, 2005).
Infeksi V. parahaemolyticus pada manusia terkait dengan galur patogenik
dari bakteri ini. Galur bakteri penyebab gastroenteritis pada manusia pertama kali diisolasi dari sampel klinis penderita kasus KLB di Calcutta-India tahun 1994-1996 yaitu V. parahaemolyticus O3:K6 (Okuda et al. 1997). V.
parahaemolyticus O3:K6 ini bersifat pandemik di negara Asia Tenggara akan
tetapi bukan merupakan galur yang sama dengan galur O3:K6 yang masuk melalui turis-turis Asia (international travellers) yang terjadi pada tahun 1982-1993 (Chiou et al. 2000; Okuda et al. 1997; Vuddhakul et al. 2000). Galur O3:K6 juga teridentifikasi di USA pada tahun 1998 dan menyebabkan kasus KLB (416 orang) setelah mengkonsumsi tiram mentah (Daniels et al. 2000). Selain itu galur ini pada produk yang sama juga menyebabkan kasus KLB di Connecticut, New Jersey, dan New York (CDC, 1999). Galur patogen lainnya yang dominan
dan menyebabkan peningkatan KLB di dunia adalah O4:K68 dan O1:K
untypeable (KUT), dimana galur ini dilaporkan terkait dekat dengan galur O3:K6
(Martinez-Urtaza et al. 2004).
Faktor Virulen Vibrio parahaemolyticus
Beberapa hasil penelitian melaporkan bahwa V. parahaemolyticus patogenik umumnya diisolasi dari 90% sampel klinis dan hanya sekitar 1-2% dari lingkungan maupun produk perikanan (Kelly dan Dan Stroh, 1988; Miyamoto et
al. 1969; Sakazaki et al. 1968). V. parahaemolyticus patogenik umumnya
dihubungkan dengan kemampuan bakteri ini memproduksi hemolisis. Berdasarkan kemampuannya memproduksi hemolisis , terdapat 3 jenis hemolisis yang dihasilkan oleh V. parahaemolyticus yaitu thermolabile hemolysin (TLH),
thermostable direct hemolysin (TDH), dan TDH related hemolysin (TRH).
Thermolabile hemolysin (TLH) adalah protein dengan berat molekul
berturut-turut 47.5 dan 45.3kDa yang memiliki aktivitas phospholipase/lyso
phospholipase . Hemolisis ini terdapat pada semua galur V. parahaemolyticus
akan tetapi peranannya dalam patogenesis tidak diketahui secara pasti (Bhunia, 2008). Gen penyandi tlh banyak digunakan untuk mengidentifikasi V.
parahaemolyticus pada sampel dengan menggunakan metode berbasis molekuler.
Infeksi oleh V. parahaemolyticus penyebab gastroenteritis pada manusia terkait dengan keberadaan thermostable direct hemolysin (TDH). Kejadian ini dikenal dengan istilah fenomena Kanagawa (KP) dan disebut sebagai faktor virulen. Fenomena Kanagawa (KP+) merupakan aktivitas β hemolisis pada media agar Wagatsuma (mengandung sel darah merah manusia) yang ditandai dengan pembentukan zona bening di sekitar koloni pada media agar setelah diinkubasi pada suhu 37°C selama 18-24 jam (Joseph et al. 1982; Miyamoto et al. 1969; Sakazaki et al. 1968). Keberadaan KP+ terkait dengan produksi TDH yang dapat menyebabkan lisis pada membran sel darah merah melalui pembentukan pori sehingga beberapa ion masuk ke dalam sel dan terjadi pembengkakan sel yang mengakibatkan kematian sel karena ketidakseimbangan ion (Bhunia, 2008). Mekanisme hemolisis yang disebabkan oleh TDH diawali dengan tahap pengikatan membran sel darah merah, selanjutnya terbentuk pori trans membran
yang pada akhirnya mengakibatkan gangguan pada membran sel (Honda et al. 1992). Thermostable direct hemolysin (TDH) bersifat stabil terhadap panas (100°C; 10 menit) dan aktivitas hemolitiknya tidak dapat ditingkatkan dengan penambahan lesitin. Hal ini yang menunjukkan bahwa TDH memiliki aktivitas langsung terhadap sel darah merah (Nishibuchi dan Kaper, 1995). Aktivitas TDH menyebabkan peningkatan konsentrasi kalsium dalam sel sehingga memicu sekresi ion klorida melalui sel intestinal.
Thermostable direct hemolysin (TDH) merupakan protein (terdiri dari 165
asam amino) yang tidak memiliki lipida dan karbohidrat dengan berat molekul 42 kDa. Thermostable direct hemolysin (TDH) terdiri dari 2 sub unit identik yang masing-masing memiliki berat molekul 21 kDa (Honda dan Iida, 1993; Miyamoto
et al. 1980; Takeda et al. 1978 dalam Levin, 2009). Gen penyandi TDH pertama
kali dikloning oleh Kaper et al. (1984) yang disebut tdh1, kemudian ditemukan gen tdh2 oleh Hida dan Yamamoto (1990). Nishibuchi dan Kaper (1990) melaporkan bahwa semua KP+ pada sampel klinis galur V. parahaemolyticus umumnya mengandung gen tdh1 dan tdh2 dan jika galur V. parahaemolyticus menunjukkan aktivitas hemolisin yang rendah pada agar Wagatsuma maka diduga hanya memiliki 1 gen tdh. Thermostable direct hemolysin (TDH) disebut sebagai faktor virulen pertama V. parahaemolyticus dan digunakan untuk mengidentifikasi galur patogenik V. parahaemolyticus (Cook et al. 2002; Okuda et al. 1997).
V. parahaemolyticus patogenik umumnya terkait erat dengan KP+, akan
tetapi Honda et al. (1987 dan 1988) melaporkan bahwa ditemukan V.
parahaemolytic patogenik pada sampel klinis pasien KLB di Maldives pada
tahun1985 yang ditandai dengan Fenomena Kanagawa negatif (KP-). Galur ini diketahui tidak memproduksi TDH tetapi memproduksi TDH-related hemolysin (TRH) yang menyebabkan gangguan kesehatan pada manusia. Shirai et al. (1990) menyatakan bahwa 52 galur dari sampel klinis 8 pasien diare hanya memproduksi TRH sehingga disebut juga sebagai gen patogen V. parahaemolyticus. Gen trh umumnya dikaitkan dengan V. parahaemolyticus yang menunjukkan hasil urease yang positif sehingga sering dijadikan indikator untuk identifikasi V.
parahaemolyticus patogenik walaupun tidak mutlak (Suthienkul et al. 1995;
tdh sebesar 68%. Gen trh diketahui lebih labil terhadap panas karena inaktivasi
gen trh dapat dilakukan pada suhu 65°C selama 15 menit.
Selain kemampuan memproduksi hemolisin, aktivitas urease diduga terkait dengan V. parahaemolyticus patogenik. Umumnya V. parahaemolyticus tidak memproduksi urease (Okuda et al. 1997; Osawa et al. 1996). Kelly dan Stroh (1989) melaporkan bahwa isolasi dari sampel klinis pasien gastroenteritis di Canada menunjukkan semua sampel memberikan hasil urease positif (Uh+) dan hasil Kanagawa negatif. Produksi urease oleh V. parahaemolyticus pada umumnya dikaitkan dengan keberadaan faktor virulen TRH sehingga diduga dapat digunakan untuk identifikasi V. parahaemolytic patogenik yang memproduksi gen
trh ( Osawa et al. 1996). Aktivitas urease dan TRH dilaporkan saling terkait
karena adanya hubungan secara genetis antara gen urease (ureC) dan gen trh pada kromosom dari galur V. parahaemolyticus patogenik (Iida et al. 1997; Park et al. 2000). Namun demikian, gen urease tidak berpengaruh terhadap regulasi ekspresi gen tdh dan trh (Nakaguchi, 2003).
Genom V. parahaemolyticus O3:K6 diketahui memiliki dua gen sistim
sekresi tipe III (TTSS) yaitu TTSS1 dan TTSS2 yan terdapat pada kromoson I dan II. TTSS merupakan factor virulen penyebab diare yang dimiliki oleh bakteri seperti Shigella, Salmonella, dan enteropathogenic E. coli (EPEC). Secara umum mekanisme sistim sekresi tipe III (TTSS) ini terlibat dalam perpindahan protein efektor bakteri menuju sitoplasma sel inang dari bakteri tersebut (Makino et al. 2003). Hal ini diduga juga merupakan faktor virulen yang terdapat pada V.
parahaemolyticus.
Salah satu faktor virulen yang dimiliki bakteri Gram negatif termasuk V.
parahaemolyticus adalah kemampuan menempel pada sel epitel inangnya. V. parahaemolyticus diketahui dapat memproduksi sel yang berasosiasi dengan
hemaaglutinin (HA) pada saat menempel di mukosa usus (Yamamoto dan Yokota, 1989). Hemaaglutinin merupakan glikoprotein antigenik yang berperan dalam proses terikatnya virus pada sel yang terinfeksi sehingga menyebabkan sel darah merah menggumpal. Selain itu pili dari V. parahaemolyticus diduga memiliki peranan dalam hal menempelnya reseptor epitel usus (Nakasone dan Iwanaga, 1990).
Identifikasi V. parahaemolyticus Patogenik
Metode untuk identifikasi V. parahaemolyticus patogenik telah banyak dikembangkan. Identifikasi V. parahaemolyticus patogenik dapat dilakukan dengan metode Kanagawa (KP+) yaitu dengan melihat aktivitas β hemolisis yang ditandai dengan terbentuknya zona bening di sekitar koloni bakteri yang ditumbuhkan pada media agar Wagatsuma. Metode ini memiliki kelemahan antara lain waktu preparasi dan analisis yang panjang, interpretasi hasil analisis yang kurang akurat, sensivitas rendah, dan belum tersedianya media analisis untuk mengidentifikasi faktor virulen TRH. Kendala - kendala metode konvensional untuk mengidentifikasi V. parahaemolyticus patogenik tersebut mendorong pengembangan metode identifikasi dengan hasil yang lebih akurat, sensitifitas tinggi dan tepat serta waktu analisis yang pendek.
Identifikasi V. parahaemolyticus patogenik berbasis pendekatan molekuler
seperti polymerase chain reaction (PCR) telah banyak dilakukan. Metode PCR telah banyak dikembangkan untuk pengujian-pengujian mikrobiologi karena memiliki beberapa keunggulan diantaranya tingkat akurasi dan sensitifitas tinggi, ketepatan hasil uji tinggi, waktu pengujian relatif cepat dan dapat digunakan untuk pengujian komponen yang jumlahnya sangat sedikit.
Metode PCR adalah suatu metode enzimatis untuk melipatgandakan secara eksponensial suatu sekuen nukleotida tertentu dengan cara in vitro. Metode ini pertama kali dikembangkan oleh Kary B. Mullis pada tahun 1985 dan memiliki tingkat sensitifitas tinggi sehingga dapat digunakan untuk melipatgandakan satu molekul DNA dan dapat dilakukan dalam waktu singkat. Proses PCR memiliki 4 komponen utama yaitu cetakan DNA (DNA template), merupakan fragmen DNA yang akan dilipatgandakan; oligonukleatida primer yaitu sekuen oligonukletida pendek (15-25 basa nukleotida) yang digunakan untuk mengawali sintesis rantai DNA; deoksiribonukleotida trifosfat (dNTP) yang terdiri dari dATP, dCTP, dGTP, dTTP; dan enzim DNA polimerase merupakan enzim yang melakukan katalisis reaksi sintesis rantai DNA. Enzim yang digunakan adalah Taq DNA polimerase karena enzim ini tahan terhadap suhu tinggi yang diperlukan untuk tahap pemisahan rantai cetakan DNA (denaturasi) sehingga tidak dibutuhkan penambahan enzim pada tiap siklus PCR (Yuwono, 2006). Selain itu komponen
lain yang berperan pada proses PCR adalah senyawa buffer yang berperan dalam proses penempelan primer (annealing) dimana di dalam senyawa buffer terkandung 10-50 mM Tris-HCl (pH 8.3-8.8), KCl, MgCl2, dan komponen lain
seperti gelatin dan deterjen non ionik seperti Tween 20 untuk mempertahankan kestabilan enzim Taq DNA polimerase.
Metode PCR didasarkan atas 3 tahapan untuk reaksi sintesis DNA (pelipatgandaan fragmen DNA). Tahapan-tahapan pada proses PCR dimulai dari : a. Denaturasi; merupakan tahap awal PCR yang bertujuan untuk memisahkan
rantai DNA yang berantai ganda menjadi rantai tunggal karena pembuatan
copy DNA oleh enzim Taq DNA polimerase membutuhkan DNA berantai
tunggal. Denaturasi dilakukan pada suhu 94°C selama 1-2 menit.
b. Annealing (penempelan); merupakan tahap penempelan primer pada cetakan DNA yang telah berantai tunggal. Primer akan membentuk jembatan hidrogen dengan cetakan DNA pada daerah sekuen yang komplementer dengan sekuen primer. Annealing dilakukan pada suhu 55°C selama 1-2 menit. Suhu 55°C yang digunakan pada tahap annealing memiliki spesifitas reaksi amplifikasi yang lebih tinggi akan tetapi efisiensinya menurun. Amplifikasi lebih efisien pada suhu 37 °C tetapi umumnya terjadi penempelan primer di tempat yang salah. Primer yang digunakan dalam PCR ada dua yaitu oligonukleotida yang memiliki sekuan identik dengan salah satu rantai cetakan DNA pada ujung 5’fosfat dan oligonukleotida yang identik dengan sekuen pada ujung 3’OH pada rantai cetakan DNA lain.
c. Ekstensi; merupakan tahap pembentukan polimerisasi rantai DNA yang baru berdasarkan informasi yang ada pada cetakan DNA. Setelah terbentuk polimerisasi, rantai DNA yang baru akan membentuk jembatan hidrogen dengan cetakan DNA. Tahap ekstensi dilakukan pada suhu 72°C yang merupakan suhu optimum aktivitas Taq DNA polimerase selama 1-2 menit. DNA rantai ganda yang terbentuk antara rantai cetakan DNA dengan rantai DNA hasil polimerasi selanjutnya didenaturasi kembali pada suhu 94°C untuk memperoleh rantai DNA tunggal yang baru dan berfungsi sebagai cetakan reaksi polimerasi selanjutnya.
Reaksi polimerasi ini diulangi kembali sebanyak 25-30 siklus dan pada akhir siklus akan diperoleh molekul DNA rantai ganda hasil polimerasi dalam jumlah yang lebih banyak dibandingkan dengan jumlah cetakan DNA yang digunakan. Selanjutnya DNA hasil polimerasi di elektroforesis pada gel agarose dan divisualisasikan (Sambrook et al. 1989).
Identifikasi V. parahaemolyticus patogenik menggunakan metode PCR
dilakukan dengan cara mengamplifikasi gen tdh dan trh V. parahaemolyticus. Amplifikasi gen tdh dan trh dilakukan menggunakan sepasang primer spesifik dari masing-masing gen penyandi tersebut. Salah satu sekuen nukleotida dan protokol PCR yang digunakan untuk mengidentifikasi V. parahaemolyticus patogenik adalah hasil penelitian dari Tada et al. (1992) yaitu untuk target gen tdh amplifikasi dilakukan pada ukuran amplikon 251 bp dan untuk gen trh pada ukuran amplikon 250 bp.
Sekuen nukleotida baik gen tdh maupun gen trh bakteri V.
parahaemolyticus secara keseluruhan adalah 570 bp. Beberapa hasil penelitian
melaporkan bahwa sekuen nukleotida gen tdh V. parahaemolyticus memiliki keragaman yang relatif sedikit (kurang dari 33%) sedangkan gen trh memiliki sekuen yang lebih bervariasi (gen trh1 dan trh2). Hasil penelitian yang dilakukan oleh Tada et al. (1992) menggunakan beberapa kombinasi pasangan primer untuk gen tdh dan trh serta optimasi protokol PCR terutama untuk penentuan suhu
annealing (penempelan primer), diketahui bahwa pasangan primer untuk
identifikasi gen tdh dan trh V. parahaemolyticus pada ukuran amplikon 251 bp dan 250 bp memberikan hasil dengan spesifisitas dan sensitifitas yang tinggi. Spesifisitas primer dikonfirmasi dengan analisis Southern blot hybridization menggunakan probe gen tdh dan trh. Sedangkan sensitifitas metode PCR dikofirmasi dengan menggunakan DNA genom V. parahaemolyticus WP1 dan AQ4037 yang merupakan galur V. parahaemolyticus penghasil gen tdh dan trh.
Amplifikasi sekuen nukleotida untuk mengidentifikasi gen tdh dan trh bakteri V. parahaemolyticus dengan target gen tdh dan trh pada ukuran amplikon yang berbeda juga telah dilakukan. Bej et al. (1999) melaporkan identifikasi V.
parahaemolyticus dapat dilakukan dengan metode multiplex PCR dimana sekuen
pasangan primer pada ukuran amplikon masing-masing 269 bp (Nishibuchi dan Kaper, 1985) dan 500bp (Honda dan Iida, 1993; Honda et al. 1991). Sementara itu, penelitian Rosec et al. (2009) menyebutkan bahwa untuk identifikasi V.
parahaemolyticus patogenik berdasarkan gen tdh menggunakan sekuen primer
VP21 dan VP22 pada ukuran amplikon 400 bp dan gen trh dengan pasangan primer S1 dan S2 pada ukuran amplikon 460 bp.
METODE PENELITIAN
Tempat dan Waktu Penelitian
Survei dan pengambilan sampel dilakukan di tambak udang tradisional dan intensif yang berlokasi di pantai utara Jawa Barat. Pengujian V.
parahaemolyticus patogenik pada sampel udang tambak dilakukan di
Laboratorium Mikrobiologi dan Laboratorium Bioteknologi Balai Besar Riset Pengolahan Produk dan Bioteknologi Kelautan dan Perikanan, Badan Riset Kelautan dan Perikanan, Kementerian Kelautan dan Perikanan, Slipi, Jakarta Pusat. Penelitian ini dilakukan sejak bulan Agustus 2010 hingga Maret 2011.
Bahan dan Alat
Bahan yang digunakan dalam penelitian ini adalah udang vaname (Litopenaeus vannamei) yang berasal dari tambak tradisional dan insentif. Sebagai kontrol positif digunakan isolat V. parahaemolyticus penghasil TDH dan TRH yang merupakan koleksi Fakultas Farmasi Universitas Andalas-Padang yang berasal dari Kyoto University. Media untuk isolasi dan konfirmasi V.
parahaemolyticus presumtif adalah media cair tryptic soy broth (TSB), alkaline saline pepton water (ASPW), media agar thiosulfate citrate bile salt sucrose
(TCBS), tryptic soy agar (TSA), motility test medium (MTM), triple sugar iron (TSI), dan media untuk pewarnaan gram. Seluruh media, kecuali untuk pewarnaan gram dan TCBS ditambahkan NaCl sampai 3%. Media untuk konfirmasi V.
parahaemolyticus menggunakan API 20E biochemical test kit. Bahan- bahan
untuk isolasi DNA antara lain media cair tryptic soy broth (TSB) +2.5%NaCl, TE
buffer, Sodium Dodecyl Sulphate (SDS), Cetyiltrimethyl Ammonium Bromide
(CTAB), Natrium Chloride (NaCl), proteinase-K, sodium asetat, RNase, fenol, kloroform, isoamil alkohol, isopropanol, etanol 70%. Bahan- bahan untuk elektroforesis DNA meliputi bufer TBE, agarosa, dan sybr safe gold.
Bahan-bahan yang digunakan untuk amplifikasi gen tdh dan trh antara lain Go Taq
PCR, 3mM MgCl2, dan dNTP (masing-masing 0.4 mM dATP, dCTP, dGTP, dan
dTTP), DNA/RNA free water, cetakan DNA, standard DNA ladder 100bp, dan primer oligonukleotida untuk gen tdh dan trh yang masing-masing memiliki ukuran amplikon 251 bp dan 250bp (Tada et al. 1992). Urutan basa primer untuk gen tdh dan trh dapat dilihat pada Tabel 7.
Tabel 7. Primer oligonukleotida yang digunakan untuk deteksi gen tdh dan trh
Target DNA amplikon Ukuran Urutan basa (5’-3’)
GGT ACT AAA TGG CTG ACA TC (forward) Gen tdh 251bp CCA CTA CCA CTC TCA TAT GC (reverse)
GGC TCA AAA TGG TTA AGC G (forward) Gen trh 250bp CAT TTC CGC TCT CAT ATG C (reverse)
Tada et al. (1992)
Alat-alat yang digunakan dalam penelitian ini adalah peti berinsulasi, termometer, botol plastik steril, plastik steril, mikroeppendorf 1.5 ml, 200µl, erlenmeyer 250 ml, 125 ml, timbangan digital, stomacher, vorteks, tabung reaksi bertutup, jarum ose, cawan petri, botol pengencer, refrigerator, refrigerator -20oC, sentrifius, inkubator 37°C, autoklaf, laminar air flow, mikropipet beserta tip ukuran 10, 100, 200, dan 1000µl, DNA/RNA/protein analyser, PCR system
thermal cycler (Gene AMP PCR 9700), perangkat elektroforesa (Model B1A; owl separation system), gel documentation (EC 250-90; EC apparatus coorporation),
dan UV transillumination (UVT 20 M; Herolab, BDA Digital Biometra). Perangkat lunak (software) yang digunakan adalah apiwebTM (Biomeireux) untuk
menganalisis hasil uji biokimia dengan API 20E biochemical test kit.
Pelaksanaan Penelitian
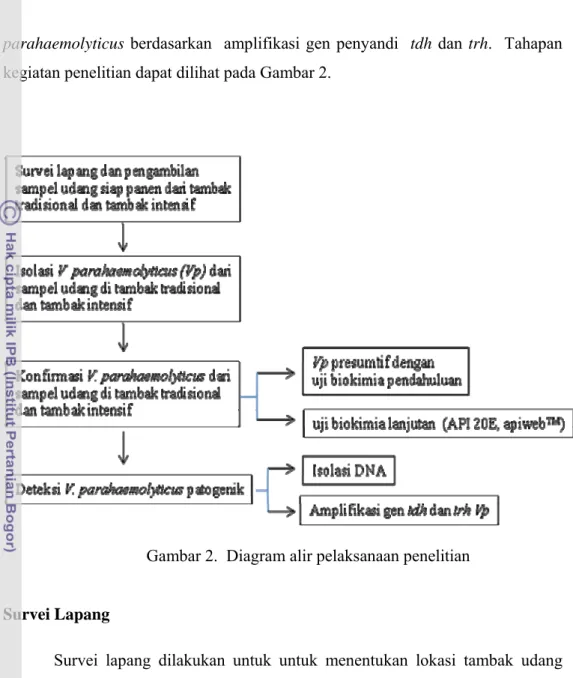
Penelitian dilakukan dalam beberapa tahapan yang meliputi (1) survei lapang lokasi tambak tradisional dan intensif, (2) pengambilan sampel udang dari tambak tradisional dan intensif hasil survei, (3) isolasi V. parahaemolyticus dari sampel udang, (4) karakterisasi sifat fenotipik berdasarkan identifikasi isolat secara biokimiawi, (5) identifikasi V. Parahaemolyticus patogenik dari isolat V.
parahaemolyticus berdasarkan amplifikasi gen penyandi tdh dan trh. Tahapan
kegiatan penelitian dapat dilihat pada Gambar 2.
Gambar 2. Diagram alir pelaksanaan penelitian
Survei Lapang
Survei lapang dilakukan untuk untuk menentukan lokasi tambak udang yang berpotensi mengandung V. parahaemolyticus baik di tambak tradisional dan intensif. Pengumpulan data dan informasi pada survei lapang meliputi :
a. Pengumpulan informasi lokasi dan jumlah tambak udang (tradisional dan intensif) yang masih beroperasi.
b. Pengumpulan informasi waktu panen tambak udang sebagai penentuan pengambilan sampel udang.
c. Pengumpulan informasi yang terkait dengan kondisi lingkungan tambak seperti salinitas, pH, suhu, luas areal tambak, waktu panen, dan data dukung lainnya .