Efficient whole plant regeneration of common bean (Phaseolus
6
ulgaris
L.) using thin-cell-layer culture and silver nitrate
Maria H. Cruz de Carvalho
a,*, Bui Van Le
b,c, Yasmine Zuily-Fodil
a,
Anh T. Pham Thi
a, Kiem Tran Thanh Van
baLaboratoire de Biochimie et Physiologie de l’Adaptation Ve´ge´tale,case7019,UMR7632,Uni6ersite´ Paris7Denis-Diderot,2place Jussieu,
75251Paris Cedex,France
bInstitut de Biotechnologie des Plantes,UMR0569,Uni6ersite´ de Paris-Sud,91405Orsay Cedex,France cNational Uni6ersity HCM City,227Nguyen6an Cu,2nd district,Ho Chi Minh City,Viet Nam
Received 5 June 2000; accepted 13 July 2000
Abstract
A method was designed to optimize rapid and high frequency direct shoot regeneration (without intermediate callus) of the commercially important common bean, Phaseolus 6ulgaris L., using the transverse thin cell layer (tTCL) method. The pretreatment of seeds with 10mM TDZ significantly increased bud regeneration frequency on tTCL. A 2-week culture of tTCLs on 10mM TDZ followed by a reduction in the TDZ concentration (1mM) was needed to achieve optimal bud induction and further development of the neo-formed buds. An incubation period greater than 2 weeks of tTCLs with 10mM TDZ concentration resulted in inhibitory effects on the development of shoots and roots. Shoot development was enhanced by 10mM BAP and 10 mM AgNO3 leading to 100% well developed shoots. Regenerated plants developed into true-to-type fertile plants. © 2000
Published by Elsevier Science Ireland Ltd. All rights reserved.
Keywords:Transverse thin cell layers; Common bean; Direct shoot regeneration; Thidiazuron; Silver nitrate; Particle bombardment
www.elsevier.com/locate/plantsci
1. Introduction
The common bean (Phaseolus 6ulgaris L.) is an annual, predominantly self-pollinated legume that constitutes a major source of dietary protein throughout both Latin America and Eastern Africa [1]. However, it is a crop which is very sensitive to drought [2,3], as well as to a vast array of pathogens such as bacteria, virus, fungus and insects, which largely reduce yield. The production of transgenic bean, through genetic engineering, with the introduction of drought resistance or defence response genes should be highly beneficial as it could raise the yield or improve the seed quality of this important legume. However, trans-formation procedures can only be successful if an
efficient protocol of regeneration has previously been established. Therefore, the first major objec-tive of the present study was to develop a reliable and highly efficient regeneration protocol which would enable large-scale production of true-to-type bean plants. Some bean regeneration proce-dures have been described from shoot apex cultures [4,5], from cotyledonary and primary leaves node explants [6] devoid of axillary meris-tems [7,8] or from cotyledonary node explants with axillary buds and a portion of the cotyledons [9], from explants consisting of the petiole and part of the blade of juvenile leaves [10] and from embryo-derived callus [11]. The problem underly-ing these procedures is a low frequency of regener-ation. High-frequency induction of direct shoot formation from intact seedlings has been estab-lished by Malik and Saxena [12] using thidiazuron (TDZ) and N6-benzylaminopurine (BAP).
Never-* Corresponding author. Tel.: +33-1-44273617; fax: + 33-1-44276068.
E-mail address:[email protected] (M.H. Cruz de Carvalho).
theless, such a regeneration procedure using in-tact seedlings is not suitable for transformation since two phenomenons are likely to occur: re-generants developed on a voluminous inoculum might escape from the selective conditions and/ or, the killing of the non-transformed tissue un-der selective conditions might block the regeneration process. Hence, our second objec-tive was to achieve a successful direct shoot re-generation using transverse thin cell layers, thus avoiding intact seedling or voluminous inoculum and therefore raising the compatibility of the proposed protocol with transformation proce-dures.
In the present paper, we report on the mor-phogenetic potential of P. 6ulgaris cv. Carioca transverse thin cell layers (tTCLs) to regenerate multiple shoots directly (without a callus phase). The TCL method [13 – 15] has already been used to regenerate other leguminous species such as Psophocarpus tetragonolobus [16], soybean [17], and cowpea [18]. Furthermore, we show that the use of thidiazuron (TDZ), a substituted pheny-lurea derivative with a high cytokinin-like activ-ity, and silver nitrate (AgNO3) a potent ethylene action inhibitor, results in highly efficient differ-entiation and development of shoots.
Up to now, regarding genetic transformation attempts with this crop, no Agrobacterium -medi-ated transformation has been successfully achieved except with another species, Phaseolus
acutifolius A. Gray [19]; there are some cases
where stable transgenic plants were obtained by direct gene transfer through particle bombard-ment [20 – 23]. In this work, a preliminary assay to assess the feasibility of the combination of this system with transformation by biolistic was also undertaken.
2. Materials and methods
2.1. Preparation of seedlings and tTCLs culture
Seeds of P. 6ulgaris L. cv. Carioca (Brazil)
were washed thoroughly with distilled water and Tween-20 (10%). They were then sonicated for 5 min and rinsed three times with sterile distilled water. Subsequently they were surface sterilized with 70% ethanol for 2 min followed by sodium hypochloride (6%) for 15 min. After five rinses
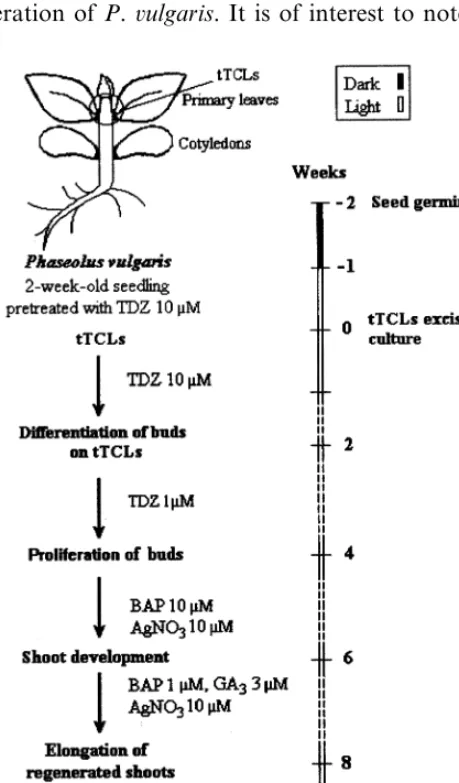
with sterile distilled water, they were allowed to germinate aseptically on half strength Murashige and Skoog (MS) medium [24] solidified with 0.4% agar – agar (Difco). The effect of the pre-treatment of the seedlings by 10 mM thidiazuron (TDZ) on subsequent regeneration expression was assessed by supplying the germination media with this compound. In addition, the effect of light on the same regeneration expression was evaluated by using seedlings germinated in the dark or under a cool fluorescent light (100 mmol m−2 s−1) at 2591°C.
Thin cell layers (0.3 – 0.5 mm thick) were ex-cised transversely (tTCLs) from 2-week-old seedlings at four zones on the plantlet: epicotyl, hypocotyl, cotyledons and roots and were cul-tured on Petri dishes (10 cm in diameter) with 20 ml MS medium supplemented with Gam-borg’s vitamin mix [25] (termed MSB5) and dif-ferent concentrations of TDZ or BAP (1 – 10
mM). From this preliminary assay, the responsive zone was determined and used for subsequent study. All culture media were solidified with 0.6% agar – agar and adjusted to pH 5.7 with 0.1 N NaOH or 0.1 N HCl before autoclaving at 121°C for 15 min. The tTCLs were cultured in a 16-h photoperiod (100 mmol m−2 s−1) at 259 1°C with 60% humidity. Every 7 – 10 days tTCLs were subcultured on fresh culture medium. In an attempt to improve regeneration efficiency and/ or development of regenerated buds, an ethylene action inhibitor, silver nitrate (AgNO3), was tested.
For each treatment 10 – 15 seedlings were used (i.e. approximately 150 tTCLs) and each experi-ment was repeated at least twice. Observation was made every week and treatment effects were quantified on the basis of the number of regener-ants obtained on all tTCLs originated from one original seedling. Statistical significance (rejection at P\0.05) of the different treatments used was assessed by the Kruskal – Wallis one-way Anova using the software Statistix© for Windows (ver-sion 2.0, Analytical Software).
2.2. Scanning microscope obser6ations
2.3. Transformation procedure and gus assay
Explants to be bombarded consisted of the api-cal and subapiapi-cal areas of the seedling (1/1.5 cm) prepared from 2-week-oldPhaseolusseedlings ger-minated as described previously. They were inocu-lated (apex to the top) on MSB5 medium containing 3 mM TDZ and 2.5% agar – agar and left at 22°C for 24 h before bombardment was undertaken.
The pBI211 plasmid vector was used [26]. This plasmid contains the chimeric gene encoding the GUS reporter gene under the control of the CaMV35S promoter. DNA-coated particles were prepared by resuspending tungsten particles (M10, Biorad) in sterile water and adding DNA to a final concentration of 0.01 mg DNA/mg tungsten with 170 mM spermidine and 18.8 M CaCl2. The sus-pension was left on ice for 10 min and then centrifuged. Precipitated DNA-coated particles were resuspended in 100 ml 100% ethanol and mixed by sonication (Tomy Seiko Co. Ltd., Japan). Each Petri plate containing 10 – 20 ex-plants was bombarded three times (each shot de-livering 150 – 225 mg DNA).
After 48 h, bombarded and control explants were tested for gus gene expression. The histo-chemical analysis for b-glucuronidase activity was done according to Jefferson and co-workers [27]. Explants were incubated overnight at 37°C in a 2 mM 5-bromo-4-chloro-3-indolyl glucuronide (X-gluc) solution buffered with 50 mM sodium phos-phate buffer at pH 7.
3. Results
3.1. Factors affecting germination and bud regeneration from tTCLs
In order to improve the frequency of bud regen-eration in tTCLs several factors were analysed separately or simultaneously.
3.1.1. Germination
The germination percentage of seeds was higher when it occurred in the dark as opposed to light (85 and 50%, respectively). Germinating seedlings were maintained in the dark for 1 week and then were transferred to light where they remained for another week before tTCLs were excised.
Further-more, tTCLs excised on seedlings that had grown in the dark for 1 week neoformed a higher number of buds than those which had not been subjected to dark conditions (data not shown). Thus, seeds were systematicaly allowed to germinate in the dark for 1 week before being transferred to light for another week.
3.1.2. Determination of the seedling’s responsi6e
zone
Transverse TCLs were excised from 2-week-old seedlings and cultured in MSB5 medium supple-mented with TDZ or BAP. After 2-week culture, proliferating tTCLs showing shoot organogenesis were obtained with tTCLs excised exclusively at the epicotyl, more specifically from a zone between the primary leaves internode and the apex of the seedling (approximately 5 – 10 responsive tTCLs per seedling). On tTCLs excised at the cotyle-donary nodes, the frequency was B1%. Trans-verse TCLs from the hypocotyl, cotyledons and roots turned into dark brown calli and did not regenerate. Therefore, all subsequent treatments were effected on tTCLs excised at the epicotyl zone.
3.1.3. Induction medium
The optimal induction medium for bud prolifer-ation on tTCLs was assessed using TDZ or BAP at a concentration range of 0 – 10 mM supple-mented with MSB5medium. After 6-week culture, the number of buds neoformed was scored. Analy-sis of data by Kruskal – Wallis one-way Anova was undertaken and the result indicates that the differ-ential response of tTCLs regarding the growth regulator used was highly significant (P=0.0001). Maximal bud induction was obtained with a con-centration of 10 mM for both growth regulators but the average number of buds obtained on TDZ supplemented medium was 46.2 (per initial seedling) as opposed to 25.3 buds obtained on BAP supplemented medium (Table 1). Therefore, tTCLs bud primary induction medium for subse-quent experiments consisted of MSB5 medium supplemented with 10 mM TDZ.
3.1.4. Pretreatment of seeds
Table 1
Effect of induction medium on the regeneration of buds inP.
6ulgarisL. cv. Carioca
Concentrationa Averageb number of bud primordia per
original seedling
aConcentration of TDZ or BAP supplemented to MSB 5
medium.
bAverage of three experiments each using 10 seedlings
(total of 150 tTCLs excised at the seedlings responsive zone). Data scored after 6 weeks culture. Figures in parenthe-ses represent the range observed.
medium (MSB5+10 mM TDZ) was found to be optimal for bud induction without callus forma-tion. However, it was observed that a prolonged exposure to this TDZ concentration had an in-hibitory effect on further development of shoots and roots. Therefore, the optimal duration of tTCL culture on primary induction medium was assessed. Following 0-, 1-, 2-, 3- and 4-week cul-ture on this high TDZ medium, tTCLs were trans-ferred to a secondary medium consisting of MSB5 medium supplemented with a lower concentration of TDZ (1 mM) or to MSB5 medium without any growth regulator (Table 2). Statistical analysis of data indicate that the differences between the du-ration of tTCL culture on primary induction medium were significant (P=0.0002). Also, ex-tremely significant differences were found between the secondary media tested (P=0.0000). Hence, optimal bud regeneration and avoidance of TDZ inhibitory effects was achieved by a three-step procedure: a seed pretreatment with 10 mM TDZ and culture in the dark for 1 week, excision and culture of tTCLs on a primary induction medium consisting of MSB5+10 mM TDZ for 2 weeks followed by a transfer of proliferating tTCLs to a secondary medium of reduced TDZ concentration (MSB5+1 mM TDZ) (Table 2).
3.3. Mode of regeneration
Buds neoformed directly on the surface of the responsive tTCL without callus formation as can be observed from scanning electron microscopy TDZ. Following this time, tTCLs were excised and
cultured on primary induction medium (MSB5+ 10mM TDZ). After 6 weeks culture the number of neoformed buds was scored. Analysis of data indi-cate that the differences found are significant (P= 0.0005). The seed pretreatment raised by 48% the average number of buds obtained (per initial seedling).
3.2. Optimal procedure for maximal bud neoformation on tTCLs
A seed preatreatment with 10 mM TDZ fol-lowed by tTCL culture on primary induction
Table 2
Regenerative response of P. 6ulgaris L. cv. Carioca tTCL as modulated by the pretreatment of seeds, the duration of tTCLs exposure to 10mM TDZ and the subsequent culture medium (with or without 1mM TDZ)
No. weeks on TDZ (10mM)a Averageb number of regenerated bud primordia per original seedling
MSB5 MSB5+TDZ (1 mM)
6.3 (1–14) 29.7 (26–30)
1 28.2 (21–35) 32.8 (17–50)
2 5.5 (2–9) 31 (14–54) 36.4 (31–40) 68.5 (36–134)
23.3 (19–30) 52.3 (23–80) 57 (20–95)
4 54.3 (49–60)
aMedium in which excised tTCLs were initially cultured.
bAverage of three experiments each using 10 seedlings (total of 150 tTCLs excised at the seedlings responsive zone). Data
scored after 6 weeks of culture. Figures in parentheses represent the range observed.
cSeeds were germinated in normal germination medium.
Table 3
Effect of silver nitrate (AgNO3) and N6-benzylaminopurine
(BAP) on shoot development of P. 6ulgaris L. cv. Carioca
Shoot developing media % of Range of shoot
length (cm)
aAverage percentage of shoots which developed from
ini-tial buds on 6-week-old tTCLs (N=265). Data scored after 2 weeks culture.
3.5. Rooting and transplantation of the neoformed shoots
Rooting was obtained on MSB5 containing 1
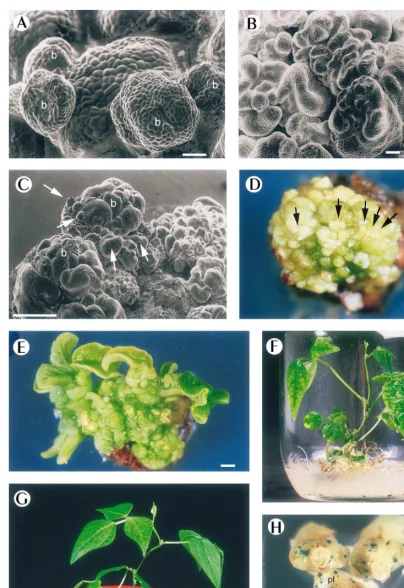
mM NAA and 10mM AgNO3(Fig. 1F) on 80% of the regenerated plants. After 2 – 3 weeks, they were transferred to the greenhouse in 8-cm pots con-taining a mixture of peat and vermiculite (50/50, v/v) (Fig. 1G). Roots were previously washed in order to eliminate any remaining culture medium. Maximal humidity was assured during the first week by placing an inverted glass beaker over each plant before transfer to field conditions. The sur-vival rate of rooted plantlets transferred to soil was 95% and these were fertile with normal pod development.
3.6. Expression of b-glucuronidase following bombardment
In order to determine the capacity of explants to be transformed, gene transfer was undertaken us-ing a biolistic approach. Explants of 1/1.5 cm consisting of the seedling zone shown to be re-sponsive to bud regeneration were bombarded with tungsten beads containing a binary vector with the uidA (GUS) gene under the transcrip-tional control of the CaMV35S promoter. Two days after the bombardment, explants were incu-bated overnight at 37°C with X-gluc and the strong blue staining was observed (Fig. 1H). The bombarded explants tested were excised to tTCLs and these remained viable (data not shown).
4. Discussion
The lack of ability to regenerate common bean with high frequency has been the main reason of the failure to produce transgenic plants efficiently. Transfer of genes into beans by theAgrobacterium system has been attempted, but no regeneration of transformed plants was achieved [29]. Stable trans-genic bean plants were obtained using an electrical acceleration device for the delivery of DNA to embryos [20]. However, the tissue culture protocol was time-consuming and with a low regeneration frequency (an average of two shoots/explant har-vested) leading to a low frequency of transgenic plants (0.03%). More recently, stable transfor-mants by bombardment of DNA were attained by (Fig. 1A – C). No callus occurred on proliferating
tTCLs (Fig. 1D). Further buds originated at the base of neoformed shoots leading to a high density of buds on tTCLs (Fig. 1C).
3.4. Effect of AgNO3 on shoot de6elopment
Full development of shoots was obtained when bud primordia were transferred to MSB5 medium containing 10 or 20 mM BAP which allowed full leaf development as opposed to TDZ under the same conditions (data not shown). However, only a small number developed. Based on previous results [28], an inhibitor of ethylene action (AgNO3) was added to a medium with or without BAP. Analysis of data indicate that the combina-tion of BAP and AgNO3at equimolar concentra-tions (10 mM) significantly raised the number of developed shoots. An average of 100% developed shoots were scored with a length range of 1.8 – 2.9 cm indicating that these conditions were optimal for shoot development (Table 3, Fig. 1E). Other concentrations of AgNO3(1 mM, 50mM) tested in combination or not with BAP had no effect (data not shown).
Fig. 1. A – G. Direct regeneration of P. 6ulgaris (cv. Carioca) plants from transverse TCLs. A, B, C, Scanning microscope
observation of tTCLs. (A) Newly formed buds, b, arose directly without callus formation after 10 days culture. Bar=100mm. (B) Clusters of shoot buds continue to proliferate after 2 weeks culture, bar=100mm, and after 3 weeks culture, newly formed buds (arrows, C) arose at the base of existing ones (b). Bar=1 mm. (D) Four-week-old tTCLs presenting multiple buds (arrows). Bar=1 mm. (E) Cluster of shoots on developing medium at 6-week stage. Bar=1 mm. (F) Well-developed shoot cultured separately on rooting medium at 9-week stage. Bar=5 mm. (G) RegeneratedP.6ulgarisplant at 11-week stage. Bar=3 cm. (H) Gus expression on 2-week-oldP. 6ulgarisseedling explants comprising the responsive zone of bud regeneration, after particle
bombardment with the pBI211 vector carrying theuid Agene under the control of the 35SCaMV promoter,pl, primary leaves,
other researchers [22,21], but in one case, one bombarded embryonic axis gave rise to one trans-formed plant (six plants transtrans-formed out of the 319 bombarded embryonic axes) since no multiple shoot induction was possible [22] and in the other, only 9% regenerated shoots were obtained after the bombardment of 3079 embryonic axes, with an average transformation frequency of 0.9% [21].
In this study, we utilised an efficient and repro-ducible method (Fig. 2) to obtain a high number of well-developed common bean regenerants. The TLC system which has been successfully used for the regeneration of different plants, including re-calcitrant leguminous [16 – 18], ligneous plants [30], dicotyledonous [13,31] and monocotyledonous species [32 – 36], is perfectly adapted for the regen-eration of P. 6ulgaris. It is of interest to note that
as opposed to the findings of Malik and Saxena [12] using intact seedlings ofP.6ulgaris, the tTCLs
system permits high frequency shoot neoformation and plant regeneration without the need of mor-phological integrity. A recent work, using more classical, voluminous explants such as individual cotyledons or hypocotyl segments from eucalyptus plants [37], presented a callus phase and a rela-tively high time interval for bud differentiation, 7 – 8 weeks instead of 2 weeks with tTCLs.
Our study has described the efficacy of thidi-azuron (TDZ) on direct multiple shoot formation from tTCLs. TDZ cytokinin activity has been shown to be comparable or even superior to com-mon adenine-based cytokinins [38,39]. Since then, TDZ has been successfully used in many tissue culture protocols of different grain leguminous known to be somewhat recalcitrant to regenera-tion procedures such as pea, chickpea and lentil [40], peanut [41], faba bean [6] cowpea [18] and common bean [6,12]. The mechanism by which TDZ induces morphogenetic responses equivalent to amino purine cytokinines is still not fully understood.
In this context, an interesting feature was that 2-week exposure to high TDZ concentration (10
mM) was sufficient to achieve optimal bud induc-tion on P.6ulgaristTCLs. Similarly, other authors
have demonstrated that a continuous exposure of tissue explants to TDZ is not needed since cultures after exposure to TDZ showed cytokinin auton-omy [12,42]. Furthermore, our results demonstrate the inhibitory effect of a prolonged exposure (4 weeks) to a high concentration of TDZ (10 mM) on shoot and root development. This could be due to the fact that TDZ, similar to other synthetic cytokinines, is less susceptible to enzymatic degra-dation by the plant when compared to the endoge-nous amino purine cytokinines [43]. Furthermore, it has also been suggested that TDZ may be involved in the synthesis and/or accumulation of cytokinines in tissue cultures [39,42].
In this study, we showed that the addition of 10
mM silver nitrate (AgNO3) to the medium with BAP (10 mM) enhanced the number of developed shoots per explant as well as the length range of shoots (Table 3). In a recent report, on the regen-eration of two eucalyptus species (Eucalyptus nitens and E. globulus), no significant effect was found with the use of silver ions and although shoot regeneration occurred from callus-derived
seedling explants, the frequency achieved was not very important (20 – 35%) [37]. Plant cell tissues and whole plants grown in culture produce ethyl-ene which has been shown to be involved in plant cell differentiation [44]. In our work, the use of AgNO3, a potent inhibitor of ethylene action [45] with BAP had a synergistic effect on enhanced shoot regeneration in P. 6ulgaris. When BAP was
used alone, only 63.8% of the shoots developed (with a length range of 1.3 – 1.7 cm) whereas when it was applied in combination with AgNO3, 100% of the shoots developed (1.8 – 2.9 cm length range). Similar results were obtained on the promotion of morphogenesis on cotyledonary node explant ofP.
6ulgaris cultured on MS medium supplemented
with 1 mg/l BAP, by the use of ethylene inhibitors [28], and it has also been suggested that the com-mon recalcitrance of cowpea to regeneration could be due to ethylene [46]. Ethylene production could be the main reason of the failure to regenerate efficiently leguminous species.
Preliminary results conducted on gene transfer by biolistic of P. 6ulgaris explants (excised from 2-week-old seedling), originated strong transient expression of the uidA (GUS) reporter gene (Fig. 1H), opening the way to devise reproducible trans-formation procedures for this species using tTCLs. Recent reports have shown that the same bean cultivar (P. 6ulgaris cv. Carioca) had a shoot apex morphology favourable for the efficient delivery of genes by biolistic [23], but shoot apex cultures would regenerate only 6 – 8 shoots per apex. In our preliminary assay, bombarded explants main-tained their capacity to give rise to tTCLs which respond positively to bud regeneration (results not shown). This tTCL method should enable the production of large-scale transformed material. TCLs system (transverse or longitudinal) has al-ready been successfully associated with transfor-mation procedures [47 – 49]. Recently, high temperature tolerance has been improved in trans-formed tobacco plants by gene silencing of a chloroplast omega-3 fatty acid desaturase [50]. It should be extremely valuable to test this achieve-ment on bean plants, since drought/high tempera-tures are major environmental problems involving this crop.
Since we emphasise the necessity of a highly productive regeneration method in order to raise the frequency of transformed plants, the notewor-thy aspects of this protocol are: (1) tTCLs are
simple to produce and integrity of seedlings is not needed for the induction of multiple bud prolifera-tion, hence they can be submitted to the selective conditions associated with the screening of trans-formed material; (2) the high frequency of regener-ation (an average of 68.5 well developed shoots per seedling) should increase the frequency of fully transformed shoots; (3) shoot apex explants have proven to be suitable to gene delivery by biolistic and tTCLs proliferation capacity was not lost after bombardment.
Acknowledgements
The authors wish to thank Professor P. Gadal and Dr J. Vidal for their encouraging support, R. Boyer (CNRS) for the tissue culture photographs and M. Grasset (Centre Interuniversitaire de Mi-croscopie Electronique, Universite´ Paris VI) for the scanning electron microscope photographs. Thanks are also due to M. Jeanneau, N. Thao Do My and E. Sadik for interesting discussion and finally, to Professor J. Vieira da Silva for his valuable help on the statistical treatment of the data. M.H. Cruz de Carvalho is supported by a grant from the ‘Sub-programa Cieˆncia e Tecnolo-gia do 2° Quadro Comunitario de Apoio’ (PRAXIS XXI/ BD/ 9149/ 96, Portugal).
References
[1] P.H. Graham, P. Ranalli, Common bean (Phaseolus
6ulgarisL.), Field Crops Res. 53 (1997) 131 – 146. [2] A.J. Haterlein, Bean, in: I.D. Teare, M.M. Peet (Eds.),
Crop Water Relations, Wiley, New York, 1983, pp. 157 – 185.
[3] M.H. Cruz de Carvalho, D. Laffray, P. Louguet, Com-parison of the physiological responses ofPhaseolusand
Vigna cultivars when submitted to drought conditions, Environ. Exp. Bot. 40 (3) (1998) 197 – 207.
[4] K.K. Kartha, K. Pahl, N.L. Leung, L.A. Morginski, Plant regeneration from meristems of grain legumes: soybean, cowpea, peanut, chickpea and bean, Can. J. Bot. 59 (1981) 1671 – 1679.
[5] I.S. Martins, M.R. Sondahl, Multiple shoot formation from shoot apex cultures of Phaseolus 6ulgaris L, J.
Plant Physiol. 115 (1984) 205 – 208.
[7] P. McClean, K.F. Grafton, Regeneration of dry bean (Phaseolus 6ulgaris L.) via organogenesis, Plant Sci. 60
(1989) 117 – 122.
[8] D. Mariotti, G.S. Fontana, L. Santini, Genetic transfor-mation of grain legumes: Phaseolus 6ulgaris L. and P.
coccineusL, J. Genet. Breed. 43 (1989) 77 – 82.
[9] C.I. Franklin, T.N. Trieu, R.A. Gonzales, R.A. Dixon, Plant regeneration from seedling explants of green bean (Phaseolus 6ulgaris L.) via organogenesis, Plant Cell
Tissue Organ Cult. 24 (1991) 199 – 206.
[10] K.A. Malik, P.K. Saxena, Regeneration in Phaseolus
6ulgarisL.: Promotive role of N6-benzylaminopurine in
cultures from juvenile leaves, Planta 184 (1991) 148 – 150. [11] M.A. Zambre, J. De Clercq, E. Vranova´, M. Van Mon-tagu, G. Angenon, W. Dillen, Plant regeneration from embryo-derived callus inPhaseolus6ulgarisL. (common
bean) and P. acutifolius A. Gray (tepary bean), Plant Cell Reports 17 (8) (1998) 626 – 630.
[12] K.A. Malik, P.K. Saxena, Regeneration in Phaseolus
6ulgaris L.: High-frequency induction of direct shoot
formation in intact seedlings by N6-benzylaminopurine
and thidiazuron, Planta 186 (1992) 384 – 389.
[13] K. Tran Thanh Van,In6itroofde no6oflower, bud, root and callus differentiation from excised epidermal tissue, Nature 246 (1973) 44 – 45.
[14] K. Tran Thanh Van, Control of morphogenesis, Annu. Rev. Plant Physiol. 32 (1981) 129 – 311.
[15] K. Tran Thanh Van, C. Gendy, Thin cell layer (TCL) method to programme morphogenetic patterns, Plant Tissue Cult. Manage. H4 (1996) 1 – 25.
[16] K. Tran Thanh Van, H. Lie-Schricke, J.L. Marcotte, T.H. Trinh, Winged bean (Psophocarpus tetragonolobus
(L.) DC), in: Y.P.S. Bajaj (Ed.), Biotechnology in Agri-culture and Forestry, vol. 2 Crop I, Springer, Berlin, 1986, pp. 556 – 567.
[17] F. Jullien, R. Wyndaele, Precociousin6itroflowering of
soybean cotyledonary node, J. Plant Physiol. 140 (1992) 251 – 253.
[18] B. Van Le, M.H. Cruz de Carvalho, Y. Zuily-Fodil, A.T. Pham Thi, K. Tran Thanh Van, Plant regeneration from the cotyledonary nodes of Vigna unguiculata (L.) Walp: Influence of shoot tip removal and thin cell layer explants, Plant Cell Tissue Organ Cult., submitted. [19] W. Dillen, J. De Clercq, A. Goossens, M. Van Montagu,
G. Angenon,Agrobacterium-mediated transformation of
Phaseolus acutifolius A. Gray, Theor. Appl. Genet. 94 (2) (1997) 151 – 158.
[20] D.R. Russell, K.M. Wallace, J.H. Bathe, B.J. Martinell, D.E. McCabe, Stable transformation of Phaseolus 6ul
-garis via electric-discharge mediated particle accelera-tion, Plant Cell Reports 12 (1993) 165 – 169.
[21] F.J.L. Araga˜o, L.M.G. Barros, A.C.M. Brasileiro, S.G. Ribeiro, F.D. Smith, J.C. Sanford, J.C. Faria, E.L. Rech, Inheritance of foreign genes in transgenic bean (Phaseolus6ulgarisL.) co-transformed via particle
bom-bardment, Theor. Appl. Genet. 93 (1996) 142 – 450. [22] J.W. Kim, T. Minamikawa, Transformation and
regen-eration of French bean plants by the particle bombard-ment process, Plant Sci. 117 (1996) 131 – 138.
[23] F.J.L. Araga˜o, E.L. Rech, Morphological factors influ-encing recovery of transgenic bean plants (Phaseolus
6ulgaris L.) of a Carioca cultivar, Int. J. Plant Sci. 158
(2) (1997) 157 – 163.
[24] T. Murashige, F. Skoog, A revised medium for rapid growth and bioassay with tobacco tissue cultures, Phys-iol. Plant. 15 (1962) 473 – 497.
[25] O. Gamborg, R. Miller, K. Ojima, Nutrients require-ments of suspension cultures of soybean root cells, Exp. Cell Res. 50 (1968) 151 – 158.
[26] R.A. Jefferson, Assaying chimeric genes in plants: the GUS gene fusion system, Plant Mol. Biol. 5 (1987) 387 – 405.
[27] R.A. Jefferson, T.A. Kavangh, M.W. Bevan, GUS fu-sions: beta glucuronidase as a sensitive and versatile marker in plants, EMBO J. 6 (1987) 3902 – 3907. [28] R.A. Hoyos, G.L. Hosfield, Ethylene inhibitors added to
in 6itro culture media of common bean promotes
mor-phogenesis of cotyledonary node explants, Annu. Rep. Bean Improv. Coop (BIC) 38 (1995) 99 – 100.
[29] P. McClean, P. Chee, J. Simental, R.F. Drong, J. Slighton, Susceptibility of dry bean (Phaseolus 6ulgaris
L.) to Agrobacterium infection: Transformation of cotyledonary hypocotyl tissues, Plant Cell Tissue Organ Cult. 24 (1991) 131 – 138.
[30] K. Tran Thanh Van, B. Van Le, Current status of thin cell layer method for the induction of organogenesis or somatic embryogenesis, in: S.J. Mohan, P.K. Gupta, R.J. Newton (Eds.), Somatic Embryogenesis in Woody Plants, vol. 6, Kluwer, Dordrecht, 1999.
[31] B. Van Le, N.T. Do My, M. Jeanneau, S. Sadik, S. Tu, J. Vidal, K. Tran Thanh Van, Rapid plant regeneration on a C4dicot species:Amaranthus edulis, Plant Sci. 132
(1998) 145 – 154.
[32] P. Lakshmanan, C.-S. Loh, C.-J. Goh, An in 6itro
method for rapid regeneration of a monopodial orchid hybridArandaDeborah using thin section culture, Plant Cell Reports 14 (1995) 510 – 514.
[33] B. Van Le, M. Jeanneau, N.T. Do My, J. Vidal, K. Tran Thanh Van, Rapid regeneration of whole plants in large crabgrass (Digitaria sanguinalis L.) using thin cell layer culture, Plant Cell Reports 18 (1998) 166 – 172.
[34] B. Van Le, T.N. Duong, K. Tran Thanh Van, Plant production via shoot regeneration from thin cell layer pseudo-bulblet explants ofLilium longiflorum in6itro, C. R. Acad. Sci. Paris 322 (1999) 303 – 310.
[35] B. Van Le, N.T. Hang Phuong, L.T. Anh Hong, K. Tran Thanh Van, High frequency shoot regeneration from Rhynchostylis gigantea (Orchidaceae) using Thin Cell Layer, Plant Growth Regul. 28 (3) (1999) 179 – 185. [36] D. Tan Nhut, B. Van Le, M. Tanaka, K. Tran Thanh Van, Shoot induction and plant regeneration from re-ceptacle tissues of Lilium longiflorum, Sci. Hortic. 1520 (2000) 1 – 8.
[37] S. Bandyopadhyay, K. Cane, G. Rasmussen, J.D. Hamill, Efficient plant regeneration from seedling ex-plants of two commercially important temperate euca-lypt species — Eucalyptus nitensandE.globulus, Plant Sci. 140 (1999) 189 – 198.
[38] M.C. Mok, D.W.S. Mok, D.J. Armstrong, K. Shudo, Y. Isogai, T. Okamoto, Cytokinin activity of N-phenyl-N%
[39] J.C. Thomas, F.R. Katterman, Cytokinin activity in-duced by thidiazuron, Plant Physiol. 81 (1986) 681 – 683.
[40] K.A. Malik, P.K. Saxena, Thidiazuron induces high frequency shoot regeneration in intact seedlings of pea (Pisum sati6um), chickpea (Cicer arietinum) and lentil (Lens culinaris), Aust. J. Plant Physiol. 19 (1992) 731 – 740.
[41] R. Gill, P.K. Saxena, Direct somatic embryogenesis and regeneration of plants from seedling explants of peanut (Arachis hypogaea): promotive role of thidi-azuron, Can. J. Bot. 70 (1992) 1186 – 1192.
[42] S.C. Capelle, D.W.S. Mok, J.E. Turner, Effects of
thidiazuron on cytokinin autonomy and the
metabolism of N6-(D2-isopentenyl) [8-14C]adenosine in
callus cultures of Phaseolus lunatus L, Plant Physiol. 73 (1983) 796 – 802.
[43] M.C. Mok, D.W.S. Mok, J.E. Turner, Biological and biochemical effects of cytokinin-active phenylurea derivatives in tissue culture systems, Hortscience 22 (1987) 1194 – 1197.
[44] E.C. Pua, G.L. Chi,De no6oshoot morphogenesis and plant growth of mustard (Brassica juncea) in 6itro in
relation to ethylene, Physiol Plant. 88 (1993) 467 – 474.
[45] E.M. Beyer, A potent inhibitor of ethylene action in plants, Physiol. Plant. 58 (1976) 268 – 271.
[46] M.S. Brar, M.J. Moore, J.M. Al-Khayri, T.E. More-lock, E.J. Anderson, Ethylene inhibitors promote in
6itro regeneration of cowpea (Vigna unguiculata L.), In
Vitro Cell Dev. Biol. Plant 35 (3) (1999) 222 – 225. [47] T.H. Trinh, S. Mante, E.-C. Pua, N.-H. Chua, Rapid
production of transgenic flowering shoots and F1 progeny from Nicotiana plumbaginifolia epidermal peels, Bio/Technology 5 (1987) 1081 – 1084.
[48] P.J. Charest, L.A. Holbrook, J. Gabard, V.N. Iyer, B.L. Miki, Agrobacterium mediated transformation of thin cell layer explants from Brassica napus L, Theor. Appl. Genet. 75 (1988) 438 – 445.
[49] B. van Le, S. Tu, M. Jeanneau, N.T. Do My, J. Vi-dal, K. Tran Thanh Van, Transformation of a C4
monocot: Digitaria sanguinalis (Large crabgrass) using Thin Cell Layer, in: A. Altman, S. Izhar and M. Ziv (Eds.), Plant Biotechnology andin Vitro Biology in the 21st Century. Proceedings of the IX International Congress on Plant Tissue and Cell Culture, 1998. [50] Y. Murakami, M. Tsuyama, Y. Kobayashi, H.
Ko-dama, K. Iba, Trienoic fatty acids and plant tolerance of high temperature, Science 287 (2000) 476 – 479.