L
Journal of Experimental Marine Biology and Ecology 245 (2000) 111–125
www.elsevier.nl / locate / jembe
Effects of increasing current velocity, turbidity and
particle-size selection on the feeding activity and scope for growth
of Ruditapes decussatus from Ria Formosa, southern
Portugal
a ,* b
P. Sobral , J. Widdows
a
ˆ
Departamento de Ciencias e Engenharia do Ambiente, FCT, Universidade Nova de Lisboa, Quinta da
Torre, 2825-114 Monte Caparica, Portugal
b
Centre for Coastal Marine Sciences, Plymouth Marine Laboratory, Prospect Place, The Hoe, Plymouth,
PL1 3DH, UK
Received 14 April 1999; received in revised form 7 October 1999; accepted 18 October 1999
Abstract
This study examines the feeding activity of the infaunal bivalve Ruditapes decussatus L., in relation to increasing current velocities and concentrations of suspended particulate matter (SPM).
21
An annular flume generated free-stream velocities of 0.6, 3, 8, 17, 34 and 36 cm s (estimated shear stress ranging from 0.001 to 3.8 Pa) and feeding rate was measured in terms of algal cell
21 21
depletion (clearance rate) in the water column. Maximum clearance rate, ca. 2.5 l h ind for 21
standard 0.3 g dry tissue mass individuals, occurred at the lower current velocities, up to 8 cm s 21
and declined with increasing velocities, specially above 17 cm s when shear stresses were 21 .0.89 Pa and sediment was eroding and moving. At low current velocities (,8 cm s ) suspension feeding caused a significant depletion in the algal concentration in the water column
21
within 10 cm of the bed. At very low current velocities (,1 cm s ) the vertical profile showed that the lowest algal concentrations occurred at a height of 10 cm. Therefore the inhalent and exhalent siphons of the clam draw in and eject water at different heights in the water column, thus minimising the intake / recirculation of algal cell depleted water This strategy allows the maintenance of a higher filtration rate and scope for growth when compared with suspension feeders where inhalent and exhalent water currents are at the same height above the bed. At slow current velocities R. decussatus therefore maintains high feeding rates, in part by regulating the distribution of algal cells in the water column. There was a marked decline in clearance rates of clams with increasing concentration of suspended particulate matter, and this was accompanied by copious mucus production. Small particles (3–7mm) were retained at a lower efficiency at lower
21
SPM concentrations (ten compared to 100 mg l ) probably reflecting adaptation to a turbid
*Corresponding author. Tel.: 1351-21-294-8500; fax: 1351-21-294-8554.
E-mail address: [email protected] (P. Sobral)
natural environment containing a mixture of phytoplankton, suspended benthic algal cells and silt.
2000 Elsevier Science B.V. All rights reserved.
Keywords: Current velocity; Feeding activity; Particle selection; R. decussatus; Scope for growth; Turbidity
1. Introduction
Water currents are essential for the supply of suspended food particles, including phytoplankton, microphytobenthic organisms and resuspended sediment particles to sessile suspension-feeding bivalves. Current velocity influence sediment dynamics, as well as settlement of the larval stage, and the feeding and growth of adult benthic animals (Kirby-Smith, 1972; Walne, 1972; Wildish and Peer, 1983; Wildish and Kristmanson, 1997).
Furthermore, hydrodynamics are important in determining the settlement of bivalve ´
larvae (Andre et al., 1993) and thus recruitment of species to the benthos. However, relatively few studies have examined the performance of marine organisms in response to a wide range of current velocities. Wildish et al. (1987, 1992), Wildish and Kristmanson (1985) and Wildish and Miyares (1990) have examined the feeding responses to flow-rate in Mytilus edulis and Placopecten magellanicus. The influence of flow velocity on growth was studied on Argopecten radians by Cahalan et al. (1989), on Mercenaria mercenaria and Crassostrea virginica by Grizzle et al. (1992) and on Modiolus modiolus by Lesser et al. (1994).
˜
In the Ria Formosa lagoon system, freshwater input is minimal (Falcao and Vale, 1990), and the tidal water movement is the principal process generating currents. Tidal processes together with storm generated waves and currents have a major role in the evolution of the barrier islands of the Ria Formosa system (Pilkey et al., 1989).
The lagoon is largely filled with an accumulation of fine sediment in salt marshes and the tidal-delta sands. This fine sediment is readily resuspended by tidal currents and storm conditions and therefore turbidity and the concentration of suspended particulate matter in estuarine and coastal systems show tidal, seasonal and inter-annual variation. R. decussatus an infaunal bivalve of tidal flats in the Ria Formosa, is consequently exposed to wide variations in both water currents and suspended particulate matter (SPM). In this dynamic system resuspension of sediment is a common feature and the ability of clams to filter and select the appropriate food particles will determine their performance and growth. Several studies indicate that suspended bottom material is an important food source for suspension feeding bivalves such as M. edulis (Kiørboe et al., 1980, 1981; Bayne et al., 1987), Spisula subtruncata (Møhlenberg and Kiørboe, 1981) and Ostrea edulis (Grant et al., 1990).
2. Materials and methods
Clams were collected at the sampling site in Ria Formosa at low tide, and allowed to acclimatise to laboratory conditions (temperature 2008C, salinity 32‰) at the Plymouth Marine Laboratory (PML), UK, where all the experiments were performed. Clams were
3
maintained in a system containing 3 m of recirculating seawater and they were fed with 21
a culture of Isochrysis galbana at a rate of approximately 3000 cells ml .
2.1. Effect of current velocity
Current velocities were generated using an annular flume (Fig. 1), for details see Widdows et al. (1998a). The flume has a 10-cm wide circular channel of 64-cm outer diameter, a maximum water depth of 35 cm, and a maximum volume of 60 l. The rotating smooth drive plate on the water surface can create free stream velocities up to
21
80 cm s . Water can be sampled at up to seven different heights above the bed via 1-mm bore tubes connected to syringes.
A small electromagnetic (EM) current sensor (Valeport 800–175) was used to quantify the free-stream velocities (at 10 cm height) and velocity-height profiles (2 to 16 cm above the bed) from which to estimate the shear stress on the bed (Pa). These vertical profiles were also confirmed by video tracking of neutrally buoyant particles close to the bed (Widdows, unpublished observations).
Sediment was introduced in the flume to a depth of 5 cm and the clams allowed to bury overnight prior to each experiment. The sandy sediment (364-mm mean diameter
615 S.E.) was collected from Whitsand Bay, Cornwall, and washed several times under running water.
Four groups (n513 clams per group; mean length 34.660.2 mm, n552) were buried in the sediment within the quadrant just upstream of the 1-mm bore sampling tubes for measuring the vertical profile in the water column. Algal cells were introduced
Fig. 1. (A) Annular flume used to study the effects of current velocity on the clearance rates of Ruditapes
21 into the flume in order to obtain a concentration of approximately 15 000 cells ml . The feeding rates of clams were then measured at six current velocities of 0.6, 3, 8, 17, 34
21
and 36 cm s (equivalent to bed shear stresses of 0.001, 0.02, 0.11, 0.89, 1.68 and 3.8 Pa, respectively).
In each experimental run, 30 min after addition of the algal cells, two water samples separated by a 2-min interval, were collected for algal cells counts at three different heights (2, 10 and 20 cm above the sediment), and this was repeated every 30 min for 2 h. Cells were counted with a Coulter Counter Model D, equipped with a 140-mm aperture tube. Clearance rates, or the volume of water cleared of suspended particles, were calculated from the exponential decline in algal cell concentrations (for details see Widdows et al., 1998a). This was based on the concentrations at the upper height, which reflected the longterm mixed algal cell concentrations in the flume. Samples from lower levels reflected small scale cell depletion at lower current velocities.
21 Controls, without clams, were run with the addition of algal cells, at 0.6 cm s and at
21
24 cm s . Algal settlement in the controls was very low (equivalent to a clearance rate 21
of ,0.25 l h ) and was subtracted from clearance rate values for each group of clams. Clearance rates values were standardised to an animal of 0.3 g dry tissue mass.
During each experimental run at a specific current velocity water samples were collected from all six heights (2, 5, 10, 15, 20 and 25 cm above the sediment). This provided a vertical profile of cell concentrations in order to determine algal cell depletion in the water column in relation to current velocity. Control counts (without
21 animals), for these profiles were again made at free stream velocities of 0.6 cm s and
21 24 cm s .
2.2. Turbidity
To test the influence of turbidity on the clearance rate of R. decussatus, three SPM 21
concentrations, 10, 100 and 300 mg l were used. These were obtained by adding fine surface mud (mean diameter 12 mm) to seawater to make up the required SPM concentration. Mud was collected from the banks of the Lynher river, at Whacker Quay, Cornwall, and then held at 58C and in the dark until required for the experiment.
SPM concentrations were monitored with an optical back-scatter probe (OBS-3 D&A Instrument), previously calibrated for the same mud. Calibration was achieved by fitting a regression line to the relationship between OBS readings (volts) and the concentration
21
of suspended material (mg l ) in samples taken for gravimetric analysis on GF / C filters.
The turbidity experiments were performed in a closed system consisting of sixteen 2-l beakers each containing one clam, plus one control beaker without an animal. These were placed on a multi-point magnetic stirrer. A mud suspension was added to each beaker to the required concentration and turbidity level monitored with the OBS sensor. Algal cells were also added to the fine sediment to provide a concentration of 10 000
21
At 30-min intervals over a period of 1 h 30 min, one 20-ml sample was taken from each beaker with a syringe and the volume of particles measured by means of a Coulter Counter Multisizer fitted with a 100-mm orifice tube. At the two highest seston concentrations dilutions (1:10 and 1:20) were made to avoid coincidence counts and blockage of the orifice tube. Samples were always stirred prior to counting.
Clearance rates, minus control values, were calculated over two consecutive time intervals and weight standardised to a 0.3 g dry weight animal.
2.3. Particle size selection
The suspended particle-size selection experiments were performed using the flume 21 and 13 animals buried in sediment. Two SPM concentrations (10 and 100 mg l ) were
21 studied. These concentrations were obtained and algal cells (10 000 cells ml ) were added in the same manner as for the turbidity experiments.
Samples were taken from two heights in the water column, measured (by volume), over a wide range of channels / particle sizes (2.4 to 9.6 mm diameter) with a Coulter Counter Multisizer fitted with a 100-mm orifice tube and averaged. Dilution (1:10) was needed at the higher seston concentration to avoid coincidence counts.
Size selection was quantified by examining the relative depletion of different particle sizes over a period of 1 h 30 min. The retention efficiency was calculated per channel using the formula 12(V /V ), V , and V being the volumes measured at two different2 1 1 2 sampling times (i.e. at the beginning and after 1 h 30 min), and then expressed as the percentage of the larger channels / particle sizes (i.e. 9.6-mm equivalent spherical diameter) were maximum volumes registered. For this calculation only particles of diameters between 2.4mm to 9.6mm were considered because larger particles introduce considerable variability into the counts, due to their relative high volumes and relatively low numbers.
3. Results
3.1. Effect of current velocity on clearance rate and sediment movement
21 21
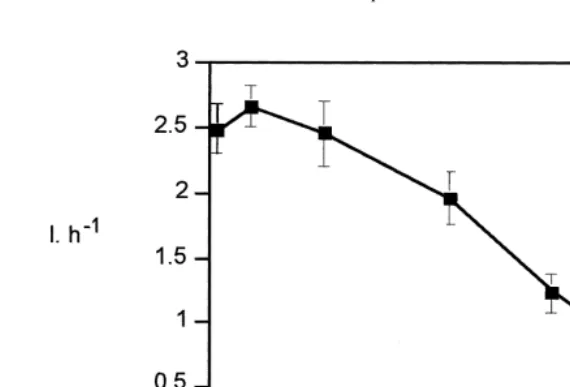
Fig. 2 shows that the maximum clearance rate (c. 2.5 l h ind ) occurs at the lower 21
current velocities (i.e. below 8 cm s ) and declines with increasing velocities,
21 21
particularly above 17 cm s (i.e. shear stresses .0.9 Pa). At velocities of 24 cm s the clearance rate declines to c. 50%. A significant linear relation between clearance
21 21
rates (l h ) and current velocity (cm s ) is explained by the equation y5 20.066x1
2.828, with r50.98 (n56).
The current velocity and the shear stress needed to induce bed movement are clearly 21 related to sediment granulometry. At free stream current velocities ,12 cm s , or bed shear stresses of less than c. 0.4 Pa there was no movement of surface sediment
21
particles. At 17 cm s (0.89 Pa) some resuspension of faeces was observed, together 21
21 21
Fig. 2. Variation of clearance rate (l h ind 6S.E.) of Ruditapes decussatus (0.3 g dw) with current velocity 21
(cm s ) in the annular flume.
saltation of sand grains on the bottom leading to the formation of small slowly moving sand ripples. At this current velocity the siphons were being bombarded by particles and the clams responded by occasional ‘coughing’ or ejection of material from the mantle
21
cavity via the inhalant siphon. At a free stream velocity of 36 cm s (3.8 Pa), there was continuous resuspension of sediment, migrating sand ripples and movement of the bed sediment, which kept covering and uncovering the clams. Two individuals could not stay in position and were transported to the next quadrant of the flume. At this high current velocity clams were experiencing continuous bombardment of the siphons with sand grains and some would keep them constricted at the tip while others would simply close their valves. On several occasions the inhalant siphons were observed to eject sediment higher than 5 cm into the water column. At the end of the experimental runs, strings of mucus bound sediment particles (i.e. material rejected as pseudofeces) were observed on the sediment surface.
3.2. Effect of current velocity on algal cell depletion in the water column
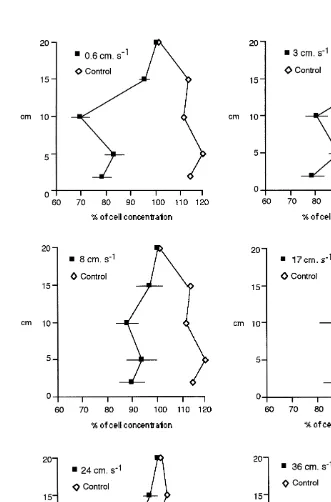
Vertical profiles of cell concentrations in the water column at different free-stream velocities are shown in Fig. 3. In all flume runs with clams present and at all current velocities, the maximum cell concentration occurred at a height of 20 cm above the bed. Consequently, all cell concentrations are expressed as a percentage of the cell concentration at 20-cm height. The results presented in Fig. 3 illustrate that algal depletion near the bed was more evident at the lower current velocities, especially at 0.6
21
Fig. 3. Vertical profiles of algal cell concentrations (mean6S.E.) after depletion by Ruditapes decussatus (j) 21
from the control profiles, reflecting the lower clearance rates and the greater vertical mixing.
21
At the lowest current velocity measured (0.6 cm s ) the algal cell depletion was greatest at 10 cm above the sediment surface (Fig. 3) but this was less pronounced at 3
21 21
cm s and was beginning to disappear by 8 cm s . Algal cell depletion at 5 and at 21
10-cm height at 0.6 and 3 cm s is significantly different from the cell concentrations at 21
15 cm (t-test, P,0.01 for 10 cm at 0.6 cm s and P,0.05 for the others).
At higher current velocities differences in cell concentrations at 5 cm and 15 cm are not significant. Algal cell concentrations found at 10 cm are significantly different (t-test, P,0.05) from the cell concentrations found higher (15 and 20 cm) in the water column
21
at all current velocities except at 17 cm s where differences where not significant due to higher variability (Fig. 3).
4. Effect of turbidity on clearance rate and particle-size selection
There was a marked decline in clearance rates by clams with increasing SPM 21
concentration from 10 to 300 mg l (Fig. 4). This decline shows a significant linear relationship for the range tested and is fitted by the equation y5 20.003x11.426, with
21
r50.99 (n53). At concentrations of 100 mg SPM l and above, visual monitoring of clam behaviour was not possible due to the high turbidity. However, at the end of the experiment the presence of mucus-containing particles was observed on the foot, mantle edge and on the bottom of the beakers.
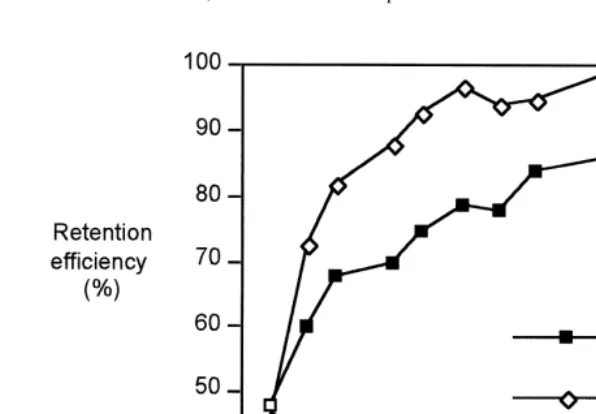
Fig. 5 shows that particles smaller than 3 mm are not retained efficiently (i.e.
21 21
Fig. 4. Influence of suspended particulate material and turbidity (mg l ) on the clearance rate (l h 6S.E.) of
Fig. 5. Variation of retention efficiency (%) with particle size by Ruditapes decussatus in response to two SPM concentrations (algal cells added).
,60–70% retention), and that particle retention efficiency by the gill increases with increasing particle diameter. Algal cells .6 mm diameter were filtered with ca. 80–100% efficiency. The particle-size selection efficiency by the clams is different for
21
the two SPM concentrations tested (Fig. 5). The 100-mg SPM l curve shows the typical relationship, with maximum retention efficiency occurring at particle sizes
21
.4–5 mm. At the lower concentration of 10 mg SPM l , the retention of smaller particles was less effective below ,8mm.
5. Discussion
5.1. Current velocity
The results of this study demonstrate that current velocity has a marked effect on the clearance or feeding rates of R. decussatus and that this species follows the general pattern of response observed for other bivalves. Previous studies have recorded a similar gradual decline in the suspension feeding and / or growth of bivalves with increasing
21
current velocity from 1 to .15 cm s (mussel M. edulis) (Wildish and Miyares, 1990), giant scallop Placopecten magellanicus (Wildish et al., 1987, 1992), bay scallop Argopecten irradians (Kirby-Smith, 1972; Cahalan et al., 1989). Grizzle et al. (1992) found a positive correlation between flow speed and growth in Mercenaria mercenaria
21 21
for speeds of 2–8 cm s with maximum growth at 2–4 cm s and decreased growth in 21
Crassostrea virginica at low speeds (1 cm s ).
21
occur at the lower current velocities (i.e. ,8 cm s ; Fig. 2). These experimental findings are in agreement with field observations of clams living on intertidal banks in
21
enclosed sheltered areas, where currents are not strong (i.e. ,12 cm s ) and just below the threshold for erosion of sandy sediments (Sobral and Widdows, unpublished data). As the clams live buried with their siphons open at the sediment surface, any currents that can resuspend small organic particles, particularly the microphytobenthos, will have a beneficial effect. In contrast, the suspension of large sediment particles (i.e. sand grains) will have an inhibitory effect on their feeding rate and valve gape due to the physical disturbance (i.e. bombardment, displacement of substrate and impairment of gill function). Consequently, as current velocity increases, both in the laboratory and in the field, the decline in clearance rate is a response to the combined effect of increased shear stress, turbidity and physical disturbance. The clams were able to cope with the mobile sediment by maintaining their position at the sediment surface and maintaining some
21
pumping activity. At 36 cm s , the highest current velocity tested, clearance rate is reduced to c. 10% of the maximum value, but this was still sufficient to meet the animal’s oxygen requirements. For example, such low clearance or ventilation rates will
21 21
provide oxygen at the rate of c. 50 mmoles O h2 (i.e. 0.25 l h 3200mmoles O2 21
l ), which is in excess of the routine rate of oxygen consumption by R. decussatus (16 21
mmoles O h2 ; Sobral and Widdows, 1997a).
This study has not only recorded significant algal cell depletion in the water column 21
above the clams at low current velocities (,8 cm s ), but it has also provided evidence of further structure to the vertical profile (Fig. 3). Algal cell or seston depletion at low current speeds is a well established phenomenon over mussel beds, as a result of a faster rate of removal by suspension feeders than replenishment via vertical diffuse
´
transport (e.g. Mytilus edulis, Frechette et al., 1989; Newell and Shumway, 1993). However, the present study with siphonate clams has provided new evidence of enhanced algal depletion at a height of 10 cm above the bed. A similar situation has been recorded for the siphonate cockle Cerastoderma edule at low current velocities (Navarro and Widdows, unpublished data). This phenomenon is explained by the jetting of algal cell depleted water from the exhalent siphon to a height of ca. 10 cm above the bed. Furthermore, this structure in the water column is maintained due to the lower turbulence and vertical mixing associated with infaunal bivalves buried in sediment with
´
a relatively smooth surface. Andre et al. (1993) described the hydrodynamics and flow patterns produced by C. edule of similar size. They showed that jetting from the exhalent
21
siphon can be as high as 7 cm at current velocities of 1 cm s , similar to those used in the present study. The ability to expel food depleted water at a different level to that used by the inhalant current is likely to be an important adaptation by clams and cockles living in sheltered environments with low current speeds. This situation is in contrast to epifaunal mussels (M. edulis), which cause algal cell depletion immediately above the mussel bed. Mussels do not maintain a specific orientation to the bed or the current and consequently have both their inhalent and exhalent openings at various levels. Such a lack of consistent orientation may result in a degree refiltering of depleted water and
21
thus reduced filtration rates (mg h ), under conditions of low currents (Widdows et al., 1998b and unpublished observations).
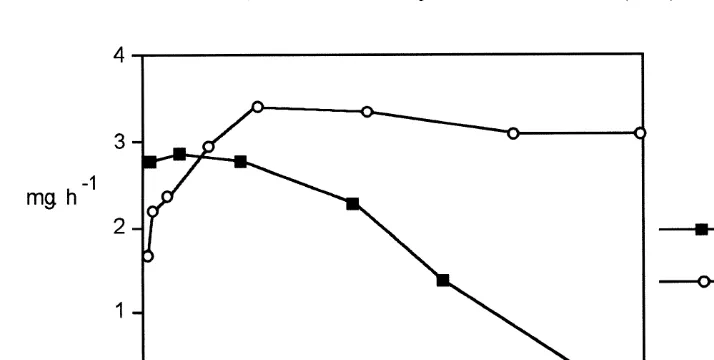
21 21
Fig. 6. Filtration rates of Ruditapes decussatus and Mytilus edulis (mg h ind ) for different current 21
velocities at the same SPM concentration (ca. 1 mg l ).
21
filtration rate at the lower current velocities (,8 cm s ), in comparison to the minimal decline in filtration rate by R. decussatus. This species difference is due to the more marked separation of inhalent and exhalent water by R. decussatus with the clam pumping from less depleted water. Clams maintain high filtration rates over a wide range
21
of low current velocities (e.g. ,10 cm s ), whereas mussels, 4 cm length, in the same flume conditions reported in this study, maintain a constant clearance and filtration rate
21
over a wide range of higher current velocities up to .40 cm s (Widdows, 21 unpublished data). In fact mussel larvae are able to settle at flow velocities ,1.2 m s and can be found in cooling effluent pipes where flow velocities are strong (1.5–2.5
21
m s ) as reported by several authors in Rajagopal et al. (1996). Therefore clams appear to be better adapted to sheltered environments with low currents, whereas mussels appear to be better adapted to conditions of higher current velocities thus avoiding seston depletion and refiltering of depleted water.
Scope for growth values calculated for the two species at a current velocity of ca. 0.6 21
cm s are shown in Table 1. These values were calculated using food ingestion rates 21
Table 1
21
Comparison between energy budgets and scope for growth (J h ) of Ruditapes decussatus with: (1) water intake and output at different levels (natural situation) and (2) water intake and output at the same level, and (3) Mytilus edulis (0.3 g dw) water intake and output at approximately same level (natural situation) at a
21 21 a
current velocity of ca. 0.6 cm s and a 100% algal ration of 1 mg l
Species CR C AE A R SFG
21 21 21 21 21
(l h ) (J h ) (%) (J h ) (J h ) (J h )
R. decussatus
(1) Intake and output at different levels 2.4 43.1 43 18.5 3.6 14.9
R. decussatus
(2) Intake and output at same level 2.4 30.4 43 12.1 3.6 8.5
M. edulis 1.8 32.5 40 13.0 2.1 10.9
a
CR, clearance rate; C, energy consumed; AE, absorption efficiency; A, energy gain; R, energy loss through respiration; SFG, scope for growth.
5.2. Turbidity and particle-size selection
This study shows that the clearance rates of R. decussatus decline with increasing SPM (Fig. 4). These findings are in agreement with other studies in the literature for M. edulis, C. edule and Venerupis pullastra, filtering purely algal suspensions, (Foster-Smith, 1975); for M. edulis feeding on resuspended fine mud, (Widdows et al., 1979); for O. edulis, (Grant et al., 1990); for Mya arenaria, (Grant and Thorpe, 1991); for C. edule, (Iglesias et al., 1992; Navarro and Widdows, 1997), all feeding on mixtures of algal cells and suspended silt.
21
Filtration rate (mg h ) by R. decussatus, based on the product of clearance rate and SPM concentration, has been calculated for two different scenarios (Fig. 7). Firstly, filtration rates have been calculated from the regression of clearance rates at different
21
current velocities (Fig. 2) for a constant SPM of 20 mg l (mean SPM at the sediment
21
Fig. 7. Relationship between filtration rate (FR, mg h ) of Ruditapes decussatus and current velocity for two
21 21
surface by an OBS probe at the field sampling site in June 1995; Sobral and Widdows, 21 unpublished data). At a constant seston concentration, filtration rate (mg h ) is
21 21
maintained below 8 cm s , but declines at current velocities .17 cm s and ceases at 21
current velocities of ca. 43 cm s . Secondly, in an environment where there is significant erosion and resuspension of sediment and microphytobenthos with increasing current velocity, filtration increases with resuspension up to SPM concentrations of ca.
21
100 mg l . Filtration rate then declines with increasing concentration as a result of lower clearance rates under high SPM conditions, through the overloading of the gill suspension feeding mechanisms. Under these prevailing SPM conditions, feeding is
21 21
inhibited at ca. 480 mg l at a corresponding current velocity of 27 cm s .
Jørgensen (1981) observed that mucociliary mechanisms in M. edulis serve to clean the gills and other mantle cavity surfaces of excess particulate material. This was also recorded in R. decussatus when filtering at the highest SPM concentration tested (300
21
mg l ). While production of mucus is part of the normal feeding process, excessive production in response to high SPM levels may be detrimental and is accompanied by a
21 21
low clearance rate (i.e. ,0.5 l h ). These observations indicate that high SPM concentrations are capable of lowering the trophic performance of clams.
The addition of low concentrations of silt to algal cell suspensions, which more closely resemble natural seston, has been found to enhance the clearance rates of M. edulis (Kiørboe et al., 1980, 1981; Bayne et al., 1987) and V. corrugatus (Stenton-Dozey and Brown, 1994). In the present study, however, when R. decussatus was fed at the
21
lowest resuspended silt1algal cell concentration (10 mg l ) the clearance rate of R. 21
decussatus was lower (c. 1.4 l h ; Fig. 4) than when feeding on algal cells only (c. 2.5 21
l h ; Fig. 2). Similar high values were recorded when measuring the clearance of algal cell from suspension in beakers without sediment (Sobral and Widdows, 1997a,b). When clams are buried in sediment, as in the flume, and thus under environmental conditions, clearance rates may be slightly higher (Fig. 2). However, an initial study, showed that there were no significant differences between clearance rates of clams held in or out of sediment and measured in beakers. The difference found here may be due to the longer recovery time following handling disturbance in the flume experiments (i.e. overnight), compared to the shorter recovery time in the beaker experiments (2 h).
Particle-size selection by R. decussatus (Fig. 5) shows that particles smaller than 3
mm in diameter, which includes bacteria and clay particles, are retained with low efficiency (i.e. ,75%). Algal cells, such as phytoplankton, which play an important role in the nutrition of the clams, and other particles in the range from 3 to 8mm are efficiently retained (70 to 100% retention).
The pattern of particle retention efficiency shown by R. decussatus at the higher 21
seston concentration (100 mg l ) ensures the effective retention of particles in the 21
range 327mm (Fig. 5). This more effective particle retention at 100 mg l compared 21
con-centrations. A. irradians was found to increase the retention efficiency of smaller particles (,4mm) with increasing seston concentrations, by trapping particles through the production of mucus, probably in the same way as R. decussatus. On the contrary C. virginica is less efficient in retaining small particles with increasing seston con-centrations. Baldwin (1995) also found that larvae of C. virginica preferably select particles .10 mm when fed complex natural suspensions.
The pattern of retention efficiency for particles of $4mm by R. decussatus agrees
˚
with the results for 13 bivalve species (Møhlenberg and Riisgard, 1978). In the lower range (particles ,4 mm) this study recorded some variability in retention efficiency between species, probably reflecting size distribution of SPM in the natural environment. 0. edulis and C. edule common in estuarine areas where fine sediment occurs, both revealed a steep decline in retention efficiency for particles below 3 mm diameter, similar to the relationship found for R. decussatus. This may represent an adaptation of the clams to a richer and more turbid environment, in contrast to epifaunal bivalves which are better adapted to open coastal waters with lower particles concentrations. [RW]
References
Andre, C., Jonsson, P.R., Lindegarth, M., 1993. Predation on settling bivalve larvae by benthic suspension feeders: the role of hydrodynamics and larval behaviour. Mar. Ecol. Prog. Ser. 97, 183–192.
Baldwin, B.S., 1995. Selective particle ingestion by oyster larvae (Crassostrea virginica) feeding on natural seston and cultured algae. Mar. Biol. 123, 95–107.
Bayne, B.L., Hawkins, A.J.S., Navarro, E., 1987. Feeding and digestion by the mussel Mytilus edulis L. (Bivalvia: Mollusca) in mixtures of silt and algal cells at low concentrations. J. Exp. Mar. Biol. Ecol. 111, 1–22.
Cahalan, J.A., Siddall, S.E., Luckenbach, M.W., 1989. Effects of flow velocity, food concentration and particle flux on growth rates of juvenile bay scallops Argopecten irradians. J. Exp. Mar. Biol. Ecol. 129, 45–60.
˜
Falcao, M., Vale, C., 1990. Study of the Ria Formosa ecosystem: benthic nutrient remineralization and tidal variability of nutrients in the water. Hydrobiologia 207, 137–146.
Foster-Smith, R.L., 1975. The effect of concentration of suspension on the filtration rates and pseudofaecal production for Mytilus edulis L., Cerastoderma edule (L.) and Venerupis pullastra (Montagu). J. Exp. Mar. Biol. Ecol. 17, 1–22.
´
Frechette, M., Butman, C.A., Geyer, W.R., 1989. The importance of boundary layer flows in supplying phytoplankton to the benthic suspension feeder Mytilus edulis L. Limnol. Oceanogr. 34, 19–36. Grant, J., Enright, C.T., Griswold, A., 1990. Resuspension and growth of Ostrea edulis: a field experiment.
Mar. Biol. 104, 51–59.
Grant, J., Thorpe, B., 1991. Effects of suspended sediment on growth, respiration and excretion of the soft-shell clam (Mya arenaria). Can. J. Fish. Aquat. Sci. 48 (7), 1285–1292.
Grizzle, R.E., Langan, R., Howell, W.H., 1992. Growth responses of suspension-feeding bivalve molluscs to changes in water flow: differences between siphonate and nonsiphonate taxa. J. Exp. Mar. Biol. Ecol. 162, 213–228.
Iglesias, J.I.P., Navarro, E., Alvarez Jorna, P., Armentia, I., 1992. Feeding, particle selection and absorption in cockles Cerastoderma edule (L.) exposed to variable conditions of food concentration and quality. J. Exp. Mar. Biol. Ecol. 162, 177–198.
Jørgensen, C.B., 1981. Feeding and cleaning mechanisms in the suspension feeding bivalve Mytilus edulis. Mar. Biol. 65, 159–163.
Kiørboe, T., Møhlenberg, F., Nøhr, O., 1980. Feeding, particle selection and carbon absorption in Mytilus
Kiørboe, T., Møhlenberg, F., Nøhr, O., 1981. Effect of suspended bottom material on growth and energetics in
Mytilus edulis. Mar. Biol. 61, 283–288.
Kirby-Smith, W., 1972. Growth of the bay scallop: the influence of experimental currents. J. Exp. Mar. Biol. Ecol. 8, 7–18.
Lesser, M.P., Witman, J.D., Sebens, K.P., 1994. Effects of flow and seston availability on scope for growth of benthic suspension-feeding invertebrates from the Gulf of Maine. Biol. Bull. 187, 319–335.
˚
Møhlenberg, F., Riisgard, H.U., 1978. Efficiency of particle retention in 13 species of suspension feeding bivalves. Ophelia 17 (2), 239–246.
Møhlenberg, F., Kiørboe, T., 1981. Growth and energetics in Spisula subtruncata (Da Costa) and the effect of suspended bottom material. Ophelia 20, 79–90.
Navarro, J.M., Widdows, J., 1997. Feeding physiology of Cerastodermaedule in response to a wide range of seston concentrations. Mar. Ecol. Prog. Ser. 152, 175–186.
Newell, C.R., Shumway, S.E., 1993. Grazing of natural particulates by bivalve molluscs: a spatial and temporal perspective. NATO ASI Ser. G33, 85–148.
Palmer, R.E., Williams, L.G., 1980. Effect of particle concentration on filtration efficiency of the bay scallop
Argopecten irradians and the oyster Crassostrea virginica. Ophelia 19 (2), 163–174.
Pilkey, O.H., Neal, W.J., Monteiro, J.H., Dias, J.M.A., 1989. Algarve baffler islands: A noncoastal-plain system in Portugal. J. Coastal Res. 5 (2), 239–261.
Rajagopal, S., Nair, K.V.K., Azariah, J., van der Velde, G., Jemmer, H.A., 1996. Chlorination and mussel control in the cooling conduits of a tropical coastal power station. Mar. Env. Res. 41 (2), 201–221. Sobral, P., Widdows, J., 1997a. The influence of hypoxia and anoxia on the physiological responses of the clam
Ruditapes decussatus L. from southern Portugal. Mar. Biol. 127, 455–461.
Sobral, P., Widdows, J., 1997b. Effects of elevated temperatures on the physiological responses and resistance to air exposure of the clam Ruditapes decussatus, from southern Portugal. Scientia Marina 61, 163–171. Stenton-Dozey, J.M.E., Brown, A.C., 1994. Short-term changes in the energy balance of Venerupis corrugatus (Bivalvia) in relation to tidal availability of natural suspended particles. Mar. Ecol. Progr. Ser. 103, 57–64. Walne, P.R., 1972. The influence of current speed, body size and water temperature on the filtration rates of
five species of bivalves. J. Mar. Biol. Assoc. UK 52, 345–374.
Widdows, J., Fieth, P., Worrall, C.M., 1979. Relationships between seston available food and feeding activity in the common mussel Mytilus edulis. Mar. Biol. 50, 195–207.
Widdows, J., Brinsley, M.D., Bowley, N., Barrett, C., 1998a. A benthic annular flume for in situ measurement of suspension feeding / biodeposition rates and erosion potential of intertidal cohesive sediments. Est. Coastal Shelf Sci. 46, 27–38.
Widdows, J., Brinsley, M.D., Salkeld, P., Elliott, M., 1998b. Use of annular flumes to determine the influence of current velocity and bivalves on material flux at the sediment-water interface. Estuaries 21 (4A), 552–559.
Wildish, D.J., Kristmanson, D.D., 1985. Control of suspension feeding bivalve production by current speed. Helgolander Meeresunters 39, 237–243.
Wildish, D.J., Kristmanson, D.D., 1997. Benthic Suspension Feeders and Flow, Cambridge University Press, UK.
Wildish, D.J., Miyares, M.P., 1990. Filtration rates of blue mussels as a function of flow velocity: preliminary experiments. J. Exp. Mar. Biol. Ecol. 142, 213–219.
Wildish, D.J., Peer, D., 1983. Tidal current speed and production of benthic macrofauna in the lower Bay of Fundy. Can. J. Fish. Aquat. Sci. 40 (suppl. 1), 309–321.
Wildish, D.J., Kristmanson, D.D., Hoar, R.L., DeCoste, A.M., McCormick, S.D., White, A.W., 1987. Giant scallop feeding and growth responses to flow. J. Exp. Mar. Biol. Ecol. 113, 207–220.