C3H apoE(
− / −
) mice have less atherosclerosis than C57BL
apoE(
− / −
) mice despite having a more atherogenic serum lipid
profile
D.C. Grimsditch
a,*, S. Penfold
a, J. Latcham
b, M. Vidgeon-Hart
c, P.H.E. Groot
a,
G.M. Benson
aaDepartment of Vascular Biology,SmithKline Beecham Pharmaceuticals,New Frontiers Science Park (North),Third A6enue,Harlow,Essex, CM19 5AD, UK
bDepartment of Laboratory Animal Sciences,SmithKline Beecham Pharmaceuticals,The Frythe,Welwyn,Herts AL6 9AR, UK cDepartment of Safety Assessment,SmithKline Beecham Pharmaceuticals,The Frythe,Welwyn,Herts AL6 9AR, UK
Received 29 March 1999; received in revised form 17 September 1999; accepted 23 September 1999
Abstract
Wild-type C57BL mice are known to be susceptible to diet-induced atherosclerosis, whilst C3H mice are resistant. We investigated the effect of these background strains on the hyperlipidaemia and atherosclerosis that develops in mice deficient in apolipoprotein E (apoE(− / −)). Male and female apoE(− / −) mice on C3H/HeNHsd (C3H) and C57BL/6J (C57) backgrounds were fed atherogenic Western diet for 12 weeks. Serum cholesterol and triglyceride concentrations were measured and atherosclerosis quantified in the aortic sinus. C3H apoE(− / −) mice fed normal diet had 1.5 – 2 fold higher serum cholesterol levels than C57 apoE(− / −) mice and 4 – 5 fold higher serum triglyceride concentrations. Feeding Western diet caused a 4 – 5 fold increase in serum cholesterol in all mice, but levels of triglyceride were either attenuated or were unaffected in C3H apoE(− / −) and C57 apoE(− / −) mice, respectively. C3H apoE(− / −) mice had approximately 2 fold higher serum cholesterol and 4 fold higher triglyceride concentrations than the C57 apoE(− / −) mice throughout the study. Serum triglyceride concentrations were 35 – 108% higher in male C3H apoE(− / −) than female C3H apoE(− / −) mice. Most of the lipids were present in the very low density lipoprotein (VLDL)/chylomicron fraction in both strains of mice whether they were fed normal or Western diet. Notwithstanding the lower plasma lipid concentrations, atherosclerotic lesion areas were more than 2-fold larger in C57 apoE(− / −) than in C3H apoE(− / −) mice (males 68911×103vs 3096×103, females 102912×103vs 4198×103mm2,
mean9SEM). © 2000 Elsevier Science Ireland Ltd. All rights reserved.
Keywords:Atherosclerosis; C3H; C57BL; Female; Lesion area; Male; Western diet
www.elsevier.com/locate/atherosclerosis
1. Introduction
Inbred normal mice are relatively resistant to the development of diet-induced atherosclerosis, and rather extreme diets, enriched in saturated fat, cholesterol and bile salts have to be used to initiate this process. Notwithstanding this disadvantage, the availability of
numerous genetically well-defined inbred strains has made the mouse an attractive species in which to search for genes linked with susceptibility for developing atherosclerosis. The work of Paigen et al. [1,2] on many strains of inbred mice has established ranking in the susceptibility for atherosclerosis as follows: C57BR/
cdJ\C57L/J\SM/J\C57BL/6J\SWR/J\C58/J\ 129/J\DBA/2J\AKR/J\BALB/c\NZB/BINJ, HRS/J, A/J, C3H/HeJ, SJL/J, CBA/J. Normal C57BL/
6J (C57) mice are, therefore, more susceptible to diet-induced atherosclerosis than C3H/He (C3H) mice. When fed a diet containing approximately 1% choles-terol and 15% fat, C57 mice develop small atheroscle-* Corresponding author. Tel.: +44-1279-627023; fax: +
44-1279-627049.
E-mail address: david –c–[email protected] (D.C. Grims-ditch)
rotic lesions in the aortic root after only 7 weeks, whereas lesions are absent in C3H mice fed the same diet for up to 1 year, although total serum cholesterol concentrations in C3H mice are usually higher than those in C57 mice [1 – 3]. Differences in the susceptibil-ity of these strains to developing atherosclerosis has been attributed to at least eight genes (Ath1 –Ath8), for which the approximate gene loci have been iden-tified [4].
The recent development of technology for gene si-lencing and gene transfer has generated many novel strains of mice with much greater susceptibility to developing atherosclerosis. Among these the knockout of the apoE gene has been shown to give one of the most severe atherogenic phenotypes. ApoE is a major constituent of very low density lipoproteins (VLDL) and chylomicrons and is an important ligand for the receptor mediated uptake of these lipoproteins from the blood into the liver, as reviewed by Hofker et al., [5]. Furthermore, apoE is implicated in HDL metabolism and reverse cholesterol transport. ApoE(− / −) mice were initially produced by two groups [6,7]. The apoE(− / −) mice were subsequently bred into the C57 background and have now been studied extensively. ApoE(− / −) mice have impaired clearance of cholesterol-ester enriched VLDL and chy-lomicrons from the blood which results in hyperlipi-daemia and the development of atherosclerotic lesions [5]. It is reported that apoE(− / −) mice develop spontaneous lesions when fed normal chow diet and this process is further accelerated by feeding a high fat/high cholesterol diet [6,8,9]. Complex lesions that include a fibrous cap can be seen in mice as young as 15-weeks-old and these are not just confined to the aortic sinus but are found in the aortic arch, the major branches of the aorta and the pulmonary and carotid arteries. The atherosclerotic lesions of apoE(− / −) mice exhibit a similar distribution, microscopic ap-pearance and cellular composition to those found in humans [9].
As mentioned above, wild-type C57 mice are rela-tively susceptible to diet-induced atherosclerosis. For the present study we have bred the apoE(− / −) mu-tation into the C3H strain, which has a low suscepti-bility for atherosclerosis, to address the question as to whether the apoE knockout is so dominant in its effect on atherosclerosis that the difference in suscepti-bility between the C57 and C3H strains is lost. Here we report the results of a study in which we compared plasma lipid and lipoprotein concentrations and the amount of atherosclerosis that developed in the aortic root of C57- and C3H apoE(− / −) mice fed a West-ern-type diet. As female mice of the C57 strain are reported to be more susceptible to diet-induced atherosclerosis than males, these studies were per-formed using both sexes [10].
2. Materials and methods
2.1. Materials
Wako cholesterol CII kit (no. 270-54399/54499) was purchased from Wako Chemicals GmbH, Germany. Boehringer-Mannheim (Peridochrom) triglyceride kit (no. 701904) and EDTA (sodium salt) were from Boehringer-Mannheim GmbH, Germany. Pyruvic acid (sodium salt), cholesterol standard solutions, glycerol gelatine (for slide coverslipping) and fumaric acid were from Sigma, USA. Isopropyl alcohol, D-glucose, NaHCO3, KCl and NaCl were from Fisher, UK. Oil
Red O (certistain grade, for lipid staining), dextrin, gelatine (for slide coating), Mayer’s haematoxylin, L-glutamic acid, glycerol (for triglyceride assay stan-dard), NaN3, CaCl2, MgSO4.7H2O and Na2HPO4were
obtained from Merck/BDH, UK. All chemicals were of analytical grade. Buffered formal saline (4% w/w formaldehyde solution) was from Pioneer Research Chemicals, UK. Butterfly-25 cannulae were from Venisystems, Ireland, Microvette CB-300 blood tubes from Sarstedt, Germany, 96-well tissue culture plates from Costar, USA and OCT compound from Bayer Diagnostics, Germany.
2.2. Diet
Normal mouse diet (RM1) was purchased from Spe-cial Diet Services, UK. Western diet was from Hope Farms, The Netherlands. Western diet was essentially the same as that used by Nishina et al. [11] and contained the following constituents (w/w): cocoa but-ter (15%), cholesbut-terol (0.25%), sucrose (40.5%), corn-starch (10%), corn oil (1%), cellulose (5.95%), casein (20%), 50% choline chloride (2%), methionine (0.2%) and mineral mixture (5.1%).
2.3. Mice
ApoE(− / −)tm1Unc
mice [7] were purchased from the Jackson laboratory, having been backcrossed for six generations into a C57BL/6J background and un-dergone three generations of brother/sister mating. The colony underwent a further eight generations of breeding and expansion before being used in the present study. C57BL/6J apoE(− / −) mice were also backcrossed for six generations into C3H/HeNHsd mice. The C3H/HeNHsd apoE(− / −) mice were then bred for two generations before being used in the present study.
2.4. Methods
Male and female apoE(− / −) mice on C3H/
were randomised to eight cages (five per cage) with regard to age (C3H: age=8.1 weeks, C57: age=7.79 0.3 weeks, mean9S.D.) and body weight. Body weights for male and female C3H apoE(− / −) and C57 apoE(− / −) mice were respectively: 25.791.5, 20.991.8, 23.391.8 and 19.491.7 g (mean9S.D.). Mice received food and water ad libitum and were housed in a room that was lit from 06:00 h to 18:00 h and maintained at 21°C. The mice were fed powdered normal diet for 2 weeks prior to receiving the Western diet. All of the mice were weaned onto Western diet over 4 days and were fed the diet for 12 weeks. Body weights were measured at weekly intervals both before and during the study. Food consumption per cage was measured both prior to and throughout the study.
2.5. Serum lipid determination
Blood samples were taken from the tails of non-fasted mice at least 4 h after the start of the light phase when food consumption would be expected to be mini-mal. Blood samples were taken just prior to treatment and at 4 weekly intervals throughout the study. Serum was prepared and aliquots were either frozen (−20°C) awaiting analysis for cholesterol and triglycerides or stored at 4°C for lipoprotein profile analysis. Total cholesterol and total triglyceride concentrations were measured enzymatically, using commercial kits from Wako Chemicals and Boehringer-Mannheim, respec-tively, and a 96-well plate reader (Thermomax, Molecu-lar Devices, UK). In weeks 0, 4, 8 and 12 of the study, equal volumes of serum were pooled from all of the mice in each group for lipoprotein analysis. Lipoprotein profiles were analysed by size-exclusion chromatogra-phy using a SMART™ micro-FPLC system (Pharma-cia, Sweden) and a method similar to that described by Hennes et al. [12]. Briefly, pooled serum samples were filtered (0.2 mm filter, Anotop 10, Whatman, UK) and
30ml of serum was applied to a Superose 6, PC 3.2/30
column. The column was eluted with a solution con-taining NaCl (150 mM), EDTA (1 mM) and sodium azide (7.7 mM) at pH 8.0. The first 920 ml of column
eluent represented the void volume. The lipoproteins were contained in the next 42 fractions of 20 ml each.
The flow rate of the mobile phase was 65 ml/min. The
cholesterol and triglyceride content of the fractions was measured as described above. VLDL/chylomicrons (VLDL/Chy), intermediate/low density lipoproteins (IDL/LDL) and high density lipoproteins (HDL) were present in fractions 1 – 16 (0.92 – 1.24 ml), 17 – 29 (1.24 – 1.5 ml) and 30 – 42 (1.5 – 1.76 ml), respectively. In addi-tion, individual terminal serum samples from five of the mice in each of the four groups were separated into 20-ml fractions and their cholesterol concentration
mea-sured as described above.
2.6. Perfusion fixing of hearts and aortae
The mice were killed by cervical dislocation. The hearts and aortae were immediately perfused in situ with oxygenated Krebs – Henseleit buffer at 37°C under a pressure of approximately 110 cm of water via a cannula inserted in the left ventricle and an outlet created by cutting the right atrium. After 30 min the buffer was replaced with buffered formal saline at 37°C and the perfusion was continued for a further 30 min. The hearts and aortae were then removed, cleaned of extravascular fat and stored in formalin until they were processed.
2.7. Sectioning and staining of the aortic sinus
Cross-sections of the aortic root were taken as de-scribed previously [13,14]. Briefly, the hearts were bi-sected just below the level of the atria and the base of the heart plus aortic root were taken for analysis. After equilibrating the tissue in OCT compound overnight the hearts were immersed in OCT compound on a cryostat chuck (Bright Instrument Company Ltd, UK) with the aorta facing the chuck. The tissue was frozen by surrounding the chuck with dry ice. The hearts were then sectioned perpendicular to the axis of the aorta, starting within the heart and proceeding in the direction of the aorta. After the aortic root had been identified by the appearance of the three-valve leaflets, alternate 10-mm sections were taken and mounted on gelatinised
slides. Sections were air dried for 1 h and subsequently rinsed briefly in 60% isopropyl alcohol. The sections were stained with Oil Red O, counterstained with Mayer’s haematoxylin, cover slipped using glycerol gelatine and sealed with nail varnish.
2.8. Quantification of atherosclerosis in the aortic root
the lesions and the areas stained red were obtained by calibrating the software using an image of a micrometer slide taken at the same magnification.
2.9. Statistical analysis
Non-baseline adjusted serum cholesterol and triglyce-ride, body weight and food consumption data were analysed at weeks 0, 4, 8 and 12 using a mixed-model, repeated-measures ANOVA. Multiple comparison least significant difference (LSD) tests were then performed on the serum triglyceride data. Body weight and food
consumption data were natural log transformed prior to ANOVA to normalise variance. Atherosclerotic le-sion area and oil red O area data were also analysed by ANOVA followed by a multiple comparison LSD test. Significance was assumed if P00.05. Analyses were performed using SAS software (version 6.11) running on a PC.
The procedures involving animals in this study were subject to both internal review and UK Home Office regulations.
3. Results
3.1. Body weight and food consumption
The body weights of mice in all four groups increased throughout the study as expected. The weights of C3H apoE(− / −) mice were significantly greater (7 – 24%) than those of the C57 apoE(− / −) mice both prior to and 4, 8 and 12 weeks after the start of Western diet feeding (P00.0017 for each timepoint). Male mice of both strains were heavier (18 – 33%) than their female littermates both prior to and throughout the study (P=0.0001). Male mice ate more than females when fed the normal diet (5.1 vs 4.8 g, P=0.011). As ex-pected, food consumption fell in all 4 groups when the mice were fed the Western diet but it did not differ significantly between the sexes and strains throughout this period (3.390.2 g, mean9S.D.).
3.2. Serum cholesterol and triglyceride concentration
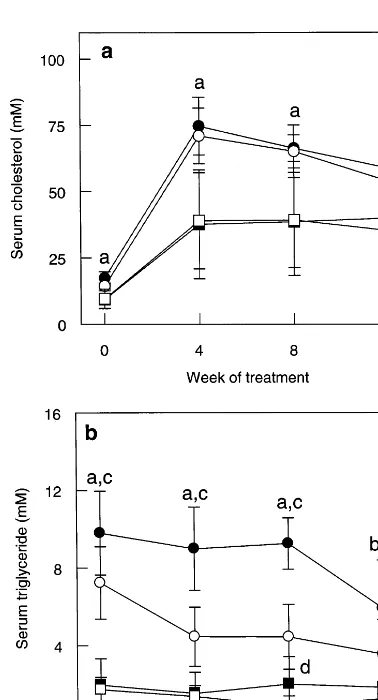
When fed normal diet, C3H apoE(− / −) mice had higher serum cholesterol (males: 1.9 fold, females: 1.5 fold) and very much higher triglyceride (males: 5 fold, females: 4.2 fold) concentrations than C57 apoE(− / −
) mice (P=0.0001, see Fig. 1). There was no difference in the serum cholesterol concentrations of male and female mice of either strain or between the serum triglyceride levels of male and female C57 apoE(− / −) mice. However, male C3H apoE(− / −) mice did have 35% higher serum triglyceride concentrations than their female littermates (P=0.0004).
Total serum cholesterol was increased dramatically in all four groups of mice after 4 weeks of feeding the Western diet, with C3H apoE(− / −) mice maintaining a 1.5 – 2 fold higher serum cholesterol concentration than C57 apoE(− / −) mice (P=0.0001, see Fig. 1a). There were no significant differences in the serum cholesterol concentrations of male and female mice of either strain throughout the study.
Total serum triglyceride concentration decreased rather than increased in all four groups following the switch to Western diet (see Fig. 1b). C3H apoE(− / −) mice had 3 – 6 fold higher serum triglyceride concentra-Fig. 1. Total serum cholesterol (a) and triglyceride (b) concentrations
in male and female apoE(− / −) mice on either C3H or C57 back-grounds fed Western diet for 12 weeks. Total cholesterol and triglyc-eride concentrations (mM) were measured in blood taken from non-fasted mice as described in ‘Methods’ (mean9S.D.). C3H apoE(− / −) mice had significantly higher serum cholesterol and triglyceride concentrations than C57 apoE(− / −) mice throughout the study (a: P=0.0001, b: P00.0033). Male C3H apoE(− / −) mice had higher serum triglyceride concentrations than female C3H apoE(− / −) mice throughout the study (c:P00.0027). Male C57 apoE(− / −) mice had higher serum triglyceride concentrations than female C57 apoE(− / −) mice at week 8 (d: P=0.024). =Male C3H apoE(− / −) mice (n=10); =male C57 apoE(− / −) (n=
Table 1
Percentage distribution of serum lipoprotein cholesterol and triglyceride in male and female apoE(−/−) mice on either C3H or C57 backgrounds before and after 12 weeks of being fed Western dieta
Cholesterol Triglyceride
Sex Total VLDL/Chy IDL/LDL HDL VLDL/Chy IDL/LDL HDL
Strain Total
Week
(%) (%) (%)
(mM) (mM) (%) (%) (%)
0 C3H Male 17.492.4 80 14 6 9.892.2 98 2 0
Female 14.491.1 72 12 16 7.291.9 99 1 0
Male 9.493.3 59 26 15 2.091.4
C57 99 1 0
Female 9.693.7 73 20 6 1.790.6 85 10 5
Male 62.7912.8 8892 1091 290.5 5.992.5 95 3
12 C3H 2
Female 51.994.4 9091 991 190.2 3.691.8 98 2 0.5 Male 39.2919.4 8890.3 1190.6 290.5 1.891.9
C57 95 3 2
Female 37.998.2 9190.5 890.3 290.3 1.390.3 98 2 0
aEqual volumes of serum were pooled from all of the mice in each group at week 0 and 12 of the study. Individual serum samples from five mice in each group were used to produce cholesterol lipoprotein profiles at week 12. Pooled and individual samples were fractionated using a Superose 6 gel filtration column and the fractions analysed for cholesterol and triglycerides as described in ‘Methods’. Total serum cholesterol and triglyceride concentrations are expressed as mean9S.D.
tions than C57 apoE(− / −) mice throughout the study (P00.0033 for each timepoint). Male C3H apoE(− / −) mice had significantly higher (66 – 108%) serum triglyceride concentrations than female C3H apoE(− / −) mice at weeks 4 – 12 (P00.0027 for each time-point). In contrast, male C57 apoE(− / −) mice had higher triglyceride levels than female C57 apoE(− / −) mice only at week 8 (134%, P=0.024).
3.3. Lipoprotein profiles
Lipoprotein profiles were acquired from serum sam-ples taken from normal diet fed mice and again from the mice after 4, 8 and 12 weeks of Western diet feeding. Equal volumes of serum from all the mice in each group were pooled prior to analysis at all time points, except week 12 when individual serum samples from five mice from each group were fractionated. Lipoprotein profile data for weeks 0 and 12 are shown in Table 1. The cholesterol and triglyceride lipoprotein profiles were very similar in weeks 4, 8 and 12, so only the data from week 12 were included in Table 1.
In male and female C3H apoE(− / −) and female C57 apoE(− / −) mice fed normal diet, the vast major-ity (72 – 80%) of cholesterol was present in the VLDL/
Chy fraction, whilst in male C57 apoE(− / −) mice fed chow, a much smaller percentage (59%) of the total serum cholesterol was found in this fraction (see Table 1). After 12 weeks of feeding Western diet the VLDL/
Chy fraction represented even more (88 – 91%) of the total serum cholesterol in all mice. The concentrations of HDL cholesterol were small and represented only 1 – 2% of total serum cholesterol in all four groups of mice.
Serum triglyceride was almost exclusively found in the VLDL/Chy fraction when the mice were fed either
normal or Western diet. Only a negligible amount of triglyceride was present in the IDL/LDL and HDL-sized particles.
3.4. Areas and lipid content of atherosclerotic lesions
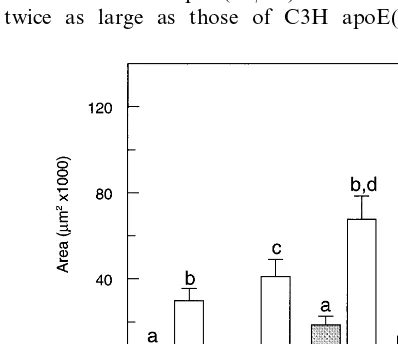
The total cross-sectional areas of lesions in both male and female C57 apoE(− / −) mice were more than twice as large as those of C3H apoE(− / −) mice
Fig. 2. Total sectional area of atherosclerotic lesions and cross-sectional area of lesion stained with ORO in male and female apoE(− / −) mice on either C3H or C57 backgrounds fed Western diet for 12 weeks. Areas (mm2) were measured as described in
(malesP=0.0088, females P=0.0001, see Fig. 2). The total cross-sectional area of atherosclerotic lesions was 51% greater in female C57 apoE(− / −) than in male C57 apoE(− / −) mice (P=0.016) and although the area of lesions tended to be larger in female C3H apoE(− / −) than male C3H apoE(− / −) mice, this difference did not reach significance.
The total area of lesion stained with ORO in male C57 apoE(− / −) mice was higher than in male C3H apoE(− / −) mice (P=0.002) but there was no signifi-cant difference when these stained areas were expressed as percentages of the total cross-sectional area of lesion. There was no difference in the ORO stained area between females of either strain. The percentage of lesion stained by ORO in male C57 apoE(− / −) mice was twice that in female C57 apoE(− / −) mice (27% vs 14%, P=0.0008). There was no difference in ORO area between the sexes for C3H apoE(− / −) mice.
4. Discussion
It is well known that when some inbred strains of mice are fed an atherogenic diet they are more suscepti-ble to developing atherosclerosis than others [1]. Nor-mal C57 mice are known to be an atherosclerosis susceptible strain whilst normal C3H mice are resistant. The present study was designed to investigate whether the difference in the susceptibility of C57 versus C3H mice is maintained when the apoE gene is silenced, a knockout known to greatly increase the susceptibility for atherosclerotic lesion formation. We measured total and lipoprotein cholesterol and triglyceride in serum and quantified lesion formation in the aortic sinus of both male and female mice.
The differences in serum lipids caused by the intro-duction of the apoE null mutation into C57 mice have been described previously. We have found some inter-esting differences in serum lipid levels that depend on the strain of mouse into which the apoE(− / −) muta-tion is introduced. C3H- and C57 apoE(− / −) mice fed a normal diet had serum cholesterol levels that were 3 – 6 fold higher than those found in normal C3H and C57 mice fed a similar diet [3,15]. Others have found slightly higher serum cholesterol concentrations in C57 apoE(− / −) mice fed a chow diet than those reported here (:14 – 16 vs :9.5 mM, respectively) [9,16]. The reasons for this difference are not clear but may relate to composition of the diet used or the number of generations of backcrossing the mice had undergone. When C3H- and C57 apoE(− / −) mice were fed West-ern diet containing 0.25% cholesterol, serum cholesterol concentrations increased enormously to approximately 2.8% and 1.5% w/v in the serum, respectively. Com-pared with normal C3H and C57 mice fed a diet containing 4 – 5 times as much cholesterol, the apoE(−
/ −) mice in the present study had approximately 5 – 15 fold higher serum cholesterol levels [3,15]. Cholesterol levels in the C57 apoE(− / −) mice fed Western diet were slightly lower than those reported previously for C57 apoE(− / −) mice fed atherogenic diets, although a direct comparison is difficult because different diets and lengths of treatment were used [9,16].
Very large differences in diet-induced hypercholes-terolaemia were found between the two strains of knockout mice. C3H apoE(− / −) mice had 1.5 – 2 fold higher serum cholesterol concentrations than C57 apoE(− / −) mice when fed either normal or Western diet. When a diet containing approximately 1% choles-terol, 15% fat and 0.5% cholic acid was fed to normal C3H and C57 mice, Nishina et al. [2] also reported that C3H mice had higher serum cholesterol concentrations than C57 mice, although another group [15] found that C57 mice had significantly higher cholesterol concentrations.
C57 apoE(− / −) mice fed normal diet had serum triglyceride concentrations that were similar to those reported by others in C57 apoE(− / −) mice [16] and half those of wild-type C57 mice [15] fed a similar diet. Interestingly, triglyceride levels in C3H apoE(− / −) mice were 5-fold higher than those found in normal C3H mice [15]. When mice were fed the Western-type diet, serum triglyceride concentrations tended to decline in the C3H apoE(− / −) whilst they remained unal-tered in the C57 apoE(− / −) mice. Serum triglyceride levels in C3H- and C57 apoE(− / −) mice fed Western diet were 2 – 3 fold higher and half those in their respective background strains fed an atherogenic diet [15]. The triglyceride concentrations of C57 apoE(− / −) mice fed Western diet were only half those reported by Tangirala et al. [16].
As with the serum cholesterol concentrations there were large differences in serum triglyceride levels be-tween the two strains of mice. Serum triglyceride levels were 3 – 6 fold higher in C3H- than in C57 apoE(− / −) mice when fed either chow or Western diet. These results were consistent with those obtained from some studies using normal C3H and C57 mice. Normal C3H mice are reported to have higher serum triglyceride levels than C57 mice although the magnitude of the difference was larger in the apoE(− / −) mice than in the normal mice (3 – 6 fold vs 1 – 2 fold, respectively) [2,15].
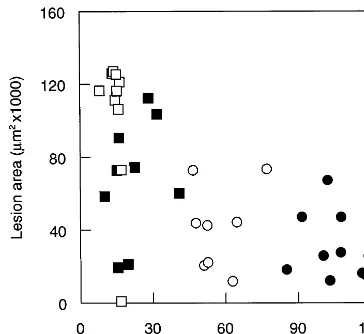
Fig. 3. Atherosclerotic lesion areas versus serum triglyceride exposure in male and female C3H- amd C57 apoE(− / −) mice fed Western diet for 12 weeks. Triglyceride exposure was calculated as the area under the serum triglyceride curve for each mouse. =Male C3H apoE(− / −) mice (n=10); =male C57 apoE(− / −) (n=9); =female C3H apoE(− / −) (n=8);=female C57 apoE(− / −) (n=10).
reported to have relatively small lesions of between 1.8×103and 15
×103
mm2[1,4,17]. In contrast to these
data the lesions in the aortic root of male and female C57 apoE(− / −) mice were 68911×103 and 1029
12×103
mm2 (mean9SEM), respectively, which were more than twice as large as those of sex-matched C3H apoE(− / −) mice (3096×103and 4198×103
mm2).
Silencing the apoE gene in the C3H and C57 mice, therefore, greatly increased the cross-sectional area of atherosclerotic lesions, by more than 30 fold and 7 – 10 fold, respectively. The relative difference in lesion areas between the two strains was, however, reduced from around 10 fold in the background strains to only 2-fold in the apoE(− / −) mice.
Female C3H- and C57 apoE(− / −) mice were more susceptible to developing atherosclerosis than their male littermates, although the difference between the male and female C3H apoE(− / −) mice was not sig-nificant. A number of other groups have also reported that female mice are more susceptible to developing atherosclerosis than males [10,17] and that when female C57 mice were given testosterone they were shown to have reduced lesion formation [10].
We have previously reported that serum cholesterol exposure is directly related to the size of atherosclerotic lesions in apoE3 Leiden mice [14]. In light of this observation and the results for serum cholesterol from the present study, we might have expected atheroscle-rotic lesions to be larger in C3H apoE(− / −) mice than in C57 apoE(− / −) mice. Paradoxically, how-ever, we found that the atherosclerotic lesion areas in the C57 apoE(− / −) mice were more than twice as large as those in C3H apoE(− / −) mice. In previous studies in a variety of inbred strains, little or no correla-tion was found between serum cholesterol concentra-tion and atherosclerosis [15]. Although there appeared to be an inverse relationship between serum cholesterol exposure and atherosclerotic lesion size for the groups of mice in the present study, we were not able to show this statistically because there was no correlation within each group.
Surprisingly, the apoE(− / −) mice with the lowest serum triglyceride concentrations had the largest atherosclerotic lesions. Serum triglyceride exposure was calculated from the area under the serum triglyceride curve (AUC) and compared with the size of the lesions in the individual mice (Fig. 3). However, we were not able to demonstrate a relationship between serum triglyceride exposure and atherosclerosis statistically be-cause there was no correlation within each group. It is probable that there are underlying differences in lipo-protein metabolism that result in the greater accumula-tion of triglyceride-rich lipoproteins in the serum of male vs female C3H apoE(− / −) mice and C3H- vs C57 apoE(− / −) mice and that these lipoproteins may be less atherogenic. It is believed that certain types of serum cholesterol concentrations than female C57 mice
fed the same diet [3]. This sex difference was not seen when C57 mice were fed an atherogenic diet or in C3H mice fed either diet. Qiao et al. also found no difference between the sexes in C57 mice fed an atherogenic diet, although male C3H mice did have higher serum choles-terol concentrations than females [17]. The same group also found that male C57 apoE(− / −) mice had higher cholesterol levels than females when fed chow.
When C3H- and C57 apoE(− / −) mice were fed either chow or Western diet, 84 – 94% and 98 – 99% of cholesterol, respectively was present in the non-HDL (VLDL/Chy/IDL/LDL) fractions and negligible amounts were in HDL. These results are consistent with those reported in the literature for apoE(− / −) mice [18]. However, in wild-type C3H and C57 mice fed a normal diet, the main proportion (83 – 87%) of the cholesterol is reported to be in HDL and only following the feeding of an atherogenic diet does serum choles-terol increase mainly in the non-HDL fraction [4]. Even when they are fed an atherogenic diet, it is reported that only 67 – 88% of cholesterol is in the non-HDL fractions in male and female C3H and C57 mice [17]. The main difference in the distribution of cholesterol across the lipoprotein profiles of normal and apoE(− / −) mice is, therefore, the much higher proportion of HDL cholesterol in normal mice.
Atherosclerotic lesions are reported to be either ab-sent or very small (B1000 mm2) in normal C3H mice
hypertriglyceridaemia in humans that are not linked with low HDL are not associated with an increased risk for developing atherosclerosis [19]. It is possible that the triglyceride-rich lipoproteins in C3H mice were chylomicrons because the mice were not fasted before blood samples were taken in this study. However, this is unlikely because there was no increase in serum triglyc-eride levels when the mice were transferred from the normal diet containing 2.6% fat to the Western diet containing 16% fat. It is probable, therefore, that these particles are VLDL. Whatever the nature of the triglyc-eride-rich lipoproteins in the C3H apoE(− / −) mice, it is likely that the triglycerides they contain will be hydrolysed by lipoprotein lipase (LPL) before they can be removed from the blood. Renier et al. reported that the expression and activity of LPL is 2 – 3 fold higher in the peritoneal macrophages taken from wild-type C57 mice than from normal C3H mice [20]. They proposed that high LPL secretion would accelerate the degrada-tion of VLDL and so facilitate the uptake of its rem-nants by the macrophages. This process would be expected to result in the conversion of macrophages into foam cells and so accelerate atherosclerosis. The model proposed by Renier et al. provides a good expla-nation for the lower serum cholesterol and triglyceride concentrations and greater atherosclerosis we found in C57- than in C3H apoE(− / −) mice.
Differences in the inflammatory and oxidative re-sponses to atherogenic diets could also contribute to the difference in atherosclerosis susceptibility between C3H and C57 apoE(− / −) mice. In C3H and C57 mice, these diets have been shown to increase gene expression of macrophage-colony stimulating factor (M-CSF), monocyte chemotactic protein (MCP-1), serum amyloid – A, heme oxygenase and NF-kB in C57
but not C3H mice [21]. The levels of conjugated dienes, markers of lipid oxidation, were also greater in C57 than in C3H mice. The macrophages of C57 mice have been reported to have a lower neutral cholesterol es-terase activity than those of C3H mice and, conse-quently, a reduced release of free cholesterol which could result in the accumulation of cholesterol esters and increased foam-cell formation [3].
It is reported that HDL cholesterol concentrations and susceptibility to atherosclerosis are negatively cor-related and that in mice, HDL levels are influenced by the four genesAth1 –Ath4 [4]. HDL is thought to have an anti-atherogenic activity by virtue of its ability to inhibit the pro-atherogenic oxidation of LDL [22] and its role in reverse cholesterol transport, whereby excess cholesterol is removed from cells and carried to the liver for excretion in the bile [23]. Nishina et al. re-ported that when normal C3H and C57 mice were fed chow diet, most (71 – 87%) of the cholesterol was in the HDL fraction and very little (13 – 29%) in the non-HDL fractions. On feeding an atherogenic diet, there was a
marked increase in serum cholesterol mostly in the non-HDL fractions and, in particular, the VLDL frac-tion [2]. In normal C57 mice there was a reducfrac-tion in HDL cholesterol, a decrease that was associated with changes in the HDL apolipoproteins A-I, A-II and C [2,24]. In contrast, when normal C3H mice were fed the same atherogenic diet it caused no significant change in the levels of HDL cholesterol. A number of other groups have also reported that normal C57 mice have lower concentrations of HDL cholesterol than C3H mice when fed an atherogenic diet [3,17,24]. In the present study, when mice of either sex or strain were fed Western diet, more than 88% of cholesterol was present as VLDL/Chy and only 1 – 2% was in the HDL fraction. We did not find a difference in HDL choles-terol levels between groups that could account for the observed difference in lesion development between C3H- and C57 apoE(− / −) mice.
The fact that the relative susceptibility of the back-ground strains to develop atherosclerosis is maintained in apoE(− / −) mice shows that the difference in the susceptibility of the two strains to atherosclerosis is probably not dependent on apoE. It is, however, inter-esting that the difference in susceptibility between the background strains was increased in this model.
Atherosclerotic lesion development in normal mice is very slow and frequently absent even after many months of feeding an atherogenic diet. However, with the development of the C3H apoE(− / −) mouse we now have a model that develops significant lesions and shows a more pronounced difference when compared with C57 apoE(− / −) mice. It is now our intention to investigate the genetic basis of the difference in atherosclerosis susceptibility between C3H- and C57 apoE(− / −) mice.
In conclusion, the marked difference in susceptibility for dietary-induced hyperlipidaemia and atherosclerosis in C57 versus C3H mice is not lost but is in fact increased by silencing the apoE gene.
Acknowledgements
The authors would like to thank Kim Prescott for her expert technical assistance and gratefully acknowl-edge Philip Overend, Edward Jay and Brian Bond for their expert help with and comments on the statistical analysis of the data.
References
[2] Nishina PM, Wang J, Toyofuku W, Kuypers FA, Ishida BY, Paigen B. Atherosclerosis and plasma and liver lipids in nine inbred strains of mice. Lipids 1993;28:599 – 605.
[3] Ishii I, Ito Y, Morisaki N, Saito Y, Hirose S. Genetic differences of lipid metabolism in macrophages from C57BL/6J and C3H/
HeN mice. Arterioscler Thromb Vasc Biol 1995;15:1189 – 94. [4] Paigen B. Genetics of responsiveness to high-fat and
high-choles-terol diets in the mouse. Am J Clin Nutr 1995;62:458S – 62S. [5] Hofker MH, van Vlijmen BJ, Havekes LM. Transgenic mouse
models to study the role of APOE in hyperlipidemia and atherosclerosis. Atherosclerosis 1998;137:1 – 11.
[6] Plump AS, Smith JD, Hayek T, Aalto-Setala K, Walsh A, Verstuyft JG, Rubin EM, et al. Severe hypercholesterolemia and atherosclerosis in apolipoprotein E-deficient mice created by homologous recombination in ES cells. Cell 1992;71:343 – 53. [7] Piedrahita JA, Zhang SH, Hagaman JR, Oliver PM, Maeda N.
Generation of mice carrying a mutant apolipoprotein E gene inactivated by gene targeting in embryonic stem cells. Proc Nat Acad Sci USA 1992;89:4471 – 5.
[8] Zhang SH, Reddick RL, Piedrahita JA, Maeda N. Spontaneous hypercholesterolemia and arterial lesions in mice lacking apolipo-protein E. Science 1992;258:468 – 71.
[9] Nakashima Y, Plump AS, Raines EW, Breslow JL, Ross R. ApoE-deficient mice develop lesions of all phases of atherosclero-sis throughout the arterial tree. Arterioscler Thromb 1994;14:133 – 40.
[10] Paigen B, Holmes PA, Mitchell D, Albee D. Comparison of atherosclerotic lesions and HDL-lipid levels in male, female, and testosterone-treated female mice from strains C57BL/6, BALB/c, and C3H. Atherosclerosis 1987;64:215 – 21.
[11] Nishina PM, Verstuyft J, Paigen B. Synthetic low and high fat diets for the study of atherosclerosis in the mouse. J Lipid Res 1990;31:859 – 69.
[12] Hennes U, Gross W, Edelmann A. Changes of lipoprotein patterns in hamsters under different metabolic states analyzed following separation by micropreparative chromatography on SMART™ System from multiple micro plasma samplings. Sci Tools 1992;36:10 – 2.
[13] Paigen B, Morrow A, Holmes PA, Mitchell D, Williams RA. Quantitative assessment of atherosclerotic lesions in mice. Atherosclerosis 1987;68:231 – 40.
[14] Groot PH, van Vlijmen BJ, Benson GM, Hofker MH, Schiffel-ers R, Vidgeon-Hart M, et al. Quantitative assessment of aortic atherosclerosis in APOE*3 Leiden transgenic mice and its rela-tionship to serum cholesterol exposure. Arterioscler Thromb Vasc Biol 1996;16:926 – 33.
[15] Paigen B, Morrow A, Brandon C, Mitchell D, Holmes P. Variation in susceptibility to atherosclerosis among inbred strains of mice. Atherosclerosis 1985;57:65 – 73.
[16] Tangirala RK, Casanada F, Miller E, Witztum JL, Steinberg D, Palinski W. Effect of the antioxidant N,N%-diphenyl 1,4-phenylenediamine (DPPD) on atherosclerosis in apoE-deficient mice. Arterioscler Thromb Vasc Biol 1995;15:1625 – 30. [17] Qiao JH, Xie PZ, Fishbein MC, Kreuzer J, Drake TA, Demer
LL, et al. Pathology of atheromatous lesions in inbred and genetically engineered mice: genetic determination of arterial calcification. Arterioscler Thromb 1994;14:1480 – 97.
[18] Ishibashi S, Herz J, Maeda N, Goldstein JL, Brown MS. The two-receptor model of lipoprotein clearance: tests of the hy-pothesis in ‘knockout’ mice lacking the low density lipoprotein receptor, apolipoprotein E, or both proteins. Proc Nat Acad Sci USA 1994;91:4431 – 5.
[19] Assmann G. Lipid Metabolism Disorders and Coronary Heart Disease. Mu¨nchen: MMV Medizin Verlag GmbH, 1993:98. [20] Renier G, Skamene E, DeSanctis JB, Radzioch D. High
macrophage lipoprotein lipase expression and secretion are as-sociated in inbred murine strains with susceptibility to atherosclerosis. Arterioscler Thromb 1993;13:190 – 6.
[21] Shih DM, Welch C, Lusis AJ. New insights into atherosclerosis from studies with mouse models. Mol Med Today 1995;1:364 – 72.
[22] Hayek T, Oiknine J, Dankner G, Brook JG, Aviram M. HDL apolipoprotein A-I attenuates oxidative modification of low density lipoprotein-studies in transgenic mice. Eur J Clin Chem Clin Biochem 1995;33:721 – 5.
[23] Breslow JL. Mouse models of atherosclerosis. Science 1996;272:685 – 8.
[24] Ishida BY, Blanche PJ, Nichols AV, Yashar M, Paigen B. Effects of atherogenic diet consumption on lipoproteins in mouse strains C57BL/6 and C3H. J Lipid Res 1991;32:559 – 68.