Summary In a greenhouse experiment, potted coastal Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) seedlings were grown in miniature ‘‘Nelder’’ (Nelder 1962) plots where growing space varied from 265 to 2555 cm2 per plant. After thirty weeks, mean plant height, crown biomass and branch number increased significantly (P = 0.0141) with decreasing growing space (increasing plant density). Differences in height growth became apparent about six weeks after sowing. Further-more, horizontally reflected radiation measured within the Nelder plots showed a decrease in red:far-red ratio (R:FR) from 1.2 at the lowest density to 0.71 at the highest. Plant height was strongly inversely correlated with estimated phytochrome pho-toequilibrium values (r2 = 0.893). Field measurements made in a three-year-old variable density plantation also showed a decrease in R:FR with increasing planting density from 300 to 3,000 trees ha−1. These results support the hypothesis that young Douglas-fir seedlings are able to detect, through the phytochrome system, the presence of nearby seedlings owing to the depletion of R relative to FR in the spectra reflected by the foliage of the adjacent plants. They then adjust their growth allometry in a way that reduces the possibility of being over topped by these future competitors.
Keywords: competition, photomorphogenesis, phytochrome, planting density, reforestation, shade avoidance.
Introduction
In young forest plantations, trees planted at high densities frequently show more rapid height and diameter growth than trees planted at low densities. The growth response (the ‘‘den-sity effect’’) often manifests itself long before seedlings are tall enough to shade one another, so it is not simply a response to shade. It occurs even when competition from other species is well controlled. The effect is ephemeral, typically lasting for a few to several years, and has been observed in several species (Cameron et al. 1991, DeBell and Giordano 1994, Knowe and Hibbs 1996).
It has recently been reported that early height and diameter growth of coastal Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) seedlings also increase with planting density (Scott et al. 1992, Scott et al., Weyerhaeuser Co., Federal Way WA, unpublished results). They established 46 variable density test
plantations in western Washington and Oregon beginning in 1985. On each site, seedlings were planted at six densities, ranging from about 3,000 trees ha−1 (2.0 m between seedlings) to 300 trees ha−1 (5.8 m between seedlings). In winter 1990, 1,220 plots were established across the 11 oldest tests. Seed-ling height and stem diameter at breast height (DBH) were measured in these plots. Taking the average seedling height at the 3,000 trees ha−1 stocking density as 100%, heights of trees at decreasing stocking densities were 91, 83, 85, 80 and 79%. Comparable values for DBH were 100, 91, 83, 81, 76 and 67%. Subsequent observations across the entire range of sites has confirmed these early results. Height growth differences typi-cally became visible during the third year after planting, when the seedlings were less than 1 m tall, and well before they were shaded by their neighbors.
The mechanism(s) triggering these differences in early growth have not been elucidated. One recent hypothesis is that plants are able to perceive the presence of nearby future com-petitors by sensing, through phytochrome, the spectral proper-ties of the radiation reflected by the foliage of adjacent plants (Ballaré et al. 1987, 1990, Smith et al. 1990, Ballaré et al. 1992, 1994, 1995). This radiation is depleted in the red (R; λ ~660 nm) region of the spectrum because of preferential absorption of red light by chlorophyll. Far-red (FR; λ ~730 nm) radiation, in contrast, is not diminished because virtually all FR incident on foliage is reflected or transmitted (Smith 1994). Hence, light reflected from foliage has a reduced R:FR ratio com-pared with solar radiation. The degree of reduction in incident reflected R:FR is a function of the proximity or density, or both, of competitors, thereby providing plants with an un-equivocal ‘‘early warning signal’’ (Ballaré et al. 1987) of their presence long before shading occurs.
It is well established that plants are able to perceive R:FR radiation signals through the phytochrome system by means of alterations in phytochrome photoequilibria (ϕ) (reviewed by Mancinelli 1994) and to transduce these signals into photo-morphogenic responses (Morgan et al. 1980, Schäfer and Briggs 1986, Mohr 1988, Smith et al. 1990) through changes in gene expression (Quail 1994, Quail et al. 1995). The first demonstration of this effect in trees was in radiata pine (Pinus radiata D. Don.) seedlings (Morgan et al. 1983). When ex-posed to artificial light with an altered R:FR ratio, these seed-lings showed marked changes in the timing and intensity of
Evidence for red:far red signaling and photomorphogenic growth
response in Douglas-fir (
Pseudotsuga menziesii
) seedlings
GARY A. RITCHIE
The George R. Staebler Forest Resources Research Center, Weyerhaeuser Company, Centralia, WA 98531, USA
Received February 13, 1996
flushing, shoot elongation and fasicle development. More re-cent work has suggested that phytochrome perception of R:FR signals from neighboring trees may affect needle orientation in Scots pine (Pinus sylvestris L.) (Galinski 1994). Gilbert and coworkers (1995) provide convincing evidence that ϕ exerts a strong influence on growth and crown architecture of poplar (Populus trichocarpa×deltoides ‘Beaupré’).
This paper reports results of an experimental test of this hypothesis with Douglas-fir seedlings that were grown in vari-able density plots in a greenhouse. Growth, biomass allocation and R:FR responses to a density gradient were measured. In addition, horizontally reflected R:FR radiation was monitored in three-year-old Douglas-fir plantations planted at three den-sities.
Methods
Greenhouse experimental layout
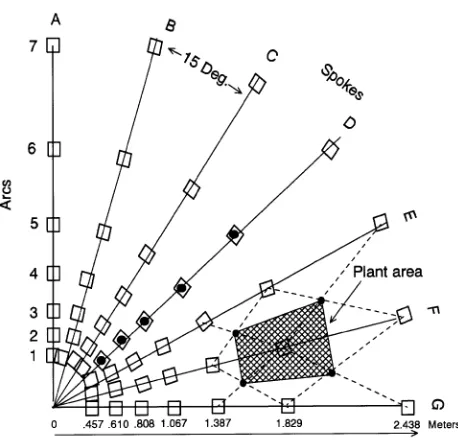
The experiment was conducted in a commercial greenhouse in Rochester, WA (lat. 46°75′ N, long. 123°20′ W, elev. 65m). The layout was based on a Nelder (1962) fan design. Four fans, each approximately 2.4 m square (Figure 1), were placed on greenhouse benches that were covered with 0.6-cm mesh hard-ware cloth. Each fan contained 49 plastic pots (10 cm2× 12 cm deep). The fans had seven spokes, each 15° apart and radiating from the origin. Pots were placed at successively greater dis-tances (arcs) along the spokes so that plants in successive arcs had progressively greater growing areas (Table 1). Spokes were labeled ‘‘A’’ through ‘‘G’’, arcs were labeled ‘‘1’’ through ‘‘7’’. Pots in Spokes A and G and Arcs 1 and 7 contained border plants that were not included in the measurements.
Plant culture
Douglas-fir seeds from a wild coastal Washington source were stratified at 4 °C in early December 1994. Sterile pots were filled with a mix comprising 1/1 (v/v) peat /perlite, and seeds were sown, one per pot, on February 16, 1995. After sowing, seeds were covered lightly with vermiculite to prevent drying and were protected with bird netting. Under-bench heating maintained the soil at about 18 °C during the germination period. On March 28, 1995, immediately following seedling emergence, the netting was removed and the fans were assem-bled. Seeds that did not emerge were replaced with seedlings of the same seed lot that had been sown in reserve. Fans were placed on the benches, and arrayed in random directions.
The plants were irrigated and fed every three days using a commercial overhead boom irrigation system. This ensured that all cultural treatments were applied evenly across the test. Feeding was with a commercial fertilizer (N,P,K; 21,5,20) at 150 ppm N, alternated with calcium nitrate supplemental feed. The greenhouse roof was a double layer of polyethylene film. In early April, moss growth was observed colonizing the surface of many of the pots. This was removed by hand and the soil surface covered with grit to prevent further moss coloniza-tion.
Measurements and harvest
Plant height was measured on all seedlings in Spokes B and F of all four fans every other week after sowing. Only two spokes were measured to avoid confounding the experiment by exces-sive touching of the plants (Braam and Davis 1990). Two measurements were made. The first was from the soil line to the base of the cotyledons (hypocotyls). The second measure was taken from the cotyledons to the growing tip (epicotyls).
Plants in three fans were harvested during the week of September 15, 1995. At that time, total plant height, stem diameter (two measurements averaged), number of branches, oven dry weight of plant top, oven dry weight of root system, and total oven dry weight of branches were determined. Total plant weight and root:shoot ratio were calculated from these
Table 1. Distance of each plant from the origin, distance from the previous plant, and growing area per plant in each Nelder fan.
Arc Distance Distance from Area per from previous plant origin (m) arc (m) cm2
1 0.457 --
--2 0.610 0.152 265
3 0.808 0.198 490
4 1.067 0.259 826
5 1.387 0.320 1432
6 1.829 0.472 2555
7 2.438 0.610
measurements. Several of the plants exhibited abnormal, stunted, development. These were noted on the data sheets and their measurements were not included in the analysis. One fan was retained for subsequent spectral measurements.
Spectral measurements
From August through October 1995 a recently calibrated LI-1800 spectroradiometer (Li-Cor, Inc., Lincoln, NE) fitted with an integrating cylinder (Ballaré et al. 1987) with a ± 10° acceptance angle was used to measure horizontally reflected radiation spectra coming from 360° at the intersect of each arc along Spoke D of Fan 3 (black dots in Figure 1). The integrat-ing cylinder was used because measurements made with tradi-tional cosine corrected planar sensors do not adequately characterize the radiation conditions at the photoreceptive sites in even-aged plant populations (Ballaré et al. 1990). The pro-cedure was to remove each plant from its position and to replace it with the sensor, which was located at roughly mid-crown height ~ 15--20 cm above the bench surface. Then a triple scan was run at 5-nm band intervals from 400 to 800 nm. The R:FR ratio was calculated as the ratio of photon irradiance in the bands at 655--665 nm (R) over irradiance in the bands 725--735 nm (FR) (Smith 1994). A black shroud was placed between the sensor and the persons making the measurements to prevent their presence from being sensed. During the August measurements, the sensor tip was accidentally chipped. A new tip was fabricated and used to make additional measurements at the same positions during October.
Data analysis
Initially, scatter plots were constructed with the raw data, in which each dependent variable was plotted against the plant growing area, which served as a surrogate for plant growing density (greater area = lower density). These plots revealed that the relationships tended to be logarithmic. Therefore, the plant growing areas were log transformed and subjected to linear regression analysis using the model: dependent variable = a + b log(plant area).
These analyses yielded P and r2 values for each regression. Regressions were evaluated by plotting residuals against ex-pected values. These analyses were conducted with pooled data from all three fans.
Field measurements
A variable density plantation had been established during February and March 1993 on an Astoria soil at Marcuson Creek near Dryad, WA (lat. 46°54′ N, long. 123°25′ W, elev. 125 m). One set of 0.4-ha blocks contained two-year-old Douglas-fir (transplanted (1 + 1) nursery stock) planted at 300, 1,400 and 3,000 trees ha−1. Before planting, the site had been sprayed to eliminate brush competition and fenced to prevent browsing by big game and rabbits. Brush was controlled by hand cutting during the three growing seasons following plant-ing. The site index (50-year height) was about 43 m.
The Li-Cor spectroradiometer with integrating cylinder was used to measure horizontally reflected R:FR radiation on sev-eral days during the growing season in the low density (300
trees ha−1), medium density (1,400 trees ha−1) and high density (3,000 trees ha−1) blocks. In each case, the tripod-mounted sensor was placed directly over the top of one seedling. Shorter-than-average seedlings were selected so that the sensor could be placed at roughly mid-crown for trees of average height, which was about 0.75 m. The instrument was progra-med to perform triple scans, which were averaged. Scans were run on several occasions in the high, medium, and low density blocks to gain information on spectral properties associated with stand density. The R:FR ratios were calculated as de-scribed above.
Results
Greenhouse experiment
Twenty-one trees were stunted (about 11% of total). Fifteen were in Fan 1, with Fan 2 and Fan 4 having only three each. There was no pattern to the distribution of stunted trees within fans. The reason for stunting is unknown, but it could have reflected seed size or vigor, or mutation. Stunted tree measure-ments were removed from the data base before analysis.
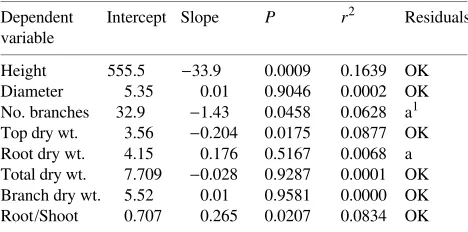
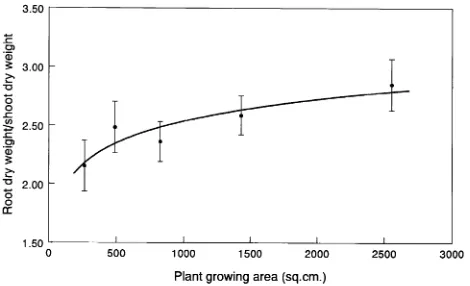
The slopes of the linear regressions on log(area) were sig-nificant for height, number of branches, top weight, and root:shoot ratio (Table 2). Among these, residual analyses were acceptable for all except branch number, where there was an indication of unequal variance. The decrease in plant height with increasing growing area (decreasing plant density) is shown in Figure 2 (all points shown). Virtually all of the height growth difference occurred in the epicotyls. A log curve of the type y=a+bln(x) was fitted through the regression. Mean height, mean branch number and mean top dry weight de-creased with increasing growing area in a similar manner (Figure 3). In contrast, the relationship of root:shoot ratio to growing area gave a positive slope, indicating that top growth increased relative to root growth as density increased (Fig-ure 4).
The ratio of R:FR radiation decreased as plant density in-creased (Figure 5). Some differences in the absolute values of R:FR across measurement dates were observed. These may have reflected differences in plant size, sun angle, sky condi-tions, or the change in sensor tips. Nevertheless, the trend was
Table 2. Linear regressions of dependent variables on plant growing area (log transformed).
Dependent Intercept Slope P r2 Residuals variable
Height 555.5 −33.9 0.0009 0.1639 OK Diameter 5.35 0.01 0.9046 0.0002 OK No. branches 32.9 −1.43 0.0458 0.0628 a1 Top dry wt. 3.56 −0.204 0.0175 0.0877 OK Root dry wt. 4.15 0.176 0.5167 0.0068 a Total dry wt. 7.709 −0.028 0.9287 0.0001 OK Branch dry wt. 5.52 0.01 0.9581 0.0000 OK Root/Shoot 0.707 0.265 0.0207 0.0834 OK
1
always for a logarithmic increase in R:FR with decrease in plant density.
Trends in R:FR in the field
Three patterns emerged from field measurements of horizontal R:FR radiation. First, R:FR varied diurnally, being high at dawn and dusk and reaching a daily minimum value during midday. Second, sky conditions affected R:FR. Diurnal curves were smooth on clear days and less so on cloudy or rainy days. The midday minima were often lower on clear, sunny days than on cloudy, rainy days. Finally, regardless of sky condi-tions, R was depleted relative to FR with increasing density (e.g., at midday on May 24, 1995, R:FR = 1.038, 0.939 and 0.792 at 300, 1,400 and 3,000 trees ha−1, respectively).
Discussion
This work was undertaken to gain a clearer understanding of the mechanism(s) that trigger the density effect in young Douglas-fir plantations. Several hypotheses have been sug-gested to explain the higher early growth rates associated with higher planting densities including the following.
(1) Animal browsing Assuming a relatively constant number of browsing animals in a given setting, a high-density plantation would have a lower ratio of animals to seedlings than a low-density plantation and would, presumably, suffer less brows-ing.
(2) Weed and brush competition Stands of trees planted at high densities would capture the site and its resources more rapidly compared with low-density stands. This would impede the development of weed communities and result in faster early tree growth.
(3) Mycorrhizal associations Most forest trees, including Douglas-fir, form mycorrhizal associations. A dense plantation might form more rapid connections among tree roots and mycorrhizal hyphae than a sparse plantation (Perry et al. 1992,
Figure 2. Final heights of individual Douglas-fir seedlings in all fans combined, as a function of plant growing area. Measurements were made approximately 205 days after seeds were sown. Log curve was fitted and plotted by computer.
Figure 3. Means (± 1 SE) of Douglas-fir seedling height, branch number, and top dry weight as a function of growing area. Data were collected approximately 205 days after seeds were sown. Log curve was fitted and plotted by computer.
Figure 4. Mean root/shoot ratios of Douglas-fir seedlings growing in all fans combined as a function of plant growing area. Measurements were made approximately 205 days after sowing. Log curve was fitted and plotted by computer.
Amaranthus and Perry 1994), hence providing more carbon for the mycorrhizal mat and promoting its growth and vigor. This would result in greater benefits to the trees and more rapid early growth.
(4) Root grafting Root grafting would occur sooner in dense plantings than in sparse plantings. Trees could then share and exploit site resources more efficiently and early growth would be enhanced.
(5) Nitrogen mineralization Compared to roots in a sparse plantation, roots in a dense plantation would exploit the rhi-zosphere more rapidly and increase the rate of nitrogen miner-alization, through root exudates, leading to more rapid early tree growth.
(6) Perception and avoidance of future competition Trees growing in close proximity with other trees are able to ‘‘per-ceive’’ the presence of these future competitors. This is fol-lowed by a change in growth programing that focuses on height growth in order to avoid overshadowing (Aphalo and Ballaré 1995). The mechanism of perception might be through R:FR signaling, or by root contact (Mahall and Callaway 1991).
This experiment failed to reject hypothesis No. 6 as it related to R:FR signaling. However, it did not test whether root inter-actions, competing brush, or animals have effects on early height growth in young stands. Further research will be re-quired to determine the significance of these potential effects. Despite the fact that the Nelder variable density planting was carried out in miniature in a greenhouse, its results are in concert with both the field observations of Scott et al. (1992) and with ‘‘growth waves’’ often observed in full-scale field Nelder plantings (Figure 6). In the field, increases occurred in height and stem diameter. In the greenhouse, height, branch number, and top weight increased, whereas stem diameter did not. However, if diameters varied across densities in the same
proportion as heights (about 10%), then the maximum differ-ences in diameter would have been on the order of only 0.5 mm. It is possible that the hand method used to measure stem diameter may not have been of sufficient precision to detect differences at this scale.
Results of both greenhouse and field measurements of R:FR radiation supported the R:FR signaling hypothesis. In both cases, R was depleted relative to FR as plant density increased. Differences in R:FR are perceived by the plant through the phytochrome system. Evidence from a wide range of plants species (summarized by Smith 1982, 1994) indicates that stem extension (height) growth is linearly and negatively related to phytochrome photoequilibrium (ϕ):
ϕ=Pfr/P, (1)
where Pfr is the phytochrome fraction in the far-red absorbing form, and P is the total phytochrome.
To test this with the height data obtained from the green-house study, ϕ values were estimated from the R:FR values measured on August 23 (Figure 5, August 23 values) based on the relationship provided by Smith (1982, Figure 2). Plant height, plotted as a function of estimated ϕ, yielded the pre-dicted linear relationship with a strong negative slope (Fig-ure 7) providing further support for the interpretation that the observed growth response was mediated by phytochrome.
Once the R:FR signal is perceived and transduced, plants presumably alter their internal growth program in such a man-ner as to gain greater light harvesting ability in crowded situ-ations (i.e., greater height) and improve their competitive position. This might occur by increasing the efficiency of either carbon assimilation or use, or both, or by changing carbon allocation patterns, or both. The final plant biomass distributions across the six densities provide insight as to what
may have occurred in the greenhouse study. Root weights decreased and total plant weights remained relatively constant, whereas top weights increased with density, resulting in a decreasing root:shoot ratio (Figure 4). This suggests that an increase in carbon allocation to above-ground parts, relative to below ground parts, may have occurred as a response to the R:FR signals and altered ϕ. This is consistent with observa-tions on soybean (Glycene max (L.) Merr.) (Kasperbauer 1987), and red beet (Beta vulgaris L.) (Hole et al. 1984).
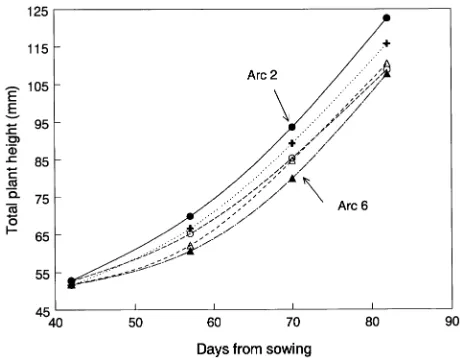
Other interpretations of these results cannot be ruled out, however. It is possible, for example, that subtle gradients in temperature, vapor pressure deficit, air movement, or other factors could have occurred within the fans and might have been implicated in the observed growth responses. However, this does not seem likely for the following reason. Weekly height measurements begun when the seedlings were just emerging from the cotyledon stage are shown in Figure 8. These growth curves show that the seedlings in Arcs 2 (highest density) and 6 (lowest density) were beginning to differentiate themselves from those in the other arcs as early as 45 days after sowing when the seedlings were only about 5.5 cm tall. It seems unlikely that, at this diminutive size, they could have modified the environment around seedlings growing 15 cm away sufficiently to cause them to alter their growth patterns. Nevertheless, this explanation cannot be discounted without further experimentation.
If the density effect evident in the field trials is, in fact, triggered by R:FR signaling among seedlings, it represents a considerable extension of the reported physical distances across which such signals are perceived. Field measurements of R:FR were made at various distances from artificial cano-pies constructed of tobacco (Nicotiana tobacum L.) or mustard (Sinapis alba L.) plants with a spectroradiometer equipped with a cosine corrected sensor (Smith et al. 1990). Red deple-tion and far-red enhancement were detected at a maximum
distance of 30 cm from these canopies. If R:FR signaling is triggering the density effect in our plantations, these signals must be propagated for a distance of at least 6 m, for that is the distance between seedlings planted at 3,000 trees ha−1. Inde-pendent measurements, made with the integrating cylinder adjacent to a 2.5-m-tall Douglas-fir hedge, showed a measur-able R depletion and FR enhancement out to approximately 3 m from the hedge (unpublished data).
A perplexing question that bears on this interpretation in-volves the role of lesser vegetation. Presumably such plants would also deplete red light energy, depressing the R:FR reflected from their foliage and sending ‘‘false signals’’ to the Douglas-fir seedlings. How would Douglas-fir seedlings dis-criminate among signals from plants that do not represent potential competitors and those that do (i.e., other tree spe-cies)? Several possible explanations come to mind. First, sig-nal timing may be important. Trees may be sensitive to the R:FR signal only during early spring, when their shoots are elongating, but before weeds and deciduous plants are in full leaf. Five-minute exposure of soybean plants to altered R:FR was sufficient to induce changes in morphogenesis (Kasper-bauer 1987), and elongating shoots are apparently the sites of R:FR perception in other species (Morgan et al. 1980, Child and Smith 1987, Ballaré et al. 1990). Alternatively, the phyto-chrome mechanism may be sufficiently sophisticated to dis-criminate among the reflectance properties of the leaves (Woolley 1971, Lee and Graham 1986) of different plants. Phytochrome is not a single molecular species, but is com-prised of a broad family of photoreceptors encoded by multiple divergent genes (Quail 1994). To complicate matters further, the ability of some plants to respond to phytochrome is de-pendent on the presence of blue or ultraviolet (UV-A) light (Mohr 1994). Such ‘‘coaction’’ between plant pigment systems has been demonstrated in Scots pine seedlings by Fernbach and Mohr (1990). Such richness of information-gathering
Figure 7. Relationship of Douglas-fir seedling final height to estimated phytochrome photoequilibria. Numbers in parentheses are arc num-bers from the origin of the Nelder fan. Each point is a mean ± 1 SE of all plants from all fans (about 25 plants per point).
leads to speculation that not only detection, but identification, of neighboring plants might be occurring in nature.
From a practical standpoint, the growth promoting effects of R:FR signaling can be exploited in forest management by increasing planting density and developing early thinning and silvicultural prescriptions to maintain proper stocking densi-ties. This is now being done in certain areas of the coastal Pacific Northwest. It is doubtful that greenhouse or nursery production could be enhanced, however, because typical grow-ing densities are already capturgrow-ing the density effect.
The implications of this phenomenon to modeling competi-tion in forest stands are obvious. The incorporacompeti-tion into such models, of algorithms simulating sophisticated information-gathering systems (Aphalo and Ballaré 1995) such as R:FR signaling, would tend to enhance their predictive ability be-yond that of models based solely on passive plant responses to resource limitations. This could lead to fundamentally new ways of thinking about the biology of plant competition and interaction, and should stimulate a great deal of additional research.
Acknowledgments
The capable technical assistance of James Keeley and Patty Ward is gratefully acknowledged. I also appreciate the generous cooperation of the Weyerhaeuser Company Rochester Regeneration Center staff in growing plants and for providing space to conduct this study. I thank Dr. Carlos Ballaré, University of Buenos Aires, Argentina for advice and for providing a prototype of the integrating cylinder used in the spectroradiometric measurements. Weyerhaeuser colleagues Drs. Wm. Scott, Roger Timmis, Christine Dean, Wm. Carlson, and Peter Farnum, along with Steven Duke and Paul Figueroa reviewed earlier drafts of this paper. Drs. R. van den Driessche, R. Guy, C. Harrington, T. Terry and R. Edmonds contributed to hypothesis development. Prof. Harry Smith, Univ. of Leicester, UK, and two anonymous referees provided reviews of the submission manuscript. A special thank you to Dr. Barbara Yoder, Oregon State University, who planted the early ideas for this work and provided me with two seminal papers on the topic of R:FR signaling. This research and its publication were sup-ported by Weyerhaeuser Company.
References
Amaranthus, M.P. and D.A. Perry. 1994. The functioning of ectomy-corrhizal fungi in the field: linkages in space and time. Plant Soil 159:133--140.
Aphalo, P.J. and C.L. Ballaré. 1995. On the importance of informa-tion-acquiring systems in plant--plant interactions. Funct. Ecol. 9:5--14.
Ballaré, C.L., Ana L. Scopel and R.A. Sánchez. 1990. Far-red radia-tion reflected from adjacent leaves: an early signal of competiradia-tion in plant canopies. Science 247:329--332.
Ballaré, C.L., Ana L. Scopel and R.A. Sánchez. 1995. Plant photomor-phogenesis in canopies, crop growth, and yield. HortScience. 30:1172--1181.
Ballaré, C.L., Ana L. Scopel, R.A. Sánchez and S.R. Radosevich. 1992. Photomorphogenic processes in the agricultural environment. Photochem. Photobiol. 56:777--788.
Ballaré, C.J., Ana L. Scopel, E.T. Jordan and R.D. Vierstra. 1994. Signaling among neighboring plants and the development of size inequalities in plant populations. Proc. Nat. Acad. Sci. USA 91:10094--10098.
Ballaré, C.L., R.A. Sánchez, Ana L. Scopel, J.J. Casal and C.M. Ghersa. 1987. Early detection of neighbour plants by phytochrome perception of spectral changes in reflected sunlight. Plant Cell Environ. 10:551--557.
Braam, J. and R.W. Davis. 1990. Rain-, wind-, and touch-induced expression of calmodulin and calmodulin-related genes in Arabi-dopsis. Cell 60:357−364.
Cameron, D.M., S.J. Rance, R.M. Jones and D.A. Charles-Edwards. 1991. Trees and pasture: a study on the effects of spacing. Agrofor. Today 3:8--9.
Child, R. and H. Smith. 1987. Phytochrome action in light-grown mustard: kinetics, fluence-rate compensation and ecological sig-nificance. Planta 172:219--229.
DeBell, D.S. and P.A. Giordano. 1994. Growth patterns of red alder.
In The Biology and Management of Red Alder. Eds. D.E. Hibbs, D.S. DeBell and R.F. Tarrant. Oregon State University Press, pp 116--130.
Fernbach, E. and H. Mohr. 1990. Coaction of blue/ultraviolet-A light and light absorbed by phytochrome in controlling growth of pine (Pinus sylvestris L.) seedlings. Planta 180:212--216.
Galinski, W. 1994. Non-random needle orientation in 1-year-old Scots pine (Pinus sylvestris L.) seedlings when adjacent to non-shading vegetation. Trees 8:160--164.
Gilbert, I.R., G.P. Seavers, P.G. Jarvis and H. Smith. 1995. Photomor-phogenesis and canopy dynamics. Phytochrome-mediated proxim-ity perception accounts for the growth dynamics of canopies of
Populus trichocarpa × deltoides ‘Beaupré.’ Plant Cell Environ. 18:475--497.
Hole, C.C., T.H. Thomas, A. Barnes, P.A. Scott and W.E.F. Rankin. 1984. Dry matter distribution between shoot and storage root of carrot, parsnip, radish and red beet. Ann. Bot. 53:625--631. Kasperbauer, M.J. 1987. Far-red light reflection from green leaves and
effects on phytochrome-mediated assimilate partitioning under field conditions. Plant Physiol. 85:350--354.
Knowe, S.A. and D.E. Hibbs. 1996. Stand structure and dynamics of young red alder as affected by planting density. For. Ecol. Manage. 82:69--85.
Lee, D.W. and R. Graham. 1986. Leaf optical properties of rainforest sun and extreme shade leaves. Am. J. Bot. 73:1100--1108. Mahall, B.E. and R.M. Callaway. 1991. Root communication among
desert shrubs. Proc. Nat. Acad. Sci. USA 88:874--876.
Mancinelli, A.L. 1994. The physiology of phytochrome action. In
Photomorphogenesis in Plants, 2nd Edn. Eds. R.E. Kendrick and G.H.M. Kronenberg. Kluwer Academic Press, Dordrecht, Boston, London, pp 211--269.
Mohr, H. 1988. Control of plant development: signals from without----signals from within. Bot. Mag. Tokyo 101:79--101.
Mohr, H. 1994. Coaction between plant pigments. In Photomorpho-genesis in Plants, 2nd Edn. Eds. R.E. Kendrick and G.H.M. Kronenberg. Kluwer Academic Press, Dordrecht, Boston, London, pp 353--373.
Morgan, D.C., T. O’Brien and H. Smith. 1980. Rapid photomodula-tion of stem extension in light-grown Sinapis alba L.: studies on kinetics, site of perception, and photoreceptor. Planta 150:95--101. Morgan, D.C., D.A. Rook, I.J. Warrington and H.L. Turnbull. 1983. Growth and development of Pinus radiata D. Don: the effect of light quality. Plant Cell Environ. 6:691--701.
Perry, D.A., T. Bell and M.P. Amaranthus. 1992. Mycorrhizal fungi in mixed-species forests and other tales of positive feedback, redun-dancy and stability. In The Ecology of Mixed-Species Stands of Trees. Eds. M.G.R. Cannell, D.C. Malcom and P.A. Robertson. Special Publication Series British Ecological Society, No. 11. Blackwell Scientific Publications, pp 151--179.
Quail, P.H. 1994. Phytochrome genes and their expression. In Photo-morphogenesis in Plants, 2nd Edn. Eds. R.E. Kendrick and G.H.M. Kronenberg. Kluwer Academic Press, Dordrecht, Boston, London, pp 71--104.
Quail, P.H., M.T. Boylan, B.M. Parks, T.W. Short Yong Xu and D. Wagner. 1995. Phytochromes: photosensory perception and sig-nal transduction. Science 268:675--680.
Schäfer, E. and W.R. Briggs. 1986. Photomorphogenesis from signal perception to gene expression: mini review. Photobiochem. Photo-biophys. 12:305--320.
Scott, W., R. Meade and R. Leon. 1992. Observations from 7- to 9-year old Douglas-fir variable density plantation test beds. Weyerhaeuser Forestry Research Field Notes 92-2, Centralia, WA, 2 p.
Smith, H. 1982. Light quality, photoperception and plant strategy. Annu. Rev. Plant Physiol. 33:481--518.
Smith, H. 1994. Sensing the light environment. In Photomorphogene-sis in plants, 2nd Edn. Eds. R.E. Kendrick and G.H.M. Kronenberg. Kluwer Academic Press, Dordrecht, Boston, London, pp 377--416. Smith, H., J.J. Casal and G.M. Jackson. 1990. Reflection signals and the perception by photochrome of the proximity of neighboring vegetation. Plant Cell Environ. 13:73--78.