PENGGUNAAN VERMIKOMPOS DALAM MENINGKATKAN
MUTU INOKULUM CENDAWAN MIKORIZA ARBUSKULA
UNTUK JATI MUNA (Tectona grandis Linn f.)
ASRIANTI ARIF
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2006
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis Penggunaan Vermikompos dalam Meningkatkan Mutu Inokulum Cendawan Mikoriza Arbuskula untuk Jati Muna (Tectona g randis Linn f.) adalah karya saya sendiri dibawah bimbingan komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, September 2006
Asrianti Arif NIM. E051030311
ABSTRAK
ASRIANTI ARIF. Penggunaan Vermikompos dalam Meningkatkan Mutu Inokulum Cendawan Mikoriza Arbuskula untuk Jati Muna (Tectona grandis Linn f.). Dibimbing oleh IRDIKA MANSUR, DEDY DURYADI S. dan SRI WILARSO BUDI R.
Mutu inokulum CMA merupakan hal penting yang perlu diperhatikan sehingga dapat mengurangi dosis inokulum yang diaplikasikan ketanaman dan mempercepat pengaruhnya terhadap tana man. Inokulum CMA dengan mutu yang baik dapat diperoleh jika persyaratan produksi inokulum dapat terpenuhi seperti pemilihan tanaman inang yang sesuai, media pertumbuhan, penambahan pupuk dan lingkungan yang cocok untuk perbanyakan cendawannya. Tujuan dari penelitian ini adalah untuk mempelajari respon jenis CMA terseleksi G. etunicatum dan Glomus sp. terhadap penambahan vermikompos, menguji penambahan vermikompos dalam meningkatkan mutu inokulum CMA, serta menguji formulasi inokulum CMA dan vermikompos dalam meningkatkan pertumbuhan semai jati Muna. Penelitian ini merupakan percobaan faktorial dengan RAL menggunakan 2 faktor perlakuan. Faktor pertama yaitu jenis inokulum CMA yang terdiri dari tiga taraf : 1) tanpa inokulasi CMA , 2) inokulasi CMA jenis Gl. etunicatum dan 3) inokulasi CMA jenis Glomus sp. Faktor kedua adalah formulasi media dengan vermikompos terdiri dari 100% zeolit, 90% zeolit dicampur 10% vermikompos, 80% zeolit dicampur 20% vermikompos, 70% zeolit dicampur 30% vermikompos, dan 60% zeolit dicampur 40% vermikompos.
Pada perbanyakan inokulum CMA, jenis G. etunicatum terlihat lebih toleran terhadap penambahan vermikompos sampai 40% (v/v) media dibandingkan jenis
Glomus sp. Akan tetapi jumlah propagul infektif inokulum kedua jenis CMA masih
belum dapat melampaui jumlah propagul pemberian pupuk hyponex merah. Setelah uji efektivitas ke semai jati Muna, formulasi inokulum CMA dengan vermikompos memberikan respon pertumbuhan dan serapan hara terbaik dibandingkan dengan kontrol.
Perlakuan formulasi inokulum CMA jenis G. etunicatum dengan vermikompos 40% dan Glomus sp. dengan vermikompos 40% memberikan respon yang sama terhadap pertumbuhan semai jati Muna dan merupakan perlakuan terbaik.
© Hak cipta milik Asrianti Arif, tahun 2006 Hak cipta dilindungi
Dilarang mengutip dan memperbanyak tanpa izin tertulis dari Institut Pertanian Bogor, sebagian atau seluruhnya dalam bentuk apa pun, baik cetak, fotokopi, mikrofilm dan sebagainya
PENGGUNAAN VERMIKOMPOS DALAM MENINGKATKAN
MUTU INOKULUM CENDAWAN MIKORIZA ARBUSKULA
UNTUK JATI MUNA (Tectona grandis Linn f.)
ASRIANTI ARIF
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Ilmu Pengetahuan Kehutanan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2006
Judul Tesis : Penggunaan Vermikompos dalam Meningkatkan M utu
Inokulum Cendawan Mikoriza Arbuskula untuk Jati Muna (Tectona grandis Linn f.)
Nama : Asrianti Arif
Nomor Pokok : E.051030311
Program Studi : Ilmu Pengetahuan Kehutanan
Disetujui, Komisi Pembimbing
Dr.Ir. Irdika Mansur, M.For.Sc Ketua
Dr.Ir. Dedy Duryadi Solihin, DEA Dr.Ir. Sri Wilarso Budi R, M.S. Anggota Anggota
Diketahui,
Ketua Program Studi Dekan Sekolah Pascasarjana Ilmu Pengetahuan Kehutanan
Dr.Ir. Rinekso Soekmadi, M.S. Dr.Ir. Khairil Anwar Notodiputro, M.S.
KATA PENGANTAR
Bismillahirrahmanirrahim
Puji syukur penulis panjatkan kepada Allah SWT, karena berkat rahmat dan karuniaNya jualah sehingga karya ilmiah ini berhasil diselesaikan. Karya ilmiah berjudul Penggunaan Vermikompos dalam Meningkatkan Mutu Inokulum Cendawan Mikoriza Arbuskula untuk Jati Muna (Tectona grandis Linn f.) ini disusun sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada Sekolah Pascasarjana Institut Pertanian Bogor.
Pada kesempatan ini penulis menyampaikan terima kasih dan penghargaan yang sebesar-besarnya kepada Bapak Dr.Ir. Irdika Mansur, M.For.Sc. sebagai ketua komisi pembimbing, Bapak Dr.Ir. Dedy Duryadi S. , DEA. sebagai anggota komisi pembimbing, dan Bapak Dr.Ir. Sri Wilarso Budi R., MS. sebagai anggota komisi pembimbing atas kesungguhan, kesabaran dan kebijakannya serta banyak memberi masukan dan saran yang sangat bermanfaat selama membimbing penulis.
Ucapan terima kasih penulis sampaikan pula kepada :
1. Rektor UNHALU dan Dekan Fakultas Pertanian UNHALU yang telah memberi izin kepada penulis untuk melanjutkan studi S2.
2. Direktur Proyek DUE-Like Batch II UNHALU yang telah memberikan beasiswa kepada penulis.
3. Prof. Dr.Ir. Syafrida Manuwoto mantan Dekan Sekolah Pascasarjana IPB, DR.Ir. Khairil Anwar Notodiputro, MS. Dekan Sekolah Pascasarjana IPB yang memberi kesempatan kepada penulis untuk menyelesaikan program S2. 4. Ketua Program Studi IPK
5. Dr.Ir. Iskandar Zulkarnaen Siregar Kepala Laboratorium Silvikultur FAHUTAN IPB dan penguji luar komisi.
6. Teman-teman seperjuangan di Klub Mikoriza Silvikultur, terutama Bapak Ir. Abimanyu D. Nusantara, M.P, ibu Dr.Ir. Yudhy Harini Bertham, M.P, La Ode Alimudin, SP. M.Si., Ibu Ir. Luluk Setyaningsih, M.Si., juga kepada
Ibu Dr.Ir. Gusti Ayu, M.Si., dan Bapak Dr.Ir. La Ode Safuan, M.P, yang telah banyak memberikan arahan dan bimbingan kepada penulis.
7. Seluruh rekan-rekan Program Studi Ilmu Pengetahuan Kehutanan, terutama Ira Taskirawati, S.Hut. M.Si., Asriyanni, S.Hut. M.Si. dan Sedek SP. atas bantuannya kepada penulis.
8. Teman-temanku Asniah, Wiwin, Ai Asiyah (grup Kwek-kwek) yang banyak membantu dan memberi semangat, Wardana (spesialis komputer) yang banyak memberi ilmunya, Sukma, Hafida Nur, Hajra (doyan fashion), Ida (tim sukses P6), Nirwan dan Bubun (kelucuannya selama di P6), Yuli (suka bawa oleh-oleh wingko semarang), dan semua kru P6, kepada sahabatku Rajab (trima kasih atas bantuannya) dan kepada semua pihak yang tidak dapat disebutkan satu persatu.
Ucapan terima kasih yang dalam penulis sampaikan kepada kedua orang tuaku (Bapak Arief La Adi dan ibu Harni), Almarhum Nenekku, Paman-pamanku Drs. Faad Maonde, MS., Mantri La Iynu, Ir. Dani, Tante -tanteku Ir. Husna, Husriah, Sanaria, Sumira dan kakakku Asriany Arif, adik-adikku Dr. Yuniati Arif, Abdul Rahman, Muh. Fadillah atas ketulusan, kesabaran dalam memberi dorongan da n semangat untuk penulis.
Penulis menyadari bahwa tulisan ini masih jauh dari sempurna, untuk itu kritik dan saran yang bersifat konstruktif sangat penulis hargai. Semoga karya ilmiah ini bermanfaat.
Bogor, September 2006
RIWAYAT HIDUP
Penulis dilahirkan di Ujung Pandang pada tanggal 15 November 1975 dari ayah Arief La Adi dan ibu Harni. Penulis merupakan putri kedua dari lima bersaudara.
Tahun 1993 penulis lulus dari SMA Negeri I Mawasangka dan pada tahun yang sama masuk Universitas Haluoleo pada Program Studi Agronomi, Jurusan Budidaya Pertanian, Fakultas Pertanian.
Penulis menjadi staf pengajar pada Fakultas Pertanian Unhalu sejak tahun 2001 dan pada tahun 2003 penulis mendapatkan kesempatan untuk melanjutkan pendidikan ke Program Pascasarjana IPB pada Program Studi Ilmu Pengetahuan Kehutanan. Beasiswa pendidikan Pascasarjana diperoleh dari Proyek Due-Like Batch II UNHALU.
DAFTAR ISI
Halaman
DAFTAR ISI ... vi
DAFTAR TABEL ... vii
DAFTAR GAMBAR ... viii
DAFTAR LAMPIRAN ... ix
PENDAHULUAN ... 1
TINJAUAN PUSTAKA ... 7
Cendawan mikoriza arbuskula ... 7
Klasifikasi Cendawan mikoriza arbuskula ... 8
Peranan CMA ... 10
Vermikompos ... 11
Peran CMA dan vermikompos ... 12
Perbanyakan inokulum dan potensi inokulum CMA ... 13
Jati ... 14
Penyebaran dan klasifikasi jati ... 15
Deskripsi buah dan benih jati ... 15
Tempat tumbuh jati ... 16
Jati Muna ... 17
METODOLOGI PENELITIAN ... 19
Waktu dan tempat ... 19
Bahan dan alat ... 19
Metode penelitian ... 19
HASIL dan PEMBAHASAN ... 28
Hasil ... 28
Pembahasan ... 39
SIMPULAN DAN SARAN ... 53
Simpulan ... 53
Saran ... 53
DAFTAR PUSTAKA ... 55
DAFTAR GAMBAR
Halaman
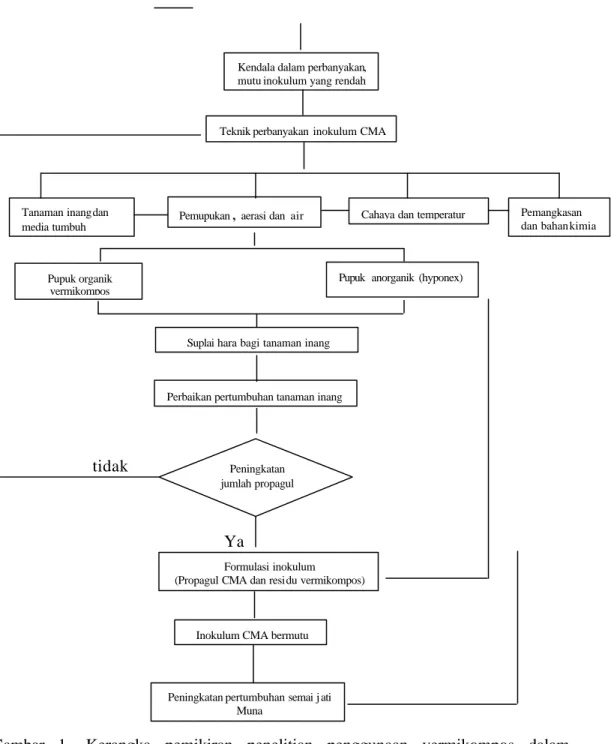
1. Kerangka pemikiran penelitian ... 5
2. Perbanyakan inokulum CMA ... 20
3. Kegiatan pengujia n potensi inokulum ... 23
4. Penyapihan dan pemeliharaan semai di polibag ... 25
5. Histogram pengaruh jenis inokulum CMA dan formulasi media vermikompos terhadap kolonisasi akar tanaman inang P. javanica ... 29
6. Histogram pengaruh jenis inokulum CMA dan formulasi media vermikompos terhadap jumlah spora tanaman inang P. javanica ... 29
7. Hifa dan vesikel CMA pada akar P. javanica ... 30
8. Grafik pengaruh formulasi inokulum CMA terhadap tinggi semai jati Muna 33 9. Semai jati Muna yang diinokulasi CMA pada umur 12 MST ... 33
10. Histogram pengaruh formulasi inokulum CMA terhadap diameter semai jati Muna umur 12 MST ... 34
11. Grafik pengaruh dosis formulasi inokulum CMA terhadap tinggi semai jati Muna ... 34 12. Histogram pengaruh dosis formulasi inokulum CMA terhadap diameter semai jati Muna umur 12 MST ... 35
DAFTAR TABEL
Halaman 1. Hasil analisis sifat fisika dan kimia tanah beberapa lokasi penyebaran
populasi jati Muna di Kabupaten Muna ... 18 2. Rekapitulasi hasil analisis sidik ragam terhadap kolonisasi akar, jumlah
spora, dan pertumbuhan tanaman inang P. javanica ... 28 3. Pengaruh jenis inokulum CMA dan formulasi media vermikompos
terhadap bobot kering tanaman P. javanica ... 30 4. Potensi inokulum cendawan mikoriza arbuskula per 100 gram media
zeolit ... 31 5. Rekapitulasi hasil analisis sidik ragam terhadap peubah pertumbuhan,
kolonisasi akar dan jumlah spora semai jati Muna ... 32 6. Pengaruh inokulum CMA terhadap bobot kering dan nisbah pucuk akar
semai jati Muna ... 36 7. Pengaruh formulasi inokulum CMA terhadap bobot kering akar dan
bobot kering akar terinfeksi semai jati Muna ... 36 8. P engaruh formulasi inokulum CMA dan dosis formulasi inokulum
terhadap kolonisasi akar pada semai jati Muna ... 37 9. Pengaruh formulasi inokulum CMA dan dosis formulasi inokulum
terhadap jumlah spora pada semai jati Muna ... 38
DAFTAR LAMPIRAN
Halaman 1. Layout plot penelitian perbanyakan inokulum CMA dengan inang
P. javanica di rumah kaca selama tiga bulan ... 60 2. Layout percobaan uji efektivitas formulasi inokulum CMA pada semai
jati Muna di persemaian rumah kaca selama tiga bulan ... 61 3a. Informasi kandungan hara dari vermikompos ... 63 3b. Informasi kandungan hara hyponex merah ... 63 4. Informasi komposisi zeolit Produksi PT Inti Agro Persada Industri Jakarta 64 5. Hasil analisa sifat fisika dan kimia tanah yang digunakan untuk
media semai jati Muna ... 65 6. Hasil analisa kandungan hara formulasi inokulum CMA yang
diberikan ke semai jati Muna ... 66
7. Hasil analisa jaringan daun semai jati Muna ... 67 8. Tabel nilai MPN untuk seri pengenceran 10 kali (Halvorson dan Ziegler
1933) ... 68 9. Tabel contoh perhitungan jumlah propagul dengan metode The Most
Probable Number (MPN) ... 69 10. Pengaruh jenis CMA dengan formulasi media vermikompos terhadap
kolonisasi akar dan jumlah spora CMA tanaman P. javanica ... 70 11a. Pengaruh formulasi inokulum CMA terhadap tinggi dan diameter semai
jati Muna ... 71 11b. Pengaruh dosis formulasi inokulum terhadap tinggi dan diameter semai
jati Muna ... 71 12. Klasifikasi tingkat kolonisasi akar ... 72
PENDAHULUAN
Latar Belakang
Pemanfaatan cendawan mikoriza arbuskula (CMA) dalam produksi semai di daerah-daerah tropis telah banyak diketahui dan diuji. Diantara jenis pohon yang diuji, sebagian besar adalah pohon legum yang pertumbuhannya cepat seperti
Paraserianthes falcataria, Acacia mangium, Acacia crassicarpa, Sesbania grandiflora , Ca ssia siamea, Ca ssia suratensis, Parkia roxburgii, Enterolobium cylocarpum, Pterocarpus sp., dan pohon-pohon non legum seperti Gmelina arborea, Tectona grandis, Ceiba pentandra dan spesies -spesies pohon pioneer yang
merupakan pohon-pohon yang sangat responsif terhadap inokulasi CMA (Setiadi 2001). Pada umumnya semai yang terinfeksi mikoriza menunjukkan respon pertumbuhan yang lebih bagus daripada yang tidak terinfeksi.
Kondisi tanah tropis dicirikan dengan rendahnya unsur hara, temperatur ekstrim, kemasaman tanah tinggi dan kekeringan serta kehadiran beberapa patogen. Dengan demikian penggunaan CMA dapat memberikan keuntungan pada semai di pembibitan (nurseryes) sebelum dipindahkan ke lapangan (Feldmann dan Idczak 1992). Telah diketahui bahwa CMA merupakan salah satu agen hayati yang berasosiasi dengan akar dari suatu tumbuhan hidup yang terutama bertanggung jawab untuk transfer hara (Brundrett 2004). Peran CMA sebagai mikroorganisme alam adalah membantu penyerapan unsur hara terutama hara P dan hara lain seperti Zn, Cu, Ni, NH4+ dan NO3-, membantu penyediaan hara dari yang tidak tersedia menjadi tersedia bagi tanaman, membantu tanaman untuk dapat bertahan pada kondisi kekeringan karena adanya hifa-hifa cendawan yang mampu menembus pori-pori tanah dan memperluas daerah penyerapan air, dan sebagai proteksi dari serangan patogen akar (Brundrett et al. 1994 ; Smith dan Read 1997).
Namun pemanfaatan CMA sebagai agen hayati masih sangat terbatas. Hal ini disebabkan oleh beberapa kendala seperti spesifikasi dari CMA itu sendiri, terbatasnya jumlah inokulum yang efektif untuk diaplikasikan di lapangan, ketidakkonsistenan mutu inokulum dan pengaruhnya terhadap tanaman memerlukan waktu yang lama dibandingkan pupuk anorganik. Respon yang lambat pada pertumbuhan tanaman mengakibatkan konsumen lebih tertarik menggunakan pupuk
anorganik. Selain itu informasi terbatas dan kurangnya sosialisasi ke masyarakat tentang CMA.
Oleh sebab itu perbaikan mutu inokulum merupakan hal penting sehingga dapat mengurangi dosis inokulum jika diaplikasikan ke tanaman dan dapat mempercepat responnya. Inokulum CMA dengan mutu yang baik dapat diperoleh jika persyaratan produksi inokulum dapat terpenuhi seperti pemiliha n tanaman inang yang sesuai, media pertumbuhan, penambahan pupuk dan lingkungan yang cocok untuk perbanyakan cendawannya. Menurut Mansur (2002), perbanyakan inokulum CMA dapat dikatakan berhasil jika kolonisasi CMA intensif (dapat diamati dari hifa yang mengolonisasi jaringan perakaran) dan jumlah spora yang dihasilkan tinggi (kurang lebih 100 spora per 10 g media).
Masalah yang cenderung banyak diulas dan sering terjadi dalam produksi inokulum CMA adalah kondisi hara media yang digunakan menjadi penyebab kegagalan terjadinya infeksi atau kolonisasi cendawan mikoriza pada akar inangnya (Sieverding 1991; Bagyaraj 1991; Marschner 1992; Brundrett 1994; Smith dan Read 1997) . Sehingga perlu dicobakan berbagai jenis pupuk dan dosis yang tepat dalam rangka meningkatkan mutu inokulum CMA. Penggunaan inokulum CMA yang baik merupakan langkah efisien dalam menunjang peningkatan pertumbuhan tanaman di pembibitan dan keberhasilan pada saat pemindahan kelapangan.
Salah satu alternatif pupuk yang dapat ditambahkan pada in okulum CMA adalah pupuk organik vermikompos. Vermikompos dihasilkan dari kemampuan beberapa cacing tanah dalam mengkonsumsi residu organik seperti limbah rumah tangga, limbah industri seperti bubur kayu, residu panen seperti sayuran, daun-daunan, dedak padi, dedak jagung, kotoran ternak, kompos dan sebagainya (Ndegwa
et al. 1999). Vermikompos merupakan pupuk berkualitas tinggi sebab mengandung
berbagai unsur hara yang dibutuhkan oleh tanaman sebelum mendapatkan manfaat dari asosiasinya dengan cendawan. Sela in itu vermikompos yang merupakan pupuk organik juga dapat membantu memperbaiki sifat fisik dan kimia tanah (Samosir 1994). Sehingga beberapa formulasi penggabungan media dan pupuk vermikompos perlu dicoba untuk mencari formulasi media yang tepat tanpa mengganggu proses infeksi dan perkembangan kolonisasi CMA.
Penelitian yang mengombinasikan CMA dan vermikompos masih terbatas pada bagaimana meningkatkan pertumbuhan dan produktivitas tanaman, selain itu pemanfaatannya masih terbatas pada tanaman pertanian dan hortikultura (Sáinz et al. 1998; Vasanthi dan Kumaraswamy 1999; Rajkhowa et al. 2000). Selama ini belum ada penelitian yang memanfaatkan vermikompos sebagai pupuk dalam perbanyakan inokulum CMA. Oleh sebab itu penelitian ini mencoba untuk melihat peluang pemanfaatan vermikompos dalam meningkatkan mutu inokulum CMA dan mencoba mendapatkan suatu formulasi inokulum CMA yang tidak hanya mengandung propagul CMA akan tetapi dapat juga digunakan sebagai pupuk untuk suplai unsur hara awal bagi tanaman.
Formulasi inokulum yang diperoleh akan dicobakan pada tanaman kehutanan seperti jati Muna (Tectona grandis L.f.) sebagai salah satu tanaman kehutanan unggulan yang dapat digunakan dalam rehabilitasi hutan-hutan yang sudah rusak dan gundul. Verbenaceae merupakan salah satu famili pohon yang dikenal bersimbiosis dengan CMA (Turjaman et al. 2004). Jenis komersil dari famili ini contohnya adalah jati (Tectona grandis L.f.) yang memiliki prospek cerah. P roduk berbahan baku kayu jati memiliki pangsa pasar luas karena termasuk kayu berkualitas tinggi, tergolong ke dalam kelas kuat II dan kelas awet I-II, yang merupakan kayu yang paling banyak dipakai untuk segala jenis kontruksi bangunan, mebel, kerajinan dan lain-lain (Pandit dan Ramdan 2002). Sulawesi Tenggara khususnya Kabupaten Muna adalah salah satu daerah penghasil jati dan di kenal ada 2 jenis jati yaitu jati Muna dan jati Malabar Muna. Namun populasi jati Muna sekarang sudah semakin berkurang, dari 70.000 ha menjadi 1000 ha (MENHUT1 2005). Hal tersebut mengindikasika n perlunya penyediaan semai untuk program rehabilitasi hutan guna mempertahankan populasi jati Muna yang sudah semakin langka.
Rumusan Masalah
Penelitian ini terdiri dari dua tahap, dimana penelitian pertama dilakukan untuk me nguji pengaruh aplikasi vermikompos terhadap kolonisasi dan pembentukan spora CMA, dengan mengombinasikan vermikompos dan zeolit untuk mencari formulasi yang tepat tanpa menghambat perkembangan CMA. Vermikompos digunakan sebagai sumber hara bagi tanaman, pengganti hyponex merah yang sering digunakan dalam produksi inokulum sebagai pupuk dengan ketersediaan P yang
rendah. Jenis CMA yang diperbanyak adalah Glomus etunicatum dan Glomus sp. (endogenous), dimana jenis CMA yang berbeda juga memiliki kemampuan yang berbeda dalam mentolerir pemberian pupuk. Sehingga permasalahan yang harus dijawab pertama adalah bagaimanakah respon kedua jenis CMA tersebut terhadap penambahan vermikompos. Pertanyaan kedua adalah apakah penambahan vermikompos dapat meningkatkan mutu inokulum CMA.
Penelitian kedua dilakukan untuk menguji pengaruh inokulum hasil perbanyakan terhadap semai jati Muna. Inokulum hasil perbanyakan mengandung CMA dan residu vermikompos yang dapat digunakan untuk memenuhi kebutuhan hara pada awal pertumbuhan tanaman, sehingga pertanyaa n ketiga yang harus dijawab adalah apakah formulasi inokulum CMA dan vermikompos dapat meningkatkan pertumbuhan jati Muna di persemaian. Adapun kerangka pemikiran pemecahan masalah dari penelitian yang dilakukan disajikan pada Gambar 1.
Kondisi lahan yang kurang mendukung untuk pertumbuhan semai
Input alternatif teknologi agen hayati cendawan mikoriza arbuskula (CMA)
`
tidak
Ya
Gambar 1. Kerangka pemikiran penelitian penggunaan vermikompos dalam meningkatkan mutu inokulum cendawan mikoriza arbuskula untuk jati Muna
Tujuan pene litian
Tujuan dari penelitian ini adalah:
Peningkatan pertumbuhan semai j ati Muna
Suplai hara bagi tanaman inang Kendala dalam perbanyakan, mutu inokulum yang rendah
Inokulum CMA bermutu
Teknik perbanyakan inokulum CMA
Tanaman inang dan media tumbuh
Cahaya dan temperatur Pemangkasan dan bahan kimia Pemupukan, aerasi danair
Pupuk organik vermikompos
Perbaikan pertumbuhan tanaman inang
Pupukanorganik (hyponex)
Peningkatan jumlah propagul
Formulasi inokulum (Propagul CMA dan residu vermikompos)
1. Untuk menguji respon jenis CMA terseleksi G. etunicatum dan Glomus sp. terhadap penambahan vermikompos.
2. Untuk menguji penambahan vermikompos dalam meningkatkan mutu inokulum CMA
3. Untuk menguji formulasi inokulum CMA dan vermikompos dalam meningkatkan pertumbuhan semai jati Muna .
Manfaat Penelitian
Hasil penelitian ini diharapkan dapat memberikan informasi dan masukan mengenai kombinasi vermikompos yang tepat dalam perbanyakan inokulum CMA, serta formulasi yang tepat dalam meningkatkan pertumbuhan semai jati Muna.
Hipotesis
1. Kedua jenis CMA memiliki respon yang berbeda terhadap penambahan vermikompos
2. Penambahan vermikompos dapat meningkatkan mutu inokulum CMA
3. Aplikasi formulasi inokulum CMA dan vermikompos dapat meningkatkan pertumbuhan semai jati Muna di persemaian
TINJAUAN PUSTAKA
Mikoriza dari asal katanya mykes yaitu cendawan atau jamur, dan rhiza yang berarti akar, kedua kata tersebut berasal dari bahasa Yunani. Banyak definisi yang dikemukakan berkaitan dengan cendawan mikoriza arbuskula. Menurut Sieverding (1991), bahwa mikoriza adalah simbiosis mutualisme antara cendawan tular tanah dan akar pada tanaman tingkat tinggi. Definisi mikoriza secara luas dikemukakan oleh Brundrett (2004), yang mencakup seluruh keragaman mikoriza sebagai suatu asosiasi simbiotik yang esensial bagi satu atau kedua mitra, antara suatu cendawan (terspesialisasi untuk hidup dalam tanah dan tumbuhan) dan akar (atau organ yang mengadakan kontak-substrat lainnya) dari suatu tumbuhan hidup, yang terutama bertanggung jawab untuk transfer hara. Mikoriza terjadi dalam suatu organ tumbuhan yang terspesialisasi dimana hubungan kontak dekat berasal dari perkembangan cendawan tumbuhan yang tersinkronisasi. Kategori utama VAM adalah asosiasi linear dan coiling, selanjutnya juga disarankan bahwa asosiasi mikoriza ditetapkan dan diklasifikasikan terutama melalui kriteria anatomi yang diatur oleh tumbuhan inang karena bentuk yang dikontrol cendawan beragam menurut tumbuhan.
CMA dapat berasosiasi dengan sebagian besar tumbuhan yang termasuk
Angiospermae, Gymnospermae, Pteridophyta, dan Bryopita. Tanaman kelompok dicotyledonous 83% dan kelompok monocotyledonous 79% berasosiasi dengan
CMA (Smith dan Rea d 1997; Sieverding 1991) dan sedikit alga (Alexopoulus
et al. 1996). Cendawan ini memiliki selang ekologis yang luas dan dapat dijumpai
dalam sebagian ekosistem yang meliputi hutan hujan rapat, lahan hutan terbuka, semak, savana, padang rumput, bukit pasir dan semi gurun. Tetapi cendawan ini jarang ditemukan pada hutan temperate yang dikuasai oleh konifer, areal yang amat basah seperti lahan padi disawah merupakan habitat yang kurang disukai (Setiadi 1989). Sebagian besar CMA bersifat acidophilic (senang kondisi masam) dengan kisaran pH 3.5– 6.0 (mikroba mesophilic), untuk jenis Gigaspora sp. dapat tumbuh dan berkecambah dengan baik pada pH 4-6 dan Glomus sp. secara umum pada pH 6-8, pada keadaan aerobik, mesothermal dengan kisaran suhu optimum 22-30°C, menyukai kelembaban kurang dari 80% dan tidak suka cahaya (Pelczar dan Chan 1986). Cahaya dan temperatur merupakan unsur iklim yang sangat mempengaruhi proses infeksi mikoriza arbuskula. Temperatur optimum bagi perkembangan spora
Glomus spp. memiliki distribusi yang luas, Gigaspora dan Sclerocystis spp. adalah umum dijumpai pada tanah-tanah tropis. Acaulospora dijumpai dapat beradaptasi dengan lebih baik pada tanah dengan pH <5,0. Kenyataannya, CMA berhubungan dengan jenis partikel tanah; Glomus mosseae dengan tekstur yang baik, subur, pH tanah tinggi; Acaulospora laevis dengan tekstur kasar, pH asam;
Gigaspora pada tanah pasir (Bagyaraj 1991).
Klasifikasi cendawan mikoriza arbuskula (CMA)
Menurut Alexopoulus et al. 1996 bahwa cendawan mikoriza arbuskula tergolong kedalam ordo Glomales dan memiliki 6 genus yaitu Acaulospora,
Entrophospora, Gigaspora, Glo mus, Sclerocystis, Scutellospora. Adapun sistem
klasifikasinya tergolong kedalam: kingdom : Fungi
phylum : Zygomycetes ordo : Glomales kelas : Zygomycetes
famili : Acaulosporaceae, Glomaceae, dan Gigasporaceae yang memiliki genus Gigaspora dan Scutellospora
genus : Acaulospora, Entrophospora, Gigaspora, Glomus, Sclerocystis,
dan Scutellospora
Namun Schubler et al. (2001), menegaskan bahwa setelah melalui tes analisis DNA, CMA ordo Glomales tidak masuk kedalam kelas Zygomycetes, Basidiomycetes, dan Ascomycetes tetapi berdiri sendiri. Glomeromycota memiliki 4 ordo yaitu:
1. Glomales memiliki 2 famili Glomeraceae (Glomus grup A) dan Glomeraceae (Glomus grub B).
2. Archaeosporales memiliki 2 famili Archaeosporaceae dan Geosiphonaceae 3. Paraglomales memiliki famili Paraglomaceae
4. Diversisporales dengan famili Gigasporaceae, Acaulosporaceae, dan Diversisporaceae
Sedangkan dalam INVAM (2006) dinyatakan bahwa cendawan mikoriza arbuskula (CMA) adalah salah satu tipe cendawan mikoriza dan termasuk ke dalam
golongan endomikoriza. Cendawan mikoriza arbuskula termasuk ke dalam golongan Glomeromycota , dengan ordo Glomales yang mempunyai 2 sub-ordo, yaitu Gigasporineae dan Glomineae. Gigasporineae dengan famili Gigasporaceae mempunyai 2 genus yaitu Gigaspora dan Scutellospora. Glomineae mempunyai 4 famili yaitu Glomaceae dengan genus Glomus, famili Acaulosporaceae dengan genus Acaulospora dan Entrophospora, Paraglomaceae dengan genus Paraglomus dan Archaeosporaceae dengan genus Archaeospora.
Cendawan tidak mampu melengkapi daur hidupnya tanpa adanya akar tanaman inang. Sporanya dapat berkecambah dan tumbuh tanpa tanaman inang, akan tetapi pertumbuhannya sangat terbatas. Masih terlalu sedikit yang diketahui mengenai mekanisme molekuler yang menentukan pensinyalan dan pengenalan antara CMA dengan tanaman inangnya. Pada tahap pertama dari pengenalan inang, hifa CMA sebelum membentuk appresorium akan membentuk percabangan hifa yang ekstensif di dekat akar inang sebelum, appresorium merupakan struktur yang digunakan untuk menembus akar tanaman. Akar inang diketahui melepaskan molekul-molekul sinyal yang memicu percabangan hifa. Ditemukan pada eksudat akar Lotus japonicus senyawa strigolaktone, strigol. Konsentrasi rendah strigogalakton 5-deoksi-strigol, sorgogalakton dan strigol alami, serta analog sintetisnya (GR24) ditemukan menginduksi percabangan hifa dari spora Gigaspora margarita yang sedang berkecambah (Akiyama et al. 2005).
CMA tidak mengubah morfologi eksternal akar tanaman tingkat tinggi, hifa tumbuh antara kedua dan dalam sel kortek dengan cara penetrasi pada lapisan dan menginvasi membran plasma. CMA menghasilkan coils, struktur yang menyebar menyerupai haustorium yang disebut arbuskula dan pembengkakan terminal yang disebut vesikel. Vesikel dibentuk juga antara atau dalam sel inang dan berfungsi sebagai penyimpanan energi untuk digunakan oleh cendawan ketika suplai metabolit inang rendah. Arbuskula adalah hifa yang menyebar sepanjang lapisan sel inang yang menginvasi membran sel inang. Hifa yang terspesialisasi ini menciptakan suatu area permukaan yang besar antara sel membran plasma inang dan melibatkan transfer metabolit dua arah oleh dua mitra. Arbuskula hidup hanya dalam jangka waktu beberapa hari sebelum terdisintegrasi (Alexopoulus et al. 1996; Smith dan Read 1997).
Peranan CMA
Peran dari CMA adalah membantu penyerapan hara sehingga dapat meningkatkan pertumbuhan tanaman. Dalam mendapatka n sumber karbohidrat, CMA memberikan keuntungan pada mitranya. Hifanya menyebar dalam tanah menyerap air, fosfor dan hara lainnya (Alexopoulus et al. 1996). CMA yang menginf eksi sistem perakaran tanaman inang akan memproduksi hifa secara intensif sehingga tanaman bermikoriza akan mampu meningkatkan kapasitasnya dalam menyerap unsur hara dan air (Brundrett et al. 1994). Hal ini juga didukung oleh Smith dan Read (1997), yang menyatakan bahwa mikoriza berpengaruh dalam meningkatkan penyerapan P, Zn, Cu, Ni, NH4+ dan kemungkinan NO3- .
Hasil penelitian Corryanti dan Rohayati (2000) menunjukkan bahwa terjadi respon pertumbuhan tanaman jati yang relatif lebih baik terlihat dari peningkatan tinggi, diameter dan berat keringnya. Namun terdapat perbedaan respon yang disebabkan oleh perbedaan isolat CMA yang diinokulasikan. Respon pertumbuhan terbaik dihasilkan oleh tanaman yang diinokulasi dengan jenis Glomus aggregatum, Mycofer, Aca ulospora sp. dan Glomus manihotis.
CMA mampu meningkatkan daya tahan tanaman terhadap serangan patogen tanah (Brundrett et al. 1994). Peranan CMA dalam menekan perkembangan patogen tanah terutama disebabkan kolonisasi awal pada perakaran tanaman sehingga mampu meningkatkan ketahanan tanaman. Secara normal CMA mampu meningkatkan penyerapan fosfor dan mineral hara lainnya sehingga peningkatan ketahanan tanaman merupakan efek tidak langsung pada peningkatan ketersediaan hara. Dengan demikian penurunan serangan penyakit diduga terdapat hubungan dengan peningkatan ketersediaan fosfor (Setiadi 2000). Selanjutnya Alexopoulus et al. (1996) menyatakan asosiasi mikoriza memberikan kontribusi untuk ketahanan dari serangan patogen akar dan nematoda dengan memproduksi antibiotik. Selaras dengan penelitian Bertha et al. (2005) menemukan bahwa kombinasi G. mosseae BEG 12 dan P. fluorescens A6RI efisien dalam menekan penyakit busuk akar
Rhizoctonia solani yang merupakan penyakit tular tanah.
Meningkatkan resistensi tanaman terhadap kekeringan adalah juga salah satu peran dari CMA (Auge 2001). Penelitian Por cel dan Ruiz -Lozano (2004) menemukan bahwa akar bermikoriza me ngakumulasi prolin yang lebih banyak
daripada akar tidak bermikoriza, sebaliknya pucuk tanaman bermikoriza mengakumulasi lebih sedikit prolin daripada pucuk tanaman tidak bermikoriza. Diduga simbiosis mikoriza tampaknya terlebih dulu meningkatkan regulasi osmotik di akar-akarnya, hal tersebut membantu mempertahankan landaian (gradient) potensial air yang memungkinkan untuk masuknya air dari tanah ke dalam akar. Kondisi demikian memungkinkan lebih tingginya potensial air daun pada tanaman bermikoriza selama periode kering dan dan melindungi tanaman terhadap cekaman oksidatif, dan pengaruh akumulatif tersebut meningkatkan toleransi tanaman terhadap kekeringan.
Vermikompos
Vermikompos (vermicompost) dihasilkan dari kemampuan beberapa cacing tanah dalam mengkonsumsi residu organik seperti limbah rumah tangga, limbah industri seperti bubur kayu, residu panen seperti sayuran, daun-daunan, dedak padi, dedak jagung, kotoran ternak, kompos dan sebagainya (Ndegwa et al. 1999; Palungkun 1999). Selanjutnya Nuryati (2004) menyatakan bahwa vermikompos berarti campuran kotoran cacing sebagai hasil buangan pencernaan bahan organik yang berwarna kehitam-hitaman berperan sebagai pupuk penyubur tanah. Vermikompos dapat meningkatkan hara dalam tanah karena mengandung nitrogen, fosfor , kalium dan unsur-unsur mikro seperti sulfur, boron, dan zinc, meningkatkan kapasitas tukar kation. Vermikompos juga mengandung berbagai hormon tumbuh bagi tanaman seperti auxin, sitokinin, giberellin (Nuryati 2004), menyediakan energi untuk aktivitas mikroorganisme, meningkatkan porositas tanah, meningkatkan kemampuan mengikat air, menstabilkan struktur ta nah seperti mengurangi pemadatan tanah, meningkatkan infiltrasi, dan menurunkan pengaruh logam-logam berat (Samosir 1994). Selanjutnya dinyatakan juga bahwa bahan organik mengurangi keracunan kation-kation seperti Al3+ dan Fe3+ pada tanah-tanah masam dan bereaksi dengan ion-ion racun seperti Cd2+ dan Hg2+ serta kation-kation unsur mikro lain yang berada pada konsentrasi tinggi dan mengurangi ketersediaannya, juga menyerap banyak air 70-80%. Ini juga disebabkan karena pori mikro pada agregat-agregat tanah menjadi lebih besar sehingga menambah kemampuan tanah untuk mengikat air dan mendukung pertumbuhan akar tanaman (Samosir 1994).
Marinari et al. (1999) , menunjukkan bahwa pada tanaman jagung (Zea mays), penambahan vermikompos dapat meningkatkan aktivitas enzim-enzim tanah yang menguntungkan seperti asam fosfatase, dehydrogenase dan protease BAA. Aktivitas enzim tersebut berkorelasi dengan sifat fisik tanah seperti porositas, yaitu meningkatkan pori makro dari 50-500 µm dan merangsang aktivitas biologi tanah.
Peran CMA dan vermikompos
Hasil-hasil penelitian tentang vermikompos terhadap peningkatan produktivitas tanaman telah banyak diteliti khususnya tanaman pertanian dan hortikultura akan tetapi penelitian tentang produksi inokulum CMA dengan formulasi media vermikompos belum banyak diteliti. Penelitian Cavender (2002), pada tanaman sorgum yang ditanam pada media gambut dan mineral menunjukkan bahwa vermikompos merangsang kolonisasi CMA pada akar sorgum walaupun hasilnya tidak konsisten. Penambahan vermikompos dapat meningkatkan bobot kering akar dan pucuk tanpa kehadiran CMA, sedangkan pemberian vermikompos tidak steril sebesar 20% dapat meningkatkan bobot kering akar dan pucuk pada media gambut.
Sebaliknya hasil penelitian Sáinz et al. (1998), pada dua jenis tanaman yang berbeda cengkeh merah dan ketimun yang diinokulasi dengan CMA jenis
Acaulospora menunjukkan bahwa terjadi perbedaan respon CMA dengan jenis
yang sama akan tetapi berbeda inang terhadap taraf pemberian vermikompos. Pada cengkeh merah, kolonisasi akar oleh CMA meningkat dan terbaik pada pemberian vermikompos 10% dari volume media jika dibandingkan dengan perlakuan lainnya. Selanjutnya Fitriatin et al. (2004), juga menunjukkan bahwa tanaman jagung manis yang ditumbuhkan pada tanah Ultisol dengan pemberian vermikompos 50 g/tan dan inokulasi CMA berpengaruh nyata terhadap derajat infeksi akar yang memiliki nilai tertinggi (72,00%), populasi total bakteri tanah (99,98 x 106 CFU/g tanah). Peningkatan juga terjadi pada bobot kering akar, bobot kering tanaman (113,13 g/pot) dan tinggi tanaman.
Perbanyakan inokulum dan potensi inokulum CMA
Kolonisasi akar dan pembentukan spora dapat berlangsung dengan optimal jika persyaratan dalam perbanyakan inokulum terpenuhi. Adapun tiga faktor utama yang
mempengaruhi pembentukan asosiasi CMA adalah jenis cendawan itu sendiri, lingkungan dan tanaman inangnya. Sumber inokulum yang digunakan dapat berupa spora, akar terinfeksi dan inokulum ta nah yang mengandung semua struktur cendawan mikoriza arbuskula baik spora maupun miselium eksternal dan miselium internal yang ada pada akar (Brundrett 1996).
Setiap spesies CMA mempunyai kemampuan yang berbeda-beda dalam
mengolonisasi akar, akan tetapi bagaimana pengaruhnya terhadap tanaman apakah sinergistik atau kompetitif sulit untuk ditentukan karena menunjukkan kesamaan dalam struktur morfologi (Gunawan 1993) . Klironomos dan Hart (2002), menyatakan bahwa perbedaan sumber inokulum yang digunakan dalam mengolonisasi akar menghasilkan perbedaan respon antara kelompok Glomineae dan
Gigasporineae. Isolat Glomus sp. dan Acaulospora sp. dapat mengolonisasi akar
dengan menggunakan seluruh tipe propagul sebaliknya isolat Gigaspora sp. dan
Scutellospora sp. yang diuji terbatas hanya pada spora dan akar terinfeksi.
Penelitian Basrudin (2005), menyatakan bahwa jenis inang berpengaruh terhadap kolonisasi dan pembentukan spora, hal ini kemungkinan berkaitan dengan eksudat akar. Jenis inang yang berbeda juga menghasilkan eksudat akar yang berbeda begitu pula dengan umur inang sehingga mempengaruhi pembentukan CMA. Dalam usaha mendapatkan hasil optimal dan infeksi yang efektif maka perlu adanya suatu isolat yang mampu hidup dan dapat beradaptasi dengan kondisi setempa t sesuai dengan tanaman lokal tersebut. Mansur (2002) mengemukakan bahwa isolasi CMA dari tanaman lokal akan lebih efektif untuk meningkatkan pertumbuhan tanaman lokal tersebut dari pada digunakan isolat dari luar daerah tersebut. Hal ini disebabkan karena CMA adalah mahluk hidup dengan daya adaptasi terhadap inang dan lingkungan yang relatif spesifik. Sehingga untuk mendapatkan hasil yang memuaskan sebaiknya digunakan isolat lokal terseleksi yang dapat dikembangkan sendiri. Penelitian Auge et al. (2004) me ndapatkan adanya pengaruh lingkungan, khususnya radiasi surya dan suhu terhadap respon tanaman terhadap CMA.
Penggunaan inokulum yang bermutu dapat meningkatkan efisiensi inokulum CMA dalam pengelolaan tanah dan tanaman. Simanungkalit (2004) , menyatakan bahwa potensi inokulum adalah kemampuan inokulan untuk mengolonisasi akar pada suatu kondisi tertentu. Potensi inokulum pada inokulan campuran sering dinyatakan
sebagai jumlah spora per satuan bobot inokulan tersebut. Padahal inokulan campuran tersebut juga mengandung hifa dan mikoriza yang memiliki kemampuan untuk mengolonisasi akar. Sehingga untuk menyatakan jumlah spora sebagai potensi inokulum kurang tepat karena tidak memperhitungkan apakah sporanya masih hidup, mati ataupun dorman. Dalam menentukan jumlah semua propagul CMA persatuan volume atau bobot tanah maka dapat dilakukan dengan Uji MPN (Most Probable
Number). Porter (1979) telah mengadaptasi metode MPN yang biasa digunakan
dalam memperkirakan jumlah organisme mikrobiologi untuk memperkirakan jumla h propagul CMA, kemudian metode ini dikembangkan lagi oleh Sieverding (1991).
Jati
Jati memiliki prospek yang baik, dimana produk berbahan baku kayu jati memiliki pangsa pasar luas karena termasuk kayu berkualitas tinggi, yaitu termasuk kedalam kelas kuat II dan kelas awet I-II, yang merupakan kayu yang paling banyak dipakai untuk segala jenis kontruksi seperti tiang, balok, jembatan, rangka atap, kosen pintu dan jendela, tiang, papan bendungan dalam air tawar, bantalan kereta api, sebagai wadah dalam produk industri kimia, dan kayu lapis (Pandit dan Ramdan 2002; Sumarna 2001). Tanaman jati juga tergolong sebagai tanaman yang berkhasiat obat, bunganya dapat digunakan sebagai obat bronchitis, membersihkan kantung kencing dan lain-lain. Buahnya dapat berfungsi sebagai obat diuretik, dan ekstrak daunnya dapat menghambat kinerja bakteri tuberkolosa. Daunnya dapat digunakan sebagai pewarna kain dan limbahnya dapat diproses menjadi briket arang (Sumarna 2001).
Penyebaran dan klasifikasi jati (Tectona grandis Linn f.)
Jati termasuk famili Verbenaceae, nama daerah jati (Indonesia); Sagun (India); Lyiu (Burma); Mai Sak (Thailand), Teak (Inggris), Teck (Perancis), Teca (Spanyol), Java Teak (Jerman). Jati memiliki persebaran yang cukup luas, meliputi sebagian besar India, Myanmar, Laos, Kamboja, bagian barat Thailand dan Indo-China. Jati telah dikembangkan di Afrika (Sudan, Kenya, Tanzania, Tanganyika, Uganda, Lower Guinea, Ghana, Nigeria, Afrika Barat), New Zealand, Australia, Kepulauan Fiji, Taiwan, Kepulauan Pasifi k. Di Benua Amerika, jati dikembangkan di Jamaica, Panama, Argentina, Puertorico,
Kepulauan Tobaqo, dan Suriname. Di Indonesia, jati tumbuh baik di Jawa Timur, Jawa Tengah dan Jawa Barat. Jati juga ditemukan dipulau-pulau kecil seperti Muna, Bawean, Kangean, Lombok, Sumbawa dan Bali. Batas-batas persebaran jati secara alamiah adalah 25o 30’ LU – 9o LS dan 73o BB – 103o 30’ BT (Mahfudz 2004; Tini dan Amri 2003).
Dalam sistem klasifikasi, tanaman jati mempunyai penggolongan yaitu: divisi : Spermatophyta ordo : Verbenales kelas : Angiospermae sub-kelas : Dicotyledoneae famili : Verbenaceae genus : Tectona
spesies : Tectona grandis Linn f.
Deskripsi buah dan benih jati
Buah keras, terbungkus kulit, berdaging lunak tidak merata (tipe buah batu). Ukura n buah bervariasi 5-20 mm, umumnya 11-17 mm. Struktur buah terdiri dari kulit luar tipis yang terbentuk dari kelopak, lapisan tengah (mesokarp) tebal seperti gabus, bagian dalamnya (endokarp) keras dan terbagi menjadi 4 ruang biji. Jumlah buah perkg bervar iasi sekitar 1100– 3500 butir, dengan rata -rata 2000 buah/kg. Benih jati berbentuk oval, ukuran kira-kira 6 x 4 mm. Jarang dijumpai dalam ke empat ruang berisi benih seluruhnya, umumnya hanya berisi 1– 2 benih, seringkali hanya satu benih yang tumbuh menjadi anakan (Rachmawati et al. 2002). Buah yang terbentuk akan masak sekitar bulan Nopember dan akan jatuh sekitar bulan Februari atau April. Buah jati termasuk ringan, antara 1,10– 2,80 g tergantung jenisnya (Sumarna 2001).
Tempat tumbuh jati
Secara umum, tanaman jati membutuhkan iklim dengan curah hujan minimum 750 mm/th, optimum 1000-1500 mm/th, dan maksimum 2500 mm/th (walaupun demikian, jati masih dapat tumbuh di daerah dengan curah hujan 3750 mm/th). Suhu udara optimal jati yaitu suhu udara minimum berkisar antara 13-17°C dan suhu maksimum berkisar antara 39-43°C. Adapun kondisi kelembaban
lingkungan tanaman jati yang optimal sekitar 80% untuk fase vegetatif dan antara 60-70% untuk fase generatif. Intensitas cahaya yang dibutuhkan cukup tinggi, 75-100% (Sumarna 2001; Mahfudz 2004)
Curah hujan secara fisik dan fisiologis berpengaruh terhadap sifat gugurnya daun deciduous dan kualitas produk kayu. Di daerah dengan musim kemarau panjang, jati akan menggugurkan daunnya dan lingkaran tahun yang terbentuk tampak artistik.
Secara geologis, tanaman jati tumbuh di tanah dengan batuan induk berasal dari berbagai jenis. Pertanaman jati akan tumbuh lebih baik pada lahan dengan kondisi fraksi lempung, lempung berpasir, atau pada lahan liat berpasir. Jati memerlukan kondisi solum lahan yang dalam dan keasaman tanah (pH) sekitar 6.0. Namun, ada kasus pada beberapa kawasan pertanaman jati dengan tingkat pH rendah (4-5), dijumpai tanaman jati dengan pertumbuhan yang baik. Karena tanaman jati sensitif terhadap rendahnya nilai pertukaran oksigen dalam tanah maka pada lahan yang berporositas dan memiliki drainase baik akan menghasilkan pertumbuhan baik (Sumarna 2001).
Unsur kimia pokok (makro element) yang penting dalam mendukung pertumbuhan jati yaitu N, P, K dan Ca. Tanaman jati akan tumbuh baik jika kandungan N pada tanah pada permukaan (top soil) antara 0,13– 0,072% dan pada lahan di bawahnya dengan ketebalan hingga 1 meter antara 0,0056 – 0,05%. Sedangkan rataan N yang dibutuhkan oleh tanaman jati sekitar 0,0039%, P antara 0,022– 0,108% atau 19-135 mg/100 g tanah, K berkisar 0,54– 1,80% (45-625 ppm/100 g) dan pada lahan di bawahnya (under top soil) antara 0,40– 1,13% (113-647 ppm/100 g), dan Ca harus lebih dari 9,27% (Sumarna 2001). Penelitian Restu (2006) mendapatkan ba hwa campuran perlakuan 2 g tepung tulang sapi, 2,5 g zeolit dan 0,5 g kaolin memberikan pengaruh terbaik terhadap peningkatan peubah pertumbuhan semai jati.
Adapun persyaratan bibit untuk reboisasi yang memenuhi kriteria mutu fisik dan fisiologi meliputi bibit normal adalah bibit berbatang tunggal, sehat dan pada pangkal batangnya berkayu, tinggi bibit diukur mulai dari pangkal batang sampai titik tumbuh teratas dihitung dengan satuan cm. Untuk tinggi batang dipersyaratkan antara 20-50 cm (MENHUT 2004).
J ati Muna
Jati Muna mulai dikembangkan permulaan awal abad ke 15, pada masa pemerintahan kerajaan Buton dengan Raja Sugi La Ende. Bibit jati diperoleh dari Jawa dan didatangkan oleh pelayar-pelayar dari Maluku. Pembudidayaan jati pertama kali di daerah Wasolangka dan kemudian populasinya menyebar sampai ke wilayah Muna (Saleh 1997). Pulau-pulau yang tersebar di Kabupaten Muna antara lain Matakidi, Wakuru, Raha dengan luas wilayah 12,81% . Wilayah Kabupaten Muna pada umumnya memiliki jenis tanah Mediteran , Ren sina dan
Litosol sehingga jati Muna tumbuh pada jenis-jenis tanah tersebut (BPS SULTRA
2004).
Populasi jati Muna tumbuh menyebar pada jenis tanah masam (pH rendah) sampai pada tanah-tanah berkapur. Informasi mengenai sifat kimia dari sebagian jenis-jenis tanah di daerah Muna disajikan pada Tabel 1.
Tabel 1. Hasil analisis sifat fisika dan kimia tanah beberapa lokasi penyebaran populasi jati Muna di Kabupaten Muna
Lokasi Peubah
Wakuru Matakidi Raha
pH (H2O) 4,9 6,2 5,8 C-Organik (%) 0,88 2,27 0,94 N 0,07 0,18 0,09 P tersedia (ppm) 12,75 10,65 2,40 K (me/100 g) 1,75 1,22 0,16 KTK(me/100 g) 5,01 13,58 5,71 Pasir (%) 51 15 67 Debu (%) 19 39 22 Liat (%) 30 46 11 Sumber : Haryanti 2005
Pada umumnya morfologi jati Muna sama dengan morfologi jati lainnya dan informasi mengenai morfologi yang lebih spesifik belum ada publikasi yang membahas secara detil.
METODOLOGI PENELITIAN
Waktu dan tempat
Penelitian ini dilaksanakan di rumah kaca Laboratorium Ekologi Hutan dan Laboratorium Silvikultur Fakultas Kehutanan IPB. Percobaan dilaksanakan pada bulan Juni 2005 sampai Juni 2006.
Bahan yang digunakan adalah benih Pueraria javanica , benih jati Muna (Tectona grandis L.f.), inokulum CMA jenis Glomus etunicatum terse leksi (eksotik) dengan kode NPI 126 (diperbanyak dari inokulum mycofer) di Laboratorium Silvikultur, inokulum CMA jenis Glomus sp. (endogenous) yang diisolasi dari bawah tegakan jati Muna (koleksi laboratorium Agronomi Fakultas Pertanian UNHALU Kendari) , vermikompos, zeolit, tanah Latosol, KOH 25%, HCl 2% (0,1 N), trypan blue, asam laktat, glyserol, larutan PVLG dan melzer, hyponex merah, gelas plastik berwarna, dan polibag.
Alat-alat yang digunakan adalah saringan spora (63µm, 125µm, 250µm , dan 500 µm), pinset spora, sentrifuse, timbangan analitik, oven, mikroskop binokuler Nikon YS100, mikroskop stereo binokuler Carton NSWT , Mikroskop Monookuler FCL 15 EX-N, kaca obyek dan gelas penutup.
Metode Penelitian
Penelitian terdiri dari dua tahap yang dilakukan secara berurutan yaitu :
Perbanyakan inokulum CMA
Media zeolit yang digunakan untuk mengecambahkan benih tanaman inang dicuci terlebih dahulu, disterilkan kemudian dimasukkan pada bak kecambah. Benih inang P. javanica direndam dengan klorox 5% selama ± 5 menit kemudian dicuci sampai bersih dengan air mengalir. Perendaman benih dengan air panas selama ± tiga menit kemudian dengan air dingin selama 24 jam. Selanjutnya benih dikecambahkan selama ± satu minggu atau sampai muncul 2 helai daun. Menyiapkan media tanam zeolit dan mencampur dengan vermikompos sesuai formulasi yang telah ditentukan. Media dimasukkan ke dalam gelas plastik berwarna yang sebelumnya telah dilubangi dibawahnya dan dilapisi lagi dengan gelas berisi zeolit yang tidak dilu bangi, berfungsi sebagai tempat air bagi kultur. Membuat lubang pada tengah media dan mengisi dengan inokulum sebanyak 10 g, kemudian tanaman P. javanica yang telah memiliki 2 - 3 helai daun dipindahkan dengan hati-hati, setelah itu lubang tadi ditutup kembali dengan zeolit. Penyiraman dilakukan setiap hari dan disesuaikan dengan kebutuhan tanaman. Khusus perlakuan tanaman inang dengan inokulasi CMA tanpa vermikompos pemberian larutan hara hyponex merah (25-5-20)
dilakukan seminggu sekali dengan konsentrasi 1 g/l air dan dibe rikan sebanyak 5 ml. Kultur disusun sesuai layout penelitian kemudian dipelihara selama tiga bulan di rumah kaca. Pemeliharaan kultur seperti penyiraman, penyiangan gulma dan pengendalian hama dilakukan secara manual. Setelah kultur berumur tiga bulan sejak inokulasi maka dilakukan pengecekan untuk mengetahui pertumbuhan dan perkembangan spora dan dilakukan pengeringan untuk merangsang pembentukan spora lebih banyak.
Gambar 2. Perbanyakan inokulum CMA menggunakan tanaman inang P. javanica selama tiga bulan di rumah kaca
Rancangan Penelitian
Penelitian ini merupakan percobaan faktorial dengan RAL menggunakan 2 faktor perlakuan. Faktor pertama yaitu jenis inokulum CMA yang terdiri dari tiga taraf : 1) tanpa inokulasi CMA (Mo), 2) inokulasi CMA jenis G.
etunicatum (Mb) dan 3) inokulasi CMA jenis Glomus sp. (Mk). Faktor kedua
adalah formulasi media dengan vermikompos terdiri dari K0 (100% zeolit), K1 (90% zeolit dicampur 10% vermikompos), K 2 (80% zeolit dicampur 20% vermikompos), K3 (70% zeolit dicampur 30% vermikompos), dan K4 (60% zeolit dicampur 40% vermikompos). Setiap kombinasi perlakuan diulang 3 kali dan setiap unit percobaan terdapat 5 pot kultur sehingga didapat 225 pot kultur. Adapun model statistik yang digunakan adalah
Yijk = µ + Mi + Kj + (MK )ij + eijk
Keterangan :
Yijk = Nilai pengamatan pada faktor M (jenis inokulum) taraf ke i, faktor K (formulasi media ) taraf ke j dan ulangan ke k
µ = Komponen aditif dari rataan
Mi = Pengaruh utama faktor M (jenis inokulum) pada taraf ke i Kj = Pengaruh utama faktor K (formulasi media ) pada taraf ke j
(MK)i j = Komponen interaksi dari faktor M (jenis inokulum) pada taraf ke i dan faktor K (formulasi media ) pada taraf ke j
eijk = Pengaruh acak yang menyebar normal (0,s2) (Mattjik dan Sumertajaya 2002)
Peubah yang diamati meliputi kolonisasi akar dan jumlah spora. Kolonisasi akar diukur berdasarkan keberadaan struktur CMA dalam akar, struktur CMA dapat dilihat dibawah mikroskop setelah dilakukan pewarnaan dengan trypan blue menggunakan metode Phillips dan Hayman (1970) yang dimodifikasi. Proses pewarnaan akar yaitu sampel akar dicuci bersih dari sisa-sisa tanah, merendam sampel akar dengan larutan KOH 2,5% selama 24 jam atau sampai akar kelihatan putih dan jernih, kemudian sampel akar dicuci bersih untuk menghilangkan larutan KOH, merendam dalam larutan HCl 2% (0,1 N) selama 24 jam dan membuang kelebihan HCl, selanjutnya merendam dengan larutan trypan blue 0,05% (campuran larutan asam gliserol, asam laktat, dan aquades) selama 24 jam.
Perhitungan kolonisasi akar dilakukan dengan cara mengambil secara acak potongan-potongan akar yang telah diwarnai sepanjang 1 cm, dan menyusun pada kaca obyek sebanyak 10 potongan akar, kemudian mengulangi sampai mendapatkan tiga preparat kaca obyek. Mengamati kolonisasi CMA dengan tiga bidang pandang dan mencatat total bidang pandang potongan akar yang terkolonisasi CMA dari 10 potongan akar tersebut. Selanjutnya persentase akar yang dikolonisasi CMA dihitung berdasarkan rumus :
% Kolonisasi CMA =
_ _ _ _ _
_ _ _
Jumlah bidang pandang yang terkolonisasi CMA
Jumlah total bidang pandang X 100%
(Rajapakse dan Miller 1992).
Menghitung jumlah spora pada akhir pengamatan setelah proses pengeringan tanaman inang selama seminggu. Pemisahan spora dilakukan dengan metode tuang saring basah (Brundrett 1994), yaitu mengambil sebanyak
20 g sampel inokulum pada pot kemudian disaring dengan memakai saringan 63µm, 125µm, dan 250µm. Saringan disusun dari ukuran terbesar hingga terkecil. Kemudian spora hasil saringan 63µm dipisahkan pada cawan plastik dan dilakukan perhitungan spora dibawah mikroskop stereo binokuler carton NSWT.
Peubah pertumbuhan yang diamati sebagai data pendukung meliputi bobot kering akar, bobot kering akar terinfeksi dan bobot kering total tanaman. Penimbangan bobot kering akar dan pucuk dilakukan pada akhir pengamatan, dengan cara menimbang bahan tanaman setelah dikeringkan dalam oven dengan suhu 70o C selama 2x24 jam atau sampai terjadi bobot kering yang konstan (Sitompul dan Guritno 1995). Bobot kering akar terinfeksi ditentukan dengan cara mengalikan bobot kering akar (g) dengan kolonisasi akar (%). Selanjutnya bobot kering total tanaman (g) ditentukan dengan cara menjumlahkan bobot kering akar dan pucuk.
Jumlah propagul CMA ditentukan berdasarkan metode MPN (The most
probable number ) (Porter 1979). Prosedur yang dilakuka n yaitu menyiapkan
media berupa pasir zeolit. Benih uji P. javanica disterilisasi dengan cara merendam dalam larutan klorox 5% selama ± 5 menit kemudian dibilas sampai bau klorox hilang dan mengecambahkan biji dalam bak kecambah.
Persiapan seri pengenceran medium dilakukan dengan cara menyiapkan seri pengenceran (dengan kelipatan 10) yaitu dengan mencampurkan contoh sampel uji dengan media zeolit. Untuk membuat seri pengenceran 10o yaitu sampel uji murni, 10-1 yaitu 1 bagian sampel uji murni dan 9 bagian zeolit, 10-2 yaitu 1 bagian sampel 10-1 dan 9 bagian zeolit dan seterusnya sampai pengenceran 10-7, setiap seri pengenceran dibuat 5 kali ulangan.
Penanaman kecambah pada pot yang telah berisi medium pertumbuhan sesuai dengan seri pengenceran, dan memupuk dengan larutan nutrisi hyponex merah sebanyak 1 g/l air seminggu sekali, selanjutnya memelihara pot kultur sampai ± 5 minggu
Gambar 3. Kegiatan pengujian potensi inokulum dengan menggunakan tanaman inang sorgum selama lima minggu di rumah kaca
Pemanenan dan pemrosesan akar yaitu dengan cara memotong bagian akar tanaman dan dicuci bersih kemudian dimasukkan kedalam botol vial yang berisi KOH 2,5 % dan direndam sampai akar kelihatan bersih dan jernih. Selanjutnya pemrosesan sampel akar sama dengan prosedur dalam pewarnaan akar. Memeriksa akar dibawah mikroskop dan mencatat pada tabel pengamatan bila ada infeksi d iberi tanda (+) dan bila tidak ada (-).
Cara perhitungan jumlah propagul yaitu dengan memilih tiga seri pengenceran yang menghasilkan kolonisasi akar, dimana P1 infeksi tertinggi, P2 dan P3 adalah yang jumlah infeksinya berturut -turut di bawah P1. Kemudian menentukan angka pada tabel MPN berdasarkan nilai P1, P2 dan P3 dan kombinasi dari angka dikali dengan faktor pengenceran P2. Selang kepercayaan 95 % dapat dihitung berdasarkan rumus :
Log Oa,b = log MPN ± 0.326
Uji efektivitas formulasi inokulum pada semai jati Muna
Persiapan benih
Benih jati yang digunakan adalah benih jati Muna . Benih diseleksi yaitu dengan cara memisahkan benih dari kotoran dan benih yang rusak, cukup kering, diamete r ± 1 cm, tidak terserang hama penyakit. Perlakuan benih berdasarkan metode Rizain (1999) dimodifikasi. Sebelum penyemaian, benih jati Muna dijemur kemudian direndam dalam air semalam, berturut-turut selama 4 hari kemudian direndam dalam abu sekam dengan perbandingan 1:0,7:1 (benih : abu sekam : air) selama 24 jam dan benih siap disemai.
Persiapan media perkecambahan dan media semai
Media perkecambahan benih menggunakan pasir yang telah dikeringkan dan diayak. Media pasir ditempatkan pada bak-bak kecambah dengan ketebalan 10 cm. Selanjutnya benih jati Muna ditanam satu persatu dengan pusar menghadap kebawah. Setelah itu benih ditutup dengan pasir tipis. Media tanam semai menggunakan tanah dan pasir yang dicampur dengan perbandingan (3:1), selanjutnya dimas ukkan dalam polibag dan ditimbang dengan berat 2 kg dengan ukuran polibag 20 x 20 cm.
Penyapihan dan Inokulasi CMA
Penyapihan dilakukan pada saat kecambah telah siap untuk disapih yaitu kecambah yang telah terbentuk dua daun pertama kira-kira umur 21 hari dan siap dipindahkan ke media polibag. Inokulasi dilakukan pada saat penyapihan, dengan cara memberikan formulasi inokulum CMA hasil perbanyakan sesuai perlakuan disekitar akar semai jati Muna. Kemudian semai diletakan dengan posisi akar persis mengenai inokulum yang diberikan dengan harapan pada saat spora berkecambah akan langsung menginfeksi akar. Inokulum yang akan digunakan adalah inokulum hasil perbanyakan dengan pemberian dosis yang berbeda yaitu 10 g, 15 g, dan 20 g per semai.
Pemeliharaan
Mela kukan penyiraman sesuai kebutuhan yaitu diperkirakan sampai mencapai kapasitas lapang. Pemberantasan hama penyakit juga dilakukan bila perlu.
Gambar 4. Penyapihan semai di polibag dan pemeliharaan selama tiga bulan di persemaian rumah kaca Laboratorium Ekologi Hutan, Fakultas kehutanan IPB
Rancangan Penelitian
Formulasi inokulum terpilih sebanyak enam kombinasi hasil perbanyakan tahap pertama diinokulasikan ke semai jati Muna dengan cara menaburkan formulasi inokulum pada lubang tanam sesuai dosis inokulum yang ditentukan. Kemudian semai dipindahkan ke lubang tanam dengan akar tepat mengenai inokulum tadi, sehingga diharapkan semai yang akan tumbuh akarnya langsung kontak dengan formulasi inokulum yang diberikan. Media tanah yang digunakan adalah jenis Latosol pada polibag dengan berat media 2 kg, kemudian disusun berdasarkan rancangan yang ditetapkan. Penelitian ini merupakan percobaan faktorial dengan RAK menggunakan 2 faktor perlakuan, faktor pertama yaitu formulasi inokulum CMA yang terdiri dari enam taraf yaitu formulasi G. etunicatum (A1), G. etunicatum dengan vermikompos 30% (A2), G. etunicatum dengan vermikompos 40% (A3), Glomus sp. (B1), Glomus sp. dengan vermikompos 30% (B2), Glomus sp. dengan vermikompos 40% (B3) dan kontrol. Faktor kedua adalah dosis formulasi inokulum CMA yang terdiri dari tiga taraf yaitu 10 g per semai (D1), 15 g per semai (D2) dan 20 g per semai (D3). Setiap kombinasi perlakuan diulang 3 kali sehingga didapat 57 unit percobaan. Peubah pertumbuhan yang diamati adalah tinggi semai, diameter batang, bobot kering semai (bagian pucuk dan akar), nisbah pucuk akar, kolonisasi akar dan jumlah spora. Adapun model statistik yang digunakan adalah
Yijk = µ + Mi + Dj + (MD)ij + ?k + eijk
Yijk = Nilai pengamatan pada faktor A adalah formulasi inokulum CMA taraf ke -i, faktor D (Dosis formulasi inokulum) taraf ke-j dan kelompok ke -k
µ = Komponen aditif dari rataan
Ai = Pengaruh utama faktor A (formulasi inokulum CMA ) pada taraf ke-i
Dj = Pengaruh utama faktor D (Dosis formulasi inokulum) pada taraf ke-j
(AD)ij = Komponen interaksi dari faktor A (formulasi inokulum CMA ) pada taraf ke-i dan faktor D (dosis formulasi inokulum) pada taraf ke-j
?k = Pengaruh aditif dari kelompok dan diasumsikan tidak berinteraksi dengan perlakuan
eijk = Pengaruh acak yang menyebar normal (0,s2) (Mattjik dan Sumertajaya 2002)
Pengamatan Pertumbuhan
Peubah pertumbuhan yang diamati dan diukur adalah tinggi semai, diameter batang, bobot kering semai, nisbah pucuk akar, kolonisasi akar dan jumlah spora. Peubah tinggi semai diukur dari pangkal batang (pada satu titik yang tetap dekat permukaan tanah) sampai titik tumbuh tertinggi semai pada jalur batang dengan menggunakan mistar , dan diukur 2 minggu sekali. Diameter batang diukur menggunakan jangka sorong pada ketinggian 1 cm dari permukaan tanah diukur pada awal dan akhir pengamatan. Bobot kering semai (bagian pucuk dan akar semai) dibersihkan kemudian dikeringkan dengan pengovenan pada suhu 70oC atau sampai tercapai bobot kering yang konstan kemudian ditimbang (Sitompul dan Guritno 1995). Nisbah pucuk akar ditentukan dengan membandingkan bobot kering pucuk dan bobot kering akar semai. Sebagai data pendukung yaitu menghitung kolonisasi CMA pada akhir penelitian, dimana prosedurnya sama dengan proses pewarnaan akar dengan metode Phillips dan Hayman (1970) dimodifikasi.
Pengamatan jumlah spora dilakukan dengan prosedur yaitu tanah dalam polibag dibongkar dan mengambil contoh tanah seberat 40 g. Sampel tanah dimasukkan ke dalam gelas kemudian direndam dan diaduk agar spora yang melekat pada partikel tanah dapat terlepas. Setelah tanah diaduk kemudian dituang dalam saringan bertingkat (63µm, 125µm, 250µm dan 500µm) yang
disusun dari paling terbesar sampai saringan terkecil. Kemudian saringan tadi disemprot dengan air mengalir dan diusahakan supaya bertekanan tinggi untuk melepaskan spora dari partikel tanah. Hasil saringan 63 µm diambil dan dimasukkan kedalam tabung sentrifus dan diberi larutan gula 50% dan disentrifus selama 5 menit dengan kecepatan 3200 rpm. Kemudian larutan supernatan pada bagian tengah diambil dengan memakai pipet dan dicuci dibawah air mengalir dengan saringan 63µm. Hasil saringan diambil dan dituang ke cawan petri kemudian di hitung dibawah mikroskop stereo binokuler Carton NSWT.
Pengamatan penunjang meliputi analisis media semai jati, analisis hara inokulum yang digunakan, temperatur udara rumah kaca selama penelitian, analisis jaringan daun semai jati untuk N, P, K, dan Ca diakhir penelitian.
Analisis Data
Data hasil pengamatan dari masing-masing peubah dilakukan analisis sidik ragam. Jika hasil analisis sidik ragam menunjukkan F hitung > dari F tabel maka dilanjutkan dengan uji jarak berganda Duncan pa da taraf kepercayaan 95% untuk membandingkan antar perlakuan (Mattjik dan Sumertajaya 2002). Pengolahan data dilakukan dengan menggunakan program komputer SAS V6.12 (Statistical Analysis
Hasil dan Pembahasan
Hasil Perbanyakan inokulum CMA
Jenis CMA, formulasi media vermikompos dan interaksi antara jenis CMA dengan formulasi media berpengaruh nyata terhadap kolonisasi akar tanaman inang
P. javanica , jumlah spora CMA, bobot kering akar, bobot kering akar terinfeksi, dan
bobot kering total (Tabel 2).
Tabel 2. Rekapitulasi hasil analisis sidik ragam terhadap kolonisasi akar, jumlah spora, dan pertumbuhan tanaman inang P. javanica
Peubah CMA Vermikompos CMA x
vermikompos KK (%)
kolonisasi akar 75,97*** 19,78*** 6,05** 18,35 %
Jmlh spora 55,04*** 26,52*** 9,72*** 17,32 %
Bobot kering akar 11,45*** 37,59*** 7,15*** 12,88 % Bobot kering akar
terinfeksi 14,37
** 9,04*** 3,20* 30,90 %
Bobot kering total 78,47*** 159,09*** 5,43*** 7,45 % Keterangan : ***) berpengaruh sangat nyata pada P=0.001, **) berpengaruh nyata pada P=0.01, *)
berpengaruh nyata pada P=0.05
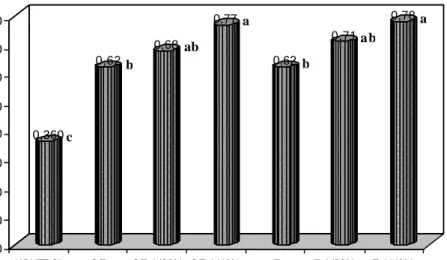
Kolonisasi akar dan jumlah spora
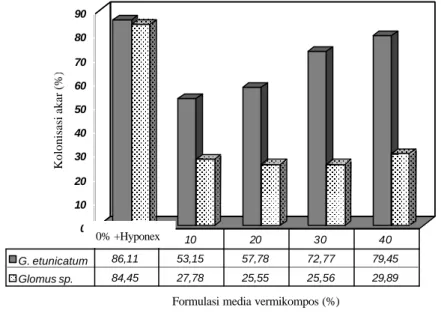
Hasil uji lanjut pengaruh interaksi antara jenis CMA dan formulasi media vermikompos terhadap kolonisasi akar inang P. javanica menunjukkan bahw a perlakuan terbaik dihasilkan oleh jenis CMA G. etunicatum dengan pemupukan hyponex merah. Kemudian diikuti oleh jenis Glomus sp. dengan hyponex merah dan jenis G. etunicatum dengan vermikompos 40%. Penurunan yang sangat tajam terjadi pada jenis Glomus sp. akibat pemberian vermikompos (Gambar 5).
Pada peubah jumlah spora, perlakuan terbaik dihasilkan oleh jenis Glomus sp. dengan hyponex merah yang berbeda nyata dengan perlakuan lainnya. Data menunjukkan bahwa terjadi penurunan yang sangat signifikan pada jumlah spora kedua jenis CMA seiring dengan penambahan vermikompos (Gambar 6). Akan tetapi jenis G. etunicatum mampu menghasilkan kolonisasi akar dan jumlah spora
yang lebih baik dibandingkan jenis Glomus sp. Hal ini diduga karena G.
etunicatum lebih toleran terhadap penambahan vermikompos (Gambar 5 dan 6).
0 10 20 30 40 50 60 70 80 90 Kolonisasi akar (%)
Formulasi media vermikompos (%)
G. etunicatum 86,11 53,15 57,78 72,77 79,45 Glomus sp. 84,45 27,78 25,55 25,56 29,89
0 10 20 30 40
Gambar 5. Pengaruh jenis inokulum CMA dan formulasi media vermikompos terhadap kolonisasi akar tanaman inang P. javanica pada bulan ketiga
0 100 200 300 400 500 600 700 800 900 Jumlah spora
Formulasi media vermikompos (%)
G. etunicatum 345 124 95 148 95
Glomus sp. 889 13 8 12 10
0 10 20 30 40
Gambar 6. Pengaruh jenis inokulum CMA dan formulasi media vermikompos terhadap jumlah spora CMA tanaman inang P. javanica pada bulan ketiga
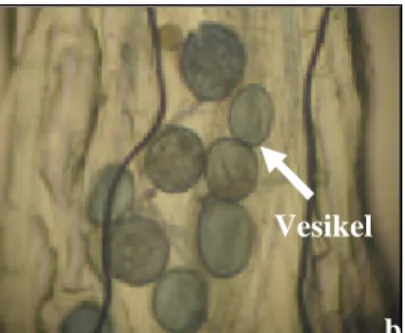
0% +Hyponex
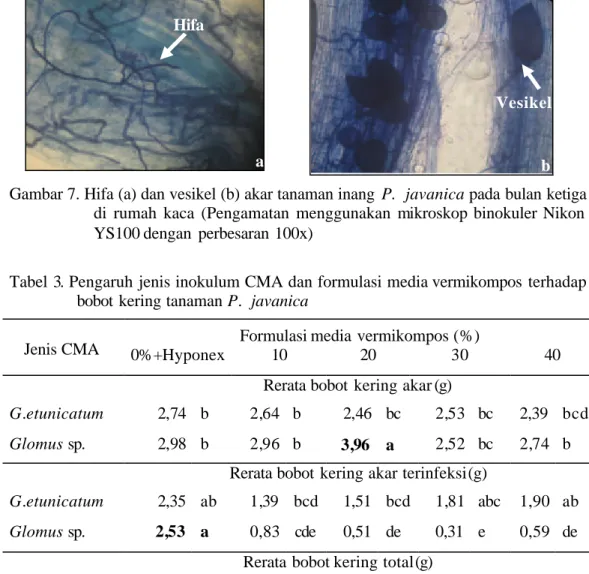
Gambar 7. Hifa (a) dan vesikel (b) akar tanaman inang P. javanica pada bulan ketiga di rumah kaca (Pengamatan menggunakan mikroskop binokuler Nikon YS100 dengan perbesaran 100x)
Tabel 3. Pengaruh jenis inokulum CMA dan formulasi media vermikompos terhadap bobot kering tanaman P. javanica
Formulasi media vermikompos (%)
Jenis CMA 0%+Hyponex 10 20 30 40
Rerata bobot kering akar (g)
G.etunicatum 2,74 b 2,64 b 2,46 bc 2,53 bc 2,39 bcd
Glomus sp. 2,98 b 2,96 b 3,96 a 2,52 bc 2,74 b Rerata bobot kering akar terinfeksi (g)
G.etunicatum 2,35 ab 1,39 bcd 1,51 bcd 1,81 abc 1,90 ab
Glomus sp. 2,53 a 0,83 cde 0,51 de 0,31 e 0,59 de Rerata bobot kering total (g)
G.etunicatum 11,93 d 14,41 c 16,77 b 16,91 b 16,28 b
Glomus sp. 11,33 de 15,61 bc 18,74 a 16,95 b 15,41 bc Keterangan : Rerata sebaris dan sekolo m diikuti huruf sama menunjukkan tidak berbeda nyata
berdasarkan uji jarak berganda Duncan pada taraf kepercayaan 95%
Jenis Glomus sp. dengan penambahan vermikompos 20%, menghasilkan bobot kering akar tertinggi yang berbeda nyata dengan perlakuan lainnya , dan bobot kering akar tanaman inang terendah dihasilkan oleh jenis G. etunicatum dengan vermikompos 40%. Jenis Glomus sp. dengan pemupukan hyponex merah menghasilkan bobot kering akar terinfeksi tertinggi, sedangkan terendah dihasilkan oleh jenis Glomus sp. dengan penambahan vermikompos 30%. Bobot kering tanaman tertinggi dihasilkan oleh perlakuan jenis Glomus sp. pada penambahan vermikompos 20% dan terendah adalah jenis Glomus sp. pada pemupukan dengan hyponex merah (Tabel 3).
a b
Hifa
Pengujian potensi inokulum CMA
Tabel 4. Potensi inokulum cendawan mikoriza arbuskula per 100 g media zeolit Jumlah pr opagul infektif (KA 10%)
Perlakuan Jumlah/100 g
zeolit
Kisaran jumlah propagul *)
G. etunicatum
G. etunicatum dengan vermikompos 30% G. etunicatum dengan vermikompos 40% Glomus sp.
Glomus sp. dengan vermikompos 30% Glomus sp dengan vermikompos 40%
308 x 104 7,7 x 104 5,4 x 104 10120 x 104 0,12 x 104 0,19 x 104 145 – 652 x 104 3,6 – 16,3 x 104 0,50 – 2,50 x 104 4770 – 21440 x 104 0,41 – 0,57 x 104 0,88 – 3,96 x 104 Keterangan : *) Kisaran jumlah propagul pada selang kepercayaan 95 %
Jenis Glomus sp. menghasilkan jumlah propagul terbanyak yaitu 10120 x 104 propagul dan terendah dihasilkan oleh Glomus sp. dengan vermikompos 30% yaitu 0,12 x 104 propagul (Tabel 4).
Uji efektivitas formulasi inokulum CMA pada semai jati Muna
Hasil analisis sidik ragam pada peubah pertumbuhan semai jati Muna menunjukkan bahwa formulasi inokulum CMA berpengaruh nyata terhadap tinggi semai, diameter, bobot kering puc uk, bobot kering akar, bobot kering akar terinfeksi, bobot kering total, nisbah pucuk akar, kolonisasi akar semai jati Muna dan jumlah spora CMA . Dosis formulasi inokulum juga berpengaruh nyata terhadap tinggi semai jati, diameter dan bobot kering pucuk. Sedangkan interaksi antara jenis formulasi inokulum CMA dan dosis formulasi inokulum berpengaruh nyata terhadap kolonisasi akar semai jati Muna dan jumlah spora CMA (Tabel 5).
Tabel 5. Rekapitulasi hasil analisis sidik ragam terhadap peubah pertumbuhan, kolonisasi akar dan jumlah spora semai jati Muna
Peubah CMA DOSIS CMA x DOSIS KK (%)
Tinggi semai 2 MST 0,31tn 0,40 tn 0,68 tn 23,45 Tinggi semai 4 MST 1,72 tn 1,20 tn 1,70 tn 20,80 Tinggi semai 6 MST 3,15** 4,41 ** 1,25 tn 20,11 Tinggi semai 8 MST 5,37 *** 4,45 ** 1,27 tn 18,03 Tinggi semai 10 MST 5,65 *** 4,99 ** 1,55 tn 14,41 Tinggi semai 12 MST 4,99 *** 6,57 *** 1,40 tn 12,81 Diameter semai 4,87 ** 4,41 ** 1,01 tn 14,11
Bobot kering pucuk 8,32 *** 3,20 ** 0,34 tn 26,74 Bobot kering akar 11,56 *** 1,40 tn 0,42 tn 27,80 Bobot kering akar
terinfeksi 11,12
***
0,54 tn 0,81 tn 19,36
Bobot kering total 10,36 *** 2,67 tn 0,22 tn 25,62 Nisbah pucuk akar 4,79 *** 2,15 tn 1,86 tn 8,48
kolonisasi akar 4,77 *** 1,04 tn 3,60 *** 15,04
Jmlh spora 9,32 *** 1,24 tn 2,76 ** 7,53
Keterangan : ***) berpengaruh sangat nyata pada P=0.001, **) berpengaruh nyata pada P=0.01, *) berpengaruh nyata pada P=0.05, tn) berpengaruh tidak nyata pada P>0.05
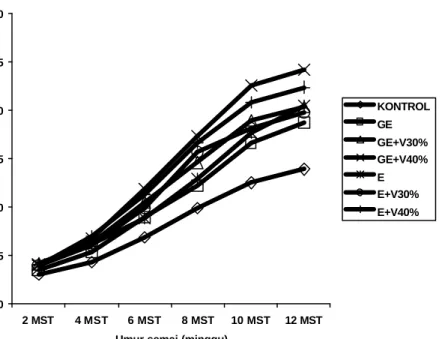
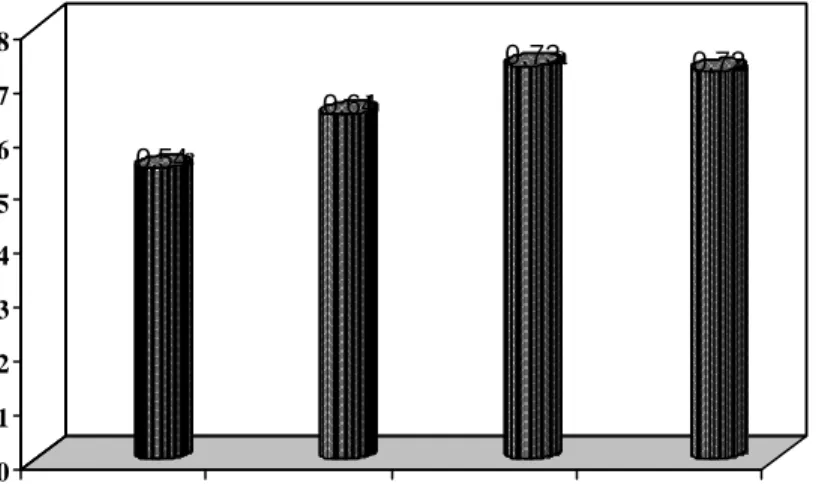
Pertumbuhan semai jati Muna
Secara umum inokulasi CMA cenderung memberikan respon pertumbuhan semai jati Muna yang lebih baik dibanding tanpa pemberian CMA (kontrol). Perlakuan formulasi inokulum CMA berpengaruh nyata terhadap tinggi dan diameter semai jati pada saat berumur 6 MST. Perlakuan formulasi inokulum G.
etunicatum dengan vermikompos 40% menghasilkan tinggi semai jati Muna yang
tidak berbeda nyata dengan formulasi inokulum Glomus sp. dengan vermikompos 40%. Sedangkan perlakuan tanpa formulasi inokulum CMA (kontrol) menghasilkan pertumbuhan semai jati Muna terendah (Gambar 8).