II-202
KEANEKARAGAMAN KEONG AIR TAWAR MARGA
FILOPALUDINA
DI
INDONESIA DAN STATUS TAKSONOMINYA
(GASTROPODA: VIVIPARIDAE)
oleh
Ristiyanti M. Marwoto * & Ayu S. Nurinsiyah **
(*Puslit Biologi-LIPI,Gedung Widyasatwaloka Jl.Raya Cibinong KM 46-** Pasca Sarjana Universitas Padjadjaran,Jatinangor)
ABSTRAK
Keong air tawar yang biasa disebut keong tutut (viviparid) sangat umum dikenal, merupakan jenis yang menyebar luas di kawasan perairan tawar di Asia Tenggara. Di Indonesia, khususnya di pulau Jawa dan Sumatera keong ini biasa dimakan oleh penduduk setempat. Keong ini menyebar luas pada beberapa tipe habitat, baik sungai, sawah, danau, kolam, rawa yang berair tenang maupun berair deras. Di Indonesia tercatat 15 jenis yang dijumpai di Jawa, Sumatera, Kalimantan, Sulawesi dan Papua. Jenis Filopaludina javanica menyebar luas hampir di semua pulau, sedangkan jenis F. costata, F. persculpta, F. crassibucca, F. lutulenta, F. rudipelis merupakan jenis-jenis endemik Sulawesi. Tercatat pula 8 jenis dari Papua yang diduga juga terbatas sebarannya. Kelompok ini termasuk suku (Family) Viviparidae dan selama ini semua jenisnya digolongkan pada marga (Genus) Bellamya. Status taksonomi kelompok ini sedang dipelajari berdasarkan karakter morfologi dan anatominya. Hasil pengamatan pendahuluan terlihat bahwa jenis – jenis dari Jawa berkelompok dengan jenis-jenis dari Sumatera dan Kalimantan dan terpisah dari jenis – jenis yang berasal dari Sulawesi dan Papua.
PENDAHULUAN
Keong air tawar genus Filopaludina Habe, 1964 yang termasuk dalam suku (Family) Viviparidae, merupakan jenis keong yang umum dikenal di Asia dan Asia Tenggara. Di Indonesia biasa disebut keong tutut, dijumpai menyebar luas hampir di berbagai tipe habitat, seperti sungai, rawa, danau, sawah, kolam baik yang berarus tenang maupun deras. Keong ini juga biasa dikonsumsi masyarakat terutama di daerah Jawa dan Sumatera namun berpotensi pula sebagai inang antara cacing Trematoda (Echinostoma) yang dapat menyebabkan penyakit echinostomiasis (Van Benthem Jutting, 1956; Berry, 1974; Sulianti, 2006).
II-203
Filopaludina penyebarannya luas meliputi daerah tropis dan subtropis, di Indonesia banyak terdapat di pulau-pulau besar seperti Pulau Sumatera, Jawa, Kalimantan, Sulawesi dan Papua. Van Benthem Jutting (1956) mengemukakan dua jenis yang disebutnya sebagai Bellamya javanica yang menyebar luas terutama di Jawa, Sumatera, Kalimantan, Sulawesi dan B. sumatrensis yang hanya dilaporkan menyebar di Jawa dan Sumatra. Di Indonesia tercatat ada 15 jenis keong tutut.. Lima jenis yang dilaporkan Sarasin & Sarasin (1898) merupakan jenis endemik Sulawesi, yakni yang disebutnya Vivipara costata, V. persculpta, V. crassibucca, V. lutulenta, V. rudipellis, empat jenis terakhir merupakan jenis baru yang dideskripsi oleh Sarasin & Sarasin (1898), sedangkan satu jenis oleh Quoy & Gaimard pada tahun 1832. Van Benthem Jutting (1963) melaporkan ada 8 jenis yang disebutnya sebagai Bellamya decipiens decipiens (Taparone, 1883), B. decipiens aruana (Boettger, 1915), B. kowiayiensis (Brazier, 1886), B. wisseli Van Benthem Jutting, 1963, B. tricostata tricostata (Lesson, 1831), B. tricostata elegans (Bavay, 1908), B. tricostata multifuniculata (Bavay, 1908), B. fragilis (Preston, 1910).
Problem taksonomi pada kelompok viviparid ini belum terpecahkan, terutama pengelompokan dalam marga (Genus) yang belum mantap karena hanya berdasarkan ukuran,bentuk dan hiasan (sculpture) cangkang. Oleh karena itu, karakter morfologi dan anatomi yang lebih lengkap perlu diamati untuk melengkapi deskripsi jenis – jenis yang dijumpai di Indonesia dan memantapkan status taksonominya.
BAHAN DAN METODE
Bahan penelitian adalah spesimen keong tutut koleksi Museum Zoologicum Bogoriense (MZB), Puslit Biologi, LIPI. Pengumpulan data dilakukan dengan mengamati dan mendeskripsi karakter morfologi cangkang, embrio dan radula, baik secara kualitatif maupun kuantitatif (Hershler & Ponder, 1998; Brandt, 1974; Bieler, 1988). Data kualitatif diperoleh dengan melakukan pengamatan bagian-bagian cangkang dewasa (Gambar 1) dan radula (Heryanto dkk., 2003; Bandel, 1984).
Data kuantitatif diperoleh dengan melakukan pengukuran cangkang dewasa (Mimpfoundi & Ndassa, 2005), pengamatan radula (Bieler, 1988)dan penghitungan embrio dalam setiap induk. Karakter cangkang dewasa yang terukur dikelompokkan dan diuji dengan analisis kluster menggunakan program SPSS 12.0. Pengamatan
II-204
radula melalui beberapa tahapan, yaitu pembuatan enzim dan pembedahan, kemudian diamati dengan Scanning Electron Microscope (SEM) (Jones et al., 1998a).
HASIL DAN PEMBAHASAN
Status taksonomi dari kelompok viviparid di Indonesia belum mantap. Pengelompokan sebagai marga (Genus) Bellamya oleh van Benthem Jutting (1956,1959 & 1963) berdasarkan tipe yang berasal dari Afrika. Brandt (1968) merevisi viviparid Thailand dan mendeskripsi marga baru Siamopaludina yang kemudian pada tahun 1974 dijadikannya sebagai subgenus dari Filopaludina. Dalam revisinya tersebut, Brandt mengelompokkan jenis B. javanica dan B. sumatrensis kedalam marga Filopaludina Habe, 1964, masing – masing sebagai Filopaludina (Siamopaludina) javanica (von dem Busch, 1844) dan F. (Filopaludina) sumatrensis (Dunker, 1852). Dia juga mengelompokkan jenis – jenis yang memiliki rusuk lingkar ke dalam marga Sinotaia Haas, 1939. Berdasarkan morfologi cangkang dewasa, jenis - jenis yang ada di Indonesia secara umum terbagi 3 yakni kelompok yang memiliki karakter morfologi marga Filopaludina (tanpa rusuk lingkar) , marga Sinotaia(memiliki rusuk lingkar agak tebal), dan marga yang bukan keduanya (cangkang dan rusuk lingkar sangat tebal). Marga yang bukan keduanya diwakili oleh populasi yang hanya dijumpai dari danau Poso yakni yang dideskripsi oleh Sarasin & Sarasin (1898) sebagai Vivipara persculpta. Karena studi revisi viviparid Indonesia belum selesai dilakukan, maka dalam makalah ini untuk sementara semua jenis viviparid dikelompokkan pada marga Filopaludinam termasuk dalam dendogram yang masih menggunakan nama genus Bellamya.
Pengamatan karakter cangkang dilakukan terhadap 35 nomor koleksi yang berasal dari Jawa, Sumatra, Kalimantan, Sulawesi dan Papua. Karakter cangkang dewasa yang diamati, diukur dan dihitung adalah, ukuran cangkang (tinggi, lebar cangkang dan tinggi seluk akhir (body whorl), tinggi, lebar mulut cangkang (aperture)), jumlah seluk (whorl), bentuk seluk, jumlah rusuk lingkar (spiral ribs), bentuk mulut cangkang, tutup cangkang (Gambar 1).
II-205
Gambar 1. Bagan pengamatan dan pengukuran keong Filopaludina (Foto: Nurinsiyah, 2008)
Jumlah anakan (embrio dalam tubuh induk betina) dihitung dan diamati bentuknya. Panjang radula diukur dan dihitung baris giginya, diamati bentuk dan susunannya (Gambar 2).
Gambar 2. Gambar rangkaian radula (gigi) Filopaludina dengan SEM (Foto: Dewi & Nurinsiyah, 2008)
Hasil pengamatan cangkang dewasa dan analisanya (Gambar 3) menunjukkan bahwa dari ke 35 nomor spesimen, terkelompok menjadi 3 kelompok, kelompok I (25 nomor koleksi) yang berasal dari Sumatra, Jawa, Kalimantan, Sulawesi dan Papua Barat . Kelompok II ( 9 nomor koleksi) berasal dari Jawa, Sulawesi dan Papua, dan kelompok III (1 nomor koleksi) berasal dari Sulawesi.
lateral
marginal
rachis
II-206
Hasil identifikasi berdasarkan karakter cangkang, menunjukkan bahwa kelompok I terdiri dari F. javanica (Jawa, Kalimantan, Sumatera, Sulawesi, Papua Barat), F. sumatrensis ( Jawa dan Sumatera), kelompok II terdiri dari F. costata, F. crassibucca, F. rudipelis, F.javanica, F. decipiens, F. tricostata, Filopaludina sp. ( Sulawesi dan Papua). Sedangkan kelompok III diduga merupakan jenis baru dari Sulawesi yang terpisah dari kelompok I dan kelompok II. Secara umum hasil pengelompokkan ini terlihat dari Gambar 3.
Hasil identifikasi juga menunjukkan bahwa no koleksi 3,10,11 adalah jenis F. sumatrensis yang menurut pengamatan Van Benthem Jutting (1956) memang berbeda dengan jenis yang disebutnya Bellamya javanica. Dalam penelitian ini karakter pembeda antara keduanya yang utama adalah adanya garis lingkar, perbedaan jumlah seluk dan ukuran rata-rata cangkang. Jenis F. javanica umumnya memiliki garis lingkar sedang dan ukuran cangkangnya relatif lebih besar dibandingkan F. sumatrensis, namun ukuran embryonic shell nya lebih kecil. Jumlah embryonic shell pada F. javanica lebih banyak ( rata-rata 14 /induk) dibandingkan F. sumatrensis (rata-rata 12/induk). Ukuran radula pada F. javanica rata – rata lebih panjang ( > 3mm) dari F. sumatrensis ( < 3mm), demikian pula jumlah baris gigi pada F. javanica lebih besar , yakni 115 baris, sedangkan pada F. sumatrensis hanya 96 baris.
II-207
Gambar 3. Dendogram kekerabatan Filopaludina di Indonesia berdasarka Karakter cangkang dewasa (Nurinsiyah, 2008).
II-208
Dendogram kekerabatan (Gambar 3) juga menunjukkan bahwa no koleksi 32 dari Papua Barat, masuk dalam Kelompok I, sejajar dengan koleksi yang memiliki karakter jenis F. javanica. Hal ini merupakan catatan baru kehadiran F. javanica di Papua Barat yang sebelumnya tidak pernah dilaporkan. Dari kelompok II, koleksi yang teridentifikasi adalah yang berasal dari Sulawesi, yakni jenis F. costata (koleksi no 24), F. crassibucca (koleksi no 25), Filopaludina lutulenta (koleksi no 26), Filopaludina sp.(koleksi no 27), sedangkan koleksi no 28 karakternya lebih mirip dengan F. costata dari Tondano. Jenis - jenis yang berasal dari Papua belum diidentifikasi dengan lengkap karena keterbatasan kunci identifikasi. Terpisahnya jenis – jenis tersebut dalam kluster perlu di pelajari lebih lanjut dengan melengkapi karakter anatomi seperti sistem reproduksi, sistem pernafasan, dan karakter-karakter pada sistem pencernaan.
Karakter embryonic shell merupakan salah satu karakter diagnostik untuk mengidentifikasi suatu spesies (Hadžišce et. al., 1976; Rintelen et. al, 2007). Hasil pengamatan embrio pada penelitian ini menunjukkan adanya karakter pembeda yang jelas yaitu keberadaan rusuk lingkar dan jumlah embrio dalam satu kantung eram. Namun karakter tersebut belum dapat digunakan sebagai diagnostik utama pembeda antara jenis F. sumatrensis dan F. javanica. Hal ini terjadi pula pada , pengungkapan kekerabatan jenis Tylomelania patriarchalis dan T. mattanensis yang dilakukan Rintelen et. al, (2007). Hasil penelitiannya menunjukkan terdapat variasi cangkang embryonic shell yang tinggi dalam brood pouch (kantung eram) pada induk yang sama. Sebaliknya Hadžišce et. al (1976) mengungkapkan adanya kesamaan pola embryonic shell pada Valvata hirsutecostata dan Valvata relicta dari Danau Ohrid Yugoslavia dengan Valvata tricarinata perconfusa dan Valvata humeralis dari Amerika Utara. Walaupun tidak dapat membedakan spesies, karakter embrio dapat digunakan untuk membedakan genera (Rintelen et. al, 2007). Selajutnya Dudgeon (1989) dalam Köhler & Glaubrecht (2003) melakukan penelitian terhadap Brotia hainanensis dan menyimpulkan bahwa variasi jumlah embrio berhubungan dengan adanya variasi pada kondisi habitat, keadaan nutrisi individu dan ukuran atau umur induk betina. Dalam penelitian ini spesimen yang diperiksa juga berasal dari beberapa tipe habitat yang berbeda sehingga variasi jumlah embriopun cukup tinggi.
II-209
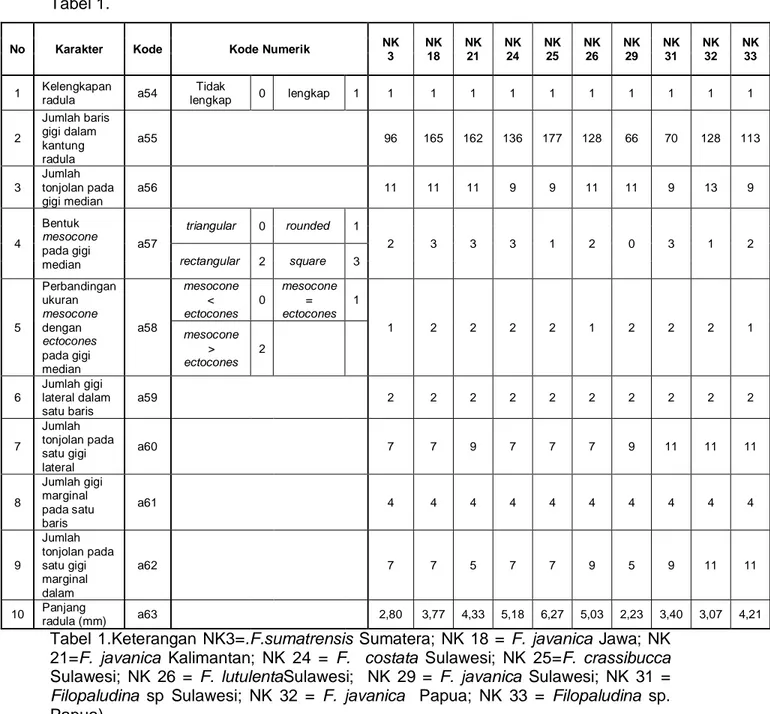
Tabel 1.
No Karakter Kode Kode Numerik NK 3 NK 18 NK 21 NK 24 NK 25 NK 26 NK 29 NK 31 NK 32 NK 33 1 Kelengkapan radula a54 Tidak lengkap 0 lengkap 1 1 1 1 1 1 1 1 1 1 1 2 Jumlah baris gigi dalam kantung radula a55 96 165 162 136 177 128 66 70 128 113 3 Jumlah tonjolan pada gigi median a56 11 11 11 9 9 11 11 9 13 9 4 Bentuk mesocone pada gigi median a57 triangular 0 rounded 1 2 3 3 3 1 2 0 3 1 2 rectangular 2 square 3 5 Perbandingan ukuran mesocone dengan ectocones pada gigi median a58 mesocone < ectocones 0 mesocone = ectocones 1 1 2 2 2 2 1 2 2 2 1 mesocone > ectocones 2 6 Jumlah gigi lateral dalam satu baris a59 2 2 2 2 2 2 2 2 2 2 7 Jumlah tonjolan pada satu gigi lateral a60 7 7 9 7 7 7 9 11 11 11 8 Jumlah gigi marginal pada satu baris a61 4 4 4 4 4 4 4 4 4 4 9 Jumlah tonjolan pada satu gigi marginal dalam a62 7 7 5 7 7 9 5 9 11 11 10 Panjang radula (mm) a63 2,80 3,77 4,33 5,18 6,27 5,03 2,23 3,40 3,07 4,21
Tabel 1.Keterangan NK3=.F.sumatrensis Sumatera; NK 18 = F. javanica Jawa; NK 21=F. javanica Kalimantan; NK 24 = F. costata Sulawesi; NK 25=F. crassibucca Sulawesi; NK 26 = F. lutulentaSulawesi; NK 29 = F. javanica Sulawesi; NK 31 = Filopaludina sp Sulawesi; NK 32 = F. javanica Papua; NK 33 = Filopaludina sp. Papua)
Walaupun dalam satu baris terdapat jumlah gigi yang sama, namun jumlah baris yang terdapat dalam satu kantung radula berbeda-beda. F. crassibucca dari habitat danau (Sulawesi) memiliki jumlah baris gigi yang paling banyak (177 baris gigi) dan radula terpanjang (6,27 mm) dibandingkan dengan 9 koleksi lainnya. Sedangkan F. javanicadari habitat sungai (Sulawesi) memiliki jumlah baris gigi yang paling sedikit (66 baris gigi) dan radula terpendek (2,23 mm) dibandingkan dengan koleksi lainnya.
II-210
Berdasarkan karakter-karakter radula yang menunjukan persamaan dan perbedaan tersebut, kesepuluh koleksi kemudian dianalisis untuk mengetahui tingkat kekerabatannya. Hasil analisis kekerabatan radula Filopaludina menunjukkan bahwa kekerabatan radula Filopaludina dipengaruhi oleh habitat. Radula Filopaludina yang menempati perairan lotik (habitat berair mengalir cepat) saling berkerabat dekat. Begitu pula halnya dengan radula Filopaludina yang menempati perairan lentik (habitat akuatik yang menggenang, tenang atau hanya terjadi sedikit aliran) saling berkerabat dekat. Keduanya saling membentuk kelompok walaupun jenisnya berbeda dan berasal dari pulau yang berbeda pula.
Troschel (1856-1863) dalam Bandel (1984) menyatakan radula sebagai karakter yang paling penting untuk mengungkap sistem natural Gastropoda dan digunakan pada tingkatan spesies (Sarasin & Sarasin, 1898; Troschel, 1857 dalam Rintelen et. al, 2007;). Namun, Sarasin& Sarasin (1898) juga menyatakan bahwa radula pada tingkatan spesies terlalu bervariasi. Michel (2000) awalnya menggunakan morfologi radula untuk membedakan spesies Lavigeria di Danau Tanganyika, Afrika. Namun setelah dilakukan penelitian lebih detil terhadap jaringan tubuhnya, diduga perbedaan radula terjadi akibat jenis tersebut mengonsumsi makanan yang berbeda.
Pengaruh substrat terhadap bentuk radula juga didukunh oleh Rintelen et. al. (2004) yang menyatakan bahwa walaupun kebanyakan Pachychilidae Sulawesi memiliki radula yang identik, tetapi perbedaan substrat yang ditempatinya diduga menyebabkan beberapa perbedaan karakter radulanya. Rintelen et. al. (2007) melakukan penelitian terhadap Tylomelania di danau-danau Sulawesi dan menemukan bentuk radula yang mirip antara dua jenis, dari danau yang berbeda tetapi pada substrat yang sama. Contohnya pada jenis yang menempati substrat batu yaitu T. insulaesacrae (Danau Towuti) dan T. zeamais (Danau Matano) dimana keduanya tidak memiliki kekerabatan yang dekat. Contoh lainnya pada jenis yang menempati substrat lumpur yaitu T. gemmifera (Danau Matano), T. wolterecki (Danau Mahalona) dan T. kristinae (Danau Towuti). Kemungkinan F. costata (dari Tondano) dan Filopaludina sp.(dari Sentani) menyukai jenis substrat yang sama, sehingga mempunyai karakter radula yang mirip.
II-211
KESIMPULANHasil penelitian menyimpulkan bahwa keong:
Keong Filopaludina di Indonesia tercatat ada 15 jenis dan masing-masing memiliki karakter morfologi yang berbeda.
F. javanica memiliki karakter morfologi yang berbeda dengan F. sumatrensis terutama pada adanya garis lingkar, perbedaan jumlah seluk, ukuran rata-rata cangkang, jumlah embryonic shell , ukuran radula dan jumlah baris gigi. Kehadiran F. javanica di Papua merupakan catatan baru karena belum
pernah dilaporkan sebelumnya, sekaligus menegaskan bahwa jenis ini merupakan jenis yang mampu menyebar luas.
Jenis – jenis yang dijumpai di Sulawesi dan Papua (selain F. javanica ) diduga merupakan jenis endemik.
DAFTAR PUSTAKA
Bandel, K. 1984. The Radulae of Carribean and Other Mesogastropoda And Neogastropoda. Zoologische Verhandelingen (214) : 1-188
Berry, A. J. 1974. The Anatomy of West Malaysian Snails of Parasitological Significance. Malayan Nature Journal (27) : 131 - 165
Bieler, R. 1988. Phylogenetic Relationships in The Gastropod Family
Architectonicidae, With Notes on The Family Mathildidae (Allogastropoda). Malacological Review, Suppl. 4: 205-240
Brandt, R.A.M. 1974. The Non-Marine Aquatic Mollusca of Thailand. Arch. Moll (105): 1-423
Hadžišce, S., C.M. Patterson, J.B. Burch and P.T. LoVerde. 1976. The Embryonic Shell Surface Sculpture of Gocea and Valvata. Malacological Review (9): 1-14.
II-212
Hershler, R. and W.F. Ponder. 1998. A Review of Morphological Characters of Hydrobioid Snails. Washington: Smithsonian Institution Press
Heryanto, A. Munandar, Ristiyanti M.M. dan Susilowati P. 2003. Keong dari Taman Nasional Gunung Halimun, Sebuah Buku Panduan Lapangan. Cibinong: Biodiversity Conservation Project-LIPI-JICA-PHKA
Jones, R.E., I. Beveridge, L.R.G. Cannon, M. S. Harvey, E. S. Nielsen, W. F. Ponder and J. Just (Editors). 1998a. Mollusca The Southern Synthesis Part A. Collingwood: CSIRO Publishing
Köhler F. and M. Glaubrecht. 2003. Morphology, reproductive biology and molecular genetics of ovoviviparous freshwater gastropods (Cerithioidea, pachycilidae) from the Philippines, wih description of a new genus Jagora. Zoologica Scripta (32): 35-59
Michel, E. 2000. Phylogeny of a Gastropod Species Flock: Exploring Speciation in Lake Tanganyika in a Molecular Framework. Advances in Ecological Research (31): 275-302
Mimpfoundi, R. and A. Ndassa. 2005. Studies On The Morphology And Compatibility Between Schistosoma hæmatobium And The Bulinus sp. Complex (Gastropoda: Planorbidae) in Cameroon. African Journal of Biotechnology 4 (9) : 1010-1016
Rintelen, V.T., A.B. Wilson, A. Meyer and M. Glaubrecht. 2004. Escalation and Trophic Specialization Drive Adaptive Radiation of Freshwater Gastropods in Ancient Lakes on Sulawesi, Indonesia. The Royal Society (271): 2541-2549 Rintelen V.T., M. Glaubrecht and P. Bouchet. 2007. Ancient Lakes as Hotspots of
Diversity: A Morphological Review of An Endemic Species Flock of Tylomelania (Gastropoda: Cerithioidea: Pachychilidae) in The Malili Lake Systwm on Sulawesi, Indonesia. Hydrobiologia 529: 11-94
Sarasin, P. und F. Sarasin. 1898. Die Süsswasser-Mollusken von Celebes. Wiesbaden: C. W. Kreidel’s Verlag
II-213
http://www.pikiran-rakyat.com/cetak/2006/052006/28/geulis/lainnya.htmVan Benthem Jutting, W.S.S. 1956. Systematic Studies On The Non-Marine Mollusca Of The Indo-Australian Archipelago. Critical Revision of The Javanese Freshwater Gastropods. Treubia 23 (2) : 259 – 477
Van Benthem Jutting, W.S.S. 1959. Catalogue of The Non-Marine Mollusca of Sumatra and of Its Satellite Island. Beaufortia 7 (83) : 41 – 191
Van Benthem Jutting, W.S.S. 1963. Non Marine Mollusca of West New Guinea, Part. 1. Mollusca from Fresh and Brackish Waters. Nova Guinea (20): 409-521