6 BAB II
TINJAUAN PUSTAKA
2.1 Gulma Tanaman Pangan sebagai Pakan Ternak
Pemanfaatan limbah inkonvensional termasuk limbah dan gulma tanaman pangan sebagai bahan pakan ternak non ruminansia telah banyak dicobakan dan cukup prospektif untuk diaplikasikan dalam upaya peningkatan sistem ketahanan pakan dalam mendukung terwujudnya sistem ketahanan pangan yang kuat (Permana, 2015). Pemanfaatan limbah sebagai bahan pakan akan dapat mengurangi biaya produksi khususnya biaya pakan yang selama ini menjadi biaya terbesar dalam usaha peternakan karena biaya pakan mencapai ±70% dari total biaya produksi. Menurut Supriyati et al. (2003), pakan merupakan kebutuhan primer dalam usaha peternakan yang mencapai 70% dari total biaya produksi. Selain itu, pakan merupakan faktor utama di dalam keberhasilan usaha pengembangan peternakan disamping faktor bibit atau genetik dan tata laksana atau manajemen (Harfiah, 2010). Penggunaan bahan pakan dari limbah dan gulma tanaman pangan juga dapat mengurangi atau mencegah pencemaran lingkungan (Mudita et al., 2009).
Gulma merupakan tumbuhan yang tumbuh di suatu tempat dalam waktu tertentu tidak dikehendaki oleh manusia (Soerjani et al. 1996). Lebih lanjut diungkapkan, gulma tidak dikehendaki karena bersaing dengan tanaman yang dibudidayakan dan dibutuhkan biaya pengendalian yang cukup besar yaitu sekitar 25-30% dari biaya produksi. Eceng gondok yang merupakan salah satu jenis gulma juga memiliki kandungan nutrien yang cukup baik, yaitu energi
7 metabolis 2096,92 kkal/kg, protein kasar 13 %, serat kasar 21,3 % (Radjiman et al., 1999). Hasil analisis kandungan nutrien daun apu yang bersumber dari sawah, menunjukkan bahwa protein kasar daun apu sebesar 14,00%; serat kasar 19,71%; lemak kasar 1,54%; abu 19,70% dan kandungan energi termetabolisnya 1444,47 kkal/kg bahan (Sumaryono, 2003). Menurut Ahmed et al. (2012), kandungan kimia serat kasar eceng gondok yaitu 60% selulosa, 8% hemiselulosa dan 17% lignin. Meryandani et al. (2009) menyatakan bahwa selulosa terbungkus dan terikat secara ikatan kovalen maupun non-kovalen pada lignin dan hemiselulosa.
Limbah inkonvensional seperti sekam padi, bulu ayam, kulit kopi, lemak hewan maupun limbah dan gulma tanaman pangan seperti batang pisang, daun apu, enceng gondok serta limbah dan gulma lainnya mempunyai kandungan nutrien yang cukup baik bagi ternak (Permana, 2015). Lebih lanjut diungkapkan, namun disisi lain bahan pakan asal limbah termasuk gulma mempunyai berbagai keterbatasan seperti kualitas nutrien yang tidak seimbang serta ketersediaan nutrient available, mineral-vitamin dan daya cerna yang rendah. Mudita et al. (2009) menyatakan bahwa pemanfaatan ransum berbasis limbah tanpa aplikasi teknologi pengolahan akan menurunkan produktivitas sapi bali maupun kambing dibandingkan dengan pemanfaatan ransum dengan aplikasi teknologi fermentasi maupun suplementasi. Dewi et al. (2014) juga menyatakan bahwa penggunaan ransum berbasis limbah dan gulma tanaman pangan tanpa tersuplementasi biosuplemen isi rumen dapat menurunkan produktivitas itik bali dibandingkan dengan penggunaan ransum berbasis limbah dan gulma tanaman pangan tersuplementasi biosuplemen isi rumen. Bagi ternak unggas seperti itik bali, serat kasar dapat dikatakan tidak dapat dicerna, dimana hanya 20%-30% serat kasar
8 yang dapat dicerna dalam saluran pencernaan belakang yaitu di sekum dan kolon yang kemungkinan dapat diserap/dimanfaatkan oleh ternak sangat kecil (Bidura, 2007). Aplikasi teknologi pengolahan pakan dan teknik suplementasi berpotensi memperbaiki kelemahan yang ada (Dewi et al., 2014).
Penelitian pemanfaatan limbah dan gulma tanaman pangan seperti batang pisang, eceng gondok, maupun gulma lainnya telah banyak dilakukan (Permana, 2015). Hasil penelitian Marlina dan Askar (2001) menunjukkan bahwa hasil analisis kandungan zat-zat makanan ransum yang diberi tambahan daun eceng gondok menunjukkan adanya peningkatan serat kasar dari 2,64 % menjadi 4,06 % dan protein yang relatif kecil. Lebih lanjut diungkapkan, hasil percobaannya menunjukkan bahwa penambahan eceng gondok sampai 10% tidak merugikan baik terhadap produksi telur maupun kualitas telurnya yang berarti peningkatan serat kasar pada penambahan eceng gondok sampai 10% tidak berpengaruh.
Marlina dan Askar (2001) mengungkapkan bahwa tingginya kandungan serat kasar dalam ransum ternak babi dapat mempengaruhi pertumbuhannya, namun berdasarkan hasil percobaan menunjukkan bahwa pencampuran eceng gondok sampai 15% ke dalam ransum babi tidak berpengaruh nyata terhadap pertumbuhan babi, tentunya akan sangat menekan harga karena akan mengurangi jumlah pemakaian konsentrat. Sehingga penambahkan penggunaan eceng gondok sebagai bahan ransum harus diatur sedemikian rupa. Namun hasil penelitian Riana dan Bidura (2002) menunjukkan penggunaan eceng gondok dalam ransum ayam buras tidak boleh lebih dari 10%, penggunaan eceng gondok 20% dan 30% mengakibatkan pertambahan bobot badan ayam buras menurun. Lebih lanjut diungkapkan, kandungan serat kasar yang tinggi merupakan faktor pembatas
9 pemanfaatan eceng gondok yang merupakan sumber daya lokal asal limbah/gulma tanaman pangan sebagai komponen ransum. Namun Bidura (2006) mengungkapkan efek negatif bahan pakan asal limbah dapat diatasi melalui aplikasi bioteknologi pakan baik melalui biofermentasi, suplementasi maupun penambahan probiotik.
2.2 Selulosa sebagai Faktor Pembatas Pemanfaatan Gulma Tanaman Pangan Faktor pembatas dalam pemanfaatan gulma tanaman pangan sebagai pakan ternak adalah rendahnya kandungan nutrient available dan kecernaan nutrien yang terutama disebabkan kandungan serat yang cukup tinggi (Permana, 2015). Serat merupakan bagian dari makanan yang sulit untuk dicerna secara enzimatis (enzim yang dikeluarkan oleh unggas) dan tidak digolongkan sebagai zat makanan (Linder, 1985), sehingga kandungan serat kasar yang cukup tinggi dalam ransum dapat menurunkan produktivitas ternak. Selulosa merupakan salah satu bahan organik yang terdapat dalam jumlah banyak di alam dan merupakan sumber energi yang sangat potensial bagi ruminansia (Arora, 1989). Puspitasari (2009) menyatakan bahwa selulosa merupakan senyawa penyusun tanaman dalam jumlah banyak yang sebagai struktur dinding sel tanaman.
Selulosa merupakan komponen utama dinding sel yang tersusun atas polimer glukosa yang berbentuk rantai linier seragam yang dihubungkan oleh ikatan β-1,4 glikosidik (Howard et al., 2003; Hermiawati et al., 2010). Struktur yang linier menyebabkan selulosa bersifat kristalin dan tidak mudah larut sehingga tidak mudah didegradasi secara kimia maupun mekanis (Dewanti et al., 2013). Namun degradasi secara sempurna polimer tersebut mampu menyediakan
10 semua potensi nutrisi yang terkandung dalam bahan pakan asal limbah inkonvensional (Mudita et al., 2014).
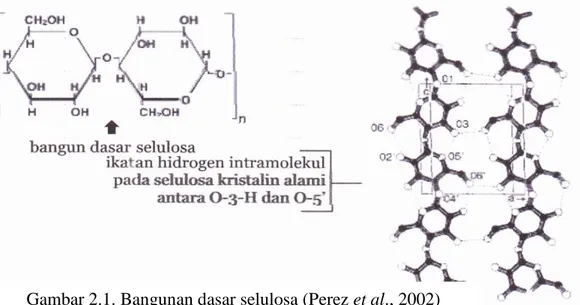
Selulosa mempunyai bobot molekul tinggi yang terdapat dalam jaringan tanaman pada dinding sel tanaman dalam bentuk mikrofibril (Suparjo et al., 2008). Selulosa adalah polimer dari β-D-glukosa dan gugus atas dan bawahnya dihubungkan dengan CH2OH (Zamora, 2005). Selulosa murni adalah homoglycan dari bobot molekul yang tinggi dengan unit berikutnya berbentuk selobiosa. Pada tanaman, ikatan selulosa dibentuk dengan cara yang tersusun untuk memproduksi kumpulan padat (mikrofibril) yang disatukan bersamaan baik oleh ikatan molekul hidrogen inter maupun intra (McDonald et al., 2002). Menurut Harjo et al. (1989), secara alamiah molekul selulosa tersusun dalam bentuk fibril yang terdiri dari beberapa molekul selulosa paralel yang dihubungkan oleh ikatan hydrogen. Lebih lanjut diungkapkan bahwa fibril-fibril tersebut akan membentu kristal dan struktur tersebut dibungkus oleh lignin yang berperan melindungi selulosa terhadap serangan enzim pemecah selulosa. Rumus molekul selulosa adalah (C6H10O5)n (Perez et al., 2002).
11 Menurut Irawadi (1990), struktur selulosa sebagai polimer karbohidrat atau polisakarida tersusun dari anhidroglukopiranosa yang memiliki rumus C6H10O5. Lebih lanjut diungkapkan bahwa selulosa diikat oleh β-1,4-glukosida alam, membentuk rantai polimer linier glukan dengan struktur rantai yang seragam. Ikatan β-1,4 glikosida pada selulosa merupakan ikatan penghubung antar unit D-glukosa (Lehninger, 1982 dalam Puspitasari, 2009). Ikatan penghubung ini dapat dipecah menjadi monomer-monomer glukosa melalui jalur hidrolisa asam dan hidrolisa enzimatis atau biologis (Harjo et al., 1989). Lebih lanjut Harjo et al. (1989) menambahkan bahwa meskipun ikatan glikosidik beta 1,4 pada serat selulosa dapat dipecah menjadi monomer-monomer glukosa dengan cara hidrolisa asam atau hidrolisa enzimatis, akan tetapi pemecahannya tidak secepat pati atau gula.
2.3 Enzim Selulase
Selulase adalah nama bagi semua enzim yang memutuskan ikatan glikosidik beta-1,4 di dalam selulosa, selobiosa, dan turunan selulosa lainnya (Denman et al., 1996). Lebih lanjut diungkapkan bahwa di alam, enzim selulase ditemukan di berbagai ekosistem, terutama ditemukan pada proses dekomposisi serasah daun, hingga keadaan anaerobik pada ruminansia. Selulase merupakan enzim yang mampu menghidrolisis selulosa menjadi gula sederhana atau glukosa dengan memutuskan ikatan glikosidik β-1,4 dalam selulosa, selodekstrin, selobiosa dan turunan selulosa lainnya (Kulp, 1975 dalam Syam, 2008).
Lynd et al. (2002) dan Beauchemin et al. (2003) menyatakan bahwa perombakan selulosa secara enzimatis berlangsung karena adanya kompleks enzim selulase yang bersifat spesifik untuk menghidrolisis ikatan β-1,4-glikosidik,
12 rantai selulosa dan derivatnya melalui beberapa tahapan yang terdiri dari enzim endo-β-glukanase/CMC-ase, eksoglukanase, dan β-glukosidase. Enzim selulase merupakan sistem enzim yang terdiri dari enzim endoglukanase, eksoglukanase dan β-glukosidase (Syam, 2008). Menurut Schlegel (1994) mekanisme pemecahan selulosa oleh selulase sekurang-kurangnya terdiri dari tiga enzim: (1) Enzim-enzim endo-β-1,4 glukanase mempengaruhi secara serentak ikatan β-1,4 di dalam makromolekul dan menghasilkan potongan-potongan besar berbentuk rantai dengan ujung-ujung bebas, (2) Enzim ekso-β-1,4 glukanase memotong mulai dari ujung-ujung rantai, disakarida selobiosa, (3) Enzim-enzim β-glukosidase menghidrolisasi selobiosa dengan membentuk glukosa. Enzim selulase adalah enzim yang dapat mengkatalisis dan menghidrolisis ikatan β 1-4 glukosidik pada selulosa (ikatan yang paling banyak di selulosa) (Bhat, 2000).
Aktivitas enzim endoglucanases atau yang sering disebut dengan CM-cellulases (carboxymethylcellulose), menghidrolisis secara acak ikatan pada serat selulosa (Syam, 2008). Lebih lanjut diungkapkan bahwa endoglukanase memotong secara acak pada situs internal amorf pada rantai selulosa polisakarida menghasilkan oligosakarida dengan panjang yang berbeda-beda dan akhirnya membentuk ujung rantai baru (Syam, 2008). Hal ini mengakibatkan rantai polisakarida yang telah terpotong (oligosakarida) mempunyai panjang rantai yang berbeda-beda (Bhat, 2000). Hasil dari hidrolisis serat selulosa adalah glukosa, cellobiose, cellotriose, dan oligosakarida yang lebih tinggi (Wood, 1985). Cellobiohydrolyase yang sering disebut dengan eksoglukanase adalah enzim pendegadasi selulosa yang ditemukan pada mayoritas fungi yang dapat mendegadasi selulosa (Wood, 1985). Lebih lanjut diungkapkan bahwa
13 eksoglukanase berperan dalam mengatur proses reduksi atau tanpa reduksi dari ujung rantai selulosa polisakarida, membebaskan melepaskan unit-unit glukosa secara berturut-turut. Eksoglukanase dapat menghidrolisis mikrokristalline namun tidak dapat menghidrolisis carboxymethylcellulose (Kim dan Kim, 1995). Enzim eksoglukanase terdiri dari 1,4- D-glucan glucanohydrolases (lebih dikenal dengan cellodextrinases) dan 1,4-D-glucan cellobiohydrolases (cellobiohydrolases). Enzim eksoglukanase adalah enzim yang aktif pada sisi crystalline selulosa (Mattinen, 1998).
β-glucoside glucohydrolases lebih dikenal sebagai β-glukosidase. β-glukosidase adalah enzim yang digunakan untuk menghidrolisis cellobiose dan pada beberapa kasus dapat menghidrolisis cello-oligosakarida menjadi glukosa (Wood, 1985). Lebih lanjut diungkapkan enzim endogluconases dan β- glukosidase dapat menghidrolisis selulosa menjadi glukosa. β-glukosidase dibutuhkan untuk menghidrolisis inhibitor cellobiose. Laju hidrolisis enzim selulase ditentukan oleh struktur enzim dan struktur substrat, dimana struktur kristal dari selulosa relatif lebih sulit dihidrolisis dibandingkan dengan struktur amorf (Coughlan, 1985).
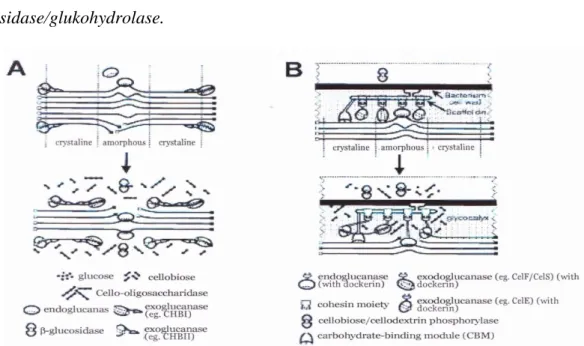
Perombakan selulosa oleh kelompok bakteri selulolitik berlangsung melalui beberapa tahapan (Bhat dan Bhat, 1997). Tahap pertama adalah menguraikan polimer selulosa secara random/acak oleh enzim carboxymethilcelulase/CMC-ase atau endo β-1,4 glukanase dengan cara memecah ikatan hidrogen yang ada di dalam struktur kristalin/amorf selulosa (ikatan internal α-1,4-glukosida) sehingga terbentuk rantai-rantai individu selulosa (oligodekstrin). Tahap kedua adalah penguraian selulosa dari ujung pereduksi dan
14 non pereduksi oleh eksoglukanase (selodektrinase dan selobiohydrolase) melalui pemotongan ujung-ujung rantai individu selulosa (ujung pereduksi dan non-pereduksi) sehingga menghasilkan disakarida dan tetrasakarida (misal selobiosa). Lynd et al. (2002) mengungkapkan hidrolisis bagian berkristal selulosa hanya dapat dilakukan secara efisien oleh enzim eksoglukanase. Tahap ketiga (terakhir) adalah tahap penguraian selobiosa menjadi glukosa oleh enzim β-glukosidase/glukohydrolase.
Aktivitas enzim didefinisikan sebagai kecepatan pengurangan substrat atau kecepatan pembentukan produk pada kondisi optimum (Lynd et al., 2002). Satu unit aktivitas enzim selulase didefinisikan sebagai jumlah enzim yang menghasilkan satu mikromol gula reduksi (glukosa) setiap menit (Lehninger, 1993 dalam Syam, 2008). Lebih lanjut diungkapkan, setiap enzim berfungsi secara optimal pada pH, suhu, dan konsentrasi substrat tertentu.
15 Faktor-faktor yang mempengaruhi aktivitas enzim yaitu:
a. Konsentrasi Substrat
Substrat yang digunakan dalam proses fermentasi berpengaruh terhadap aktivitas dan produktivitas enzim. Hasil eksperimen menunjukkan pada konsentrasi enzim yang tetap, maka pertambahan konsentrasi substrat akan menaikkan kecepatan reaksi. Akan tetapi pada batas konsentrasi tertentu, kecepatan reaksi tidak akan mengalami kenaikan walaupun konsentrasi substrat diperbesar. Substrat yang dipergunakan secara umum untuk penetapan aktivitas endoglukanase adalah CMC (Carboxymethyl cellulose) (Syam, 2008).
b. Suhu
Suhu sangat berpengaruh dan digunakan sebagai katalis enzim dalam suatu reaksi kimia. Jika pada suhu rendah reaksi kimia berlansung lambat, sedangkan pada suhu yang lebih tinggi reaksi berlangsung lebih cepat (Poedjiadi 1994 disitasi Syam, 2008). Namun, kenaikan suhu dapat menyebabkan terjadinya proses denaturasi, maka bagian aktif enzim akan terganggu dan kecepatan reaksinya pun akan menurun.
c. Part of Hidrogen (pH)
Aktivitas enzim selulase pada umumnya stabil pada kisaran pH 4,5 dan 6,5. Perubahan pH lingkungan sangat berpengaruh terhadap efektvitas bagian aktif enzim dalam membentuk kompleks enzim substrat. Jika kondisi pH rendah atau pH tinggi dapat pula menyebabkan terjadinya proses denaturasi dan akan mengakibatkan aktivitas enzim menjadi turun (Syam, 2008).
Maranatha (2008) menyatakan bahwa setiap bakteri selulolitik menghasilkan kompleks enzim selulase yang berbeda-beda, tergantung dari gen
16 yang dimiliki dan sumber karbon yang digunakan. Sumardi et al. (2010) menyatakan bahwa faktor genetik mempengaruhi besarnya produksi enzim. Lebih lanjut diungkapkan, gen setiap mikroorganisme berbeda-beda sehingga masing-masing mikroorganisme memiliki sifat yang berbeda dan dari tiap gen memiliki sifat yang spesifik untuk mengkode enzim-enzim tertentu.
Substrat Carboxy Methyl Cellulose (CMC) merupakan substrat selulosa murni yang berbentuk amorphous sehingga aktivitas enzim selulase pada substrat CMC merupakan aktivitas enzim endo-1,4-β-glukanase (Meryandini et al., 2009). Lebih lanjut diungkapkan, substrat avicel merupakan substrat selulosa yang berbentuk kristalin sehingga aktivitas enzim selulase pada substrat avicel merupakan aktivitas enzim ekso-1,4-β-glukanase. Endo-1,4-β-glukanase bekerja pada rantai dalam CMC menghasilkan oligosakarida atau rantai selulosa yang lebih pendek (Maranatha, 2008). Lebih lanjut diungkapkan, enzim ekso-1,4-β-glukanase bekerja memotong ujung rantai oligosakarida menjadi selobiosa, yaitu dua molekul glukosa yang berikatan secara β-1,4-glikosidik. Hasil penelitian Meryandani et al. (2009) menunjukkan bahwa isolat asal tanah pertanian dengan C5-1, C11-1 dan C4-4 memiliki aktivitas enzim selulase optimum pada substrat CMC berturut-turut sebesar 0,042 nkat/ml; 0,193 nkat/ml dan 0,112 nkat/ml yang menggambarkan bahwa isolat tersebut memiliki sejumlah besar endo-1,4-β-glukanase.
Hemiselulosa dan lignin akan mengganggu aktivitas enzim selulase yang hanya spesifik memotong ikatan β-1,4-glikosidik pada selulosa (Maranatha, 2008). Lebih lanjut diungkapkan, untuk meningkatkan luas permukaan substrat maka jerami padi, tongkol jagung dan kulit pisang diperkecil ukurannya sampai
17 65 mesh. Menurut Meryandani et al. (2009), hal yang menghambat aktivitas selulase pada substrat jerami padi dan tongkol jagung adalah komponen lignin karena lignin membungkus dan mengikat selulosa secara fisik sehingga menghalangi enzim selulase bekerja maksimal pada substrat.
Meryandani et al. (2009) menyatakan bahwa aktivitas enzim selulase yang lebih tinggi pada substrat limbah pertanian dibandingkan dengan selulosa sintetis kemungkinan disebabkan oleh adanya enzim hemiselulolitik pada ekstrak kasar yang diproduksi oleh bakteri. Han et al. (2003) menyatakan bahwa bakteri Clostridium cellulovorans mensintesis enzim hemiselulolitik saat tumbuh pada substrat selulosa seperti selobiosa. Lebih lanjut diungkapkan, ekspresi enzim selulase berhubungan dengan ekspresi enzim hemiselulase. Susanti (2011) yang menyatakan bahwa apabila waktu reaksi enzimatis telah mencapai optimum dalam menghasilkan produk yang maksimum maka aktivitas enzim mengalami penurunan dengan penambahan waktu inkubasi lebih lanjut. Susanti (2011) juga menyatakan bahwa produk gula pereduksi yang dihasilkan dari reaksi enzimatis sebanding dengan lama waktu inkubasi, tetapi jika sisi aktif enzim telah jenuh oleh substrat.
2.4 Mikroba Penghasil Enzim Selulase
Degradasi selulosa merupakan proses pemecahan polimer anhidroglukosa menjadi molekul yang lebih sederhana. Maranatha (2008) menyebutkan mikroba selulolitik dari kelompok bakteri mempunyai tingkat pertumbuhan cepat dan aktivitas selulolitik tinggi. Mikroba selulolitik khususnya bakteri banyak ditemukan pada tanah pertanian, hutan, jaringan hewan, saluran pencernaan herbivora baik rumen, sekum maupun kolon, rayap (air liur, sel tubuh, saluran
18 pencernaan maupun sarangnya), saluran pencernaan hewan invertebrate serta pada tumbuhan yang membusuk/mati (Anam et al., 2012). Bakteri di alam yang bersifat selulolitik antara lain: Clostridium (C. acetobutylicum, C. thermocellum), Bacillus sp., Acidothermus, Pseudomonas (P. cellulosa), Rhodothermus (Anindyawati, 2010), Erwinia, Acetovibrio, Mikrobispora, Cellulomonas, Cellovibrio, Streptomyces, Sclerotium rolfisii (Duff and Murray, 1996; Indrawati Gandjar, 2006), Fibrobacter succinogenes, Ruminococcus albus, Ruminococcus flavefaciens, Butytrivibrio fibrisolvens (Lynd et al., 2002).
Bakteri selulolitik merupakan bakteri yang mampu menghasilkan enzim selulase yang menghidrolisis selulosa menjadi produk yang lebih sederhana yaitu glukosa (Meriyandini et al., 2009). Kemampuan degradasi selulosa berbagai bakteri bervariasi yang dipengaruhi oleh jenis/spesies, substrat maupun lingkungan. Chen dan Weimer (2001) mengungkapkan bahwa bakteri selulolitik akan dominan dalam rumen ruminansia apabila ternak diberikan pakan hijauan atau pakan kaya serat.
2.5 Cacing Tanah sebagai Sumber Mikroba Selulolitik
Cacing tanah adalah salah satu binatang yang mampu mendegradasi berbagai bahan organik (Suhartanti et al., 2013). Kemampuan ini dikarenakan terkandungnya berbagai mikroba sinergis seperti protozoa, bakteri dan mikro fungi di dalam saluran pencernaannya serta terkandung berbagai enzim seperti lipase, protease, chitinase, selulase, urease dan amilase (Andika, 2015). Selain itu, di dalam saluran pencernaan cacing tanah juga terdapat mukus yang mengandung berbagai nutrien (karbohidrat, protein, bahan mineral dan bahan organik, serta berbagai asam amino) dan hormon (Pathma dan Sakthivel, 2012).
19 Lebih lanjut diungkapkan, cacing tanah juga mampu mendegradasi senyawa antinutrisi dan lignoselulosa, memproduksi antibiotika, chitinase dan glucanase, pigmen fluorescent siderophores, serta berbagai growth promotor melalui pelarutan mineral dan menekan mikroba patogen, serta memproduksi hormon 1-aminocyclopropane-1carboxylate (ACC) deaminase, dan menekan mikroba patogen.
Fuji et al. (2012) menambahkan bahwa bakteri selulolitik yang dominan pada cacing tanah Amynthun heteropoda dan Eisenia fetida adalah Burkholderia spp, Enterobacter Herbaspiririllum dan Pseidomonas, sedangkan fungi selulolitik yang dominan adalah Penicillium, Fusarium dan Staphylotrichum. Owa et al. (2013) menunjukkan dari saluran pencernaan cacing tanah Libyodrilus violaceus berhasil diisolasi bakteri Acinobacter sp, Kiebsiella sp, Bacillus sp., B. brevis, B. cereus, B. lalerosporus, B. lichenoform, P. vulgaris, Pseudomonas s, Alcaligans faecalis, Corynebacterium sp., E. cloacae, Erwinia salicie, Flavobacterium sp., F. aquartile, Micrococcus inteus, M. kristinae, M. Varians dan Proteus rennvi. Hasil penelitian yang dilakukan oleh Asnani dan Puji Lestari (2009) menunjukkan bahwa ekstrak kasar dan fraksi ammonium sulfat cacing P. excavatus mempunyai aktivitas enzim amilase, lipase dan protease. Hasil penelitian yang dilakukan oleh Susanti (2007) juga menyimpulkan bahwa dalam saluran pencernaan cacing tanah (Lumbricus terestris) terdapat bakteri selulolitik.
Hasil penelitian Suhartanti et al. (2013), bakteri selulolitik yang diisolasi dari saluran pencernaan cacing tanah terdeteksi berbentuk batang dan bersifat gram negatif sehingga dapat disimpulkan bahwa dalam saluran pencernaan cacing tanah (Lumbricus terestris) dan kascing terdapat bakteri selulotik. Bakteri
20 selulolitik dalam saluran pencernaan cacing tanah dapat mendegradasi selulosa, hasil degradasi dari bakteri tersebut sangat bermanfaat untuk meningkatkan senyawa-senyawa yang diperlukan untuk pertumbuhan tanaman atau mikroorganisme lain (Reanida et al., 2012). Bakteri selulolitik dalam saluran pencernaan mempunyai peranan penting dalam proses pendegradasi seloluosa yang hidup secara anaerob dalam saluran cerna (Suhartanti et al., 2013). Bakteri tersebut salah satunya dapat ditemukan dalam saluran cerna cacaing tanah karena hewan tersebut mamakan sisa organisme yang membusuk dalam tanah (Reanida et al., 2012). Cacing tanah dapat mencerna bahan organik melebihi dari berat badannya dalam waktu sehari (Sugiyarto et al., 2007).