KERAGAMAN STRUKTUR MORFOLOGIS DAN GEN
CYTOCHROME b
DNA MITOKONDRIA
Kryptopterus
spp. DAN
Ompok
spp. (SILURIDAE) DI DAS BATANG HARI JAMBI
ABDUL RAHMAN
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2010
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis Keragaman Struktur Morfologis dan Gen Cytochrome b DNA Mitokondria Kryptopterus spp. dan Ompok spp. (Siluridae) di DAS Batang Hari Jambi adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada Perguruan Tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Desember 2010 Abdul Rahman NIM G352080031
ABSTRACT
ABDUL RAHMAN. Variation of Morphological Structure and Cytochrome b DNA Mitochondria of Kryptopterus spp. and Ompok spp. (Siluridae) of Batang Hari Watersheds, Jambi. Under direction of Dedy Duryadi Solihin and Ridwan Affandi.
Kryptopterus spp. and Ompok spp. are locally known as glass catfish. The high variety of Glass catfish can be found in Batang Hari river. Geological history shows that Batang Hari, Musi, Indragiri and Kapuas river were originally part of an ancient river called Sunda Utara. Unfortunately, phyllogeographic analytical research of fauna on these rivers is still very limited. The objectives of this study were to find out the variation of morphological structure and cytochrome b DNA mitochondria of glass catfish on Batang Hari river. Samples were collected from four different locations from upriver to downstream. The morphological structure was measured for 12 characters and based on these characters, 12 proportions of body size were calculated. Amplification of cyt b gen mtDNA was done by PCR with specific primer: CBKR 1 and CBKR 2. The study revealed that four species of glass catfish still inhibit Batang Hari river, namely Ompok hypopthalmus, Kryptopterus limpok, K. micronema, and K. bichirris. Those glass catfish have a small variance, and can only be grouped basically on the species within the same location with PCA method. Sequencing result 927 bp partial of cyt b genes, from the primary target 1104 bp long. Based on nucleotides sequence, there is one site of distinctive nukleotide between Kryptopterus and Ompok. An amino acid was identified as distinguishing factor between Kryptopterus and Ompok from Sumatera. There are 11 of variable nucleotides on K. limpok sequences from Batang Hari river, but only one nukleotide variable found on O. hypopthalmus. Key words: variation, morphological, cytochrome b, Kryptopterus, Ompok,
RINGKASAN
ABDUL RAHMAN. Keragaman Struktur Morfologis dan Gen Cytochrome b
DNA Mitokondria Kryptopterus spp. dan Ompok spp. (Siluridae) di DAS Batang Hari Jambi. Dibimbing oleh Dedy Duryadi Solihin dan Ridwan Affandi.
Kryptopterus spp. dan Ompok spp. memiliki struktur morfologis yang mirip sehingga dikenal dengan nama lokal yang sama, “ikan lais”. Salah satu perairan yang memiliki keragaman ikan lais yang tinggi adalah daerah aliran sungai (DAS) Batang Hari. Sejarah geologis menunjukkan bahwa DAS Batang Hari, Musi, Indragiri dan Kapuas berasal dari sungai yang sama yaitu DAS Sunda Utara Purba. Akan tetapi, belum ada penelitian tentang filogeografi fauna antarkelompok sungai yang berasal dari DAS Sunda Utara Purba.
Aliran Sungai Batang Hari sangat panjang dan melewati berbagai karakter topografi dan vegetasi, serta pola pemanfaatan. Hal ini diduga akan memunculkan variasi struktur morfologis dan genetik pada populasi fauna, termasuk ikan laisdi DAS Batang Hari. Selain itu, kesamaan sejarah geologis juga diduga akan berkaitan dengan keragaman dan karakter spesifik fauna antarsungai dalam kelompok DAS Sunda Utara Purba. Penelitian ini bertujuan untuk menganalisis keragaman struktur morfologis dan gen cytochrome b (cyt b) DNA mitokondria (mtDNA) ikan lais di DAS Batang Hari.
Ikan lais dikoleksi dari empat lokasi pengambilan sampel di DAS Batang Hari yaitu Mandiangin, Sungai Bengkal, Pelayangan dan Simpang. Identifikasi spesies didasarkan pada ciri meristik. Data struktur morfologis diukur untuk 12 karakter morfometrik. Berdasarkan 12 karakter tersebut, dilakukan penghitungan 12 indeks morfometrik. Amplifikasi gen cyt b mtDNA dilakukan dengan metode
polymerase chain reaction (PCR) menggunakan primer CBKR 1:
5‟-CCCGAAAAACTCACCCCTTA-3‟ dan cbkr 2:
5‟-ATAGCCCGGTTAGAGGGTTT-3‟. Target primer ini adalah gen cyt b
sepanjang 1104 bp. Penempelan primer dilakukan pada suhu 60oC selama 45 detik.
Penelitian ini menemukan empat spesies ikan lais di DAS Batang Hari, yaitu: Ompok hypopthalmus, Kryptopterus limpok, K. micronema, dan K. bichirris.Ompok. hypopthalmus, K. limpok dan K. micronema dapat ditemukan di semua stasiun penelitian dengan jumlah sampel sesuai dengan target yang direncanakan (10 individu/lokasi). Akan tetapi jenis K. micronema hanya ditemukan di dua periode dari tiga periode pengambilan sampel. Demikian pula dengan Kryptopterus bichirris, jenis ini hanya ditemukan di satu dari tiga kali periode pengambilan sampel. Kryptopterus bichirris hanya ditemukan di stasiun Mandiangin dan Pelayangan. Berdasarkan kehadiran spesies setiap periode pengambilan sampel dan jumlah sampel yang didapatkan, perbandingan struktur morfometrik dan gen cyt b DNA mitokondria antar lokasi hanya dilakukan pada
O. hypopthalmus dan K. limpok. Kedua spesies ini merupakan ikan lais yang paling umum ditemukan di DAS Batang Hari.
Pengukuran nisbah morfometrik menunjukkan keragaman yang rendah pada intra dan antarpopulasi ikan lais di DAS Batang Hari. Nilai keragaman (varians) intra populasi pada 12 indeks struktur morfologis hanya berkisar 0.00–0.01 (χ = 0.13–1.20). Nilai varians antarpopulasi berkisar 0.01-0.51 (χ = 0.15–1.12)
sedangkan varians antarspesies pada seluruh sampel ikan lais dari DAS Batang Hari berkisar 0.01–0.26 (χ = 0.17–1.01). Analisis biplot PCA pada seluruh sampel tidak dapat mengelompokkan sampel, baik berdasarkan spesies maupun lokasi.
Berbeda dengan biplot pada seluruh sampel, biplot PCA ketika sampel dibagi menurut lokasi mampu mengelompokkan sampel berdasarkan spesies. Hasil analisis menunjukkan lebih dari 85% sampel pada setiap lokasi mengelompok berdasarkan spesies. Pengelompokan sampel berdasarkan lokasi juga mampu menunjukkan karakter spesifik sebagai penciri spesies dalam kelompok ikan lais. Ompok hypopthalmus dicirikan dengan indeks N9 (tinggi pangkal kepala/tinggi badan). Kryptopterus limpok dicirikan dengan indeks N8 (tinggi moncong/tinggi pangkal kepala). Kryptopterus bichirris dicirikan dengan indeks N11 (diameter mata/panjang kepala) dan N12 (panjang sirip dada/panjang kepala). Kryptopterus micronema walaupun terletak mengelompok, akan tetapi tidak ada indeks morfometrik penciri jenis ini.
Analisis biplot PCA antarlokasi menunjukkan hasil yang berbeda pada K. limpok dan O. hypopthalmus. Ompok hypopthalmus dapat dikelompokkan berdasarkan lokasi, sedangkan K. limpok hanya dapat dikelompokkan di lokasi S. Bengkal. Analisis biplot PCA pada O. hypopthalmus juga mampu menunjukkan indeks morfometrik penciri lokasi. Ompok hypopthalmus di Mandiangin dicirikan dengan indeks N4 (tinggi ekor/panjang baku) dan N10 (tinggi ekor/tinggi badan). Hal ini berarti O. hypopthalmus di Mandiangin memiliki struktur ekor yang lebih tinggi/tebal dibanding O. hypopthalmus dari lokasi lain. Ompok hypopthalmus di Pelayangan dicirikan dengan N6 (panjang rahang atas/panjang kepala), sedangkan
O. hypopthalmus di Simpang dicirikan dengan N8 (tinggi ‟moncong‟/tingi pangkal kepala). Tidak ada penciri O. hypopthalmus dari S. Bengkal.
Penjajaran (alignment) gen cyt b semua sampel dari Batang Hari dengan gen
cyt b semua jenis ikan lais di genbank dan Kampar menghasilkan runutan nukleotida sepanjang 927 bp. Nukleotida yang berbeda (variabel) pada genus
Kryptopterus dan Ompok tidak menunjukkan perbedaan yang signifikan (33.01% berbanding 28.91%). Berdasarkan posisi nukleotida pada triplet kodon, keragaman nukleotida terbesar secara berturut-turut adalah pada nukleotida ketiga, kesatu dan kedua dari triplet kodon. Komposisi nukleotida penyusun runutan secara umum menunjukkan sitosin merupakan basa nitrogen dengan komposisi terbesar, sedangkan basa nitrogen guanin memiliki komposisi terkecil. Terdapat satu nukleotida pembeda genus Kryptopterus dengan Ompok.
Nukleotida pembeda ini terletak pada situs ke-132 dari nukleotida gen cyt b utuh. Situs ke-132 pada Kryptopterus adalah timin, sedangkan pada Ompok adalah sitosin. Terdapat satu asam amino pembeda genus Kryptopterus dengan Ompok
dari Sumatera. Asam amino pembeda ini terletak pada situs ke-155 dari gen cyt b
utuh.
Hasil alignment intraspesies di DAS Batang Hari menunjukkan keragaman yang lebih tinggi pada K. limpok dibanding pada O. hypopthalmus. Terdapat 11 (1.19%) nukleotida yang variabel pada K. limpok, sedangkan pada O. hypopthalmus hanya ada 1 (0.11%). Ditemukan dua situs nukleotida barcodingK. limpok dan 10 situs nukleotida barcoding O. hypopthalmus dari DAS Batang Hari. Dua situs barcoding K. limpok Batang Hari terletak pada situs 30 dan ke-891 dari runutan nukleotida gen cyt b parsial.
11 nukleotida yang variabel pada K. limpok terdiri dari empat situs parsimoni dan tujuh situs singleton. Situs parsimoni merupakan situs yang memiliki minimal dua jenis (tipe) nukleotida dan setiap tipe minimal dimiliki oleh dua runutan. Empat situs parsimoni yang ditemukan konsisten membedakan K. limpok Mandiangin-Simpang dengan K. limpok S. Bengkal-Pelayangan. Empat situs parsimoni ini terletak pada situs ke-192, 591, 615 dan ke-762 dari runutan gen cyt b parsial. Substitusi pada keempat situs ini bersifat silent.
Terdapat 42 (4.53%) nukleotida yang variabel dari total runutan K. limpok
yang dialignment. 21 dari 42 nukleotida yang variabel ini merupakan situs diagnostik pembeda K. limpok Sumatera dengan genbank. Lima dari 21 nukleotida diagnostik ini (situs ke-499, 571, 616, 786 dan 884) bersifat substitusi
nonsilent sehingga menghasilkan lima situs asam amino diagnostik pembeda K. limpok Sumatera dengan genbank. Perbandingan nukleotida K. limpok Sumatera (Batang Hari dan Kampar) menunjukkan terdapat 20 (2.16%) nukleotida yang variabel. Empat dari 20 situs ini merupakan situs nukleotida diagnostik yang membedakan K. limpok Batang Hari dengan K. limpok Kampar. Empat nukleotida diagnostik pembeda K. limpok Batang Hari dengan Kampar ini terletak pada situs ke-30, 360, 388 dan ke-891 dari runutan gen cyt b parsial. Substitusi pada nukleotida ke-388 bersifat nonsilent sehingga dapat digunakan sebagai situs asam amino diagnostik pembeda K. limpok Batang Hari dengan K. limpok Kampar.
Satu nukleotida yang variabel pada O. hypopthalmus DAS Batang Hari. terdapat pada nukleotida ke-883 dari runutan gen cyt b parsial. Nukleotida pada O. hypopthalmus dari Simpang dan S. Bengkal adalah guanin, sedangkan pada O. hypopthalmus dari Mandiangin dan Pelayangan adalah adenin. Substitusi ini bersifat nonsilent. Asam amino ke-295 pada O. hypopthalmus dari Simpang dan S. Bengkal adalah alanina (GCC), sedangkan pada O. hypopthalmus dari Mandiangin dan Pelayangan adalah treonina (ACC). Alanina bersifat non polar (hidrofobi), sedangkan treonina bersifat polar. Perbandingan nukleotida O. hypopthalmus Sumatera (Batang Hari dan Kampar) menunjukkan terdapat 12 (1.29%) nukleotida yang variabel. 10 dari 12 situs ini merupakan situs nukleotida diagnostik yang membedakan O. hypopthalmus Batang Hari dengan O. hypopthalmus Kampar. Ke-10 situs nukleotida diagnostik ini bersifat substitusi
silent.
Konstruksi filogeni antara semua sampel ikan lais berdasarkan runutan gen
cyt b parsial 927 bp mampu memisahkan genus Kryptopterus dengan Ompok. Akantetapi O. hypopthalmus membentuk cluster sendiri yang terpisah dari cluster
Ompok secara umum. Adanya cluster tersendiri pada O. hypopthalmus merupakan hal yang membutuhkan penelitian lebih lanjut. Dibutuhkan penambahan jumlah sampel baik dari populasi yang sama ataupun dari populasi yang berbeda untuk membantu menjelaskan sistematika O. hypopthalmus berdasarkan data molekuler. Selain penambahan sampel, penggunaan marka genetik lain mungkin akan membantu menjelaskan clustering ini. Marka genetik yang disarankan adalah gen
cytochrome oxidase 1 (CO1) atau 16S DNA mitokondria.
Kata kunci: keragaman, struktur morfologis, cytochrome b, Kryptopterus, Ompok, Batang Hari
Hak Cipta milik IPB, tahun 2010
Hak cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tidak merugikan kepentingan yang wajar
IPB
Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya tulis dalam bentuk apapun tanpa izin IPB
KERAGAMAN STRUKTUR MORFOLOGIS DAN GEN
CYTOCHROME b
DNA MITOKONDRIA
Kryptopterus
spp. DAN
Ompok
spp. (SILURIDAE) DI DAS BATANG HARI JAMBI
ABDUL RAHMAN
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Biosains Hewan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2010
Judul Tesis : Keragaman struktur morfologis dan gen cytochrome b DNA mitokondria Kryptopterus spp. dan Ompok spp. (Siluridae) di DAS Batang Hari Jambi.
Nama : Abdul Rahman NIM : G352080031
Disetujui Komisi Pembimbing
Dr. Ir. Dedy Duryadi Solihin, DEA. Dr. Ir. Ridwan Affandi, DEA.
Ketua Anggota
Diketahui
Ketua Departemen Biologi Dekan Sekolah Pascasarjana
Dr. Ir. Ence Darmo Jaya Supena, M.Si. Prof. Dr. Ir. Khairil A. Notodiputro, M.S.
Buat (alm) Ayah, yang mengajariku banyak hal tentang hati; buat Mak, cinta, kasih sayang dan doa yang tidak pernah putus; buat @de’ dan anak-anakku, thiya dan niyya, harapan kalian yang membuat saya bertahan; untuk seluruh keluarga besar (alm) Abdul Kadir di Pasaman, Jambi dan Duri, keluarga besar Mertua di Bengkulu, terima kasih untuk bantuan moril dan materilnya. Semoga langkah tetap teguh dalam kejujuran, kemandirian dan kesederhanaan.
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah yang Maha Kuasa atas anugerah-Nya tulisan ini dapat diselesaikan. Terima kasih kepada komisi pembimbing, Dr. Ir. Dedy Duryadi Solihin, DEA dan Dr. Ir. H. Ridwan Affandi, DEA, atas arahan, saran dan bimbingannya. Terima kasih juga kepada jajaran pimpinan di Universitas Bengkulu atas izin untuk studi S2 ini, kepada Dirjen DIKTI dan jajarannya yang telah mengamanahkan beasiswa BPPS dari Rakyat Indonesia kepada saya.
Penulis juga mengucapkan terima kasih Para Nelayan di DAS batang Hari yang selalu berharap banyak agar penelitian ini bermanfaat nyata, kepada staf akademik dan non akademik di Departemen Biologi, Prof. Dr. Ir. Kadarwan Soewardi selaku penguji luar komisi ujian tesis, rekan-rekan di BSH dan di Wisma Nur, rekan-rekan di Laboratorium GMT Fakultas Peternakan, rekan-rekan alumni Unand di IPB serta seluruh pihak yang telah membantu penulis menyelesaikan S2 dan tesis ini. Semoga karya ilmiah ini bermanfaat.
Bogor, Desember 2010
RIWAYAT HIDUP
Penulis dilahirkan di Saba Singkam, Koto Nopan Rao, Pasaman, Sumatera Barat pada 20 Agustus 1981 sebagai putra dari Abdul Kadir (alm) dan Syamsimah. Pendidikan sarjana ditempuh dari tahun 1998 hingga 2004 di jurusan Biologi, Universitas Andalas, Padang. Penulis pernah menjadi staf pengajar di SMA Nusantara Jambi, SMAI Al-Falah Jambi, Lembaga Pendidikan Primagama Jambi dan Kerinci, dan Universitas Jambi. Tahun 2006, penulis diangkat menjadi dosen tetap di Universitas Bengkulu. Tahun 2008, penulis berkesempatan melanjutkan pendidikan S2 di Biosains Hewan, IPB dengan bantuan beasiswa dari Dikti (BPPS). Penulis menikah dengan Nopi Susilawati pada Februari 2008, dan telah dianugerahi dua orang putri, Aludra Fathiya R dan Hasni Haniyya R.
DAFTAR ISI
Halaman
DAFTAR ISI ... xiii
DAFTAR GAMBAR ... xvi
DAFTAR LAMPIRAN ... xvii
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan Penelitian ... 3
Manfaat Penelitian ... 4
TINJAUAN PUSTAKA ... 5
Kryptopterus spp. dan Ompok spp. ... 5
Meristik dan Morfometrik ... 6
Reproduksi dan Status Populasi ... 7
DNA Mitokondria ... 8
Cytochrome b ... 9
BAHAN DAN METODE ... 11
Waktu dan Lokasi Penelitian ... 11
Metode ... 11
Pengambilan Sampel ... 11
Pengukuran dan Pengambilan Data ... 13
Pengukuran Parameter Fisika-Kimia Perairan ... 13
Pengambilan Data Keragaman Struktur Morfologis ... 13
Pengambilan Data Keragaman Gen Cytochrome b... 14
Isolasi dan Purifikasi DNA Total dari Sampel Otot ... 14
Amplifikasi DNA Mitokondria Gen Cytochrome b ... 15
Visualisasi Pita DNA ... 15
Analisis Data ... 15
Analisis Data Parameter Fisika-Kimia Perairan ... 15
Analisis Data Keragaman Struktur Morfologis ... 16
Analisis Data Keragaman Gen Cytochrome b ... 16
HASIL ... 17
Posisi Geografis dan Administratif Lokasi Penelitian ... 17
Kondisi Perairan Lokasi Pengambilan Sampel ... 17
Keragaman Jenis dan Kelimpahan Ikan Lais di Setiap Lokasi Pengambilan Sampel ... 18
Keragaman Struktur Morfologis ... 19
Keragaman Gen Cytochrome b Berdasarkan Runutan Nukleotida ... 24
Keragaman Gen Cytochrome b Berdasarkan Runutan Asam Amino ... 29
Jarak Genetik dan Filogeni ... 31
PEMBAHASAN ... 36
Kondisi Perairan Lokasi Pengambilan Sampel ... 36
Keragaman Jenis dan Kelimpahan Ikan Lais di Setiap Lokasi Pengambilan Sampel ... 38
Keragaman Struktur Morfologis ... 39
Keragaman Gen Cytochrome b ... 40
SIMPULAN DAN SARAN ... 46
Simpulan ... 46
Saran ... 46
DAFTAR PUSTAKA ... 47
DAFTAR TABEL
Halaman
1 Jenis ikan lais pada kelompok sungai DAS Sunda Utara Purba ...2
2 Jenis dan jumlah individu ikan lais yang digunakan untuk analisis ...12
3 Jenis dan jumlah ikan lais yang ditemukan setiap periode pengambilan sampel ...19
4 Nilai varians struktur morfologis ikan lais di DAS Batang Hari ...20
5 Persentase pengelompokan spesies pada setiap lokasi pengambilan sampel ..22
6 Karakterisasi nukleotida pada beberapa perbandingan ikan lais ...24
7 Karakterisasi nukleotida pada K. limpok dan O. hypopthalmus ...25
8 Daftar situs diagnostik O. hypopthalmus dari DAS Batang Hari dengan DAS Kampar. ...26
9 Daftar situs variabel pada K. limpok dari DAS Batang Hari ...27
10 Daftar situs nukleotida pembeda K. limpok Batang Hari dengan Kampar ...28
11 Karakterisasi asam amino pada berbagai perbandingan jenis ikan lais ...30
12 Karakterisasi asam amino pada K. limpok dan O. hypopthalmus ...30
13 Jarak genetik ikan lais dari Batang Hari berdasarkan runutan gen cyt b parsial 927 bp dengan spesies pembanding dari genbank dan Kampar ...32
DAFTAR GAMBAR
Halaman
1 Peta lokasi pengambilan sampel ...11
2 Biplot PCA parameter fisika-kimia perairan lokasi pengambilan sampel ...18
3 Biplot PCA struktur morfologis seluruh sampel. ...21
4 Biplot PCA sampel ikan lais dari Pelayangan. ...22
5 Biplot PCA O. hypopthalmus ...23
6 Konstruksi filogeni seluruh ikan lais yang dianalisis berdasarkan nilai p-distance runutan nukleotida gen cyt b parsial 927 bp. ...33
7 Konstruksi filogeni seluruh ikan lais yang dianalisis berdasarkan nilai p-distance asam amino hasil translasi runutan gen cyt b parsial 927 bp. ...34
DAFTAR LAMPIRAN
Halaman
1 Peta lokasi pengambilan sampel ...53
2 Diagram kunci identifikasi jenis ikan lais yang ditemukan ...54
3 Data morfometrik baku yang diukur ...56
4 Data nisbah morfometrik yang dihitung ...57
5 Data parameter fisika-kimia lokasi pengambilan sampel ...58
6 Hasil uji t parameter fisika kimia lokasi pengambilan sampel ...59
7 Nisbah morfometrik seluruh sampel ikan lais yang ditemukan ...60
8 Biplot sampel ikan lais berdasarkan lokasi ...64
9 Biplot PCA K. limpok ...66
10 Penjajaran nukleotida gen cytochrome b parsial pada ikan lais ...67
11 Daftar situs nukleotida pembeda Kryptopterus dengan Ompok dari Sumatera ...83
12 Penjajaran asam amino hasil translasi gen cytochrome b parsial ikan lais ...84
13 Daftar perbedaan asam amino K. bichirris Batang Hari dan genbank dengan Kryptopterus secara umum ...89
14 Jarak genetik ikan lais dari Batang Hari dengan spesies pembanding berdasarkan runutan gen cyt b parsial 927 bp ...90
15 Konstruksi filogeni seluruh sampel ikan lais berdasarkan nilai distance Kimura 2 parameter dari runutan nukleotida gen cyt b parsial 927 bp ...91
PENDAHULUAN
Latar Belakang
Kryptopterus spp. dan Ompok spp. secara umum disebut dengan nama lokal “ikan lais”. Kedua genus ini sangat mirip dan hanya dibedakan dari penampakan sirip punggung. Kryptopterus spp. memiliki sirip punggung yang tereduksi dengan jari-jari sirip kurang dari empat. Ompok spp. memiliki sirip punggung normal dengan jari-jari empat atau lebih (Kottelat et al. 1993). Identifikasi secara molekuler pada ikan lais di daerah Kampar, Riau, menunjukkan kedua genus ini berbeda pada asam amino ke-155 dari runutan gen cytochrome b utuh. Asam amino ke-155 pada Kryptopterus spp. adalah valina (val) sedangkan pada Ompok
spp. adalah metionina (met) atau isoleusina (ile) (Elvyra 2009).
Daerah sebaran ikan lais terdapat di perairan tawar Indochina, Semenanjung Malaya hingga Kepulauan Sunda Besar (Sundaland). Ikan lais terutama ditemukan pada sungai-sungai yang bermuara ke arah Laut Cina Selatan dan Selat Malaka. Daerah sebaran ikan lais di Indonesia terdapat di Sumatera, Kalimatan dan Jawa, dengan sebaran utama di Sumatera dan Kalimantan. Perairan di Sumatera yang ditemukan keberadaan ikan lais adalah Sungai Musi (Weber & de Beaufort 1965), Batang Hari, Indragiri (Tan & Ng 2000), Kampar (Elvyra 2009), Rangau (Yustina 2001) dan Alas (Ng et al. 2004).
Salah satu perairan yang memiliki keragaman ikan lais yang tinggi adalah Daerah Aliran Sungai (DAS) Batang Hari. Delapan jenis Kryptopterus dan lima jenis Ompok ditemukan di sungai ini (Weber & de Beaufort 1965; Tan & Ng 2000; Tan & Kottelat 2009). Kedelapan jenis Kryptopterus tersebut adalah K. bichirris, K. kryptopterus, K. micronema, K. eugeneiatus, K. limpok, K. macrocephalus, K. palembangensis dan K. schilbeides. Lima jenis Ompok yang ditemukan adalah O. hypopthalmus, O. eugeneiatus, O. fumidus, O. leiachantus
dan O. rhadinurus.
Sungai Batang Hari merupakan salah satu sungai terbesar di Sumatera dengan panjang sekitar 600 km. Bagian hulu Sungai Batang Hari berasal dari daerah perbatasan Jambi dan Sumatera Barat. Sungai Batang Hari melintasi sebagian Sumatera Barat bagian selatan dan sebagian besar Provinsi Jambi.
Sungai ini bermuara di Selat Berhala, pantai Sumatera bagian timur (Sabiham & Hisao 1986). Sejarah geologis menunjukkan bahwa Sungai Batang Hari, bersama-sama dengan Sungai Musi, Indragiri dan Kapuas berasal dari DAS Sunda Utara Purba (Voris 2000).
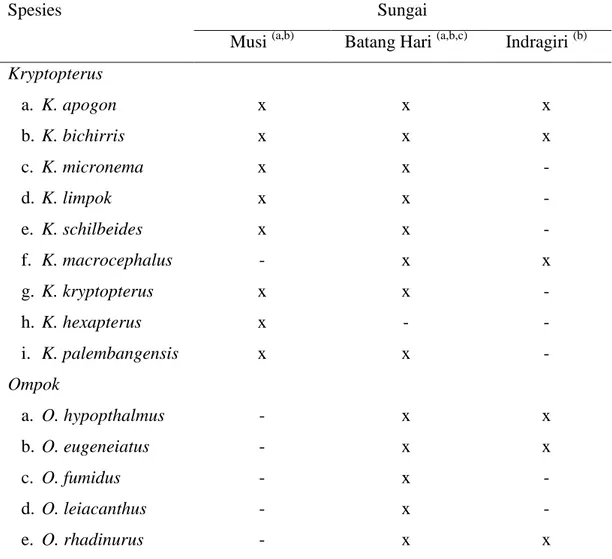
Kesamaan sejarah geologis diantara kelompok sungai DAS Sunda Utara Purba diduga akan berkaitan dengan jenis fauna air yang hidup di sungai-sungai tersebut. Hasil penelitian Weber dan de Beaufort (1965; Tan & Ng 2000; Tan & Kottelat 2009), menunjukkan bahwa jenis-jenis ikan lais yang ditemukan di kelompok sungai yang berasal dari DAS Sunda Utara Purba memiliki tingkat kesamaan jenis yang tinggi (Tabel 1).
Tabel 1 Jenis ikan lais pada kelompok sungai DAS Sunda Utara Purba
Spesies Sungai
Musi (a,b) Batang Hari (a,b,c) Indragiri (b)
Kryptopterus a. K. apogon b. K. bichirris c. K. micronema d. K. limpok e. K. schilbeides f. K. macrocephalus g. K. kryptopterus h. K. hexapterus i. K. palembangensis Ompok a. O. hypopthalmus b. O. eugeneiatus c. O. fumidus d. O. leiacanthus e. O. rhadinurus x x x x x - x x x - - - - - x x x x x x x - x x x x x x x x - - - x - - - x x - - x
*a = Weber dan de Beaufort (1965), b = Tan dan Ng (2000), c = Tan dan Kottelat (2009), x = ditemukan, - = tidak ditemukan.
3 Aliran Sungai Batang Hari sangat panjang dan melewati berbagai karakter topografi dan vegetasi, serta pola pemanfaatan. Hal ini diduga akan memunculkan variasi struktur morfologis dan genetik pada populasi fauna, termasuk ikan laisdi DAS Batang Hari. Selain itu, kesamaan sejarah geologis juga diduga akan berkaitan dengan keragaman dan karakter spesifik fauna antar sungai dalam kelompok DAS Sunda Utara Purba. Adanya keragaman dan karakter spesifik, baik secara struktur morfologis maupun genetik merupakan bagian adaptasi dari fauna tersebut terhadap lingkungan yang dihadapinya.
Informasi keragaman morfologis dan genetik pada suatu organisme sangat berguna untuk karakterisasi jenis, perkembangan dan distribusinya berdasarkan ruang dan waktu. Karakterisasi jenis, perkembangan dan distribusi populasi dibutuhkan untuk menentukan langkah konservasi dan pemanfaatan secara berkesinambungan. Selain itu, tingkat keragaman antarpopulasi, terutama keragaman genetik, dapat juga digunakan untuk memperkirakan tingkat resiko kepunahan suatu organisme (Lacy 1997). Kajian menyeluruh aspek morfologis, genetik dan ekologis dibutuhkan sebagai informasi untuk menentukan langkah pelestarian ikan lais di lingkungan alaminya, dan sebagai dasar untuk domestikasi ikan lais. Oleh sebab itu, penelitian tentang keragaman struktur morfologis dan genetik, antarspesies dan antarpopulasi ikan lais di DAS Batang Hari Jambi sangat perlu untuk dilakukan.
Salah satu marka genetik yang banyak digunakan untuk mengungkapkan keragaman genetik dalam populasi adalah gen cytochrome b DNA mitokondria (Farias et al. 2001). Gen cytochrome b telah terbukti efektif digunakan untuk melihat keragaman dan sebaran spesies ikan lais di daerah Kampar (Elvyra 2009).
Tujuan Penelitian
Menganalisis keragaman struktur morfologis dan keragaman gen
cytochrome b DNA mitokondria antarpopulasi dan antarspesies ikan lais di DAS Batang Hari, Jambi.
Manfaat Penelitian
Hasil penelitian ini diharapkan bermanfaat untuk:
1. Memperoleh data keragaman struktur morfologis dan gen cyt bKryptopterus
spp. dan Ompok spp. di DAS Batang Hari.
2. Memberikan data pembanding untuk verifikasi sistematika Kryptopterus
spp. dan Ompok spp. berdasarkan penampilan morfologis yang dikemukakan Weber dan de Beaufort (1965), dan Kottelat et al. (1993). 3. Memberikan data terbaru anggota dari jenis-jenis Kryptopterus dan Ompok
yang masih ditemukan di Sungai Batang Hari berdasarkan karakter morfologis dan genetik.
4. Data awal analisis filogeografi Kryptopterus spp. dan Ompok spp. pada kelompok sungai yang berasal dari DAS Sunda Utara Purba.
TINJAUAN PUSTAKA
Kryptopterus spp. dan Ompok spp.
Kryptopterus spp. dan Ompok spp. merupakan kelompok ikan air tawar yang termasuk dalam ordo Siluriformes, famili Siluridae. Famili Siluridae dikenal sebagai ikan bersungut (catfish) yang mengalami penciutan sirip punggung. Famili Siluridae memiliki satu atau dua pasang sungut pada rahang bawah dan rahang atas, dan sungut pada rahang atas biasanya lebih panjang (Nelson 2006). Famili Siluridae dibedakan dari famili lain dalam ordo Siluriformes berdasarkan ciri kepala dan badannya yang lebih pipih (Diogo 2005). Famili ini memiliki sirip anal sangat panjang terdiri dari 41-110 jari-jari lunak, dengan sirip perutkecil atau tidak ada, dan tidak memiliki sirip lemak (Nelson 2006). Sirip punggung tidak dilengkapi dengan jari-jari keras, dan jumlah jari-jari sirip kurang dari tujuh. Jari-jari keras pada sirip dada biasanya tidak kuat. Beberapa genera dalam famili Siluridae tidak memiliki sirip punggung (Diogo 2005).
Paling kurang ada 11 genera dan 97 spesies dalam famili Siluridae. Genera yang termasuk famili Siluridae antara lain Belodontichthys, Ceratoglanis,
Hemisilurus, Hito, Kryptopterus, Micronema, Ompok, Pterocryptis, Silurichthys,
Silurus (Parasilurus), dan Wallago (Nelson 2006). Genus Kryptoterus dan Ompok
memiliki warna dan bentuk tubuh yang mirip. Kedua genus ini secara umum disebut ikan lais (Elvyra 2009) dan hanya dibedakan dari penampakan sirip punggung (Kottelat et al. 2003). Kelompok ikan ini tersebar di perairan tawar Indochina, Thailand, Borneo dan Sumatera.
Penciri jenis (spesies) dalam genus Kryptopterus dan Ompok dapat didasarkan pada berbagai karakter. Weber dan de Beaufort (1965) mengutamakan jumlah jari-jari sirip sebagai kunci identifikasi jenis Kryptopterus. Kottelat et al.
(1993), menggunakan jumlah dan panjang sungut, dan jumlah jari-jari sirip anal untuk kunci identifikasi Kryptopterus dan Ompok. Ng (2004) menggunakan indeks perbandingan morfometrik untuk karakterisasi jenis Kryptopterus. Beberapa indeks morfometrik yang digunakan tersebut antara lain: perbandingan panjang, tinggi dan lebar kepala terhadap panjang baku, perbandingan panjang
sirip terhadap panjang baku, perbandingan panjang hidung terhadap panjang kepala dan perbandingan diameter mata terhadap panjang kepala.
Paling kurang 14 spesies Kryptopterus dan 7 spesies Ompok telah dideterminasi. Pembeda utama antar spesies dalam kedua genus ini didasarkan pada jumlah dan jarak terjauh sungut, dan jumlah sirip anal. Akan tetapi ada beberapa kelompok spesies yang memiliki sungut dan jumlah sirip anal yang mirip, sehingga diperlukan karakter lain untuk pembeda antar spesies. Karakter lain yang digunakan sebagai penciri spesies antara lain: pola warna badan, bentuk cuping sirip ekor, bentuk gigi vomer, dan beberapa indeks morfometrik. Indeks morfometrik yang digunakan adalah perbandingan kepala terhadap panjang baku, perbandingan diameter mata terhadap panjang kepala dan perbandingan panjang sirip dada terhadap panjang kepala (Kottelat et al. 1993).
Meristik dan Morfometrik
Karakter struktur morfologis telah umum digunakan dalam biologi ikan untuk mengukur tingkat perbedaan dan hubungan antar takson pada berbagai tingkatan (Turan 1999). Karakter struktur morfologis dapat dibagi atas karakter meristik dan morfometrik. Karakter meristik adalah struktur yang dapat dihitung seperti jumlah sisik, jumlah insang, pori-pori pada kepala dan sebagainya. Karakter morfometrik adalah karakter yang dapat diukur seperti panjang sirip, panjang kepala, diameter mata, atau perbandingan antar beberapa karakter yang dapat diukur (Helfman et al. 1997).
Pengukuran karakter morfometrik dapat digunakan untuk memperkirakan persediaan (ukuran populasi) ikan pada suatu lokasi (Misra & Easton 1999). Selain itu karakter morfometrik dapat digunakan untuk interpretasi habitat dan tingkah laku makan suatu spesies (Calvanti et al. 1999). Ketepatan analisis morfometrik dengan perkiraan ukuran populasi ikan dipengaruhi oleh jumlah karakter morfometrik yang dianalisis (Misra & Easton 1999). Penggunaan karakter-karakter tradisisonal banyak dikritik karena memiliki banyak bias dan kelemahan. Sebagai alternatif, perlu suatu sistem pengukuran morfometrik baru (truss system) sehingga analisis ukuran populasi ikan lebih tepat (Turan 1999).
7 Karakter meristik dan morfometrik dapat dianalisis dengan metode analisis komponen utama (PCA). PCA merupakan teknik analisis multivarian yang mampu mebedakan kelompok sama atau berbeda berdasarkan dua atau lebih karakter yang terukur (Quicke 1997). Konsep dasar PCA adalah analisis kelompok, karakter yang sama akan dikelompokkan pada satu kelompok dan karakter yang berbeda dipisahkan menjadi kelompok yang berbeda. Hasil dari PCA adalah pengelompokan fenetik atau yang sering disebut dengan fenogram (Ubaidillah & Sutrisno 2009).
Reproduksi dan Status Populasi
Usia matang gonad pada ikan lais berbeda antara jantan dan betina. Menurut Simanjuntak (2007), ikan lais betina mulai matang gonad saat panjang tubuh mencapai 115 mm, sedangkan lais jantan mulai matang gonad saat panjang tubuh mencapai 214 mm. Penelitian terbaru Elvyra (2010), menyatakan bahwa ikan lais betina baru mulai matang gonad saat mencapai ukuran 229 mm dengan berat 53.66 gram, sedangkan lais jantan mulai matang gonad saat berukuran panjang 226 mm dengan berat 48.04 gram. Ciri morfologis betina matang gonad ditandai dengan ovari dan telur berwarna kuning, mengisi hingga 2/3 rongga perut. Telur tersusun rapat dan butir telur mudah dipisahkan. Sedangkan jantan matang gonad ditandai dengan testis mengisi hingga ¼ rongga perut, gerigi testis lebar dan tebal, dan struktur testis menjadi lebih pejal (Elvyra 2009).
Pemijahan ikan lais hanya berlangsung satu musim dalam setahun (total spawner). Pemijahan mulai berlangsung pada awal musim penghujan dan mencapai puncaknya pada bulan Oktober (Simanjuntak 2007; Elvyra 2010). Namun hasil penelitian Simanjuntak (2007), dan Elvyra (2010) pada sungai yang sama menunjukkan sedikit perbedaan pada lama waktu pemijahan. Simanjuntak (2007), menyatakan bahwa pemijahan ikan lais berlangsung dari bulan Juni hingga Desember. Sedangkan Elvyra (2010), menyatakan waktu pemijahan ikan lais berlangsung lebih pendek yaitu hanya terjadi pada bulan September hingga November.
Pemijahan ikan lais dominan dilakukan di daerah rawa banjiran, sehingga ekosistem rawa banjiran memegang peranan penting dalam siklus reproduksi ikan
lais. Pemijahan ikan lais berlangsung sebelum terjadinya banjir maksimum. Hal ini merupakan strategi adaptasi ikan lais untuk mendapatkan keuntungan dari sumberdaya makanan alami yang tersedia bagi juwana ikan. Beberapa bulan kemudian ketika musim banjir besar tiba, anak-anak ikan lais sudah dapat mengkonsumsi anak-anak spesies ikan yang lain yang baru menetas (Simanjuntak 2007).
Nilai fekunditas rata-rata pada ikan lais adalah sebesar 6635 dengan diameter telur 0.77 mm. Nilai fekunditas adalah banyaknya jumlah telur yang ada dalam ovari sesaat sebelum ikan melakukan pemijahan. Tidak ada perbedaan yang signifikan pada diameter telur di ovari bagian anterior, tengah dan posterior. Hal ini memperkuat bukti bahwa ikan lais memiliki pola pemijahan serentak (total spawner). Fekunditas ikan lais hanya tergantung pada indeks kematangan gonad, dan tidak terpengaruh dengan bertambahnya ukuran panjang. Ikan lais yang telah mengalami pematangan gonad tidak akan memiliki fekunditas yang lebih tinggi walaupun memiliki ukuran tubuh yang lebih panjang (Elvyra 2010).
Belum ada usaha konservasi yang signifikan untuk melindungi ikan lais. Data IUCN (2010), menunjukkan hanya empat jenis ikan lais yang termasuk dalam daftar IUCN. Diantara keempat jenis tersebut hanya Ompok fumidus yang dikategorikan sebagai ikan lais terancam punah (vulnerable) dengan perkembangan populasi yang terus menurun. Sedangkan tiga jenis lainnya (O. leiachantus, K. lumholtzi dan K. micronema) dikategorikan sebagai tidak cukup data (data deficient). Elvyra (2009), menyarankan penangkapan ikan lais hanya dilakukan pada ikan yang telah melewati tahapan matang gonad, yaitu 229 mm pada betina dan 226 mm pada jantan. Berdasarkan waktu pemijahan, sebaiknya penangkapan hanya dilakukan setelah ikan melakukan pemijahan yaitu pada akhir bulan November dan harus dihindari penangkapan sebelum ikan melakukan pemijahan.
DNA Mitokondria
DNA mitokondria (mtDNA) merupakan DNA utas ganda yang pada umumnya berbentuk sirkuler. DNA mitokondria terdiri atas DNA utas berat (heavy strand) dan DNA utas ringan (light strand). Utas berat terdiri atas 2 gen
9 penyandi rRNA (16 S dan 12 S), 12 gen penyandi protein (NADH dehydrogenase
(ND)1, ND2, ND3, ND4, ND5, ND 4L, COI, COII, COIII, Cythochrome-b, ATPase 6 dan ATPase 8), 14 gen penyandi tRNA dan daerah bukan penyandi (dloop). Utas ringan terdiri dari 1 gen penyandi protein (ND 6) dan 8 gen penyandi tRNA (glutamic acid, proline, serine, tyrosine, cystein, asparagine, alanine dan glutaminaae). Berdasarkan fungsinya, genom mitokondria dibagi menjadi daerah penyandi (coding region) dan daerah bukan penyandi (non coding region). Daerah penyandi terdiri dari 13 gen penyandi protein, 2 gen penyandi rRNA dan 22 gen penyandi tRNA. Daerah bukan penyandi hanya ada pada daerah kontrol (control region). Daerah control region berperan dalam proses transkripsi dan replikasi genom mitokondria (Anderson et al. 1981).
Rekonstruksi pohon filogenetik dan sistematika pada berbagai kelompok hewan termasuk ikan lebih sering menggunakan analisis mtDNA dibanding DNA inti. DNA mitokondria berevolusi lebih cepat dibanding DNA inti, sehingga proses evolusi antar takson lebih mudah dideteksi (Kocher et al. 1989). Menurut Solihin (1994), mtDNA memiliki keunggulan sebagai penanda dalam keragaman genetik dan studi biologi populasi pada hewan karena: (i) mt DNA terdapat dalam jumlah kopi yang tinggi, sehingga lebih mudah diisolasi dan dipurifikasi; (ii) mtDNA berukuran relatif kecil sehingga dapat dipelajari sebagai kesatuan yang utuh; (iii) bagian-bagian mtDNA berevolusi dengan kecepatan yang berbeda sehingga dapat digunakan untuk studi sistematika dan zoogeografi; (iv) mtDNA diturunkan melalui jalur induk betina tanpa rekombinasi sehingga afinitas genetik yang diatur mtDNA merupakan refleksi dari filogeni matriarcale (garis induk betina); (v) mtDNA sangat polimorf, baik untuk intrapopulasi maupun untuk interspesies.
Cytochrome b
Gen cytochrome b (cyt b) mtDNA merupakan salah satu gen yang paling banyak dipelajari pada ikan. Ukuran gen cytochrome b pada ikan berkisar 1140 bp (Ketmaier et al. 2004; Doadrio & Perdices 2005) hingga 1143 bp (Perez et al. 2007). Seperti halnya sebagian besar gen pengkode protein, gen cyt b mtDNA
merupakan transmembran protein yang penting dalam rantai respirasi seluler organisme (Kocher et al. 1989).
Gen cyt b mtDNA dapat digunakan sebagai parameter klasifikasi pada berbagai tingkatan taksa, karena nilai mutasi pada gen ini berbeda berdasarkan posisi kodonnya (Irwin et al. 1991). Namun demikian, masih ada beberapa keraguan tentang kemampuan gen cyt b menghasilkan pohon filogeni yang akurat, apabila fragmen yang digunakan berukuran pendek (Kocher et al. 1989). Pada ikan, gen cyt b telah digunakan untuk menganalisis filogeni(Sullivan et al. 2004), sejarah dan struktur genetik antar populasi yang berbeda (Aboim et al. 2005), struktur genetik populasi yang terisolasi (Russel 2003) maupun sejarah biogeografi (Mateos et al. 2002).
Kodon (basa nitrogen) pertama dan kedua pada gen cyt b mtDNA memiliki nilai gamma yang rendah. Hal ini berarti bahwa kodon pertama dan kedua pada gen cyt b memiliki nilai substitusi yang rendah atau tidak bervariasi antar individu. Sebaliknya, kodon ketiga memiliki nilai gamma yang tinggi. Hal ini berarti bahwa kodon ketiga memiliki nilai substitusi yang tinggi, atau bervariasi antar individu (Farias et al. 2001). Akan tetapi, walaupun memiliki nilai substitusi yang tinggi, sebagian besar substitusi pada kodon ketiga bersifat saturation effect. Saturation effect adalah perubahan basa nukleotida yang tidak diikuti dengan perubahan asam amino yang ditranslasikan (Kocher et al. 1989).
Substitusi nukleotida terdiri atas substitusi transisi dan transversi. Substitusi transisi adalah substitusi basa nitrogen sejenis, purin menjadi purin atau pirimidin menjadi pirimidin. Substitusi transversi adalah perubahan nukleotida dari basa purin menjadi pirimidin atau sebaliknya. Pada gen penyandi protein, substitusi nukleotida dapat menghasilkan asam amino yang sama (substitusi silent/sinonim), maupun asam amino yang berbeda (substitusi non sinonim). Sebagian besar substitusi sinonim ditemukan akibat substitusi nukleotida pada kodon ketiga, walaupun dapat terjadi akibat substitusi nukleotida pada kodon pertama. Substitusi non sinonim dominan ditemukan akibat substitusi nukleotida pada kodon kedua dan pertama, walaupun dapat terjadi akibat substitusi nukleotida pada kodon ketiga (Nei & Kumar 2000).
BAHAN DAN METODE
Waktu dan Lokasi Penelitian
Penelitian ini dilaksanakan pada bulan Juli 2009 hingga Juni 2010. Pengambilan sampel dilakukan di DAS Batang Hari, Jambi. Analisis molekuler dilakukan di Laboratorium Biologi Molekuler, Pusat Penelitian Sumberdaya Hayati dan Bioteknologi (PPSHB), Lembaga Penelitian dan Pengabdian pada Masyarakat (LPPM), Institut Pertanian Bogor.
Metode
Pengambilan Sampel
Kryptoterus spp. dan Ompok spp. (lais) dikoleksi dari empat lokasi (titik) pengambilan sampel di DAS Batang Hari. Empat lokasi pengambilan sampel adalah: Desa Mandiangin Tebet (Mandiangin), Desa Sungai Bengkal (S. Bengkal), Desa Pelayangan dan Desa Simpang. Pengambilan lokasi pengambilan sampel didasarkan pada pola aliran Sungai Batang Hari (Gambar 1).
Gambar 1 Peta lokasi pengambilan sampel, 1: Mandiangin Tebet, 2: Sungai Bengkal, 3: Pelayangan, 4: Simpang, garis hijau = aliran sungai Batang Hari, garis kuning = aliran sungai Batang Tembesi.
Desa Mandiangin Tebet (1) dilewati Sungai Batang Tembesi. Sungai Batang Tembesi merupakan anak sungai terbesar dari DAS Batang Hari yang berasal dari daerah Taman Nasional Kerinci Seblat (TNKS). Desa Sungai Bengkal (2) dilewati aliran utama Sungai Batang Hari yang berasal dari daerah perbatasan Jambi
dengan Sumatera Barat. Sungai Batang Tembesi akan bertemu dengan Sungai Batang Hari di Desa Pelayangan (3). Sungai Batang Hari akan melewati Kabupaten Batang Hari, Kotamadya Jambi dan Kabupaten Muara Jambi sebelum bercabang menjadi dua sungai di Desa Simpang (4). Salah satu cabang melewati Taman Nasional Berbak, sehingga cabang ini disebut Sungai Berbak. Cabang yang lain melewati pelabuhan Muara Sabak dan tetap disebut Sungai Batang Hari. Kedua sungai ini akan bermuara di Selat Berhala di Pantai Sumatera Timur. Peta lokasi pengambilan sampel yang lebih rinci ditampilkan pada lampiran 1.
Pengambilan sampel dilakukan sebanyak tiga periode yang dianggap menggambarkan kondisi tahunan populasi ikan lais di DAS Batang Hari. Pengambilan sampel pada bulan Agustus 2009 mewakili kondisi surut terendah (puncak musim kemarau) di DAS Batang Hari. Pengambilan sampel pada bulan Desember 2009 mewakili awal musim penghujan, saat air Sungai Batang Hari mulai naik. Pengambilan sampel pada bulan April 2010 mewakili kondisi akhir musim penghujan (saat air Sungai Batang Hari mulai surut).
Kryptoterus spp. dan Ompok spp. yang telah dikoleksi, diidentifikasi berdasarkan Kottelat et al. (1993), dan Tan dan Ng (2000) (Lampiran 2). Jenis
Kryptopterus dan Ompok yang ditemukan beserta jumlah individu yang digunakan untuk analisis keragaman struktur morfologis dan genetik ditampilkan pada tabel 2. Sampel yang digunakan disimpan di Musium Zoologicum Bogoriense (MZB), Bogor sebagai spesimen sebaran lokasi ditemukannya ikan lais.
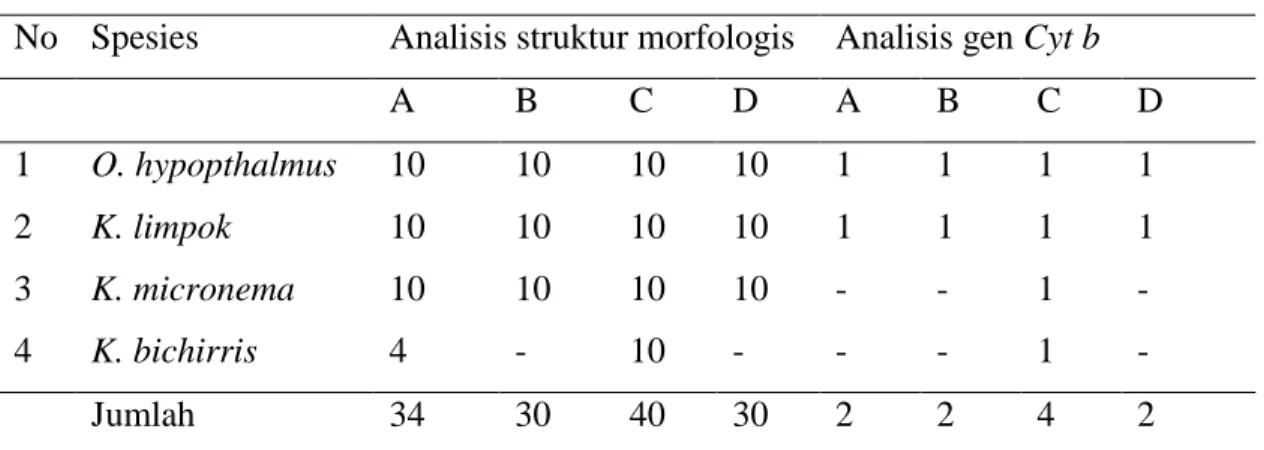
Tabel 2 Jenis dan jumlah individu ikan lais yang digunakan untuk analisis No Spesies Analisis struktur morfologis Analisis gen Cyt b
A B C D A B C D 1 O. hypopthalmus 10 10 10 10 1 1 1 1 2 K. limpok 10 10 10 10 1 1 1 1 3 K. micronema 10 10 10 10 - - 1 - 4 K. bichirris 4 - 10 - - - 1 - Jumlah 34 30 40 30 2 2 4 2
13
Perbandingan keragaman struktur morfologis dan gen cyt b antar lokasi pengambilan sampel hanya dilakukan pada K. limpok dan O. hypopthalmus karena kedua spesies ini merupakan spesies ikan lais yang paling umum ditemukan di DAS Batang Hari. K. limpok dan O. hypopthalmus dapat ditemukan sepanjang tahun di DAS Batang Hari, sedangkan K. micronema dan K. bichirris hanya dapat ditemukan saat air Sungai Batang Hari mulai naik (Desember) dan saat air Sungai Batang Hari mulai surut (April). Selain itu, K. bichirris hanya dapat ditemukan di stasiun Mandiangin dan Pelayangan dengan jumlah individu kurang dari target yang direncanakan. Target awal jumlah individu dalam penelitian ini adalah 10 individu/jenis/lokasi.
Pengukuran dan Pengambilan Data
Pengukuran Parameter Fisika-Kimia Perairan
Pada setiap lokasi pengambilan sampel dilakukan pengukuran parameter fisika kimia perairan. Parameter fisika yang diukur adalah ketinggian tempat, suhu perairan, kecepatan arus dan kecerahan. Ketinggian tempat diukur dengan GPS, suhu perairan diukur dengan termometer, kecepatan arus diukur dengan metoda bola hanyut, dan kecerahan diukur dengan secchi disk. Parameter kimia yang diukur adalah pH, oksigen terlarut (DO) dan alkalinitas. pH diukur dengan menggunakan pH meter, DO diukur dengan DO meter, dan alkalinitas diukur dengan metode titrasi asam basa.
Pengambilan Data Keragaman Struktur Morfologis
Data morfologi yang akan diukur pada setiap individu mengikuti metode baku pengukuran morfometrik ikan menurut Soewardi et al. (1995) yang telah dimodifikasi. Modifikasi yang dilakukan adalah mengganti karakter panjang total dan tinggi maksimum dengan karakter panjang sirip dada dan diameter mata (Lampiran 3). Selain itu, dilakukan penghitungan nilai tinggi bukaan mulut yang diperoleh berdasarkan perkalian nilai panjang rahang atas dengan √2 (Affandi et al. 2009).
Berdasarkan pengukuran morfometrik baku diatas, dilakukan pengukuran proporsi ukuran tubuh yang dikategorikan sebagai data nisbah morfometrik.
Penentuan nisbah data morfometrik yang dihitung mengacu pada Soewardi et al.
(1995) yang telah dimodifikasi. Modifikasi yang dilakukan adalah menghilangkan indeks yang berhubungan dengan karakter panjang total dan tinggi maksimum. Indeks panjang bagian tubuh di muka sirip punggung/panjang total diganti dengan indeks panjang bagian tubuh di muka sirip punggung/panjang baku. Dilakukan penambahan tiga indeks baru yaitu panjang sirip dada/panjang kepala, diameter mata/panjang kepala dan tinggi bukaan mulut/tinggi kepala (Lampiran 4).
Pengambilan Data Keragaman Gen Cytochrome b
Sampel yang digunakan dalam analisis keragaman genetik adalah jaringan otot yang terletak di dekat sirip ekor. Jaringan otot diambil sebesar 1 cm3 dan dimasukkan ke dalam tabung koleksi yang telah diisi etanol absolut (96%). Perbandingan sampel dengan etanol absolut dalam tabung koleksi dijaga tidak kurang dari 1:5. Sampel kemudian dibawa ke laboratorium untuk pengkodean DNA (DNA typing). Tahapan DNA typing meliputi isolasi dan purifikasi DNA total, amplifikasi gen target, visualisasi pita gen target dan sekuensing.
Isolasi dan Purifikasi DNA Total dari Sampel Otot
Isolasi dan purifikasi DNA total mengacu pada metode Sambrooks et al.
(1989) yang telah dimodifikasi Duryadi (1993). Otot ikan lais dicacah halus, dimasukkan ke dalam tabung polietilen, kemudian ditambahkan 500 μl digestion buffer. Selanjutnya sampel dihancurkan sampai halus dengan pengaduk gelas di dalam tabung polietilen. Setelah sampel cukup halus, ditambahkan lagi 250 μl
digestion buffer, digoyang sebentar, dan diinkubasi pada suhu 55ºC selama 12 jam. Setelah itu sampel disentrifugasi dengan kecepatan 6500 rpm selama beberapa detik, selanjutnya supernatan (cairan bagian atas) hasil sentrifugasi dipindahkan ke tabung polietilen baru.
Sampel kemudian ditambahkan 500 μl fenol, digoyang sampai tercampur rata, kemudian disentrifugasi dengan kecepatan 13000 rpm selama 3 menit. Supernatan dipindahkan ke tabung polietilen baru, ditambahkan 500 μl kloroform isoamil alkohol, digoyang sampai tercampur rata, kemudian disentrifugasi dengan kecepatan 13000 rpm selama 3 menit. Supernatan yang terbentuk dipindahkan ke tabung polietilen baru, ditambahkan etanol absolut dingin sebanyak 2 kali volume sampel, digoyang sebentar, kemudian disentrifugasi dengan kecepatan 13000 rpm
15 selama 5 menit. Selanjutnya etanol absolut dalam tabung polietilen tersebut dibuang. Endapan (pelet) yang tinggal dalam tabung polietilen ditambah 500 μl etanol 70%, digoyang sebentar dan disentrifugasi dengan kecepatan 13000 rpm selama 5 menit. Endapan yang terbentuk (DNA) yang diperoleh dikeringkan di udara terbuka. DNA yang diperoleh ditambahkan 100 μl larutan TE (Tris HCl - EDTA), digoyang sebentar, kemudian diinkubasi pada suhu 37ºC selama 15 menit. Sampel DNA total yang diperoleh disimpan dalam freezer.
Amplifikasi DNA Mitokondria Gen Cytochrome b
Amplifikasi gen cyt b dari mtDNA ikan lais dilakukan dengan metode
Polymerase Chain Reaction (PCR). Primer, komposisi bahan pereaksi PCR, dan kondisi reaksi PCR mengacu pada Elvira (2009). Primer yang digunakan adalah cbkr 1: 5‟-CCCGAAAAACTCACCCCTTA-3‟ dan cbkr 2: 5‟-ATAGCCCGGTTAGAGGGTTT-3‟. Target primer ini adalah gen cyt b
sepanjang 1104 bp. Komposisi (bahan) pereaksi yang digunakan dalam setiap reaksi PCR terdiri dari 10-100 ng sampel, 100 mM primer, 0.01 mM dNTP, 50 mM MgCl dan 1 unit Taq-polymerase. Kondisi PCR adalah sebagai berikut: predenaturasi selama 5 menit pada suhu 94oC, denaturasi selama 30 detik pada suhu 94oC, penempelan primer selama 45 detik pada suhu 60oC, elongation
selama 1 menit pada suhu 72oC dan post PCR pada suhu 72oC selama 5 menit. Proses PCR dilakukan sebanyak 35 siklus.
Visualisasi Pita DNA
Hasil PCR akan divisualisasi dengan metode elektroforesis pada gel agarose 1,2% dengan voltase 85 mV selama 40 menit. Hasil elektroforesis diwarnai dengan menggunakan ethidium bromide. Pita DNA yang telah diwarnai diamati dengan sinar ultraviolet. Hasil PCR yang teramplifikasi dengan baik akan dirunut (sequens).
Analisis Data
Analisis Data Parameter Fisika-Kimia Perairan
Data parameter fisika kimia perairan pagi, siang dan sore hari dirata-ratakan. Rata-rata data diolah dengan uji t pada taraf 5% untuk melihat apakah
parameter fisika kimia perairan antar lokasi pengambilan sampel berbeda nyata atau tidak. Jika hasil uji t taraf 5% berbeda nyata, uji t dilanjutkan pada taraf 1%.
Analisis Data Keragaman Struktur Morfologis
Data keragaman struktur morfologis diuji dengan metode principal component analysis (PCA) menggunakan program R (Everitt & Hothorn 2006). Hasil pengolahan data ini diharapkan akan menunjukkan perbedaan struktur morfologis antar spesies dan antar lokasi pengambilan sampel.
Analisis Data Keragaman Gen Cytochrome b
Pengolahan runutan gen cytochrome b dilakukan dengan program MEGA 4 (Tamura et al. 2007). Hasil runutan di aligment, kemudian dihitung keragaman dan jarak genetik intra dan antarpopulasi. Analisis pohon filogeni dari data runutan dilakukan dengan menggunakan metode Neighbour Joining. Data cyt b
mtDNA ikan lais yang diperoleh dibandingkan dengan data gen cyt b ikan lais dari gen bank (www.ncbi.nlm.nih.gov) dan Sungai Kampar (Elvyra 2009) untuk melihat pola standar baku isi runutan maupun variasi yang terjadi. Spesies dan kode akses ikan lais pembanding dari genbank adalah sebagai berikut:
Kryptopterus minor (AY458895), K. minor (DQ119481), K. limpok (DQ119431),
K. kryptopterus (DQ119434), K. schilbeides (DQ119482), O. bimaculatus
(DQ119433), O. bimaculatus (FJ711331), O. miostoma (DQ119435), O. pabda
HASIL
Posisi Geografis dan Administratif Lokasi Penelitian
Pengambilan sampel dilakukan pada empat stasiun yaitu: Desa Mandiangin Tebet (Mandiangin), Desa Sungai Bengkal (S. Bengkal), Desa Pelayangan dan Desa Simpang. Desa Mandiangin terletak pada ketinggian 29 mdpl dengan kordinat 02.01.923 LS dan 102.57.893 BT. Secara administratif Desa Mandiangin termasuk dalam Kecamatan Mandiangin, Kabupaten Sarolangun. Desa S. Bengkal terletak pada ketinggian 30 mdpl dengan kordinat 1.32.598 LS dan 102.43.232 BT. Secara administratif Desa S. Bengkal termasuk dalam Kecamatan Tebo Ilir, Kabupaten Muara Tebo. Desa Pelayangan terletak pada ketinggian 15 mdpl dengan kordinat 1.40.965 LS dan 103.5.851 BT. Secara administratif Desa Pelayangan termasuk dalam Kecamatan Muara Tembesi, Kabupaten Batang Hari. Desa Simpang terletak pada ketinggian 7 mdpl dengan kordinat 1.15.871 LS dan 104.05.550 BT. Secara administratif Desa Simpang termasuk dalam Kecamatan Berbak, Kabupaten Tanjung Jabung Timur.
Kondisi Perairan Lokasi Pengambilan Sampel
Analisis data parameter fisika-kimia lokasi pengambilan sampel dilakukan dengan metode Principal Component Analysis (PCA). Hasil analisis menunjukkan ke-empat stasiun pengambilan sampel terbagi dalam tiga kelompok, yaitu Mandiangin, Simpang dan gabungan S. Bengkal dengan Pelayangan (Gambar 2). Lokasi Mandiangin dicirikan dengan kecepatan arus dan kecerahan yang tinggi. Lokasi Simpang dicirikan dengan kecerahan yang tinggi. Tidak ada parameter fisika-kimia perairan yang dominan di lokasi S. Bengkal dan Pelayangan. Data parameter fisika-kimia lokasi pengambilan sampel ditampilkan pada lampiran 5.
Berdasarkan plotting data parameter fisika-kimia perairan dilakukan uji keterkaitan (korelasi) untuk melihat hubungan antarkarakter. Hasil uji korelasi Pearson menunjukkan bahwa kecepatan arus berbanding terbalik dengan tingkat alkalinitas (P = 0.02685), sedangkan pH berbanding terbalik dengan kecerahan (P= 0.02435). Korelasi antarkarakter lain seperti suhu dan DO tidak menunjukkan nilai korelasi yang signifikan.
Gambar 2 Biplot PCA parameter fisika-kimia perairan lokasi pengambilan sampel, 1=Mandiangin, 2=S. Bengkal, 3=Pelayangan, 4=Simpang.
Hasil uji t taraf 95% terhadap parameter fisika-kima perairan lokasi pengambilan sampel (Lampiran 6) menunjukkan keempat stasiun tidak memiliki parameter fisika-kimia yang berbeda nyata. Hal yang sama juga ditemukan ketika taraf uji t dinaikkan hingga 99%. Tidak ada lokasi yang memiliki parameter fisika-kimia yang berbeda nyata.
Keragaman Jenis dan Kelimpahan Ikan Lais di Setiap Lokasi Pengambilan Sampel
Ikan lais yang ditemukan dalam penelitian ini ada 4 jenis yaitu Ompok hypopthalmus, Kryptopterus micronema, K. limpok, dan K. bichirris (Tabel 3). Pengamatan selama periode pengambilan sampel menunjukkan bahwa O. hypopthalmus dan K. limpok merupakan jenis yang paling mudah ditemukan. Kedua jenis ini konsisten ditemukan setiap periode pengambilan sampel, yang berarti bahwa O. hypopthalmus dan K. limpok dapat ditemukan sepanjang tahun di DAS Batang hari. Diantara kedua spesies ini, O. hypopthalmus lebih sering ditemukan dan dengan jumlah individu tangkapan yang lebih banyak.
Kryptopterus micronema hanya ditemukan dua kali dari tiga kali periode pengambilan sampel, yaitu saat air Sungai Batang Hari mulai naik (awal musim penghujan) dan saat air Sungai Batang Hari mulai surut (akhir musim penghujan).
19 (pengambilan sampel ketiga), dan hanya ditemukan di stasiun Mandiangin dan Pelayangan.
Tabel 3 Jenis dan jumlah ikan lais yang ditemukan setiap periode pengambilan sampel
Nama jenis Periode pengambilan sampel Jumlah
Agustus 2009 Desember 2009 April 2010
A B C D A B C D A B C D O. hypopthalmus * - * - * - * * * * * * 40 K. limpok 3 * - - - * - - * * * * 40 K. micronema - - - 3 - 2 * * * * 40 K. bichirris - - - 4 - * - 14 Jumlah 134
*=Ditemukan dan dikoleksi dengan jumlah sampel maksimal 10 individu/jenis/lokasi, angka =hanya ditemukan sebanyak angka tersebut, -= tidak ditemukan, A= Mandiangin, B= S. Bengkal, C= Pelayangan, D= Simpang.
Berdasarkan periode pengambilan sampel ditemukan bahwa ikan lais dominan ditemukan saat akhir musim penghujan atau saat air Sungai Batang Hari mulai surut (April-Mei). Saat akhir musim penghujan disebut juga dengan musim puncak tangkapan ikan. Pada tahun 2010, musim puncak tangkapan ikan berlangsung pada akhir April hingga awal Mei. Menurut informasi nelayan di DAS Batang Hari, musim puncak tangkapan ikan pada tahun 2009 berlangsung satu bulan lebih cepat, yaitu pada pertengahan Maret. Musim puncak tangkapan ikan di daerah hulu (Mandiangin dan S. Bengkal) berlangsung lebih awal (lebih cepat) sekitar dua minggu dibandingkan dengan daerah hilir (Simpang).
Keragaman Struktur Morfologis
Pengukuran proporsi ukuran tubuh yang dikategorikan sebagai data nisbah morfometrik menunjukkan keragaman yang rendah pada intra dan antarpopulasi ikan lais di DAS Batang Hari. Nilai keragaman (varians) intra populasi pada 12 indeks struktur morfologis hanya berkisar 0.00–0.01 (χ = 0.13–1.20). Nilai varians antarpopulasi berkisar 0.01-0.51 (χ = 0.15–1.12) sedangkan varians antarspesies
pada seluruh sampel ikan lais dari DAS Batang Hari berkisar 0.01–0.26 (χ = 0.17– 1.01) (Tabel 4).
Tabel 4 Nilai varians struktur morfologis ikan lais di DAS Batang Hari
No Kode Sampel N1 N2 N3 N4 N5 N6 N7 N8 N9 N10 N11 N12 N13 1 Nh 0.00 0.00 0.00 0.00 0.01 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 2 Oh 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 3 Th 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 4 Kh 0.00 0.00 0.00 0.00 0.01 0.00 0.00 0.01 0.00 0.00 0.00 0.01 0.00 5 h total 0.00 0.00 0.01 0.03 0.03 0.02 0.02 0.06 0.02 0.02 0.02 0.01 0.02 6 Ni 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.01 0.01 7 Oi 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.01 8 Ti 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.01 0.01 9 Ki 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.02 0.00 0.00 0.00 0.00 0.00 10 i total 0.01 0.00 0.01 0.01 0.01 0.02 0.01 0.51 0.02 0.01 0.02 0.01 0.02 11 Nm 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.01 12 Om 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.01 0.01 0.00 0.00 0.00 0.00 13 Tm 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.04 0.00 0.00 0.00 0.02 0.01 14 Km 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.01 0.00 0.00 0.00 0.00 0.00 15 m total 0.00 0.00 0.01 0.00 0.00 0.01 0.00 0.14 0.01 0.03 0.01 0.00 0.01 16 Tb 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 17 Mandiangin 0.01 0.02 0.01 0.07 0.02 0.02 0.02 0.05 0.04 0.03 0.02 0.01 0.01 18 S. Bengkal 0.02 0.01 0.01 0.06 0.02 0.04 0.01 0.41 0.02 0.04 0.06 0.02 0.00 19 Pelayangan 0.01 0.01 0.01 0.15 0.02 0.01 0.01 0.12 0.04 0.08 0.04 0.04 0.01 20 Simpang 0.02 0.02 0.02 0.05 0.01 0.01 0.01 0.15 0.02 0.04 0.04 0.02 0.01 21 Batang Hari 0.01 0.02 0.01 0.07 0.02 0.03 0.01 0.26 0.03 0.05 0.04 0.02 0.01
*huruf awal kode sampel menunjukkan lokasi, sedangkan huruf kedua menunjukkan jenis. N= mandiangin, O= S. Bengkal, T= Pelayangan, K= Simpang, h = O. hypopthalmus, i = K. limpok, m = K. micronema, b = K. bichirris.
Indeks N8 (tinggi moncong/tinggi pangkal kepala), N4 (tinggi ekor/panjang baku), dan N10 (tinggi ekor/tinggi badan) memiliki varian tertinggi. Indeks N1 (panjang kepala/panjang baku), N3 (tinggi badan/panjang baku), dan N7 (panjang
21 hidung/panjang kepala) memiliki varian terendah. Hal ini berarti struktur tinggi merupakan karakter yang paling bervariasi diantara sampel ikan lais yang didapatkan, sedangkan karakter panjang kepala dan panjang baku relatif tidak berbeda. Berdasarkan nilai varian dapat dinyatakan bahwa struktur morfologis antarlokasi dan antarspesies ikan lais di DAS Batang Hari tidak beragam. Data lengkap proporsi ukuran tubuh (nisbah morfometrik) seluruh sampel ditampilkan pada lampiran 7.
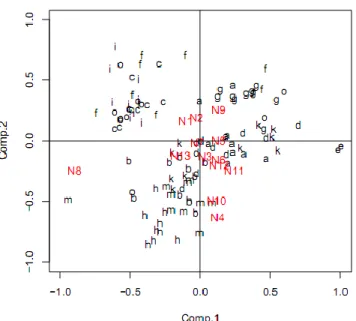
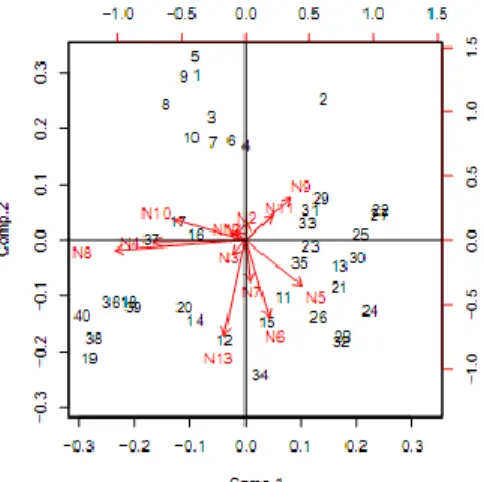
Hasil analisis keragaman morfometrik dengan metode PCA untuk seluruh sampel menunjukkan tidak ada populasi atau spesies yang mengumpul (mengelompok) secara sempurna (Gambar 3). Demikian juga dengan indeks struktur morfologis yang digunakan, tidak ada indeks yang dapat mencirikan spesies ataupun lokasi pengambilan sampel. Indeks N8 terlihat berada paling jauh dari sumbu utama PCA, akan tetapi indeks ini tidak mencirikan suatu populasi atau spesies tertentu.
Gambar 3 Biplot PCA struktur morfologis seluruh sampel, a=Nh, b=Ni, c=Nm, d=Oh, e=Oi, f=Om, g=Th, h=Ti, i=Tm, k= Kh, m= Ki, o= Km.
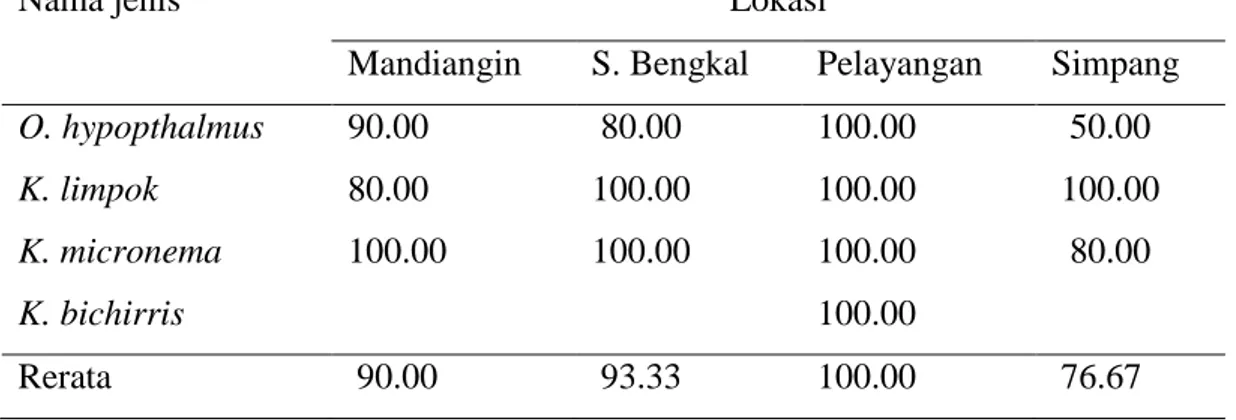
Berbeda dengan biplot pada seluruh sampel, biplot PCA ketika sampel dibagi menurut lokasi mampu mengelompokkan sampel berdasarkan spesies. Hasil analisis menunjukkan lebih dari 85% sampel pada setiap lokasi mengelompok berdasarkan spesies (Tabel 5). Pengelompokan yang sempurna
(100%) ditemukan pada sampel ikan lais yang berasal dari Pelayangan (Gambar 4). Individu ke-8 dari spesies K. micronema (nomor 28) walaupun terlihat terpisah dari kumpulan individu lain, akan tetapi masih berada dalam kuadran yang sama. Diagram biplot PCA dari lokasi Mandiangin, S. Bengkal dan Simpang ditampilkan pada lampiran 8.
Tabel 5 Persentase pengelompokan spesies pada setiap lokasi pengambilan sampel
Nama jenis Lokasi
Mandiangin S. Bengkal Pelayangan Simpang
O. hypopthalmus 90.00 80.00 100.00 50.00
K. limpok 80.00 100.00 100.00 100.00
K. micronema 100.00 100.00 100.00 80.00
K. bichirris 100.00
Rerata 90.00 93.33 100.00 76.67
Gambar 4 Biplot PCA sampel ikan lais dari Pelayangan, 1-10 = O. hypopthalmus, 11-20 = K. limpok, 21-30 = K. micronema, 31-40 = K. bichirris.
Pengelompokan sampel berdasarkan lokasi juga mampu menunjukkan karakter spesifik sebagai penciri spesies dalam kelompok ikan lais (Gambar 4, Lampiran 8). Ompok hypopthalmus dicirikan dengan indeks N9 (tinggi pangkal kepala/tinggi badan). Kryptopterus limpok dicirikan dengan indeks N8 (tinggi moncong/tinggi pangkal kepala). Kryptopterus bichirris dicirikan dengan indeks
23 N11 (diameter mata/panjang kepala) dan N12 (panjang sirip dada/panjang kepala). Kryptopterus micronema walaupun terletak mengelompok, akan tetapi tidak ada indeks morfometrik penciri jenis ini.
Analisis biplot PCA antarlokasi menunjukkan hasil yang berbeda pada K. limpok dan O. hypopthalmus. Ompok hypopthalmus dapat dikelompokkan berdasarkan lokasi (Gambar 5), sedangkan K. limpok hanya dapat dikelompokkan di lokasi S. Bengkal (Lampiran 9). Biplot PCA pada O. hypopthalmus
menunjukkan 75% individu pada spesies ini mengelompok berdasarkan lokasi. Pengelompokan tertinggi ditemukan pada O. hypopthalmus yang berasal dari Pelayangan. 90% individu O. hypopthalmus dari lokasi ini mengelompok di kuadran II. Pengelompokan terendah ditemukan pada O. hypopthalmus dari lokasi Simpang. 50% individu O. hypopthalmus di lokasi Simpang berada di kuadran I, sedangkan 50% yang lain berada di kuadran III.
Gambar 5 Biplot PCA O. hypopthalmus, 1-10 = Mandiangin, 11-20 = S. Bengkal, 21-30 = Pelayangan, 31-40 = Simpang.
Analisis biplot PCA pada O. hypopthalmus juga mampu menunjukkan indeks morfometrik penciri lokasi. Ompok hypopthalmus di Mandiangin dicirikan dengan indeks N4 (tinggi ekor/panjang baku) dan N10 (tinggi ekor/tinggi badan). Hal ini berarti O. hypopthalmus di Mandiangin memiliki struktur ekor yang lebih tinggi/tebal dibanding O. hypopthalmus dari lokasi lain. Ompok hypopthalmus di Pelayangan dicirikan dengan N6 (panjang rahang atas/panjang kepala), sedangkan
O. hypopthalmus di Simpang dicirikan dengan N8 (tinggi ‟moncong‟/tinggi pangkal kepala). Tidak ada penciri O. hypopthalmus dari S. Bengkal.
Keragaman Gen Cytochrome b Berdasarkan Runutan Nukleotida
Penjajaran (alignment) gen cyt b semua sampel dari Batang Hari dengan gen
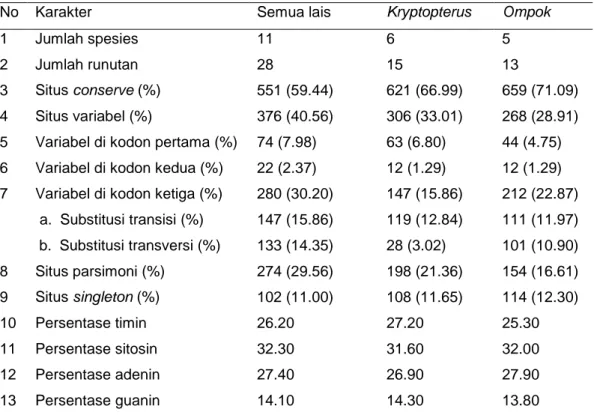
cyt b semua jenis ikan lais di genbank dan Kampar (Elvyra 2009) menghasilkan runutan nukleotida sepanjang 927 bp (Lampiran 10). Nukleotida yang berbeda (variabel) pada genus Kryptopterus dan Ompok tidak menunjukkan perbedaan yang signifikan (33.01% berbanding 28.91%) (Tabel 6).
Tabel 6 Karakterisasi nukleotida pada beberapa perbandingan ikan lais
No Karakter Semua lais Kryptopterus Ompok
1 Jumlah spesies 11 6 5
2 Jumlah runutan 28 15 13
3 Situs conserve (%) 551 (59.44) 621 (66.99) 659 (71.09) 4 Situs variabel(%) 376 (40.56) 306 (33.01) 268 (28.91) 5 Variabel di kodon pertama (%) 74 (7.98) 63 (6.80) 44 (4.75) 6 Variabel di kodon kedua (%) 22 (2.37) 12 (1.29) 12 (1.29) 7 Variabel di kodon ketiga (%) 280 (30.20) 147 (15.86) 212 (22.87)
a. Substitusi transisi (%) 147 (15.86) 119 (12.84) 111 (11.97) b. Substitusi transversi (%) 133 (14.35) 28 (3.02) 101 (10.90) 8 Situs parsimoni (%) 274 (29.56) 198 (21.36) 154 (16.61) 9 Situs singleton (%) 102 (11.00) 108 (11.65) 114 (12.30) 10 Persentase timin 26.20 27.20 25.30 11 Persentase sitosin 32.30 31.60 32.00 12 Persentase adenin 27.40 26.90 27.90 13 Persentase guanin 14.10 14.30 13.80
*situs parsimoni=ditemukan minimal dua jenis nukleotida, setiap jenis nukleotida dimiliki oleh minimal dua runutan, situs singleton=nukleotida yang berbeda hanya ditemukan pada satu runutan.
Berdasarkan posisi nukleotida pada triplet kodon, keragaman nukleotida terbesar secara berturut-turut adalah pada nukleotida ketiga, kesatu dan kedua. Berdasarkan jumlah keragaman nukleotida pada kodon pertama dan kedua, dapat dinyatakan bahwa keragaman asam amino pada genus Kryptopterus minimal sebesar 8.09%, sedangkan pada Ompok minimal sebesar 6.04%. Komposisi basa nitrogen terbesar penyusun runutan adalah sitosin, sedangkan guanin memiliki komposisi terkecil. Ditemukan satu situs nukleotida pembeda Kryptopterus dan