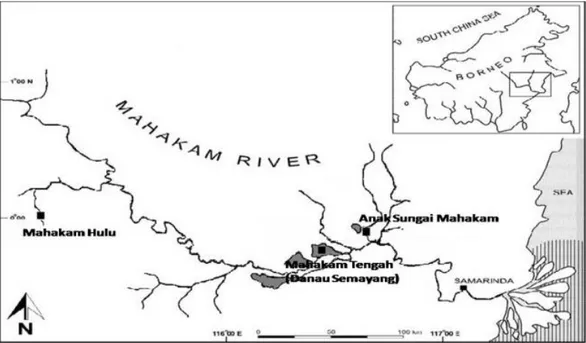
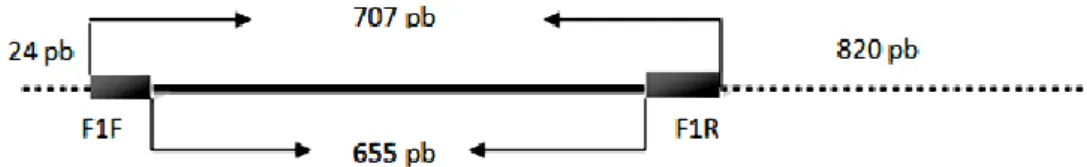
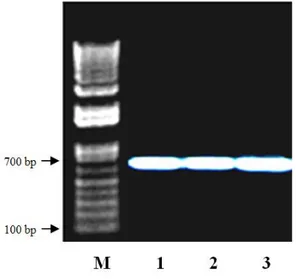
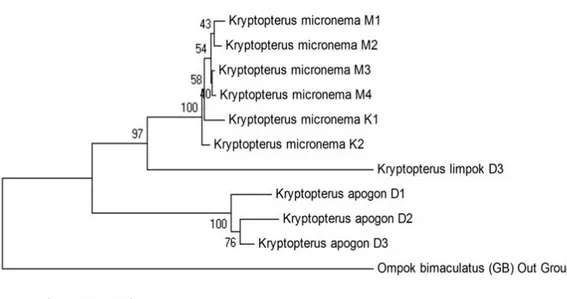
Kode batang DNA ikan lais genus Kryptopterus asal Sungai Mahakam. Kalimantan Timur]
Bebas
9
0
0
Teks penuh
Gambar
Dokumen terkait
