EFEKTIVITAS PENYERAPAN KARBONDIOKSIDA (CO2)
OLEH FITOPLANKTON (Chaetoceros sp.) PADA
FOTOBIOREAKTOR
MUHAMAD KEMAL IDRIS
SKRIPSI
DEPARTEMEN ILMU DAN TEKNOLOGI KELAUTAN FAKULTAS PERIKANAN DAN ILMU KELAUTAN
INSTITUT PERTANIAN BOGOR 2012
PERNYATAAN MENGENAI SKRIPSI DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa Skripsi yang berjudul:
EFEKTIVITAS PENYERAPAN KARBONDIOKSIDA (CO2)
OLEH FITOPLANKTON (Chaetoceros sp.) PADA
FOTOBIOREAKTOR
adalah benar merupakan hasil karya sendiri dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Semua sumber data dan informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka dibagian akhir Skripsi ini. Bogor, Februari 2012
MUHAMAD KEMAL IDRIS C54061248
oleh Fitoplankton (Chaetoceros sp.) pada Fotobioreaktor. Dibimbing oleh ALAN FRENDY KOROPITAN dan RAHMANIA DARMAWAN.
Penelitian dengan topik efektivitas penyerapan karbondioksida (CO2) oleh
fitoplankton (Chaetoceros sp.) pada fotobioreaktor, dilakukan selama 14 hari (satu siklus hidup fitoplankton). Analisis data nutrien dilakukan oleh laboratorium Produktivitas Lingkungan Hidup (Proling) MSP-IPB, sedangkan data karbon inorganik (DIC) dan karbon organik partikulat (POC) dianalisis oleh laboratorium Dinas Pekerjaan Umum, Jawa Timur. Penelitian ini dilakukan dengan
memanfaatkan gas CO2 yang diinjeksi ke dalam fotobioreaktor. Fotobioreaktor
tersebut telah diisi oleh inokulum fitoplankton air laut, yaitu Chaetoceros sp. Fotobioreaktor berkapasitas 50 liter diisi dengan 40 liter media kultur dan selama percobaan media diambil untuk keperluan analisis sebanyak 4 liter. Gas CO2 dialirkan ke dalam reaktor dengan sistem tertutup dari dasar reaktor dengan
menggunakan penyalur udara (air distributor) berpori halus. Pengukuran
dilakukan terhadap biomassa fitoplankton, parameter kualitas air dan konsentrasi gas CO2. Kultur Chaetoceros sp. dilakukan skala laboratorium dengan kondisi
parameter kualitas air terkontrol (Lampiran 4). Selama kultur berlangsung suhu terukur berkisar antara 22,14 - 26,16oC, suhu optimal terjadi pada hari kedelapan hingga ke sepuluh dengan kisaran suhu 22,39 - 24,67oC. Kisaran salinitasnya 25,34 - 27,65‰, salinitas optimalnya 26,66 - 27,65‰. pH media tumbuh berkisar antara 5,79 - 6,84, dengan pH optimal 6,27 - 6,68. Konsentrasi oksigen (DO) terlarut berkisar antara 0,52 - 1,52 ppm, DO optimalnya 1 - 1,52 ppm.
Pengukuran nutrien dikhususkan terhadap nitrat, nitrit, fosfat dan silikat. Pertumbuhan biomassa Chaetoceros sp. diikuti dengan dengan peningkatan kadar nitrit dari 0,106 µmol/kg menjadi 4,75 µmol/kg, sedangkan nitrat mengalami penurunan selama kultur dari 1,37 µmol/kg hingga mencapai 0,69 µmol/kg. Pada awal kultur, konsentrasi fosfat sebesar 10,67 µmol/kg berkurang hingga 1,94 µmol/kg diikuti dengan peningkatan biomassa Chaetoceros sp.. Silikat
mengalami penurunan dari 268,50 µmol/kg hingga 98,98 µmol/kg. Konsentrasi nitrat pada fotobioreaktor lebih rendah daripada kadar nitrit, fosfat dan silikat sehingga nitrat adalah faktor pembatas pada pertumbuhan Chaetoceros sp.. Dalam satu siklus fotobioreaktor (14 hari) Chaetoceros sp. mampu menyerap 10,56% vol. CO2 dari total 12% vol. yang diinjeksikan ke dalam fotobioreaktor
dengan laju penyerapan rata-rata 0,56% vol. CO2 per hari. Pada kondisi optimal
(hari ke-10), 1,35% vol. gas CO2 terlarut diserap oleh Chaetoceros sp. dan
© Hak cipta milik Muhamad Kemal Idris, tahun 2012 Hak cipta dilindungi
Dilarang mengutip dan memperbanyak tanpa izin tertulis dari Institut Pertanian Bogor, sebagian atau seluruhnya dalam bentuk apa pun, baik cetak, fotokopi, microfilm, dan sebagainya
OLEH FITOPLANKTON (Chaetoceros sp.) PADA
FOTOBIOREAKTOR
MUHAMAD KEMAL IDRIS SKRIPSI
sebagai salah satu syarat untuk memperoleh gelar Sarjana Ilmu Kelautan pada Departemen Ilmu dan Teknologi Kelautan
Fakultas Perikanan dan Ilmu Kelautan Institut Pertanian Bogor
DEPARTEMEN ILMU DAN TEKNOLOGI KELAUTAN FAKULTAS PERIKANAN DAN ILMU KELAUTAN
INSTITUT PERTANIAN BOGOR 2012
SKRIPSI
Judul Skripsi : EFEKTIVITAS PENYERAPAN KARBONDIOKSIDA (CO2) OLEH FITOPLANKTON (Chaetoceros sp.) PADA
FOTOBIOREAKTOR
Nama Mahasiswa : Muhamad Kemal Idris
Nomor Pokok : C54061248
Departemen : Ilmu dan Teknologi Kelautan Menyetujui, Dosen Pembimbing Utama Anggota
Dr. Alan F. Koropitan, S.Pi., M.Si. Rahmania A. Darmawan, M.Sc. NIP. 19751130 199903 1 003 NIP. 19711228 199703 2 002
Mengetahui, Ketua Departemen ITK
Prof. Dr. Ir. Setyo Budi Susilo, M.Sc. NIP. 19580909 1988303 1 003
. Tanggal Lulus: 15 Desember 2011
Radiasi matahari dipantulkan oleh permukaan bumi menuju ke luar
atmosfer, namun sebagian dari radiasi tersebut dipantulkan kembali ke permukaan bumi oleh gas rumah kaca. Hal tersebut mengakibatkan peningkatan suhu
permukaan bumi yang dikenal dengan pemanasan global. Pemanasan global dapat menyebabkan terjadinya perubahan cuaca, perubahan cuaca dalam jangka waktu yang panjang akan memicu terjadinya perubahan iklim. Oleh karena itu dilakukan berbagai upaya untuk menekan laju pemanasan global, salah satunya dengan mengurangi laju peningkatan gas karbondioksida (CO2) di atmosfer.
BPPT melakukan kultur fitoplankton yang dianggap dapat menyerap CO2
sehingga kadarnya dapat berkurang di atmosfer. Penyumbang CO2 terbesar salah
satunya adalah pabrik, oleh karena itu dilakukan kultur fitoplankton yang memanfaatkan injeksi CO2 dari cerobong asap pabrik. Namun sebelum itu perlu
dilakukan penelitian skala laboratorium untuk memastikan kemampuan
fitoplankton dalam menyerap CO2. BPPT memberikan peluang kepada penulis
untuk ikut berpartisipasi dalam penelitian ini, sehingga muncullah topik mengenai Efektivitas Penyerapan Karbondioksida oleh Fitoplankton (Chaetoceros sp.) pada Fotobioreaktor.
Penelitian ini merupakan tugas akhir yang dibuat sebagai salah satu syarat untuk memperoleh gelar kesarjanaan. Penulis mengucapkan terima kasih kepada ALLAH SWT atas nikmat sehat, iman dan islam, sehingga penulis dapat
menyelesaikan skripsi ini. Penulis juga mengucapkan terima kasih kepada kedua orang tua yang telah sabar menunggu kelulusan penulis. Terima kasih juga
kepada Bapak Dr. Alan Frendy Koropitan, S.Pi., M.Si., Ibu Rahmania A. Darmawan, M.Sc., dan Bapak Dr. Ir. Tri Prartono, M.Sc. selaku dosen
pembimbing dan penguji yang telah memberikan banyak masukan dan perbaikan selama penelitian. Ucapan terima kasih juga ditujukan kepada Bapak Dr. Ir. Richardus Kaswadji, M.Sc. yang telah memberikan saran mengenai penelitian, serta kedua orang tua, keluarga dan semua pihak yang telah membantu dalam penyusunan skripsi ini. Semoga skripsi ini dapat bermanfaat bagi semua pihak.
Bogor, Februari 2012
Muhamad Kemal Idris
i
Halaman
DAFTAR ISI ... i
DAFTAR GAMBAR ... ii
DAFTAR TABEL ... iii
DAFTAR LAMPIRAN ... iv 1. PENDAHULUAN ... 1 1.1. Latar belakang ... 1 1.2. Tujuan ... 2 2. TINJAUAN PUSTAKA ... 4 2.1. Fitoplankton Chaetoceros sp. ... 4 2.2. Fotobioreaktor ... 5
2.3. Parameter kualitas perairan ... 7
2.4. Nutrien ... 9
2.5. Sistem karbonat laut ... 13
3. METODOLOGI PENELITIAN ... 17
3.1. Waktu dan tempat ... 17
3.2. Alat dan bahan penelitian ... 17
3.3. Persiapan Penelitian ... 18
3.4. Pelaksanaan penelitian ... 22
4. HASIL DAN PEMBAHASAN ... 25
4.1. Pertumbuhan Chaetoceros sp. ... 25
4.2. Pertumbuhan Chaetoceros sp. dan penyerapan CO2 ... 27
4.3. Pengaruh nutrien (nitrat, nitrit, fosfat dan silikat) terhadap pertumbuhan Chaetoceros sp. ... 31
4.4. Pertumbuhan Chaetoceros sp. dan sistem karbonat dalam fotobioreaktor ... 35
5. KESIMPULAN DAN SARAN ... 38
5.1. Kesimpulan ... 38
5.2. Saran ... 38
DAFTAR PUSTAKA ... 39
ii
DAFTAR GAMBAR
Halaman
1. Skema penelitian ... 3
2. Chaetoceros lorenzianus ... 4
3. Kultur fitoplankton pada fotobioreaktor di PT. Indolakto yang dikembangkan oleh Pusat Teknologi Lingkungan - BPPT ... 6
4. Sistem karbonat di laut ... 14
5. Kadar relatif karbon anorganik terlarut pada kondisi pH tertentu ... 15
6. Skema fotobioreaktor ... 20
7. Tahapan kultur fitoplankton ... 21
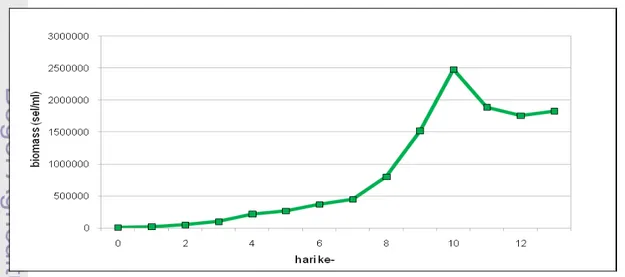
8. Pertumbuhan Chaetoceros sp. pada fotobioreaktor ... 26
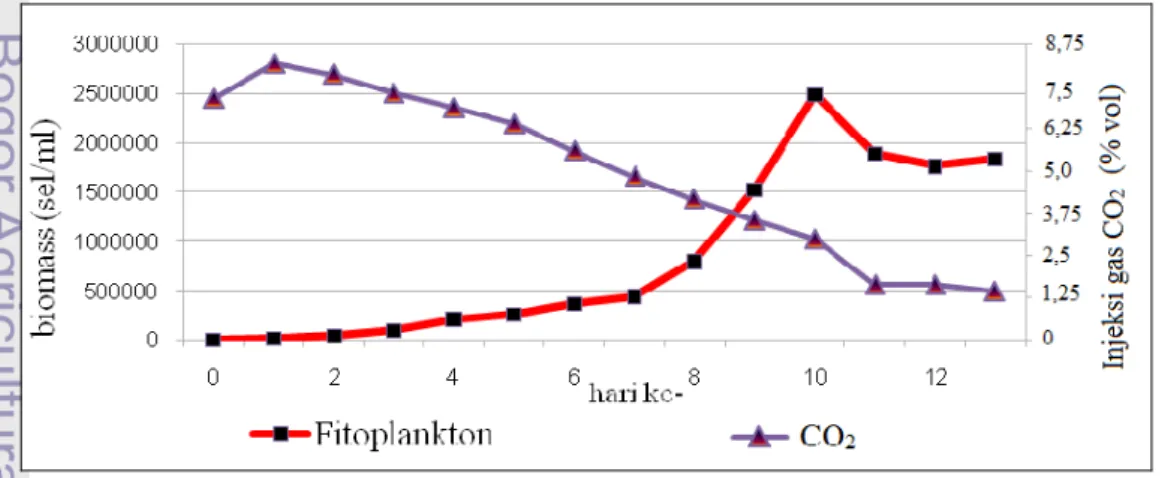
9. Pertumbuhan Chaetoceros sp. terhadap konsentrasi gas CO2 ... 28
10. Pertumbuhan Chaetoceros sp. terhadap nitrat, nitrit dan fosfat ... 32
11. Pertumbuhan Chaetoceros sp. terhadap silikat ... 34
12. Pertumbuhan Chaetoceros sp. terhadap POC dan DIC ... 36
iii Halaman 1. Klasifikasi Chaetoceros sp. ... 5 2. Bahan penelitian ... 17 3. Alat penelitian ... 18 4. Kegiatan penelitian ... 22
iv
DAFTAR LAMPIRAN
Halaman
1. Hasil pengukuran biomassa dan CO2 ... 43
2. Hasil pengukuran nutrien ... 43
3. Hasil pengukuran DIC dan POC ... 43
4. Hasil pengukuran parameter kualitas perairan ... 44
5. Pengukuran konsentrasi gas CO2 ... 45
6. Pengukuran kualitas perairan ... 46
7. Metode analisis nutrien ... 47
1 1.1 Latar belakang
Pemanasan global adalah permasalahan bersama yang dihadapi oleh seluruh umat manusia. Pemanasan global terjadi karena peningkatan suhu permukaan bumi akibat gas rumah kaca. Efek gas rumah kaca adalah peristiwa dimana radiasi cahaya matahari menembus permukaan bumi kemudian sebagian diserap dan dipantulkan kembali berupa gelombang infra merah. Gelombang inframerah yang dipantulkan bumi kemudian diserap oleh molekul gas seperti uap air (H2O), karbondioksida (CO2), metan (CH4), dan ozon (O3). Gas yang
menyerap gelombang inframerah disebut gas rumah kaca. Gelombang panas inframerah ini terperangkap dalam lapisan troposfir sehingga suhu permukaan bumi meningkat. Perubahan suhu menyebabkan kondisi atmosfer tidak stabil dan menimbulkan terjadinya anomali-anomali terhadap parameter cuaca yang
berlangsung lama. Dalam jangka panjang anomali-anomali parameter cuaca tersebut akan menyebabkan terjadinya perubahan iklim. Perubahan iklim berdampak pada kenaikan muka laut yang dapat menyebabkan erosi pantai, berkurangnya salinitas air laut dan meningkatnya resiko banjir (Susandi, 2008).
Pemanasan global menyebabkan penelitian yang berjudul Efektivitas Penyerapan Karbondioksida (CO2) oleh Fitoplankton (Chaetoceros sp.) pada
Fotobioreaktor ini penting dilakukan untuk mengurangi laju peningkatan emisi CO2 di atmosfer. Cerobong asap pabrik merupakan salah satu penyumbang gas
CO2 di atmosfer. Untuk mengurangi emisi gas tersebut digunakanlah
2
fitoplankton. Namun perlu dilakukan riset skala laboratorium di Puspitek, Serpong sebagai pendahulunya. Riset ini dilakukan dengan menginjeksi karbondioksida ke dalam fotobioreaktor saat mengkultur fitoplankton (Chaetoceros sp.). Hal tersebut bertujuan untuk menghitung berapa besar penyerapan karbondioksida yang dilakukan oleh fitoplankton. Riset yang dilakukan oleh Pusat Teknologi Lingkungan BPPT ini mengizinkan mahasiswa untuk berpartisipasi di dalamnya.
Penelitian ini dilakukan dengan memanfaatkan gas CO2 murni yang
diinjeksi ke dalam fotobioreaktor. Fotobiorektor tersebut telah diisi oleh
inokulum fitoplankton air laut, yaitu Chaetoceros sp. Fitoplankton yang dianggap mampu melakukan fotosintesis dengan bantuan cahaya lampu dengan intensitas 1500 hingga 2000 luks tentunya akan mencapai kondisi optimal pada saat menerima pasokan CO2 yang cukup. Pola perubahan konsentrasi nutrien (nitrat,
nitrit, silikat dan fosfat) dan sistem karbon (karbon organik partikulat/POC dan karbon anorganik terlarut/DIC) selama kultur akan terlihat pada fotobioreaktor, sehingga dapat dilakukan analisis secara deskriptif (studi literatur) terhadap
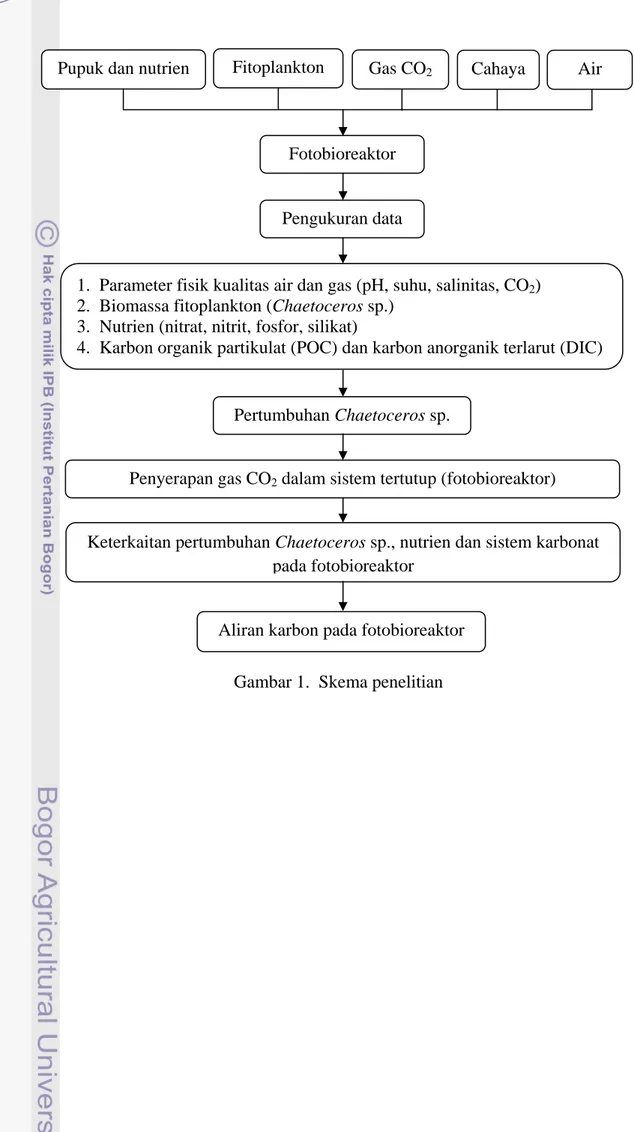
keterkaitan antara nutrien, sistem karbon dan pertumbuhan biomassa fitoplankton. Penjelasan singkat mengenai penelitian ini terlihat pada Gambar 1.
1.2 Tujuan
Penelitian ini bertujuan untuk mengkaji kemampuan Chaeoceros sp. dalam menyerap CO2 pada fotobioreaktor, serta keterkaitan antara pertumbuhan
Gambar 1. Skema penelitian Fotobioreaktor
Pengukuran data
1. Parameter fisik kualitas air dan gas (pH, suhu, salinitas, CO2)
2. Biomassa fitoplankton (Chaetoceros sp.) 3. Nutrien (nitrat, nitrit, fosfor, silikat)
4. Karbon organik partikulat (POC) dan karbon anorganik terlarut (DIC)
Pertumbuhan Chaetoceros sp.
Penyerapan gas CO2 dalam sistem tertutup (fotobioreaktor)
Pupuk dan nutrien Fitoplankton Gas CO2 Cahaya Air
Keterkaitan pertumbuhan Chaetoceros sp., nutrien dan sistem karbonat pada fotobioreaktor
4
2. TINJAUAN PUSTAKA
2.1 Fitoplankton Chaetoceros sp.
Chaetoceros sp. adalah salah satu spesies diatom. Diatom (filum Heterokontophyta, kelas Bacillariophyta) berbentuk uniseluler, walaupun demikian terdapat berbagai spesies agregat yang berkelompok dan membentuk koloni seperti rantai. Sel diatom tertutup oleh dinding sel yang terbuat dari silikat, bahan yang keras seperti gelas. Diatom adalah pabrik fotosintesis yang efisien, menghasilkan banyak makanan yang dibutuhkan makhluk hidup (makanan tersebut adalah diatom itu sendiri), serta oksigen (O2) sebagai hasil
fotosintesisnya. Diatom sangat penting di perairan terbuka, berperan sebagai produsen utama di daerah beriklim sedang dan kutub (Castro dan Huber, 2007).
Chaetoceros sp. adalah spesies fitoplankton yang tidak toksik terhadap manusia (Aunurohim et al., 2009). Chaetoceros sp. memiliki bentuk rantai memanjang yang merupakan gabungan dari beberapa sel pada tepi luarnya. Sepasang setal mengarah keluar pada sudut-sudut gabungan sel tersebut. Diameter katupnya berkisar antara 7 - 30 µm seperti pada Gambar 2.
Sumber : Johnson dan Allen (2005)
Tabel 1 menunjukkan klasifikasi Chaetoceros sp. dimana lebih dari dua lusin spesiesnya memiliki kelimpahan yang besar hampir sepanjang tahun pada perairan laut, baik di habitat muara maupun pesisir di sepanjang pantai Atlantik. Selain itu Chaetoceros sp. selalu menjadi bagian dari ledakan populasi diatom pada musim semi (Johnson dan Allen, 2005).
Tabel 1. Klasifikasi Chaetoceros sp. No. Takson Jenis
1. Kingdom Chromista 2. Filum Bacillariophyta 3. Kelas Mediophyceae 4. Orde Chaetocerotales 5. Famili Chaetocerotaceae 6. Genus Chaetoceros 7. Spesies Chaetoceros sp Sumber : Kawaroe et al. (2010)
2.2 Fotobioreaktor
Fotobioreaktor merupakan wadah atau tempat mereaksikan
mikroorganisme, dimana cahaya matahari masih dapat menembus masuk ke dalamnya. Alat ini digunakan sebagai reaktor eksperimen untuk mengetahui kemampuan fitoplankton dalam menyerap gas CO2. Selain itu juga berfungsi
untuk menurunkan gas CO2 dari sumbernya dan menghasilkan gas O2.
Fotosintesis fitoplankton mampu menyerap CO2 (dan NOx) dari cerobong
asap dimana untuk keperluan itu diperlukan teknologi pembudidaya fitoplankton salah satunya berupa fotobioreaktor. Fotobioreaktor meningkatkan produktivitas fitoplankton menjadi 2 hingga 5 kali lebih tinggi dari kondisi normalnya. Gas CO2 yang keluar dari suatu cerobong asap selanjutnya dapat langsung
6
dihubungkan ke fotobioreaktor dan dimanfaatkan oleh fitoplankton untuk pertumbuhannya melalui mekanisme fotosintesis (Setiawan et al., 2008).
Fotobioreaktor memiliki tinggi yang mencapai 160 cm dengan diameter luar 20 cm dan dalam 9,8 cm (Gambar 3). Volume reaktor 50 L serta dilengkapi dengan distributor udara, pompa, alat ukur tekanan, katup-katup dan lampu. Kecepatan injeksinya 0,03 liter per menit. Media diinjeksikan CO2 ke dalam
sistem dengan pompa. Sistem dilengkapi dengan penampungan gas untuk menyimpan karbondioksida sebelum diinjeksikan ke dalam fotobioreaktor. Sistem ini dijalankan dalam skala batch.
Gambar 3. Kultur fitoplankton pada fotobioreaktor di PT. Indolakto yang dikembangkan oleh Pusat Teknologi Lingkungan - BPPT
2.3 Parameter kualitas perairan
Pada saat melakukan kultur fitoplankton, kontrol terhadap parameter kualitas perairan perlu diperhatikan untuk menghindari terjadinya perubahan sistem metabolisme di dalam tubuh fitoplankton. Beberapa parameter tersebut antara lain suhu, salinitas, pH dan DO. Fitoplankton mampu melakukan
fotosintesis dengan mengubah energi cahaya menjadi biomassa. Pada saat kultur fitoplankton, pencahayaan harus diatur sedemikian rupa agar tidak menyebabkan terjadinya perubahan suhu. Suhu ruangan biasanya cukup bagi pertumbuhan fitoplankton (Hutagalung et al., 1997).
Suhu yang dapat ditoleransi oleh Chaetoceros sp. adalah 20 - 30 °C (pertumbuhan terjadi secara normal), sedangkan suhu optimalnya adalah 25 - 30 °C (Kawaroe et al., 2010). Suhu berpengaruh langsung terhadap laju fotosintesis tumbuhan khususnya reaksi enzimatis. Perubahan temperatur merupakan
indikator terjadinya proses perubahan kondisi kimia dan biologi perairan
(Aunurohim et al., 2009). Kenaikan suhu menyebabkan peningkatan kecepatan metabolisme dan respirasi organisme air (Kennish, 1990).
Salinitas adalah tingkat keasinan atau kadar garam terlarut dalam air. Kandungan garam pada air tawar kurang dari 0,05%. Jika lebih dari itu, air dikategorikan sebagai air payau atau menjadi saline bila konsentrasi garamnya mencapai 3 hingga 5% (Suriadikarta dan Sutriadi, 2007). Chaetoceros sp. mampu hidup pada kondisi salinitas minimal 6‰ akan tetapi yang optimal adalah 17 - 25‰ (Kawaroe et al., 2010). Perubahan salinitas secara signifikan akan
berbahaya bagi pertumbuhan organisme. Hal tersebut disebabkan karena proses osmosis di dalam sel sehingga tubuhnya akan kekurangan atau kelebihan cairan.
8
Ketidakseimbangan antara kadar larutan dalam sel (lebih pekat) dengan media lingkungannya menyebabkan cairan sel menjadi hiperosmosis, akibatnya sel membengkak dan pecah atau lisis (Yulianto, 1989).
Nilai pH adalah nilai dari hasil pengukuran ion hidrogen (H) di dalam air. Air dengan kandungan ion H+ tinggi akan bersifat asam, dan sebaliknya akan bersifat basa (Alkali). Nilai pH yang tinggi terjadi di perairan dengan kandungan fitoplankton tinggi, dimana proses fotosintesis membutuhkan banyak CO2. pH
akan mencapai 9 hingga 10, bahkan lebih tinggi jika bikarbonat diserap dari air (Svobodova et al., 1993). Stabilitas pH dipengaruhi oleh aktivitas respirasi dan fotosintesis. Respirasi akan menurunkan pH, dan sebaliknya fotosintesis
menaikan nilai pH (Malone dan Burden, 1988). Hubungan antara CO2 dengan pH
berbanding terbalik, semakin tinggi kadar CO2 maka semakin rendah nilai pH
(Sanusi, 2006).
Konsentrasi O2 terlarut adalah parameter penting dalam menentukan
kualitas perairan. Konsentrasi O2 dipengaruhi oleh keseimbangan antara produksi
dan konsumsi O2 dalam ekosistem. O2 diproduksi oleh komunitas autotrof
melalui proses fotosintesis dan dikonsumsi oleh semua organisme melalui pernapasan. Penurunan jumlah O2 dan peningkatan konsentrasi amoniak (NH3)
menjadi ancaman berbahaya bagi organisme. Konsentrasi O2 rendah akan
meningkatkan kecepatan respirasi, menurunkan efisiensi respirasi dan pertumbuhan yang dapat berakibat pada kematian massal (Izzati, 2008).
Oksigen bagi kehidupan organisme diperlukan terutama pada malam hari untuk kegiatan respirasi. Respirasi mendukung proses metabolisme organisme sehingga kandungan O2 terlarut dalam perairan sangat diperlukan bagi
kelangsungan proses pertumbuhannya (Ariyati et al., 2007). Peningkatan bahan organik juga meningkatkan konsumsi O2. Hal tersebut diakibatkan oleh
perombakan bahan organik yang dilakukan oleh mikroorganisme seperti bakteri (Wetzel, 1983). Kelarutan O2 di dalam laut dipengaruhi oleh suhu dan salinitas.
Semakin tinggi suhu dan salinitas perairan maka kelarutan O2 semakin kecil.
Pada umumnya lapisan permukaan laut mengandung O2 terlarut sebesar 4,5 - 9
mg/L (Sanusi, 2006).
2.4 Nutrien
Energi yang tersimpan dalam senyawa organik akan digunakan di dalam proses metabolisme, bila tidak digunakan akan dilepaskan sebagai panas
kemudian hilang dari sistem selamanya. Berbeda dengan energi, bahan yang berbentuk senyawa organik dapat digunakan berulang dalam suatu siklus. Bahan-bahan ini seperti nitrogen dan fosfor, awalnya berasal dari atmosfer atau
pelapukan batuan. Setelah itu terbentuk sebagai molekul anorganik sederhana, mereka akan dikonversi ke dalam bentuk lain dan dimasukkan ke dalam jaringan autotrof. Saat ini senyawa organik dipecah oleh pencernaan, pernapasan, dan dekomposisi. Hasilnya dilepaskan kembali ke lingkungan, dan siklus dimulai lagi (Castro dan Huber, 2007).
Media pertumbuhan fitoplankton menggunakan air laut maupun air tawar yang diperkaya dengan penambahan nutrien melalui pupuk. Air laut maupun tawar sudah mengandung berbagai elemen yang diperlukan fitoplankton walaupun dalam jumlah sangat sedikit. Oleh karena itu nutrien atau unsur-unsur makro dan mikro harus ditambahkan ke dalam media. Unsur-unsur makro yang diperlukan
10
secara langsung atau tidak langsung dalam pembentukan sel adalah C, H, O, N, P, S, K, Mg. Unsur mikro diperlukan dalam kadar sangat rendah sebagai katalisator, bahan dalam fungsi khusus atau regulasi osmotik ini diantaranya Fe, Mn, Cu, Zn, Mo, V, B, Cl, Co, Ca, Si, dan Na (Hutagalung et al., 1997).
Nitrat (NO3) adalah bentuk utama nitrogen di perairan alami dan
merupakan nutrien utama bagi pertumbuhan tanaman dan alga. Nitrat nitrogen sangat mudah larut dalam air dan bersifat stabil. Senyawa ini dihasilkan dari proses oksidasi sempurna senyawa nitrogen di perairan. Oksidasi amoniak menjadi NO2 dilakukan oleh bakteri Nitrosomonas, sedangkan oksidasi NO2
menjadi NO3 dilakukan oleh Nitrobacter. Kedua jenis bakteri tersebut merupakan
bakteri kemotrofik, yaitu bakteri yang mendapatkan energi dari proses kimiawi. NO3 yang merupakan sumber nitrogen bagi tumbuhan selanjutnya dikonversi
menjadi protein. Proses ini ditunjukkan sebagai berikut:
Sumber : Effendi (2003)
Kadar NO3 di perairan yang tidak tercemar biasanya lebih tinggi daripada kadar
ammonium (NH4). Kadar nitrat-nitrogen pada perairan alami hampir tidak pernah
lebih dari 0,1 mg/L. Kadar NO3 lebih dari 5 mg/L menggambarkan terjadinya
pencemaran antropogenik yang berasal dari aktivitas manusia dan kotoran hewan. Kadar nitrat-nitrogen yang lebih dari 0,2 mg/L dapat mengakibatkan terjadinya eutrofikasi perairan, yang selanjutnya menstimulir pertumbuhan alga dan tumbuhan air secara pesat (Effendi, 2003). Distribusi horisontal kadar NO3
semakin tinggi menuju ke arah pantai, dan kadar tertinggi biasanya ditemukan di perairan muara. Peningkatan kadar NO3 di laut disebabkan oleh masuknya limbah
rumah tangga atau pertanian (pemupukan) yang umumnya banyak mengandung NO3. Sedangkan distribusi vertikal NO3 di laut semakin bertambah seiring
dengan peningkatan kedalaman (Hutagalung et al., 1997).
Senyawa nitrit (NO2) yang terdapat dalam air laut merupakan hasil reduksi
senyawa NO3 atau oksidasi amoniak (NH3) oleh mikroorganisme. Selain itu
senyawa NO2 juga berasal dari hasil ekskresi fitoplankton, terutama saat
timbulnya ledakan populasi fitoplankton. Dalam air laut yang masih alami, kadar NO2 umumnya sangat rendah (kurang dari 0,1 µg/L). Distribusi vertikal kadar
NO2 semakin tinggi seiring dengan penambahan kedalaman laut dan semakin
rendahnya kadar O2. Sedangkan distribusi horisontal kadar NO2 semakin tinggi
apabila semakin menuju ke arah pantai dan muara sungai. Meningkatnya kadar NO2 di laut berkaitan erat dengan masuknya bahan organik yang mudah terurai
(baik yang mengandung nitrogen maupun tidak). Mikroorganisme mengurai bahan organik dengan memanfaatkan O2 dalam jumlah yang banyak, apabila tidak
cukup maka akan memanfaatkan NO3 sehingga NO3 berubah menjadi NO2. Oleh
karena itu senyawa NO2 menjadi salah satu indikator pencemaran (Hutagalung et
al., 1997). Pada perairan alami, NO2 biasanya ditemukan dalam jumlah yang
sangat sedikit, lebih sedikit dari NO3 karena bersifat tidak stabil dengan
keberadaan O2. Sumber NO2 dapat berupa limbah industri dan rumah tangga.
Perairan alami memiliki kadar NO2 sekitar 0,001 mg/L, sangat jarang menemukan
NO2 dengan kadar melebihi 1 mg/L. Kadar NO2 yang lebih dari 0,05 mg/L dapat
bersifat toksik bagi organisme perairan yang sangat sensitif. Sedangkan untuk keperluan peternakan, kadar NO2 10 mg/L masih dapat diterima. Bagi manusia
12
Pada perairan laut, unsur fosfor tidak ditemukan dalam bentuk bebas sebagai elemen, melainkan senyawa inorganik yang terlarut (ortofosfat dan polifosfat) dan senyawa organik berupa partikulat. Fosfat (PO4) adalah bentuk
fosfor yang dimanfaatkan oleh tumbuhan dan alga. Keberadaan fosfor relatif lebih sedikit dan mudah mengendap di kerak bumi. Fosfor merupakan unsur yang esensial bagi tumbuhan tingkat tinggi dan alga karena berpengaruh terhadap tingkat produktivitas perairan dan berperan dalam transfer energi di dalam sel (Adenosine Triphosphate dan Adenosin Diphosphate), sehingga menjadi faktor pembatas bagi tumbuhan dan alga. Ortofosfat (O-PO4) yang merupakan produk
ionisasi dari asam ortofosfat adalah bentuk fosfor yang paling sederhana dan banyak ditemukan di perairan. Kadar fosfor dalam ortofosfat jarang melebihi 0,1 mg/L, meskipun perairan tersebut eutrofik. Ortofosfat juga dapat dimanfaatkan secara langsung oleh tumbuhan akuatik (Effendi, 2003). Fosfat diadsorbsi oleh fitoplankton dan seterusnya masuk ke dalam rantai makanan. Pada air laut, kadar rata-rata PO4 sekitar 2 µg/L. Kadar ini semakin meningkat dengan masuknya
limbah rumah tangga, industri dan pertanian yang banyak mengandung PO4.
Peningkatan kadar PO4 dalam laut akan menyebabkan terjadinya ledakan populasi
fitoplankton (Hutagalung et al., 1997).
Silikon adalah salah satu unsur yang terdapat pada kerak bumi secara berlimpah. Silikon banyak ditemukan dalam bentuk silikat (SiO2). Silikat
bersifat tidak larut dalam air, baik yang bersifat asam maupun basa. Umumnya SiO2 berada dalam bentuk koloid. Sumber utama SiO2 adalah mineral kuarsa dan
feldspar, sedangkan sumber antropogenik SiO2 relatif sangat kecil. Silikon
alga terutama diatom, membutuhkan SiO2 untuk membentuk dinding sel. Pada
perairan payau dan laut, kadar SiO2 berkisar antara 1.000 - 4.000 mg/L (Effendi,
2003).
Silikat adalah nutrien yang sangat penting di laut. Tidak seperti nutrisi utama lainnya seperti PO4, NO3 dan NH4, yang dibutuhkan oleh hampir semua
plankton laut. Silikat adalah unsur kimia penting bagi biota tertentu seperti diatom, radiolarian, sillicoflagellates dan spons yang mengandung SiO2. Biota
seperti diatom adalah salah satu produsen yang paling penting di laut. Estimasi menunjukkan bahwa diatom memberikan kontribusi lebih dari 40% dari seluruh produksi primer. Oleh karena itu siklus SiO2 telah menerima perhatian ilmiah
yang signifikan dalam beberapa tahun terakhir dan banyak ilmuwan telah mempelajari perilaku SiO2 di lingkungan laut. Konsentrasi SiO2 terlarut di laut
sekitar 70,6 µmol/L dan masukan bersih SiO2 terlarut dari darat ke laut adalah (6.1
± 2.0)x102 mol setiap tahunnya. Kontribusi utama SiO2 (sekitar 80%) berasal dari
sungai (Jinming, 2010).
2.5 Sistem karbonat laut
Gas Karbondioksida (CO2) yang ada di atmosfer senantiasa
berkesetimbangan dengan CO2 terlarut di lautan. Hal tersebut menyebabkan
peningkatan konsentrasi CO2 di atmosfer berpengaruh terhadap lingkungan
perairan (Munandar, 2009). Karbondioksida dalam bentuk karbon anorganik terlarut diubah menjadi karbon organik melalui proses fotosintesis, kemudian kembali ke atmosfer melalui proses respirasi dan dekomposisi (Effendi, 2003). Penurunan CO2 dalam ekosistem akan meningkatkan pH perairan. Sebaliknya
14
proses respirasi oleh semua komponen ekosistem akan meningkatkan jumlah CO2,
sehingga pH perairan menurun (Wetzel, 1983). Pada umumnya, perairan alami mengandung CO2 sebesar 2 mg/L (Nontji, 1987).
Pada kisaran pH 8 terjadi proses hidrasi, dimana CO2 terlarut bereaksi
dengan molekul air menjadi asam karbonat (H2CO3). Selain itu komposisi
persentase kadar spesies CO2 adalah 0,4% H2CO3, 85,4% HCO3- dan 14,2% CO3
2-menunjukkan bahwa HCO3- dominan pada kondisi tersebut (Feely et al., 2001).
Lingkungan dengan pH basa memiliki konsentrasi ion hidroksida (HO-) yang cukup tinggi. Hal tersebut diakibatkan oleh serangan ion hidroksida yang bersifat elektronegatif kepada atom karbon (CO2) yang bersifat elektropositif. Setelah itu
apabila energi aktivasinya terpenuhi maka terbentuklah keadaan transisi yang merupakan penentu laju reaksi yang membentuk senyawa bikarbonat (HCO3-).
Tahap berikutnya yaitu terjadi disosiasi bikarbonat menjadi karbonat (CO32-)
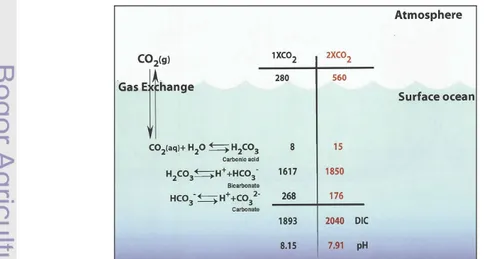
dengan melepaskan satu ion hidrogen (Munandar, 2009). Proses yang dialami oleh gas CO2 setelah difusi ke perairan laut menjadi CO2 terlarut dapat dilihat
pada Gambar 4.
Sumber : Feely et al. (2001)
Karbon anorganik terlarut terdistribusi dalam beberapa bentuk yaitu CO2
(aq), H2CO3, HCO3- dan CO3-. Kadar relatif keempat senyawa tersebut sangat
dipengaruhi oleh suhu, tekanan, pH dan salinitas (Munandar, 2009). Pada pH 8, total CO2 dominan berbentuk HCO3- dan sebagian kecil berbentuk H2CO3 dan
CO3-. Senyawa H2CO3 dominan terbentuk pada pH yang cenderung bersifat asam
(pH 6), sementara CO3- dominan terbentuk pada pH lebih besar dari 8 (Sanusi,
2006).
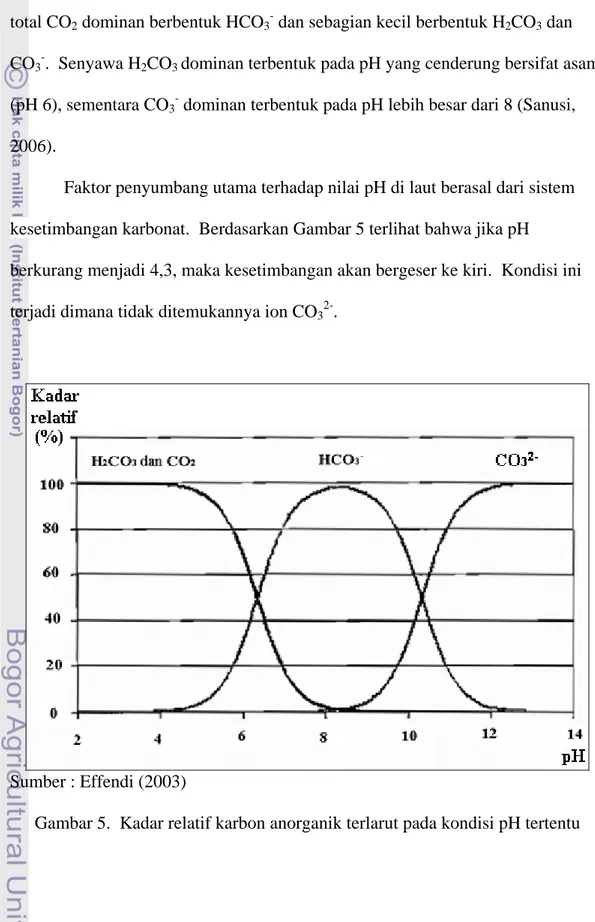
Faktor penyumbang utama terhadap nilai pH di laut berasal dari sistem kesetimbangan karbonat. Berdasarkan Gambar 5 terlihat bahwa jika pH
berkurang menjadi 4,3, maka kesetimbangan akan bergeser ke kiri. Kondisi ini terjadi dimana tidak ditemukannya ion CO32-.
Sumber : Effendi (2003)
16
Jika pH meningkat, maka kesetimbangan akan bergeser ke kanan dimana kadar CO2 dan H2CO3 mulai berkurang sehingga meningkatkan kadar HCO3-.
Pada pH 8,3 CO2 dan H2CO3 tidak ditemukan lagi, hanya terdapat ion HCO3- dan
CO32-. Oleh karena itu reaksi kesetimbangan akan berlangsung jika pH perairan
17 3.1 Waktu dan tempat
Penelitian kultur fitoplankton dilakukan selama 14 hari pengamatan untuk satu siklus hidup fitoplankton (Chaetoceros sp.). Penelitian ini berlangsung selama Bulan Agustus - Desember 2008. Penelitian bertempat di Balai Teknologi Lingkungan BPPT, Puspitek, Serpong. Selama kegiatan dilakukan pengukuran terhadap beberapa parameter kualitas air dan gas. Selain itu dilakukan analisis laboratorium seperti analisis nutrien (nitrat, nitrit, fosfat dan silikat), analisis karbon organik dan anorganik.
3.2 Alat dan bahan penelitian
Pengukuran parameter kualitas air dan gas menggunakan beberapa alat dan bahan seperti yang dapat dilihat pada Tabel 2 dan 3.
Tabel 2. Bahan penelitian
No. Nama Fungsi
1 Inokulan Chaetoceros sp. Bibit kultur fitoplankton
2 Air laut Media tumbuh fitoplankton
3 Akuades Membersihkan alat
4 Pupuk f2 Meningkatkan kadar nutrien
5 Klorin Menghilangkan HCl
6 HCl Membunuh bakteri patogen dan
kontaminan
7 Alkohol 70% Membersihkan sisa-sisa cairan kimia 8 Gas CO2 murni Gas untuk fotosintesis
9 Lugol Mengawetkan sampel fitoplankton
10 Natrium Tiosulfat Menghilangkan Klorin 11 Perangkat lunak Ms. Excel Menyajikan data
18
Tabel 3. Alat penelitian
No. Tipe alat Fungsi
1. Fotobioreaktor 50 liter Wadah kultur 2.
Portable Combination Gas Analyzer
RIKEN KEIKI Model RX-515 Alat ukur gas CO2 dan O2
3. Tabung gas CO2 murni Injeksi gas CO2
4. Kompresor Pemompa udara
5. Tempat penampungan udara
Pengunci udara agar tidak keluar dan masuk fotobioreaktor
6. Flow meter Pengatur tekanan udara
7. Haemocytometer Neubauer Improved Pengukuran biomassa fitoplankton
8. Filter gas Penyaring udara dari kotoran gas 9. Pipet tetes
Meletakkan sampel pada kaca preparat
10.
Hanna HI 9282 Multiparameter
water quality meter Alat ukur parameter fisik perairan
11. Mikroskop
Melihat jenis dan menghitung biomassa fitoplankton
12. Selang air Mengisi air ke dalam fotobioreaktor 13. Tabung reaksi Wadah pengukuran parameter fisik 14. Penyemprot cairan dan lap Membersihkan alat
Kultur murni Chaetoceros sp. diperoleh dari P2OLIPI, kemudian diperbanyak melalui kultur (scale up) di laboratorium. Konsentrasi gas CO2 diukur dengan
Portable Combination Gas Analyzer RIKEN KEIKI Model RX-515 (Lampiran 5). Parameter kualitas air seperti suhu, pH, salinitas diukur dengan HI 9282
Multiparameter water quality meter (Lampiran 6). Pengukuran biomassa fitoplankton dilakukan dengan menghitung jumlah fitoplankton menggunakan Haemocytometer Neubauer Improved. Pengunaan Haemocytometer Neubauer Improved sama seperti kaca preparat yang dilihat dengan bantuan mikroskop.
3.3 Persiapan Penelitian
Kepadatan awal kultur murni Chaetoceros sp. yang digunakan adalah 10.330 sel/ml. Media tumbuh yang digunakan adalah pupuk f2 dengan
penambahan silikat. Konstruksi fotobioreaktor dibuat dengan material akrilik berkapasitas 50 liter yang diisi dengan 40 liter media kultur dan selama percobaan media diambil untuk keperluan analisis sebanyak 4 liter. Gas CO2 dialirkan ke
dalam reaktor dengan sistem tertutup dari dasar reaktor dengan menggunakan penyalur udara berpori halus. Sebelum percobaan dimulai, terlebih dahulu dilakukan tes kebocoran terhadap air dan gas pada seluruh komponen fotobioreaktor.
Pada penelitian ini, sistem injeksi dilakukan satu kali ke dalam
penampungan gas (118 liter) yang berisi campuran gas antara CO2 dan N2 dengan
perbandingan 12% (CO2) : 88% (N2). Karbondioksida dengan konsentrasi sebesar
12 % vol. (120.000 ppm) diinjeksikan secara kontinu dengan laju injeksi sebesar 0,5 liter/menit dari penampungan gas ke dalam fotobioreaktor melalui bantuan kompresor. Konversi dari % vol. ke ppm adalah sebagai berikut: 1 % vol. dapat didefinisikan sebagai 10.000 ppm. Selain itu, sumber cahaya untuk penyinaran digunakan daylight lamp dengan intensitas penyinaran antara 1.500 dan 2.000 luks selama 12 jam dari pukul 06.00 hingga 18.00 (Setiawan et al., 2008).
Kultur Chaetoceros sp. dilakukan selama 14 hari, setelah itu hasil kultur dipanen dan tidak dimanfaatkan lagi sehingga tidak terjadi dekomposisi mineral di dalam fotobioreaktor. Fotobioreaktor dicuci kemudian digunakan untuk kultur fitoplankton yang baru, baik spesies yang sama maupun berbeda. Skema
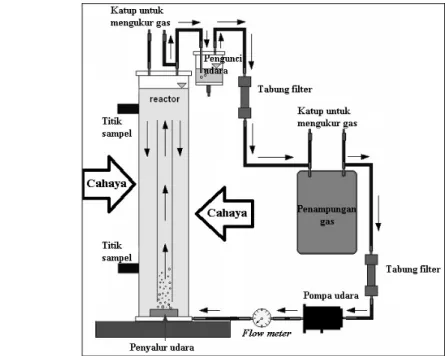
fotobioreaktor yang digunakan untuk mengkultur fitoplankton dapat dilihat pada Gambar 6.
20
Sumber : Setiawan et al. (2008)
Gambar 6. Skema fotobioreaktor
Berdasarkan Gambar 6 terlihat bahwa aliran udara yang membawa campuran gas akan mengisi fotobioreaktor secara menyeluruh sehingga membuat penyebaran CO2 terlarut yang akan dimanfaatkan oleh Chaetoceros sp. terdistribusi dengan
baik.
Pada tahap persiapan, terdapat tiga komponen penting diantaranya inokulan Chaetoceros sp., media kultur dan fotobioreaktor. Chaetoceros sp. diperbanyak (scale up) di laboratorium hingga kepadatannya 10.330 sel/ml. Media kultur yang berupa air laut disterilisasi dengan klorin yang bertujuan untuk membersihkan media kultur dari organisme yang dapat menghambat pertumbuhan Chaetoceros sp.. Setelah diaerasi dengan klorin, pada media ditambahkan
Na2S2O3 (Natrium Tiosulfat) untuk menetralisir klorin. Pupuk f2 dan
penambahan silikat dapat dimasukkan ke dalam media kultur setelah bau klorin hilang. Pemberian pupuk dilakukan untuk mempercepat pertumbuhan dengan
memberikan nutrisi yang dibutuhkan oleh Chaetoceros sp., sedangkan penambahan silikat bertujuan untuk memenuhi pembentukan cangkang
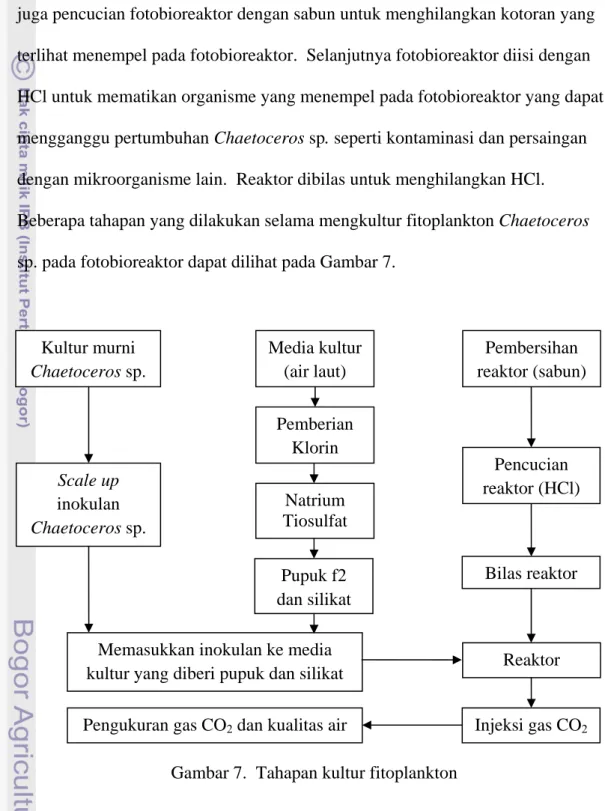
Chaetoceros sp.. Selain mempersiapkan inokulan dan media kultur, dilakukan juga pencucian fotobioreaktor dengan sabun untuk menghilangkan kotoran yang terlihat menempel pada fotobioreaktor. Selanjutnya fotobioreaktor diisi dengan HCl untuk mematikan organisme yang menempel pada fotobioreaktor yang dapat mengganggu pertumbuhan Chaetoceros sp. seperti kontaminasi dan persaingan dengan mikroorganisme lain. Reaktor dibilas untuk menghilangkan HCl. Beberapa tahapan yang dilakukan selama mengkultur fitoplankton Chaetoceros sp. pada fotobioreaktor dapat dilihat pada Gambar 7.
Gambar 7. Tahapan kultur fitoplankton Kultur murni Chaetoceros sp. Reaktor Scale up inokulan Chaetoceros sp. Natrium Tiosulfat
Memasukkan inokulan ke media kultur yang diberi pupuk dan silikat
Media kultur (air laut) Bilas reaktor Pemberian Klorin Pencucian reaktor (HCl) Pembersihan reaktor (sabun) Injeksi gas CO2
Pengukuran gas CO2 dan kualitas air
Pupuk f2 dan silikat
22
3.4 Pelaksanaan penelitian
Tahap pelaksanaan dimulai dengan memasukkan inokulan fitoplankton Chaetoceros sp. ke dalam media kultur (air laut dengan salinitas 25 - 26‰) yang telah diberi pupuk dan penambahan silikat. Setelah itu inokulan yang bercampur dengan media kultur dimasukkan ke dalam fotobioreaktor. Fotobioreaktor yang telah diisi oleh fitoplankton, kemudian diinjeksi dengan gas CO2 melalui pompa
udara, selanjutnya mengatur aliran tekanan udara (CO2) pada fotobioreaktor agar
stabil. Pengukuran gas CO2 dilakukan melalui katup yang terdapat pada pengunci
udara dengan bantuan alat RIKEN KEIKI Model RX-515, sedangkan parameter kualitas air diukur melalui sampel yang diambil dari fotobioreaktor dengan bantuan alat Hanna HI 9282 Multiparameter (Lampiran 6).
Pengukuran parameter kualitas air dilakukan setiap hari (09.00 dan 19.00) selama kultur melalui pengambilan sampel air laut pada fotobioreaktor seperti yang terdapat pada Tabel 4.
Tabel 4. Kegiatan penelitian
No. Kegiatan Waktu Tempat
1. Pengukuran suhu Setiap hari (14 hari) Puspitek, Serpong 2. Pengukuran salinitas Setiap hari (14 hari) Puspitek, Serpong 3. Pengukuran pH Setiap hari (14 hari) Puspitek, Serpong 4. Pengukuran DO Setiap hari (14 hari) Puspitek, Serpong 5. Pengukuran gas CO2 Setiap hari (14 hari) Puspitek, Serpong
6. Analisis nitrat Hari 1, 2, 4, 6, 8 dan 10 Lab. Proling, MSP-IPB 7. Analisis nitrit Hari 1, 2, 4, 6, 8 dan 10 Lab. Proling, MSP-IPB 8. Analisis fosfat Hari 1, 2, 4, 6, 8 dan 10 Lab. Proling, MSP-IPB 9. Analisis silikat Hari 1, 2, 4, 6, 8 dan 10 Lab. Proling, MSP-IPB 10. Analisis DIC Hari 1, 4 dan 10 Dinas PU Jawa Timur 11. Analisis POC Hari 1, 4 dan 10 Dinas PU Jawa Timur
Berdasarkan Tabel 2 pengukuran gas CO2 dilakukan tiga kali dalam sehari (09.00,
14.00 dan 19.00 WIB) karena tidak memerlukan sampel air laut.
Penghitungan penyerapan CO2 diukur dengan melihat aliran gas CO2 yang
terjadi selama proses kultur berlangsung. Gas CO2 yang masuk ke dalam
fotobioreaktor akan dibandingkan dengan gas CO2 yang keluar dari
fotobioreaktor, sehingga dapat diketahui berapa volume gas CO2 yang
dimanfaatkan oleh Chaetoceros sp. secara langsung. Setelah itu dilihat bagaimana keterkaitannya terhadap pertumbuhan biomassa Chaetoceros sp..
Keberadaan nutrien dalam fotobioreaktor diukur dua hari sekali (enam kali) selama kultur dengan cara mengambil 250 ml sampel kultur fitoplankton dari fotobioreaktor. Botol sampel yang digunakan adalah botol gelap atau berwarna agar terhindar dari cahaya matahari. Setelah itu sampel disimpan di dalam kulkas sebelum dikirim untuk dianalisis lebih lanjut di Laboratorium Produktivitas Lingkungan, Departemen Manajemen Sumberdaya Perairan, Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor. Metode yang dilakukan adalah spektrofotometri (detail metode ada pada Lampiran 7).
Sampel karbon anorganik terlarut (DIC) dan karbon organik partikulat (POC) diambil dari fotobioreaktor tiga kali selama kultur. Pengukuran DIC dilakukan dengan mengambil sekitar 250 ml sampel kultur fitoplankton dari fotobioreaktor lalu disaring dengan kertas saring Whatman. Hasil saringan yang berupa cairan (supernatan) diberi H2SO4 sebanyak 14 tetes lalu di simpan dalam
kulkas. Botol yang digunakan adalah botol kaca gelap dengan tutup berlapis teflon. Pada pengukuran POC, 250 ml sampel juga dimasukkan ke dalam botol gelap tanpa perlu disaring seperti pengukuran DIC. Kemudian sampel diberi 14
24
tetes H2SO4 dan disimpan dalam kulkas sebelum pengiriman ke Laboratorium Dinas Pekerjaan Umum Bina Marga, Pemerintah Propinsi Jawa Timur. Metode yang digunakan untuk mengukur DIC dan POC adalah SNI 01-6241-2000. (detail metode ada pada Lampiran 8).
25 4.1 Pertumbuhan Chaetoceros sp.
Kultur Chaetoceros sp. dilakukan skala laboratorium dengan kondisi parameter kualitas air terkontrol (Lampiran 4). Selama kultur berlangsung suhu terukur berkisar antara 22,14 - 26,16oC, suhu optimal terjadi pada hari kedelapan hingga ke sepuluh dengan kisaran suhu 22,39 - 24,67oC. Kisaran salinitasnya 25,34 - 27,65‰, salinitas optimalnya 26,66 - 27,65‰. pH media tumbuh berkisar antara 5,79 - 6,84, dengan pH optimal 6,27 - 6,68. Konsentrasi oksigen (DO) terlarut berkisar antara 0,52 - 1,52 ppm, DO optimalnya 1 - 1,52 ppm.
Salah satu faktor yang sangat penting bagi pertumbuhan biomassa Chaetoceros sp. adalah intensitas cahaya. Cahaya berperan dalam proses fotosintesis. Penelitian skala laboratorium ini mensimulasikan cahaya matahari yang biasanya digunakan oleh fitoplankton dengan daylight lamp berkapasitas 1500 - 2000 lux. Bila dikonversi ke dalam satuan energi (µmol photon.m-2.s-1) maka kapasitas penyinaran daylight lamp selama kultur berlangsung adalah 20,76 - 27,68 µmol photon.m-2.s-1. Nilai tersebut lebih kecil bila dibandingkan dengan cahaya yang optimal di perairan terbuka. Menurut Yuliana (2002), cahaya
optimal bagi proses fotosintesis fitoplankton berkisar antara 204,64 - 241,20 µmol photon.m-2.s-1.
Pertumbuhan Chaetoceros sp. dapat terlihat dari peningkatan biomassanya yang diukur melalui pengamatan mikroskop dengan bantuan Haemocytometer Neubauer Improved. Nilai biomassa ini dapat dijadikan indikator pertumbuhan yang terjadi selama satu siklus hidup Chaetoceros sp.. Pertumbuhan fitoplankton
26
secara umum dapat dibagi menjadi lima fase yang meliputi fase lag, fase
eksponensial, fase penurunan kecepatan tumbuh, fase stasioner dan fase kematian (Mata et al., 2010).
Selama kultur berlangsung tidak ditemukan adanya fase penurunan kecepatan tumbuh, hal tersebut dapat dilihat pada Gambar 8 dimana setelah fase eksponensial tidak ada peningkatan biomassa, tetapi yang terjadi adalah fase kematian yang ditandai dengan penurunan biomassa Chaetoceros sp.. Pada fase lag penambahan jumlah densitas Chaetoceros sp. cenderung linear dengan
peningkatan konstan dari 10.330 sel/ml menjadi 448.000 sel/ml pada hari ketujuh. Hal tersebut disebabkan karena sel-sel fitoplankton masih dalam proses adaptasi secara fisiologis terhadap media tumbuh sehingga metabolisme untuk tumbuh manjadi lambat.
Fase eksponensial terjadi pada hari kedelapan hingga kesepuluh dengan peningkatan biomassa Chaetoceros sp. dari 800.600 sel/ml menjadi 2.480.000 sel/ml (Gambar 8).
Fase eksponensial atau fase logaritmik yaitu fase dimana Chaetoceros sp. memiliki laju pertumbuhan tetap. Sel bereproduksi dengan cepat sehingga pertumbuhan populasi mencapai maksimal.
Pada hari kesepuluh hingga hari keduabelas terjadi fase kematian sebesar 720.000 sel/ml. Fase ini terjadi akibat kualitas fisik dan kimia kultur berada pada titik dimana sel tidak mampu lagi mengalami pembelahan (kematian). Pada hari keduabelas hingga akhir kultur terjadi kondisi dimana faktor pembatas dan kecepatan tumbuh sama karena jumlah sel yang membelah dan yang mati seimbang, kondisi ini disebut fase stasioner.
Keberhasilan kultur ditandai dengan pertumbuhan yang semakin meningkat dari kepadatan biomassa Chaetoceros sp.. Hal tersebut merupakan waktu generasi pertumbuhan Chaetoceros sp., sehingga dapat dikatakan waktu generasi merupakan waktu yang diperlukan oleh Chaetoceros sp. untuk membelah dari satu sel menjadi beberapa sel selama pertumbuhan. Pada penelitian ini kultur dilakukan selama dua minggu, bila diperpanjang tidak akan meningkatkan
biomassa karena keterbatasan faktor fisik dan kimia.
4.2 Pertumbuhan Chaetoceros sp. dan penyerapan CO2
Perubahan konsentrasi gas CO2 diamati setiap hari sesuai dengan
pengukuran biomassanya (Lampiran 1). Pertumbuhan Chaetoceros sp. dan gas CO2 memiliki hubungan korelasi negatif, dimana peningkatan biomassa diikuti
dengan penurunan konsentrasi gas CO2. Penurunan konsentrasi gas CO2 diduga
akibat difusi gas CO2 yang terjadi pada saat gas CO2 dialirkan ke dalam
28
konsentrasi karbon anorganik terlarut (DIC) pada fotobioreaktor. Karbondioksida terlarut inilah yang dimanfaatkan oleh Chaetoceros sp. dalam proses fotosintesis.
Pada awal kultur terjadi peningkatan konsentrasi CO2 dari 7,15% vol.
menjadi 8,17% vol.. Hal ini menunjukkan bahwa injeksi CO2 sebesar 7,15% vol.
tidak mengalami difusi di dalam fotobioreaktor, bahkan sebaliknya terjadi difusi dari CO2 terlarut dalam fotobioreaktor kembali menjadi gas CO2 menuju
penampungan gas.
Pada fase lag (adaptasi) hingga fase eksponensial terlihat bahwa
konsentrasi CO2 mengalami penurunan yang cenderung konstan (hari ke 2 - 10)
dengan kisaran penurunan 0,34 - 0,52% vol.. Rata-rata penurunan konsentrasi gas CO2 selama kedua fase tersebut adalah 0,574% vol. per hari. Setelah itu pada fase
kematian (hari ke 10 dan 11), konsentrasi gas CO2 mengalami penurunan terbesar
mencapai 1,35% vol. hingga menjadi 1,65% vol.. Hal tersebut diduga akibat tingginya pertumbuhan biomassa (puncak pertumbuhan) yang tidak diikuti dengan ketersediaan CO2 terlarut di dalam fotobioreaktor (keterbatasan CO2). Hubungan
pertumbuhan biomassa Chaetoceros sp. dengan konsentrasi CO2 pada
fotobioreaktor diperlihatkan oleh Gambar 9.
Pada fase stasioner konsentrasi gas CO2 tidak lebih dari 1,65% vol. dan
penurunan hanya sebesar 0,02% vol. dan 0,19% vol.. Hal tersebut diduga akibat keseimbangan antara ketersediaan CO2 dengan laju pertumbuhan biomassa
Chaetoceros sp.. Konsentrasi CO2 akhir sebesar 1,44% vol., mengalami
penurunan dari awal kultur sebesar 6,72% vol.. Bila dibandingkan dengan konsentrasi CO2 pada perairan terbuka, konsentrasi CO2 terlarut di perairan
pasifik khatulistiwa tertinggi sebesar 750 ppm dan yang rendah sebesar 150 ppm (Beardall dan Stojkovic, 2006). Konsentrasi CO2 terlarut pada fotobioreaktor
lebih rendah dengan kisaran 19 ppm hingga 78,5 ppm (Lampiran 1).
Perhitungan penyerapan CO2 pada fotobioreaktor perlu dilakukan untuk
mengetahui aliran gas CO2 yang terjadi dalam sistem tertutup seperti yang terlihat
pada Tabel 5.
Tabel 5. Penyerapan karbondioksida pada fotobioreaktor Hari
ke
Penyerapan CO2 Penyerapan
CO2 (%)
Masuk (% vol.) Keluar (% vol.) Serapan (% vol.)
0 7,15 8,17 -1,02 31,92 1 8,17 7,83 0,34 34,75 2 7,83 7,3 0,53 39,17 3 7,3 6,87 0,43 42,75 4 6,87 6,4 0,47 46,67 5 6,4 5,57 0,83 53,58 6 5,57 4,83 0,74 59,75 7 4,83 4,15 0,68 65,42 8 4,15 3,52 0,63 70,67 9 3,52 3,0 0,52 75 10 3,0 1,62 1,35 86,5 11 1,62 1,63 0,02 86,42 12 1,63 1,44 0,19 88 13 1,44 - - -
30
Berdasarkan Tabel 5, aliran gas CO2 pada sistem tertutup (fotobioreaktor)
memiliki konsentrasi awal sebesar 7,15% vol. yang dihitung selisihnya terhadap gas yang keluar dari fotobioreaktor. Kemudian dapat diketahui seberapa besar penyerapan gas CO2 yang dimanfaatkan oleh Chaetoceros sp. pada
fotobioreaktor. Sistem tertutup menyebabkan minimnya kemungkinan gas keluar dari sistem sehingga asumsi yang digunakan adalah konsentrasi gas yang keluar dari fotobioreaktor pada hari pertama akan menjadi konsentrasi gas masukan pada hari kedua, dan seterusnya. Selisih konsentrasi yang terjadi diduga akibat difusi gas CO2 ke dalam media tumbuh (air laut), hasil difusi gas tersebut secara
langsung dimanfaatkan oleh Chaetoceros sp. untuk fotosintesis karena tidak ada persaingan dengan spesies yang lain. Oleh karena itu selisih tersebut dianggap sebagai penyerapan CO2 oleh Chaetoceros sp.. Pada awal kultur (hari ke 0) tidak
terjadi penyerapan CO2, ditunjukkan dengan negatifnya nilai penyerapan -1,02%
vol.. Hal ini diduga akibat masuknya gas CO2 yang melimpah dan tidak
diimbangi dengan kepadatan biomassa Chaetoceros sp. sebesar 10.330 sel/ml. Berbeda dengan hari berikutnya, penyerapan gas CO2 meningkat sebesar 0,34%
vol. yang diduga akibat pemanfaatan CO2 oleh Chaetoceros sp. dalam
fotobioreaktor.
Pada hari kedua hingga kelima adalah fase lag (adaptasi), hasil
pengukuran outlet gas CO2 sebesar 7,83 - 5,57% vol. dengan laju penyerapan 0,43
- 0,83% vol. dari injeksi yang diberikan pada fotobioreaktor. Pada hari kesepuluh, 1,35% vol. gas CO2 terlarut dengan penyerapan sebesar 46% dari injeksi pada hari
ke-10. Selanjutnya pada fase stasioner penyerapannya berkisar antara 0,02 - 0,19% vol.. Selama kultur berlangsung penyerapan CO2 tertinggi oleh
Chaetoceros sp. terjadi pada fase eksponensial (hari kesepuluh) sebesar 1,35% vol. diikuti dengan peningkatan biomassa sebesar 960.000 sel/ml akibat kecepatan reproduksi sel yang maksimal. Pada penelitian ini, dalam satu siklus
fotobioreaktor (14 hari) Chaetoceros sp. mampu menyerap 10,56% vol. CO2 dari
total 12% vol. CO2 yang diinjeksikan ke dalam fotobioreaktor dengan laju
penyerapan rata-rata 0,56% vol. CO2 per hari.
4.3 Pengaruh nutrien (nitrat, nitrit, fosfat dan silikat) terhadap pertumbuhan Chaetoceros sp.
Nutrien dalam jumlah yang sedikit di perairan menjadi faktor pembatas. Pada penelitian ini pengukuran nutrien dikhususkan kepada nitrat, nitrit, fosfat dan silikat (Lampiran 2). Nitrat adalah senyawa anorganik yang berperan sebagai nutrisi makhluk hidup autotrof untuk tumbuh dan berkembang (Alaerts, 1987). Nitrit bersifat toksik, peningkatan konsentrasi nitrit dapat menyebabkan
penurunan biomassa Chaetoceros sp.. Fosfat berperan dalam proses metabolisme serta transfer energi di dalam sel (Boyd, 1982). Silikat lebih banyak digunakan oleh diatom dalam pembentukan dinding sel. Ketersediaan nutrien pada
fotobioreaktor sangat terbatas, hal ini dikarenakan sistem batch (tertutup), dimana tidak adanya pergerakan massa air yang membawa sejumlah nutrien, gas dan organisme lain ke dalam fotobioreaktor.
Pertumbuhan Chaetoceros sp. pada penelitian ini memiliki hubungan korelasi negatif terhadap nitrat, artinya pertumbuhan biomassa Chaetoceros sp. berbanding terbalik dengan peningkatan konsentrasi nitrat (Gambar 10).
32
Gambar 10. Pertumbuhan Chaetoceros sp. terhadap nitrat, nitrit dan fosfat
Pada awal kultur, konsentrasi nitrat berkisar antara 1,37 µmol/kg sampai dengan 1,51 µmol/kg. Namun pada hari kedelapan dan seterusnya berkurang menjadi 1,24 µmol/kg hingga 0,69 µmol/kg. Nitrat adalah hasil proses nitrifikasi oleh bakteri Nitrosomonas sp. yang mengoksidasi nitrit (Hendersen dan
Markland, 1987). Proses tersebut diduga akibat ketersediaan oksigen yang melimpah pada fotobioreaktor. Pada saat kultur, penurunan biomassa Chaetoceros sp. akan mengurangi ketersediaan oksigen pada fotobioreaktor, sehingga nitrat yang dihasilkan dari proses nitrifikasi juga berkurang. Menurut Mackentum (1969), konsentrasi nitrat yang optimal untuk pertumbuhan
fitoplankton pada perairan laut berkisar antara 3,9 mg/L (82,71 µmol/kg) hingga 15,5 mg/L (328,73 µmol/kg), sedangkan pada fotobioreaktor konsentrasi nitrat jauh lebih kecil dengan kisaran 0,69 µmol/kg hingga 1,51 µmol/kg.
Gambar 10 menunjukkan keterkaitan antara nitrit dengan pertumbuhan biomassa Chaetoceros sp. adalah korelasi positif dimana peningkatan konsentrasi nitrit diikuti dengan penurunan biomassa Chaetoceros sp.. Hal ini diduga akibat
nitrit yang bersifat toksik menyebabkan kematian Chaetoceros sp. sehingga biomassanya mengalami penurunan seperti yang terlihat pada hari kesepuluh hingga keduabelas. Pada awal kultur konsentrasi nitrit adalah 0,106 µmol/kg dan terus bertambah hingga mencapai 4,75 µmol/kg. Bila dibandingkan dengan perairan terbuka, menurut Effendi (2003) pada perairan alami konsentrasi nitrit sekitar 0,001 mg/L (0,02 µmol/kg), sulit menemukan nitrit dengan konsentrasi melebihi 1 mg/L (21,21 µmol/kg). Hal tersebut menunjukkan konsentrasi nitrit pada fotobioreaktor lebih tinggi dari perairan alami tetapi tidak menyebabkan kematian massal bagi fitoplankton karena kurang dari 0,5 mg/L (10,6 µmol/kg).
Fosfat memiliki hubungan korelasi negatif dengan kelimpahan biomassa Chaetoceros sp.. Hal tersebut terlihat dari penurunan konsentrasi fosfat sejak hari pertama kultur (10,67 µmol/kg) hingga pada akhir kultur menjadi 1,94 µmol/kg. Penurunan konsentrasi fosfat pada fotobioreaktor diduga akibat pemanfaatan fosfat oleh fitoplankton untuk transfer energi di dalam sel (Effendi, 2003). Hal tersebut menyebabkan konsentrasi fosfat pada fotobioreaktor lebih tinggi daripada konsentrasi nitrat dan nitrit. Kandungan fosfat yang terdapat di perairan
umumnya tidak lebih dari 1,03 µmol/kg, kecuali pada perairan yang menerima limbah dan pemupukan fosfat (Perkins, 1974). Pada fotobioreaktor konsentrasi fosfat berkisar antara 1,94 µmol/kg hingga 10,67 µmol/kg, lebih tinggi daripada perairan terbuka. Fosfat yang optimal untuk pertumbuhan fitoplankton berkisar antara 2,77 µmol/kg hingga 56,58 µmol/kg (Mackentum, 1969).
Pada kultur fotobioreaktor terlihat bahwa silikat dan Chaetoceros sp. memiliki hubungan korelasi negatif, artinya peningkatan kelimpahan Chaetoceros sp. diikuti dengan penurunan kadar silikat seperti yang terlihat pada Gambar 11.
34
Gambar 11. Pertumbuhan Chaetoceros sp. terhadap silikat
Hal ini diduga akibat pemanfaatan silikat oleh Chaetoceros sp. untuk
pembentukan dinding sel. Konsentrasi silikat mengalami penurunan dari 268,50 µmol/kg hingga 98,98 µmol/kg seperti yang terlihat pada Gambar 11. Pada perairan terbuka, konsentrasi silikat tertinggi sebesar 17,8 mg/L (289,43 µmol/kg) terdapat di sungai dan terendah 0,128 mg/L (2,08 µmol/kg) di perairan Okinawa (Mkadam et al., 2005).
Menurut Kamatani dan Takano (1984), konsentrasi silikat air laut Teluk Tokyo adalah 0,280 mg/L (4,55 µmol/kg). Bila dibandingkan dengan konsentrasi silikat air laut, fotobioreaktor memiliki konsentrasi silikat yang cukup tinggi. Hal ini diduga akibat pemupukan silikat pada awal kultur serta dipengaruhi oleh salinitas dengan kisaran antara 25 - 27‰. Selain itu juga diduga akibat perbedaan kepadatan diatom (Chaetoceros sp.) pada fotobioreaktor lebih tinggi karena volumenya yang terbatas. Konsentrasi nitrogen (penjumlahan antara konsentrasi nitrat dan nitrit) pada fotobioreaktor lebih rendah bila dibandingkan dengan
konsentrasi fosfat dan silikat sehingga nitrogen merupakan faktor pembatas pada pertumbuhan Chaetoceros sp..
4.4 Pertumbuhan Chaetoceros sp. dan sistem karbonat dalam fotobioreaktor Karbon sangat penting keberadaannya bagi Chaetoceros sp. sebagai pembentuk asam amino (nutrisi) yang berperan di dalam metabolisme. Karbon anorganik terlarut (DIC) adalah total karbon terlarut yang diperoleh dari
penjumlahan senyawa karbon seperti CO2, CO3, HCO3 dan H2CO3. Oleh karena
itu perubahan konsentrasi senyawa karbon sangat berpengaruh terhadap konsentrasi DIC. Semakin tinggi penyerapan CO2 oleh Chaetoceros sp. maka
akan menyebabkan penurunan konsentrasi DIC di dalam fotobioreaktor. Selama kultur Chaetoceros sp. pada fotobioreaktor, pengukuran karbon anorganik terlarut dan karbon organik partikulat hanya dilakukan tiga kali (Lampiran 3). Karbon anorganik terlarut (DIC) adalah karbon yang belum dimanfaatkan oleh fitoplankton. Hasil pemanfaatan karbon anorganik oleh fitoplankton akan meningkatkan biomassa Chaetoceros sp. dan menghasilkan karbon organik partikulat (POC). Pada penelitian ini hubungan antara
pertumbuhan Chaetoceros sp. dengan DIC dan POC terlihat pada Gambar 12. Peningkatan konsentrasi POC dari 0 µmol/kg hingga mencapai 1.550 µmol/kg diikuti dengan peningkatan biomassa Chaetoceros sp. serta penurunan kadar DIC dari 918 µmol/kg menjadi 27,36 µmol/kg. Hal ini menunjukkan bahwa karbon anorganik dimanfaatkan oleh Chaetoceros sp. untuk pertumbuhannya.
Selanjutnya, peningkatan kelimpahan Chaetoceros sp. akan menghasilkan POC melalui proses mortalitas.
36
Gambar 12. Pertumbuhan Chaetoceros sp. terhadap POC dan DIC
Hubungan antara DIC dengan POC adalah salah satu proses
kesetimbangan karbon yang tidak hanya terjadi di alam tetapi juga di dalam fotobioreaktor, terbukti dengan peningkatan konsentrasi salah satu parameter akan berdampak sebaliknya (penurunan) pada parameter yang lain sehingga total karbon tetap terjaga. Hasil pengukuran karbon pada penelitian ini sesuai dengan penelitian yang dilakukan oleh Clark dan Flynn (2000) mengenai penyerapan karbon oleh Thalassiosira pseudonana yang menunjukkan terjadinya peningkatan karbon organik (POC) seperti yang terlihat pada Gambar 13.
Jadi, pada sistem karbonat tertutup ini (fotobioreaktor), proses injeksi gas CO2 yang masuk ke dalam fotobioreaktor akan terlarut menjadi karbon anorganik
(DIC) kemudian diserap oleh Chaetoceros sp., dan menghasilkan karbon organik partikulat (POC). Mengingat bahwa pada hari ke-10 terjadi pertumbuhan optimal biomassa Chaetoceros sp., maka aliran karbon dalam sistem tertutup ini menjadi fokus pembahasan selanjutnya.
Sumber : Clark dan Flynn (2000)
Gambar 13. Hasil pengukuran karbon pada kultur Thalassiosira pseudonana
Laju injeksi sebesar 0,5 liter per menit dari penampungan gas yang berkapasitas 118 liter, artinya dalam satu hari mengalir 720 liter gas ke dalam fotobioreaktor. Gas tersebut adalah campuran antara gas CO2 (12%) dan N2
(88%). Injeksi gas CO2 pada hari ke-10 sebesar 3% vol., sedangkan yang keluar
(hari ke-11) 1,65% vol., sehingga gas CO2 yang diserap adalah sebesar 1,35% vol.
atau 9,72 liter total yang dimanfaatkan oleh Chaetoceros sp. pada hari ke-10. Selanjutnya 9,72 liter CO2 tersebut terlarut menjadi DIC yang direpresentasikan
oleh konsentrasi DIC sebesar 0,33 mgC/L. Pada waktu yang hampir bersamaan, Chaetoceros sp. melakukan fotosintetis untuk pertumbuhannya, dimana terjadi penambahan 960.000 sel/ml dari hari ke-9. Produksi sampingan aktivitas proses biologis (mortalitas) adalah POC, dimana nilainya sebesar 18,97 mgC/L pada hari ke-10. Jadi, dalam sistem tertutup ini DIC cenderung turun dan diikuti oleh peningkatan POC sejak hari ke-0. Hal ini memungkinkan akibat proses dekomposisi belum sempat terjadi dalam siklus 14 hari ini.
38
5. KESIMPULAN DAN SARAN
5.1 Kesimpulan
Kondisi optimal kultur skala laboratorium Chaetoceros sp. pada
fotobioreaktor terjadi saat suhu berkisar antara 22,39 - 24,67oC, salinitas 26,66 - 27,65‰, pH 6,27 - 6,68, dan DO 1 - 1,52 ppm. Selama satu siklus fotobioreaktor (14 hari) Chaetoceros sp. mampu menyerap 10,56% vol. CO2 yang diinjeksi ke
dalam fotobioreaktor selama kultur. Laju penyerapan rata-ratanya 0,56% vol. CO2 per hari dari masukan awal konsentrasi gas CO2 sebesar 7,15% vol. Pada
kondisi pertumbuhan Chaetoceros sp. mencapai optimal (hari ke-10), penyerapan gas CO2 terlarut mencapai 1,35% vol.. Nitrogen (nitrat dan nitrit) menjadi faktor
pembatas pada pertumbuhan Chaetoceros sp. karena memiliki jumlah minimum bila dibandingkan dengan fosfat dan silikat. Dalam sistem karbonat di
fotobioreaktor, produksi sampingannya adalah POC sebesar 18,97 mgC/L.
5.2 Saran
Sistem karbonat pada fotobioreaktor diawali dengan injeksi gas CO2,
kemudian terlarut dalam bentuk karbon anorganik (DIC) dan dimanfaatkan oleh Chaetoceros sp.. Proses mortalitas Chaetoceros sp. akan menghasilkan karbon organik partikulat (POC). Pada proses selanjutnya, POC dapat terdekomposisi kembali menjadi DIC. Oleh karena itu, untuk memutus proses ini maka sebaiknya pada saat pemanenan (setelah hari ke-14), POC dikubur ke dalam tanah.
39
Alaerts, G. dan S, Sumestri. 1987. Metode Penelitian Air. Usaha Nasional. Surabaya. 309 h.
American Public Health Association (APHA). 1989. Standard method for
examination of waste water. 12th Edition. APHA AWWA (American
Water Work Association) and WPCF (Water Pollution Control Federation). New York. 453 h.
Ariyati, R. W., L. Sya’rani, E. Arini. 2007. Analisis Kesesuaian Perairan Pulau Karimunjawa dan Pulau Kemujan sebagai Lahan Budidaya Rumput Laut. Jurnal Pasir Laut. 3(1): 27-45.
Aunurohim, D. Saptarini, D. Yanthi. 2009. Fitoplankton Penyebab Harmful Algae Blooms (HABs) di Perairan Sidoarjo. Jurnal. Biologi Fakultas Matematika dan Ilmu Pengetahuan Alam. Institut Teknologi Sepuluh November. Surabaya. 1937: 1-7.
Badan Standardisasi Nasional (BSN). 2000. Air demineral. Balai Besar Litbang Industri Hasil Pertanian. Bogor. SNI (01-6241-2000): 5-6.
Beardall, J. and S. Stojkovic. 2006. Microalgae under Global Environmental Change: Implications for Growth and Productivity, Populations and Trophic Flow. Scienceasia. 32(1): 1-10.
Boyd, C. E. 1982. Water Quality in Warm Water Fish Pond. Auburn University Agricultural Experimenta Satation. Auburn Alabama. 359 h.
Castro, P. dan M. E. Huber. 2007. Marine Biology. New York: MacGraw-Hill Higher Education. New York. 460 h.
Clark, D. R. and K. J. Flynn. 2000. The Relationship Between the Dissolved Inorganic Carbon Concentration and Growth Rate in Marine
Phytoplankton. The Royal Society, London, Proceedings B(267): 953-959.
Effendi, H. 2003. Telaah Kualitas Air. Penerbit Kanisius. Yogyakarta. 257 h. Feely, R. A., C. L. Sabine, T. Takahashi, R. Wanninkhof. 2001. Uptake and
Storage of Carbon Dioxide in the Ocean: The Global CO2 Survey.
Oceanography, 14(4): 18-32.
Hendersend, S. B. and H. R. Markland. 1987. Decaying Lakes, The Origin and Control of Cultural Eutrophication. John wiley & Sons. New York. 264 h.
40
Hutagalung, H. P., D. Setiapermana, S. H. Riyono. 1997. Metode Analisis Air Laut, Sedimen dan Biota. Buku 2. Pusat Penelitian dan Pengembangan Oseanologi Lembaga Ilmu Pengetahuan Indonesia. Jakarta. 181 h. Izzati, M. 2008. Perubahan Konsentrasi Oksigen Terlarut dan pH Perairan
Tambak setelah Penambahan Rumput Laut Sargassum plagyophyllum dan Ekstraknya. Buletin Anatomi dan Fisiologi. 16(2): 60-69.
Jinming, S. 2010. Biogeochemical Processes of Biogenic Elements in China Marginal Seas. Zhejiang University Press, Hangzhou and Springer-Verlag Berlin Heidelberg. Germany. 662 h.
Johnson, W. S. and D. M. Allen. 2005. Zooplankton of the Atlantic and Gulf Coasts, A Guide to Their Identification and Ecology. The John Hopkins University Press. Baltimore and London. 379 h.
Kamatani, A. and M. Takano. 1984. The Behavior of Dissolved Silica During the mixing of river and seawaters in Tokyo Bay. Estuarine, Coastal and Shelf Science. 19: 505-512.
Kawaroe, M., T. Prartono, A. Sunuddin, D. W. Sari, D. Augustine. 2010. Mikroalga, Potensi dan Pemanfaatannya untuk Produksi BioBahan Bakar. IPB Press. Bogor. 150 h.
Kennish, M. J. 1990. Ecology of Estuaries: Biological Aspects. CRC Press. Boston. 391 h.
Malone, R. F. and D. G. Burden. 1988. Design of Recilculating Blue Crab Shedding System. Louisiana Sea Grant College Program. Louisiana State University. Louisiana. 76 h.
Mata, T. M., A. A. Martins, N. S. Caetano. 2010. Microalgae for biodiesel production and other applications: A Review. Renewable and Sustainable Energy Reviews. 14: 217-232.
Mackentum, K. M. 1969. The Practice of Water Pollution Biology. United States Department of Interior. Federal Water Pollution Control Administration. Division of Technical Support. 411 h.
Mkadam, K. M., T. Yonaha, V. S. Ali and A. Tokuyama. 2005. Dissolved
Aluminum and Silica Release on the Interaction of Okinawan Subtropical Red Soil and Seawater at Different Salinities: Experimental and Field Observations. Geochemical Journal. 40: 333-343.
Munandar, H. 2009. Sistem Karbonat di Laut serta Peranannya bagi Biota Calsifier. Balai Konservasi Biota Laut, Pusat Penelitian Oseanografi Lembaga Ilmu Pengetahuan Indonesia. Jurnal Oseana. 34(1): 35-46.
Nontji, A. 1987. Laut Nusantara. Cetakan II. Djambatan. Jakarta. 459 h. Perkins, E. J. 1974. The Biology of Estuaries and Coastal Water. Academy
Press Co. New York. 678 h.
Sanusi, H. S. 2006. Kimia Laut. Proses Fisik Kimia dan Interaksinya dengan Lingkungan. Departemen Ilmu dan Teknologi Kelautan. Fakultas Perikanan dan Ilmu Kelautan. Institut Pertanian Bogor. Bogor. 188 h. Setiawan, A., Kardono, R. A. Darmawan, A. D. Santoso, A. H. Stani, Prasetyadi,
L. Panggabean, D. Radini, S. Sapulete. 2008. Teknologi Penyerapan Karbondioksida dengan Kultur Fitoplankton pada Fotobioreaktor. In Pertemuan Ilmiah Tahunan V Ikatan Sarjana Oseanologi Indonesia, 11 November 2008, Institut Teknologi Bandung. Bandung. 7 h.
Suriadikarta, D. A. dan M. T. Sutriadi. 2007. Jenis-jenis Lahan Berpotensi untuk Pengembangan Pertanian di Lahan Rawa. Balai Penelitian Tanah. Jurnal Litbang Pertanian. 26(3): 115-122.
Susandi, A., I. Herlianti, M. Tamamadin, I. Nurlelea. 2008. Dampak perubahan iklim terhadap ketinggian muka laut di wilayah Banjarmasin. Jurnal Ekonomi Lingkungan. 12(2): 1-2.
Svobodova Z., R. Lloyd, J. Machova, B. Vykusova. 1993. Water Quality and Fish Health. EIPAC Technical Paper No. 54, FAO. Roma. 59 h. Wetzel, R. G. 1983. Limnology. Second Edition. Saunders College Publishing,
Toronto. 859 h.
Yuliana. 2002. Hubungan antara kandungan nutrien dan intensitas cahaya dengan produktivitas primer fitoplankton di Perairan Teluk Lampung. [Tesis]. Bogor: Institut Pertanian Bogor. Bogor. 83 h.
Yulianto, K. 1989. Pengaruh Penurunan Salinitas Terhadap Laju Fotosintesis Alga Hijau (Caulerpa serrulata forsk) J. Agardh dan Valonia
Aegagropila C. Agardh. Jurnal. Balai Penelitian dan Pengembangan Sumberdaya Laut. Pusat Penelitian dan Pengembangan Oseanologi. Lembaga Ilmu Pengetahuan Indonesia. Ambon. ISBN(979-8093-04-6): 39-44.
42
Lampiran 1. Hasil pengukuran biomassa dan CO2.
Hari ke Hasil
Biomassa (sel/ml) Gas CO2 (%vol)
0 10.33 7,15 1 20.66 8,17 2 49.067,5 7,83 3 100.71 7,3 4 219 6,87 5 265.8 6,4 6 369.8 5,57 7 448 4,83 8 800.6 4,15 9 1.520.000 3,52 10 2.480.000 3 11 1.890.000 1,65 12 1.760.000 1,63 13 1.830.000 1,44
Lampiran 2. Hasil pengukuran nutrien
Hari ke-
NO3-N NO2-N PO4-P Silikat Klorofil
mg/L µmol/kg mg/L µmol/kg mg/L µmol/kg mg/L µmol/kg µg/L µmol/kg 0 0,087 1,37 0,005 0,12 1,035 10,68 16,437 268,50 11,055 0,04 2 0,093 1,47 0,009 0,19 0,908 9,37 16,25 265,44 18,789 0,07 4 0,096 1,52 0,046 0,98 0,857 8,84 14,467 236,39 79,04 0,31 6 0,091 1,44 0,052 1,12 0,799 8,24 12,428 203,06 90,307 0,35 8 0,079 1,25 0,103 2,19 0,55 5,67 9,583 156,47 189,825 0,75 10 0,044 0,69 0,223 4,75 0,188 1,94 6,06 98,98 330,49 1,30
Lampiran 3. Hasil pengukuran karbon anorganik terlarut (DIC) dan karbon organik partikulat (POC).
Hari ke- DIC (mgC/L) POC (mgC/L) DIC (µmol/kg) POC (µmol/kg)
0 11,24 0 918,02 0
4 2,86 7,66 233,50 625,49 10 0,33 18,97 27,36 1.549,62