Microfungal communities in soil, litter and casts of Lumbricus terrestris
L. (Lumbricidae): a laboratory experiment
Alexei V. Tiunov
a,∗, Stefan Scheu
baLaboratory of Soil Zoology, Institute of Ecology and Evolution, Leninsky Prospect 33, 117071, Moscow, Russia bInstitute of Zoology, Darmstadt University of Technology, Schnittspahnstrasse 3, D-64287, Darmstadt, Germany
Received 8 July 1999; received in revised form 29 November 1999; accepted 1 December 1999
Abstract
The anecic earthworm Lumbricus terrestris L. was kept in laboratory microcosms containing beech forest soil without litter, with beech leaf litter or with lime leaf litter. The structure of microfungal communities in soil, litter and fresh and aged (100 days) earthworm faeces was analysed using the washing and plating technique. The passage of mineral soil through the gut of L. terrestris affected the structure of the fungal community only little. In contrast, in the litter treatments the structure of the fungal community in fresh earthworm casts significantly differed from that in soil and litter. The majority of soil and litter inhabiting fungi survived passage through the gut of L. terrestris and the fungal community in casts consisted of a mixture of soil and litter inhabiting fungi. However, the frequency of Cladosporium spp., Alternaria spp., Absidia spp., and other taxa was strongly reduced in fresh casts. The degree of colonization of litter particles (number of isolates per number of plated particles) also decreased, but some fungi (mainly Trichoderma spp.) benefited from gut passage and flourished in fresh casts. During ageing of cast material the dominance structure of the fungal community changed. Both the degree of colonization of organic particles and the species diversity increased and approached that in soil. However, the structure of the fungal community in casts remained cast specific even after 100 days of incubation. It is concluded that the feeding and burrowing activity of L. terrestris accelerates the colonization of litter by the edaphic mycoflora but also extends the range of occurrence of litter-associated fungi into mineral soil layers. ©2000 Elsevier Science B.V. All rights reserved.
Keywords: Soil fungi; Fungal community; Anecic earthworms; Lumbricus terrestris; Litter decomposition
1. Introduction
Fungi and earthworms are important members of soil communities and their interactions are assumed to significantly affect microbial-mediated processes in soil. Although the importance of these interactions is widely accepted, they are little studied and the in-formation available is in part contradictory (Brown, 1995). Earthworms may influence soil fungi by a
∗Corresponding author. Tel.:+7-095-958-1449;
fax:+7-095-954-5534/952-2592.
E-mail address: [email protected] (A.V. Tiunov).
variety of mechanisms, including the alteration of the physical and chemical state of the environment (‘en-gineering’ sensu Jones et al., 1997), comminution and translocation of litter, dispersal of fungal propagules and grazing on fungal tissue (Visser, 1985). Various authors suggested that fungi are a major component of the diet of earthworms (Dash et al., 1984, 1986; Ed-wards and Fletcher, 1988; Tiwari et al., 1990; Doube and Brown, 1998). Earthworms have been shown to prefer food substrates colonized by certain fungal species and fungi have been found to be damaged by gut passage (Cooke, 1983; Moody et al., 1995, 1996; Marfenina and Ishchenko, 1997).
Even low levels of selective grazing by invertebrates has been shown to significantly alter the distribution and succession of fungi on litter (Parkinson et al., 1979; Newell, 1984). Therefore, it is likely that earth-worms alter the composition of fungal communities in soil and the fungal succession in decomposing lit-ter. However, there is only fragmentary information on effects of earthworms on the structure of fungal com-munities in soil. Domsch and Banse (1972) reported a strong alteration of the fungal dominance structure in casts of Lumbricus terrestris in comparison to the food substrates. In contrast, Tiwari and Mishra (1993) did not find significant differences in the structure of fun-gal communities in soil and field-collected earthworm casts. In micro- and mesocosm experiments, the pres-ence of the epigeic earthworm species Dendrobaena octaedra (McLean and Parkinson, 1998) and the endo-geic species Octolasion tyrtaeum (Scheu and Parkin-son, 1994) affected the structure of fungal communi-ties only little, although the frequency of some species changed significantly. Detailed studies aiming at sep-arating different mechanisms which may contribute to earthworm-mediated changes in the structure of fungal communities in soil are lacking.
We studied microfungal communities in fresh and aged faeces of the widespread anecic earthworm L. terrestris L. This large deep-burrowing species builds permanent vertical burrows, but feeds mainly on or-ganic materials on the soil surface. Due to the feeding and burrowing activity of L. terrestris, large amounts of mineral soil are translocated upward to the soil sur-face and mixed with litter, whereas litter material is transferred deep into the mineral soil. We addressed the following questions: how does passage through the gut of L. terrestris affect the structure of the fungal community as compared to soil and litter? Which of the soil and litter inhabiting fungal species survive, and which are damaged by gut passage? How does the fungal community in faeces change during ageing of cast material?
2. Materials and methods
2.1. Experimental design
Mineral soil, beech leaf litter and specimens of L. terrestris were obtained from a 115–120 year old
beech (Fagus sylvatica) stand, the ‘Göttinger Wald’ (Lower Saxony, Germany). Soil material was taken in June 1996 at a depth of 3–10 cm from the
min-eral soil surface and sieved (<4 mm). Overwintered
F. sylvatica leaf litter was collected from the soil surface at the same time. Lime (Tilia cordata) leaf litter was collected in May 1996 in a lime grove 30 km south of Moscow (Russia) and kept until the start of the experiment in polyethylene bags at 5◦C.
The experiment was carried out in narrow containers (inner dimensions: 650 mm high, 310 mm long, 10 mm wide). Three combinations of beech forest soil and litter were studied: soil without litter (B0 treatment),
soil+beech litter (BB treatment), and soil+lime litter
(BL treatment).
Containers were filled with soil to a level of 500 mm. The soil was compacted to a bulk density
of 0.65 kg dry weight 1000 cm−3 which is typical
for the upper soil layers at the Göttinger Wald. The moisture content of the soil was kept at the field level (62% of dry wt.). About 25 g of Tilia or Fa-gus litter was added to each BL and BB container and replenished periodically as it was consumed by earthworms. In each microcosm two adult L. ter-restris (average individual weight 6.0±0.2 g) were
placed. Containers were kept at 15◦C in
perma-nent darkness. Five replicates were set up per treat-ment. Further details are given in Tiunov and Scheu (2000).
After a preliminary period of 2 weeks, when a bur-row system was established by the earthworms, con-tainers were opened every second day and freshly de-posited faeces were collected (average age of 1 day). For the analysis of the microfungal community, fresh casts and control litter samples were collected 26–30 days after the start of the experiment from four con-tainers of each treatment. Simultaneously, control soil samples were taken from B0 (two replicates), BL and BB treatments (one replicate each). Soil and litter sam-ples were taken from the zones not affected by L. ter-restris (at least 50 mm away from the nearest burrow). Faecal material of age 100 days was obtained by incubating casts between two layers of soil separated by 0.5 mm gauze. The container with soil and casts
was incubated in a polyethylene bag at 15◦C. A plastic
2.2. Isolation of fungi
The composition of the fungal communities in the soil, litter and casts was analysed by a modified washing and plating method (Parkinson and Williams, 1961). One gram (fresh weight) of soil, cast or litter material was blended for 20 s in 100 ml sterile wa-ter. About 0.5 g of the blended material was placed in the washing apparatus (vol. ca. 100 ml) between 0.5 and 0.2 mm meshes. The material was washed
by applying a constant flow (0.5 l min−1) of sterile
water for 10 min. During the washing procedure the apparatus was gently shaken. Washed organic parti-cles of size 0.2–0.5 mm were collected on sterile filter paper. Fifty particles per sample were plated on malt extract agar containing streptomycin and tetracycline
(0.1 and 0.05 g l−1, respectively) to suppress growth
of bacteria. One particle was placed per Petri-dish. The Petri-dishes were incubated at room temperature. When two or more fungal species were growing from the single particle, pure cultures were prepared by transferring hyphal inocula to separate plates. The frequency of occurrence of fungal taxa (as percent-ages) was calculated as the number of particles with
fungal growth per total number of isolates×100. On
average, 44.5 isolates were obtained per 50 plated particles.
2.3. Statistical analysis
The main goal of the analysis was to compare the structure of whole fungal communities in the studied substrata, rather than to follow changes in abundance of single fungal species or genera. For this purpose, ordinations and discriminant analysis were used. Rare fungi (fewer than two isolates), yeast-like forms and loosely-defined groups (i.e., ‘other sterile dark’, ‘other Trichoderma’; see further) were not included in the analysis. The remaining 46 taxa represented ca. 70% of the isolates.
The logic of the mathematical analysis follows the scheme proposed by Puzachenko and Kuznetsov (1998). Based on the relative frequency of fungal taxa, the square matrix of nonparametric Gamma
correlation (analogous to Kendall τ) between the
studied samples was calculated. The distance matrix was obtained by subtracting correlation coefficients
(range 1 to−1) from 1. Thus, distances varied from
0 (perfect similarity between two samples) to 2 (no similarity). The distance matrix was analysed further by nonmetric multidimensional scaling. This ordina-tion technique ‘rearranges’ objects in n-dimensional space, so as to arrive at a configuration that best ap-proximates the observed distances. The number of meaningful dimensions was evaluated by comparing actual stress values with the theoretical exponential function of stress.
The coordinates of the samples in n-dimensional space were used for discriminant function analysis (DFA), with ‘substrate’ (soil, litter or casts) as a group-ing variable. Squared Mahalanobis distances between group centroids and the reliability of sample classifica-tion were determined. Typically, only two significant discriminant functions (canonical roots) were derived. Thus, the results of DFA are graphically presented in 2-dimensional space. For the interpretation of the dis-criminant axes in respect to the frequency of fungal taxa, linear correlations were calculated between the discriminant function scores for each sample and the relative dominance of fungal taxa. A similar procedure was used to analyse the similarity matrix between fun-gal taxa. Prior to the analysis, fungi were grouped into soil- and litter-associated species using cluster analy-sis and Ward’s minimum variance method.
All calculations were made using the STATISTICA software package. Differences between means were tested using Tukey’s honestly significant difference (HSD) at the 0.05 probability level.
3. Results
3.1. ‘Soil’ and ‘litter’ fungi
Table 1
Frequency (percentage of the total number of isolates) of dominant fungi in soil, litter and fresh casts of L. terrestrisa
Soil B0 casts Beech litter BB casts Lime litter BL casts
‘Beech litter species’
Mortierella gamsii Milko 1.6 ab 0 a 14.8 b 6.7 b 0.5 a 0 a
Trichoderma pseudokoningii Rifai 0 a 0 a 6.9 b 13.2 b 0 a 0 a
Sterile hyaline sp. 1 0 a 0 a 3.3 b 0.7 ab 0 a 0 a
Cladosporium spp. 0 0 2.6 0 0 0
Sterile dark sp. 1 0 0 1.1 1.4 0 0
‘Lime litter species’
Trichoderma koningii Oudem. 0 a 0 a 0 a 0 a 29.3 b 39.4 b
Trichoderma hamatum Bain. 0.9 a 1.4 a 4.4 a 2.2 a 9.5 ab 14.5 b
Mucor hiemalis Wehmer 0 0 5.8 3.0 5.9 3.5
Alternaria sp. 2 0 a 0.6 a 0.5 a 0 a 5.1 b 0.6 a
Alternaria alternata (Fr.) Keissler 0 0 0 0 2.4 0
Tolypocladium sp. 0 0 0 0 1.6 1.8
‘Soil species’
Acremonium spp. 11.2 a 9.0 ab 1.7 ab 5.6 ab 0.5 b 3.9 ab
Humicola sp. 1 5.7 a 5.7 a 0 b 2.0 ab 0 b 1.8 ab
Absidia cylindrospora Hagem 5.5 a 2.1 ab 0.5 b 1.4 ab 0 b 0 b
Sterile dark sp. 2 3.9 ab 6.4 a 0.7 b 4.3 ab 0 b 1.7 ab
Cylindrocarpon destructans (Zinssm.) Scholten 4.4 4.3 0 0.5 0 0.6 Mortierella minutissima van Tiegh. 3.8 a 2.6 a 0 b 3.4 a 0 b 0 b
Penicillium spp. 3.6 4.6 1.2 2.9 0.5 0.6
Volutella sp. 3.1 4.4 1.6 5.7 0 0.6
Sterile dark sp. 3 2.8 a 0 b 0.6 ab 1.2 ab 0 b 0 b
Sterile dark sp. 4 2.6 ab 3.4 b 0 c 0.7 ac 0 c 0 c
Chrysosporium sp. 1 2.6 a 0 b 0 b 0 b 0 b 0 b
Phialophora sp. 1 2.1 a 0 b 0 b 0 b 0 b 0 b
Sterile hyaline sp. 2 2.0 a 0 b 0 b 0.7 ab 0 b 0 b
Sterile hyaline sp. 3 1.9 2.0 0 1.2 0 0
Paecilomyces spp. 1.8 1.4 1.1 0 0 0
Sterile hyaline sp. 4 1.5 2.0 1.8 1.2 0 0.6
Sporothrix sp. 1.5 1.3 0 0.8 0 0.6
Chloridium sp. 1.5 0.6 0 0.7 0 0
Trichoderma polysporum (Link ex Pers.) Rifai 0.9 0.6 1.2 0 0 0
Sterile hyaline sp. 5 0 0.6 0 0 0 1.2
Chrysosporium sp. 2 0 0 0 1.5 0 0
Trichocladium sp. 0 1.4 0 0 0 0
Total number of taxa isolated 37 29 26 32 18 24
Average number of taxa per sample 22.5 a 19.0 a 14.0 bc 16.8 ab 11.5 c 11.5 c
aThe casts originated from B0 (soil only), BB (soil+beech litter) and BL (soil+lime litter) treatments. Values within rows followed
by different letters are significantly different (Tukey’s HSD test based on arcsine-transformed data, n=4, p<0.05).
Cluster analysis of the similarity matrix of 38 dominant fungal taxa singled out three main clusters, which were interpreted as ‘beech litter species’, ‘lime litter species’ and ‘soil species’. The reliability of the separation was further
con-firmed by the discriminant analysis (p<0.001).
Fungal taxa in Table 1 are grouped according to this clustering. However, the arbitrary character
of the grouping should be noted, as some ‘soil fungi’ (e.g. T. polysporum or Sterile hyaline sp. 4) were equally abundant in the beech litter, while M. hiemalis had similar occurrence in both types of litter.
and Chrysosporium sp. 2 were isolated only from the casts, but the frequency of these fungi was low and they did not reliably separate from the ‘soil fungi’.
3.2. Fresh casts
The structure of the fungal community in food sub-strates and egested materials can only be compared directly in the B0 treatment, where earthworms ex-clusively consumed mineral soil. Only a few fungal taxa were significantly affected by gut passage in this treatment (Table 1). However, Mucorales (Absidia spp. and Mortierella spp.) tended to be less frequent and other fungal species, e.g. Chrysosporium sp. 1 and Phialophora sp., were not isolated from casts. The total number of fungal taxa was lower in casts (29) than in soil (37).
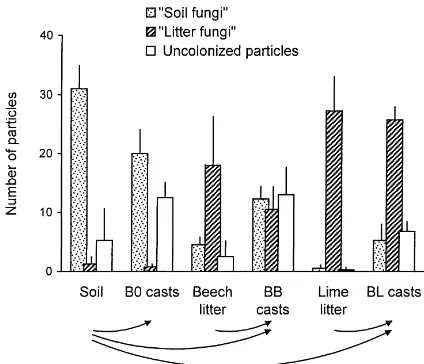
Comparing food substrate and casts is more diffi-cult in BB and BL treatments because L. terrestris consumed both soil and litter. Alterations in the struc-ture of fungal communities in casts of these treat-ments might be due to gut passage or to mixing of soil and litter materials. Generally, ‘soil fungi’ in casts ac-counted for 35 and 13% of the total number of isolates in BB and BL treatments, respectively (Fig. 1). Of
Fig. 1. Number of organic particles colonized by litter-associated fungi (‘litter fungi’), soil-associated fungi (‘soil fungi’) and un-colonized particles in soil, litter and fresh casts of Lumbricus terrestris (means+SD). The casts originated from B0 (soil only), BB (soil+beech litter) and BL (soil+lime litter) treatments. 50 particles were plated per replicate, n=4.
Fig. 2. Discriminant analysis plane of soil, litter and fresh casts of Lumbricus terrestris according to the fungal dominance structure. The casts originated from B0 (soil only), BB (soil+beech litter) and BL (soil+lime litter) treatments. Ellipses represent 95% confidence limits.
the ‘litter fungi’, Mucorales (Mortierella gamsii and M. hiemalis), Cladosporium spp., and two species of Alternaria tended to be less abundant in casts than in the litter offered as food substrate. In contrast, the fre-quency of three Trichoderma spp. was greater in casts (Table 1). However, most of these tendencies were not statistically significant.
In each of the three treatments the number of or-ganic particles from which no fungi developed was significantly greater in L. terrestris casts compared to food materials. The mean number of uncolonized par-ticles was 10% in soil, 5% in beech litter and 0.5% in lime litter, but reached 25, 26 and 14% in B0, BB and BL casts, respectively (Fig. 1).
Table 2
Squared Mahalanobis distances between group centroids and re-liability of discrimination based on data on fungal dominance structurea
B0 casts BB casts BL casts
Soil 5.3 nsb 73.8∗∗∗ 102.5∗∗∗
Beech litter 180.9∗∗∗ 19.2∗ 130.3∗∗∗
Lime litter 186.9∗∗∗ 114.7∗∗∗ 26.4∗∗
B0 casts 0 89.4∗∗∗ 86.7∗∗∗
BB casts 89.4∗∗∗ 0 100.8∗∗∗
aSee text for details. bns=not significant. ∗p<0.01.
∗∗p<0.005. ∗∗∗p<0.0001.
distinguished from each other and from both soil
and litter (p<0.01). In contrast, casts from the B0
treatment could not be discriminated from the soil (Table 2).
The two discriminant axes can be interpreted eas-ily considering the dominance structure of the fungal communities. The occurrence of dominant ‘soil fungi’ positively correlated with the first axis, but not with the second axis. ‘Beech litter fungi’ were negatively cor-related with both axes. ‘Lime litter fungi’ were nega-tively correlated with the first axis and posinega-tively with the second (Table 3).
3.3. Aged casts
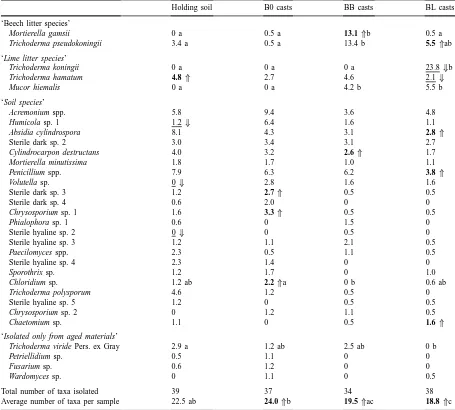
The frequency of certain fungal taxa significantly changed during the 100-day incubation of soil and cast material (Table 4). However, in most cases the changes were not uniform in the different substrates. The frequency of T. koningii, Volutella sp. and Humi-cola sp. 1 tended to decline during ageing of soil and casts. In contrast to the fresh samples, Sterile hyaline sp. 1, Sterile dark sp. 1, Tolypocladium sp. and Tri-chocladium sp. were not isolated from the aged sam-ples (Tables 1 and 4). Conversely, the frequency of A. cylindrospora, T. pseudokoningii, T. polysporum and Penicillium spp. increased in most aged substrates. T. viride, Fusarium sp., Wardomyces sp. and other fungi were isolated only from aged soil and casts. The total number of isolated taxa also increased, and the number of uncolonized particles in aged casts did not exceed 10%.
Table 3
Linear correlation (r-values) between the frequency of fungal taxa and discriminant axesa
Axis 1 Axis 2
‘Lime litter species’
Trichoderma hamatum −0.517∗∗ 0.599∗∗
Mucor hiemalis −0.515∗∗ 0.010 nsb
Alternaria sp. 2 −0.482∗ 0.435∗
Trichoderma koningii −0.472∗ 0.727∗∗∗
Tolypocladium sp. −0.221 ns 0.507∗
‘Beech litter species’
Mortierella gamsii −0.323 ns −0.646∗∗
Trichoderma pseudokoningii −0.171 ns −0.630∗∗
Sterile hyaline sp. 1 −0.313 ns −0.511∗
Sterile dark sp. 1 −0.180 ns −0.434∗
‘Soil species’
Humicola sp. 1 0.757∗∗∗ 0.077 ns Sterile dark sp. 2 0.677∗∗∗ −0.125 ns Mortierella minutissima 0.657∗∗∗ −0.264 ns
Sterile dark sp. 4 0.652∗∗ −0.014 ns
Acremonium spp. 0.636∗∗ −0.074 ns
Cylindrocarpon destructans 0.626∗∗ 0.086 ns
Absidia cylindrospora 0.613∗∗ −0.126 ns
Penicillium spp. 0.585∗∗ −0.183 ns
Sterile hyaline sp. 3 0.517∗∗ −0.094 ns
Sporothrix sp. 0.500∗ 0.074 ns
Sterile hyaline sp. 2 0.480∗ −0.134 ns
Chloridium sp. 0.480∗ −0.098 ns
Phialophora sp. 1 0.480∗ −0.019 ns
Chrysosporium sp. 1 0.448∗ −0.008 ns
Paecilomyces spp. 0.410∗ −0.138 ns aSee text for details.
bns=not significant. ∗p<0.05.
∗∗p<0.01. ∗∗∗p<0.001.
The changes in the fungal community structure in aged materials were reflected by reliable discrimina-tion between ‘fresh’ and ‘aged’ substrates. The dis-crimination was strongest between ‘fresh’ and ‘aged’
soil (p=0.003) and between ‘fresh’ and ‘aged’ casts
from BL treatment (p=0.007) whereas it was less
re-liable between ‘fresh’ and ‘aged’ casts in B0 and BB
treatments (p=0.037 and 0.054, respectively).
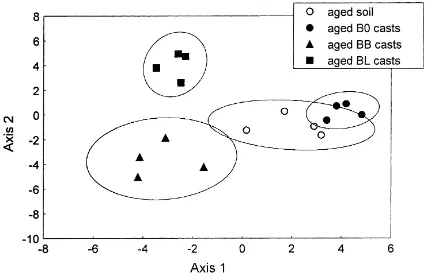
During incubation, cast materials of different treat-ments were kept in the same soil container, separated only by 2–5 cm. However, after 100 days of ageing, fungal communities in L. terrestris casts retained spe-cific features (Fig. 3). The three types of cast material were separated from each other in 2-dimensional
Table 4
Frequency (percentage of the total number of isolates) of dominant fungi in aged soil and casts of Lumbricus terrestrisa
Holding soil B0 casts BB casts BL casts
‘Beech litter species’
Mortierella gamsii 0 a 0.5 a 13.1⇑b 0.5 a
Trichoderma pseudokoningii 3.4 a 0.5 a 13.4 b 5.5⇑ab
‘Lime litter species’
Trichoderma koningii 0 a 0 a 0 a 23.8⇓b
Trichoderma hamatum 4.8⇑ 2.7 4.6 2.1⇓
Mucor hiemalis 0 a 0 a 4.2 b 5.5 b
‘Soil species’
Acremonium spp. 5.8 9.4 3.6 4.8
Humicola sp. 1 1.2⇓ 6.4 1.6 1.1
Absidia cylindrospora 8.1 4.3 3.1 2.8⇑
Sterile dark sp. 2 3.0 3.4 3.1 2.7
Cylindrocarpon destructans 4.0 3.2 2.6⇑ 1.7
Mortierella minutissima 1.8 1.7 1.0 1.1
Penicillium spp. 7.9 6.3 6.2 3.8⇑
Volutella sp. 0⇓ 2.8 1.6 1.6
Sterile dark sp. 3 1.2 2.7⇑ 0.5 0.5
Sterile dark sp. 4 0.6 2.0 0 0
Chrysosporium sp. 1 1.6 3.3⇑ 0.5 0.5
Phialophora sp. 1 0.6 0 1.5 0
Sterile hyaline sp. 2 0⇓ 0 0.5 0
Sterile hyaline sp. 3 1.2 1.1 2.1 0.5
Paecilomyces spp. 2.3 0.5 1.1 0.5
Sterile hyaline sp. 4 2.3 1.4 0 0
Sporothrix sp. 1.2 1.7 0 1.0
Chloridium sp. 1.2 ab 2.2⇑a 0 b 0.6 ab
Trichoderma polysporum 4.6 1.2 0.5 0
Sterile hyaline sp. 5 1.2 0 0.5 0.5
Chrysosporium sp. 2 0 1.2 1.1 0.5
Chaetomium sp. 1.1 0 0.5 1.6⇑
‘Isolated only from aged materials’
Trichoderma viride Pers. ex Gray 2.9 a 1.2 ab 2.5 ab 0 b
Petriellidium sp. 0.5 1.1 0 0
Fusarium sp. 0.6 1.2 0 0
Wardomyces sp. 0 1.1 0 0.5
Total number of taxa isolated 39 37 34 38
Average number of taxa per sample 22.5 ab 24.0⇑b 19.5⇑ac 18.8⇑c
aThe casts originated from B0 (soil only), BB (soil+beech litter) and BL (soil+lime litter) treatments. Bold⇑values indicate significant
increase in dominance during 100 days of incubation, underlined⇓values indicate significant decrease (t-test, p<0.05, n=4). Values within rows followed by different letters are significantly different (Tukey’s HSD test based on arcsine- transformed data, n=4, p<0.05).
significantly different in BB and BL casts and in soil
(p<0.05). In contrast, aged casts from the B0 treatment
could not be reliably separated from the soil (p=0.27).
4. Discussion
Two very different situations were modelled in our experiment. (i) L. terrestris consumed only mineral
Fig. 3. Discriminant analysis plane of aged soil and casts of Lumbricus terrestris according to the fungal dominance structure. The casts originated from B0 (soil only), BB (soil+beech litter) and BL (soil+lime litter) treatments. Ellipses represent 95% confidence limits.
from both substrates. Beech leaf litter appears to be low quality food for L. terrestris because in the BB treatment earthworm body mass decreased by 12.1% over 45 days. In contrast, Tilia litter was readily con-sumed and earthworm body mass increased by 9.1% in the BL treatment. The average quantity of litter materials in fresh casts was about 20% (w/w) in the BL treatment but less than 10% in the BB treatment (Tiunov and Scheu, 2000).
The soil washing technique, which, by removing fungal spores, isolates predominantly active mycelial fungi, was used to assess the composition of the fungal community in food materials and casts of L. terrestris. The results can be summarised as follows:
1. The passage of mineral soil through the gut of L. terrestris only weakly affected the dominance structure of the soil microfungal community. 2. The fungal community in casts deposited after
feeding on litter and soil significantly differed from that in soil and litter. The community is formed by mixing of soil and litter colonizing species and by selectively changing the occurrence of particular species due to gut passage. The composition of the fungal community in casts of L. terrestris strongly depended on the type of litter consumed.
3. Certain soil and litter inhabiting fungi (e.g. A. cylindrospora, Cladosporium spp., Alternaria spp.) were detrimentally affected by gut passage. The colonization rate (number of isolates per num-ber of plated particles) also strongly decreased in
casts. However, some fungi (mainly Trichoderma spp.) flourished in the fresh casts.
4. The dominance structure of fungi changed consid-erably during ageing of casts. Both the colonization rate and the diversity of species increased. How-ever, the fungal community in casts continued to differ from that in soil for a long time.
L. terrestris is a large earthworm species and in-gests large organic particles. Therefore, it does not conform to typical fungal feeders, defined as animals that selectively graze fungal material (Shaw, 1992). However, the composition of fungi in food material may affect its palatability to earthworms, particularly to litter feeding species like L. terrestris (Cooke, 1983; Moody et al., 1995). It is also likely that fungi are im-portant agents for the conditioning of litter in middens of L. terrestris.
The decrease in the colonization of organic particles by fungi and the decline in frequency of certain species indicate that at least some fungi were damaged by gut passage or by specific environmental conditions in fresh casts. These ‘sensitive’ species comprised about 5–10, 20–25 and 10–15% of the total number of iso-lates in lime litter, beech litter and soil, respectively. The feeding activity of L. terrestris appears to suppress early litter colonizers (Cladosporium, Alternaria) and secondary sugar fungi (Mucorales) in favour of Tri-choderma spp.
Knowledge on digestion of fungal hyphae during gut passage through earthworms is very limited; how-ever, by using fluorescence microscopy it has been concluded that fungal hyphae are digested in part by L. terrestris (Wolter and Scheu, 1999). The preference of invertebrate fungivores for darkly pigmented fungi has been stressed frequently (Shaw, 1992; Maraun et al., 1998). Digestion of Alternaria alternata and Cladosporium cladosporioides by the epigeic earth-worm Eisenia fetida has been reported by Marfenina and Ishchenko (1997). Therefore, it is likely that the decrease in frequency of Alternaria, Cladosporium and Phialophora species in casts of L. terrestris ob-served in the present study was caused by digestion of these species.
reported M. hiemalis and Trichoderma sp. to be of similar palatability to L. terrestris and two other earth-worm species. Also, the viability of spores of Tri-choderma sp. and M. hiemalis has been found to be reduced strongly by gut passage (by 99 and 90%, re-spectively), presumably due to the action of intestinal fluids (Moody et al., 1996). T. koningii and T. viride, and also Mortierella ramanniana were found to be digested by the tropical earthworm species Drawida assamensis (Tiwari et al., 1990). In contrast, Dom-sch and Banse (1972) reported a strong increase in the frequency of T. hamatumand Mortierella zonata in casts of L. terrestris. In accordance with this finding, the frequency of the three dominant Trichoderma spp. (T. pseudokoningii, T. koningii and T. hamatum) was strongly increased in casts of L. terrestris in our exper-iment. Three mechanisms might have been responsible for these changes: (i) L. terrestris may have selectively consumed litter particles colonized by Trichoderma spp.; (ii) in the gut of earthworms and in fresh casts, Trichoderma spp. might have been competitively su-perior to other fungal species, which might be due to the ability of Trichoderma spp. to colonize new sub-strates quickly but also due to damage to competing fungal species during gut passage; (iii) species of the genus Trichoderma were little damaged by gut pas-sage or balanced the damage by compensatory growth (Hedlund and Augustsson, 1995). We assume the last mechanism to be most important since the number of particles colonized by Trichoderma spp. was consis-tently higher in casts than in ingested materials.
Fungal species which occupy resources earlier may have a competitive advantage over later incom-ers (Stahl and Christensen, 1992). Thus, ‘tolerant’ species like Trichoderma may benefit from the feed-ing activity of L. terrestris. This may be of profound importance for the succession of fungi on decom-posing litter, since species of the genus Trichoderma are known to be effective competitors and produce a wide range of antifungal substances (Domsch et al., 1980).
Domsch and Banse (1972) reported that the percent-age of T. hamatum in casts of L. terrestris increased strongly during 30 days of incubation. In our experi-ment the frequency of Trichoderma spp. was not af-fected by cast incubation in B0 and BB treatments, but it was reduced in the BL treatment (from 28.5% in fresh to 16.3% in aged casts). Simultaneously, the
fre-quency of Absidia, Penicillium and other ‘soil fungi’ increased.
The role of earthworms in the dispersal of beneficial or harmful soil fungi is controversial (Edwards and Fletcher, 1988; Brown, 1995; Pattinson et al., 1997). In our study the majority of soil and litter inhabiting fungi were able to survive gut passage through L. terrestris. Therefore, we assume that the feeding and burrowing activity of L. terrestris accelerates the colonization of litter by the edaphic mycoflora but also extends the range of litter-associated fungi to mineral soil layers. The vertical movement of L. terrestris ranges from the soil surface to a depth of up to 2 m, which corresponds with the range of cast deposition (Joergensen et al., 1998). It is likely that in temperate ecosystems the role of L. terrestris and other anecic earthworm species in vertically transporting fungal propagules through the soil profile can hardly be effectively substituted by any other mechanism.
Acknowledgements
Financial support by the Volkswagen-Stiftung, Fed-eral Republic of Germany, is gratefully acknowledged. We thank Dr. Yu.G. Puzachenko for valuable advice on statistical analysis.
References
Brown, G.G., 1995. How do earthworms affect microfloral and faunal community diversity? Plant and Soil 170, 209–231. Cooke, A., 1983. The effects of fungi on food selection by
Lumbricus terrestris L. In: Satchell, J.E. (Ed.), Earthworm Ecology. Chapman and Hall, London, pp. 365–373.
Dash, H.K., Beura, B.N., Dash, M.C., 1986. Gut load, transit time, gut microflora and turnover of soil, plant and fungal material by some tropical earthworms, plant and fungal material by some tropical earthworms. Pedobiologia 29, 13–20.
Dash, M.C., Satpathy, B., Behera, N., Dei, C., 1984. Gut load and turnover of soil, plant and fungal material by Drawida calebi, a tropical earthworm. Rev. Ecol. Biol. Soil 21, 387–393. Domsch, K.H., Banse, H.J., 1972. Mykologische Untersuchungen
an Regenwurmexkrementen. Soil Biol. Biochem. 4, 31–38. Domsch, K.H., Gams, W., Anderson, T.H., 1980. Compendium of
Soil Fungi. Academic Press, London, 859 pp.
Edwards, C.A., Fletcher, K.E., 1988. Interactions between earthworms and microorganisms in organic-matter breakdown. Agric. Ecosyst. Environ. 24, 235–247.
Hedlund, K., Augustsson, A., 1995. Effects of enchytraeid grazing on fungal growth and respiration. Soil Biol. Biochem. 27, 905– 909.
Joergensen, R.G., Küntzel, H., Scheu, S., Seitz, D., 1998. Movement of faecal indicator organisms in earthworm channels under a loamy arable and grassland soil. Appl. Soil Ecol. 8, 1–10.
Jones, C.G., Lawton, J.H., Shachak, M., 1997. Positive and negative effects of organisms as physical ecosystem engineers. Ecology 78, 1946–1957.
Maraun, M., Migge, S., Schaefer, M., Scheu, S., 1998. Selection of microfungal food by six oribatid mite species (Oribatida, Acari) from two different beech forests. Pedobiologia 42, 232–240. Marfenina, O.E., Ishchenko, I.A., 1997. Earthworms preference
for soil microscopic fungi. Izvestiya Akademii Nauk Seriya Biologicheskaya 4, 504–506 (in Russian).
McLean, M.A., Parkinson, D., 1998. Impacts of the epigeic earthworm Dendrobaena octaedra on microfungal community structure in pine forest floor: a mesocosm study. Appl. Soil Ecol. 8, 61–75.
Moody, S.A., Briones, M.J.I., Piearce, T.G., Dighton, J., 1995. Selective consumption of decomposing wheat straw by earthworms. Soil Biol. Biochem. 27, 1209–1213.
Moody, S.A., Piearce, T.G., Dighton, J., 1996. Fate of some fungal spores associated with wheat straw decomposition on passage through the guts of Lumbricus terrestris and Aporrectodea longa. Soil Biol. Biochem. 28, 533–537.
Newell, K., 1984. Interactions between two decomposer basidiomycetes and a collembolan under Sitka spruce: distribution, abundance and selective grazing, abundance and selective grazing. Soil Biol. Biochem. 16, 227–233.
Parkinson, D., Visser, S., Whittaker, J.B., 1979. Effects of collembolan grazing on fungal colonization of leaf litter. Soil Biol. Biochem. 11, 529–535.
Parkinson, D., Williams, S.T., 1961. A method for isolating fungi from soil microhabitats, Plant . Plant and Soil 13, 347–355.
Pattinson, G.S., Smith, S.E., Doube, B., 1997. Earthworm Aporrectodea trapezoides had no effect on the dispersal of a vesicular-arbuscular mycorrhizal fungi, Glomus intraradices. Soil Biol. Biochem. 29, 1079–1088.
Puzachenko, Yu.G., Kuznetsov, G.V., 1998. Ecological differentia-tion of rodents in tropical semi-evergreen broad-leaved forests of North Vietnam. Zoologicheskij Zhurnal 77, 117–132 (in Russian).
Scheu, S., Parkinson, D., 1994. Effects of earthworms on nutrient dynamics, carbon turnover and microorganisms in soils from cool temperate forests of the Canadian Rocky Mountains — laboratory studies. Appl. Soil Ecol. 1, 113–125.
Shaw, P.J.A., 1992. Fungi, fungivores, and fungal food webs. In: Carroll, G.C., Wicklow, D.T. (Eds.), The Fungal Community. Marcel Dekker, New York, pp. 295–310.
Stahl, P.D., Christensen, M., 1992. In vitro mycelial interactions among members of a soil microfungal community. Soil Biol. Biochem. 24, 309–316.
Striganova, B.R., Marfenina, O.E., Ponomarenko, V.A., 1988. Some aspects of the effect of earthworms on soil fungi. Izvestiya Akademii Nauk Seriya Biologicheskaya 5, 715–719 (in Russian).
Tiunov, A.V., Scheu, S., 2000. Microbial biomass, biovolume . biovolume and respiration in Lumbricus terrestris cast material of different age. Soil Biol. Biochem.32, 265–275.
Tiwari, S.C., Mishra, R.R., 1993. Fungal abundance and diversity in earthworm casts and in uningested soil. Biol. Fertil. Soils 16, 131–134.
Tiwari, S.C., Tiwari, B.K., Mishra, R.R., 1990. Microfungal species associated with the gut content and casts of Drawida assamensis Gates. Proc. Indian Acad. Sci. (Plant Sci.) 100, 379–382. Visser, S., 1985. Role of soil invertebrates in determining the
composition of soil microbial communities. In: Fitter, A.H., Atkinson, D., Read, D.J., Usher, M.B. (Eds.), Ecological Interactions in Soil. Blackwell, Oxford, pp. 297–317. Wolter, C., Scheu, S., 1999. Changes in bacterial numbers and