6 BAB 2
TINJAUAN PUSTAKA
2.1 Definisi
Endometriosis (awalnya bernama adenomioma) ditemukan 150
tahun yang lalu oleh Rokitansky sebagai adanya kelenjar epitel dan
stroma ektopik di endometrium. Endometriosis adalah penyakit jinak yang
didefinisikan sebagai adanya jaringan kelenjar dan stroma endometrium
ektopik atau di luar dari kavum uteri dan dihubungkan dengan nyeri pelvik
dan infertilitas. Kelenjar dan stroma ekstra uterin atau lesi endometriosis
dapat ditemukan di berbagai tempat terutama ovarium dan pada
permukaan organ rongga pelvik.13-15
Dulu, implan jaringan endometriosis yang klasik dikenal sebagai lesi
yang gelap, berwarna biru-hitam, bersifat seperti bubuk yang terbakar,
berada di bawah lapisan peritoneal. Sekarang, endometriosis ditemukan
dalam berbagai jenis dan bentuk lainnya. Lesi dapat bervariasi warna, dari
merah sampai coklat, hitam, putih atau kuning; lesi juga dapat ditemukan
dalam bentuk yang jernih atau vesikel merah. Bentuk dari lesi bergantung
pada lokasi, suplai darah, perdarahan yang terjadi, ada tidaknya fibrosis,
dan dapat merefleksikan subtipe fenotipe dari penyakit ini. Pada
kenyataannya, beberapa peneliti mempercayai bahwa lesi atipikal dapat
memiliki aktivitas biokimia yang lebih kompleks dan dapat menjadi tipe
7 2.2 Epidemiologi
Pada umumnya endometriosis paling sering ditemukan pada usia
reproduksi. Insidensi pastinya belum diketahui, namun prevalensinya pada
kelompok tertentu sering ditemukan.Prevalensi endometriosis pada
ovarium masih belum pasti diketahui. Endometriosis mempengaruhi
5-10% pada semua wanita usia reproduktif dan 60-80% dari wanita infertil
atau wanita dengan nyeri pelvis, dengan usia rata-rata 25 hingga 30
tahun.4-6,11,19
Endometriosis lebih sering terjadi pada wanita yang memiliki siklus
ovulatoar yang biasanya terjadi pada umur antara 20 sampai 45 tahun
dibandingkan dengan wanita yang memiliki siklus anovulatoar yang
berumur perimenars atau perimenopause, yaitu sebesar 22%. Di Amerika
Serikat, endometriosis mempengaruhi 5-10% wanita usia reproduktif.5 Di
Indonesia sendiri ditemukan 15-25% wanita infertil yang disebabkan oleh
endometriosis.
2.3 Etiopatogenesis
Hingga kini belum diketahui secara pasti penyebab dari
endometriosis. Banyak teori yang disebut ikut berperan dalam
patogenesis endometriosis, sehingga endometriosis juga disebut sebagai
“penyakit penuh teori”.1,13,20-22 Ada berbagai teori mengenai endometriosis
diantaranya:
Teori Sampson/Teori implantasi Endometriosis Peritoneal: Teori
8
endometriosis, yang dikemukakan oleh Sampson pada tahun 1927,
mengajukan mekanisme yang paling banyak diterima mengenai
terjadinya endometriosis terjadi karena darah haid mengalir kembali
(retrograd) dari sel endometrium yang luluh melalui saluran tuba ke
dalam kavum peritoneum dan berimplantasi pada permukaan
peritoneum. Endometriosis dipercaya sebagai akibat dari defek
molekuler yang membantu survival jaringan endometrium pada
debris menstruasi di peritoneum.10,12,23 Profil ekspresi gen yang
dikarakteristikkan oleh microarray pada endometrium wanita dengan
dan tanpa endometriosis, menunjukkan bahwa beberapa gen target
progesteron mengalami disregulasi selama jendela implantasi, yaitu
saat endometrium terpapar pada kadar progesteron paling tinggi.13-14
Penelitian eksperimental Wiltz menyatakan stroma dan epitel
endometrium dapat dengan mudah dan cepat melekat pada
mesotelium (invasi transmesotelial terjadi antara 18-24 jam)24
Teori Metaplasia dikemukakan oleh Meyer, yang mengatakan bahwa
lesi endometriosis terbentuk akibat transformasi (metaplasia) dari
sel-sel epitel coelomic yang berasal dari saluran Muller, sehingga
terbentuk jaringan endometrium yang menjadi dasar terjadinya
endometriosis. Teori ini disebut teori coelom metaplasia bahwa
endometriosis berasal dari metaplasia sel khusus pada lapisan
mesotelial dari visceral dan peritoneum. Faktor hormonal atau
imunologi diperkirakan merangsang transformasi sel endometrium
9
embrio sisa dari Wolffian atau saluran Mullerian bertahan dan
berkembang menjadi lesi endometriosis. Namun, teori ini tidak
sempurna karena tidak dapat menjelaskan lesi endometriosis yang
ditemukan di daerah diluar perjalanan duktus Mullerian.25-27 Inflamasi dan Respon Imun
Data yang cukup telah menyatakan bahwa endometriosis
dihubungkan dengan sebuah keadaan inflamasi subklinis peritoneum
yang ditandai oleh peningkatan volume cairan peritoneum,
peningkatan konsentrasi sel darah putih cairan peritoneum (terutama
makrofag dengan peningkatan aktivitasnya) dan peningkatan sitokin
inflamasi, faktor pertumbuhan dan substansi penyokong
angiogenesis. Tingkat aktivasi basal yang lebih tinggi dari makrofag
peritoneum pada pasien dengan endometriosis dapat mengganggu
fertilitas dengan cara menurunkan motilitas sperma, meningkatkan
fagositosis sperma atau mengganggu fertilisasi, mungkin dengan
meningkatkan kadar sitokin seperti TNF-α. Makrofag dapat menyokong pertumbuhan sel-sel endometrium dengan cara
mensekresi growth factor dan angiogenetic factor seperti epidermal
growth factor (EGF), macrophage-derived growth factor (MDGF),
fibronektin dan adhesion molecule seperti integrin. Setelah
perlekatan sel-sel endometrium ke peritoneum, terjadi invasi dan
pertumbuhan lebih lanjut yang tampaknya diregulasi oleh matrix
10
Sitokin inflamasi memainkan peran sentral dalam regulasi proliferasi,
aktivasi, motilitas, adhesi, kemotaksis dan morfogenesis dari sel.
Beberapa sitokin seperti IL-1, IL-5, IL-6, IL-8, IL-15, monocyte
chemotactic protein-1 (MCP-1), TNF-α, transforming growth factor-β
(TGF-β) dan Regulated on Activation, Normal T-cell Expressed dan
Secreted (RANTES) telah diimplikasikan dalam patogenesis
endometriosis. Ekspresi TNF-α, IL-8, dan MCP-1 lebih tinggi pada endometriosis tingkat dini dan menurun pada endometriosis tingkat
lanjut, sementara ekspresi TGF-β menurun dengan penurunan keparahan penyakit. RANTES juga meningkat dalam cairan
peritoneum wanita dengan penyakit yang lebih berat.29-31
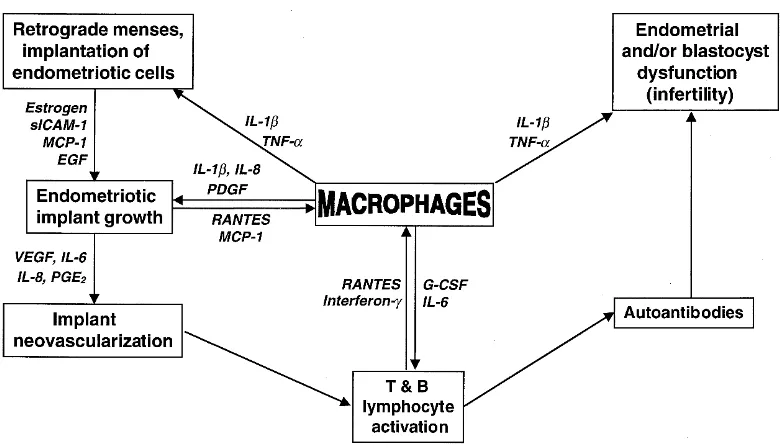
Gambar 1. Imunobiologi Endometriosis32
Sistem imun manusia sehat menyingkirkan sel-sel endometrium
11
lesi endometriosis. Proses ini mungkin difasilitasi oleh perubahan
apoptosis sel-sel endometrium yang normalnya meningkat pada
akhir siklus menstruasi tetapi proses apoptosis ini secara signifikan
menurun pada endometriosis. Dengan demikian pada wanita sehat,
sel-sel endometrium yang didiseminasi ke dalam lokasi ektopik
mungkin diprogram untuk mengalami kematian dan dengan mudah
dieliminasi oleh sistem imun.29,30,33,34
Endometriosis dapat disebabkan oleh penurunan pembersihan
sel-sel endometrium cairan peritoneum akibat penurunan aktivitas sel-sel
Natural Killer (NK)atau penurunan aktivitas makrofag. Penurunan
sitotoksisitas yang dimediasi secara seluler terhadap sel-sel
endometrium autolog telah dihubungkan dengan endometriosis.33,34
Endometriosis merupakan kondisi inflamasi dimana sejumlah besar
leukosit direkrut dari sirkulasi darah ke dalam lesi endometriosis
sehingga terjadi perubahan jumlah dan fungsi dari leukosit ini dalam
endometrium eutopik dan cairan peritoneum dan juga dalam lesi
endometriosis. Makrofag, sel NK, limfosit T, limfosit B, sel mast dan
sel dendritik meningkat dalam lesi endometriosis sebagai melalui
ekstravasasi dari sirkulasi darah ke dalam lesi endometriosis dimana
terjadi perubahan fungsi sel T regulator yang mempengaruhi
terjadinya endometriosis dan progresifitasnya.32,33,34
Peranan Makrofag, fagosit mononuklear (monosit dan makrofag)
ditemukan pada kebanyakan jaringan tubuh dan berperan vital
12
bersirkulasi yang diproduksi disumsum tulang dari progenitor mieloid
bersama adalah prekursor untuk makrofag jaringan. Monosit mampu
berdiferensiasi menjadi sel-sel efektor yang heterogen secara
morfologi dan secara fungsional, termasuk makrofag yang tinggal
dalam jaringan dan makrofag inflamasi.Selama proses inflamasi,
monosit direkrut ke jaringan yang mengalami jejas dengan cara
melekat ke endotel pembuluh darah dan mengikuti gradien
haptotaktik dan kemotaktik lokal sebelum berdiferensiasi menjadi
makrofag. Makrofag baik yang tinggal di dalam jaringan atau yang
baru direkrut adalah sumber utama kemokin dalam jaringan. 29,32,-34
Makrofag mononuklear mengikuti neutrofil ke dalam inflamasi,
memfagosit debris seluler dan material asing dan akhirnya keluar
dari tempat inflamasi. Makrofag bisa diaktifkan secara klasik (M1
makrofag) atau diaktifkan secara alternatif (M2 makrofag), tetapi ada
heterogenitas substansial dalam fenotip makrofag, karena sebagian
peran luas yang makrofag jalankan dalam respon inflamasi dan
dalam mempertahankan homeostasis jaringan. 32
Makrofag mempertahankan host dengan pengenalan, fagositosis
dan destruksi mikroorganisme yang menyerang dan juga berperan
sebagai scavenger, membantu untuk membersihkan sel-sel yang
mengalami apoptosis dan debris seluler. Makrofag mensekresikan
berbagai sitokin, faktor pertumbuhan, enzim-enzim, dan
prostaglandin yang membantu memperantarai fungsinya sendiri
13
Makrofag memiliki habitat normal pada cairan peritoneum dan jumlah
dan aktivitasnya sangat meningkat pada wanita dengan
endometriosis. Daripada bekerja sebagai scavenger (makrofag M1)
untuk mengeliminasi sel-sel endometrium ektopik, makrofag
peritoneum yang diaktifkan secara alternatif (makrofag M2) dan
monosit sirkulasi pada wanita dengan endometriosis tampaknya
menyokong endometriosis dengan mensekresi faktor pertumbuhan
dan sitokin yang menstimulasi proliferasi endometrium ektopik dan
menghambat fungsi scavengernya. 29-33
Aktivasi alternafif makrofag (makrofag M2) adalah langkah kunci
dalam perkembangan endometriosis dimana peningkatan makrofag
M2 ini akan mensekresi dan meningkatkan konsentrasi sitokin,
prostaglandin, komponen komplemen, dan faktor pertumbuhan
seperti tumor necrosis factor-β (TNF-α), IL-6, dan transforming
growth factor-β (TGF-β). Normalnya sel-sel endometriosis yang
masuk ke kavum peritoneal disingkirkan oleh makrofag. Mekanisme
aberasi pada endometriosis ini mengakibatkan tidak efektifnya
sistem pembersihan imunologis terhadap agen asing. Makrofag M2
dan peningkatan kadar sitokin mengakibatkan inisiasi, progresi dan
pertumbuhan sel-sel endometrium juga neovaskularisasi.32,34-35
Makrofag M2 lebih berperan dibandingkan makrofag M1 dalam
patogenesis endometriosis. Hal ini mungkin disebabkan oleh faktor
genetik, hormonal dan lingkungan. Sebuah penelitian menyatakan
14
reseptor estrogen yang diekspresikan pada permukaannya. Di
bawah pengaruh estrogen ini makrofag M2 akan mensekresikan
sitokin dan faktor pertumbuhan (seperti VEGF, hepatocyte growth
factor, dan TNF-α) yang berkontribusi terhadap perkembangan dan
persistensi endometriosis.30-32,35
Proses kemotaktik sel inflamasi oleh kemokin dimediasi oleh
reseptor spesifik kemokin. Fungsi kemokin sebagai regulator
motilitas dan orientasi leukosit adalah sebagai mediator proinflamasi,
imunomodulator kuat (aktivasi dan diversifikasi limfosit), modifier
biologis fungsi eritrosit dan faktor angiogenik. Kemokin berikatan dan
mengaktifkan reseptor spesifik pada permukaan leukosit.29,32-35
-kemokin CCL5 yang juga dikenal sebagai RANTES (regulated
upon activation, normal T cell expressed and secreted). RANTES
atau CCL5 dihasilkan dari stroma endometrium dan dipengaruhi oleh
estrogen yang dominan estrogen lokal dari aromatase. RANTES
memberi sinyal pada T limfosit untuk diaktifkan menghasilkan
sitokin.29,35
Hampir setiap wanita mengalami menstruasi retrograd setiap
bulannya. Dan secara fisiologis setiap sel endometrium akan
menyebabkan reaksi inflamasi dengan menghasilkan MCP-1 dan
RANTES dari sel stroma dan kelenjar endometrium yang dipicu oleh
estrogen yang dihasilkan dari proses aromatase yang diketahui
15
yang berpengaruh terhadap rekrutmen dari monosit dan makrofag ke
jaringan lesi endometriosis.29,32,35
Peningkatan jumlah makrofag ditemukan dalam cairan peritoneal
penderita dengan endometriosis. Makrofag ini juga ditemukan
memiliki efek stimulasi pada jaringan endometrium, dibandingkan
dengan makrofag wanita tanpa endometriosis yang memiliki efek
penekanan.29-35
Perubahan cairan peritoneum yang menunjukkan peningkatan
aktivitas makrofag, sekresinya adalah beberapa sitokin yang
menyebabkan terjadinya proses apoptosis patologis. Hal ini terutama
ditemukan pada endometriosis berat dengan infertilitas, dimana
terjadi proses tersebut pada sel granulosa ovarium dengan
ditemukan kadar Interleukin-6 (IL-6) dan IL-8 yang tinggi pada cairan
peritoneum. Pertumbuhan lebih lanjut dari sel endometrium akibat
menstruasi retrograd kemungkinan juga melibatkan sistem imun
penderita endometriosis. Suatu proses imunologi yang sangat
komplek dan saling terkait diduga berperan pada pertumbuhan lebih
lanjut dari sel endometrium yang terlepas. Hal ini berhubungan
dengan dijumpainya sel limfoid pada implant endometriosis. Selain
itu dijumpai juga adanya peningkatan kadar makrofag dan limfosit T
didalam cairan peritoneum. Keadaan ini mungkin merupakan salah
satu awal dari proses inflamasi yang komplek. Terjadi pula
peningkatan kadar sitokin dan growth factor yang dihasilkan oleh
16
berpengaruh pada sel induknya sendiri dan parakrin yang
berpengaruh pada sel disekitarnya atau masuk peredaran darah
maupun rongga tubuh yang cukup jauh. Para peneliti menemukan
jenis sitokin yang meningkat diantaranya adalah RANTES
(Regulated on Activation, Normal T-cell Expressed and Secreted),
IL-1 (Interleukin-1), IL-6 dan TNF (Tumor Necrosis Factor). Sedangkan
faktor pertumbuhan yang meningkat pada penderita endometriosis
diantaranya adalah VEGF (Vascular Endothelial Growth Factor). IL-1
merupakan sitokin yang memiliki peran penting dalam mengatur
inflamasi dan respon imun. IL-1 yang dihasilkan oleh aktif monosit
dan macrophage, memiliki dua macam reseptor yaitu reseptor alfa
dan beta, tetapi keduanya dapat dihambat dengan satu macam
reseptor antagonis IL-1. IL-1 beta dapat memicu faktor angiogenesis
seperti VEGF dan IL-6 sehingga terjadi pertumbuhan pembuluh
darah pada stroma endometriosis, tetapi tidak pada stroma
endometrium normal.29-34
Pada penelitian Djaganata SP, didapatkan bahwa jumlah sel NK
dengan ekspresi sel NK (CD56) adalah tidak ada perbedaan
signifikan antara jaringan endometriosis dibandingkan dengan
jaringan endometrium normal. Temuan ini menggambarkan bahwa
sel imun seluler terutama sel NK tidak merespon dengan baik
sebagaimana reaksi normal terhadap keadaan inflamasi/terinfeksi
yang seharusnya sel NK tersebut meningkat dalam jumlah dan
17
dengan endometriosis terjadi defek kuantitatif dan kualitatif pada sel
NK dikarenakan kegagalan dalam aktivasi yang disebabkan dari
menurunnya IL-2 sebagai imunodilator sel NK dan kegagalan sel NK
saat pengenalan sel target (sel endometrium ektopik), sel NK
mengenali sel target dengan mendeteksi adanya ekspresi MHC
kelas 1 pada permukaan sel target, apabila dijumpai MHC kelas 1
maka akan timbul sinyal negatif terhadap sel NK dan kemudian sel
NK akan mengekspresikan KIRs, karena itu sel NK tidak
tertarik/terikat ke jaringan lokal inflamasi (jumlah yang sama pada
wanita normal), dan tidak teraktivasi seperti keadaan normal tanpa
inflamasi. Sel NK menganggap sel endometrium ektopik adalah sel
yang normal, walaupun tidak pada tempat semestinya. Dapat
disimpulkan pada wanita dengan endometriosis mengalami
gangguan sistem imun tubuh seluler pada cairan rongga peritoneum.
Sistem imun tubuh seluler (makrofag, limfosit dan sel NK) tidak dapat
secara adekuat membersihkan jaringan endometrium ektopik yang
berada di cairan rongga peritoneum, dan justru mendukung
terjadinya proses implantasi, angiogenesis, proliferasi, dan
imortalitas sel endometrium ektopik sehingga terjadi endometriosis.
Hal ini mungkin disebabkan dari kegagalan sel NK merespon
keadaan pada endometriosis, yaitu saat pengenalan sel target,
ketidakseimbangan respon dan produksi sitokin M1 yang lebih
rendah dibandingkan M2, dan Th1 yang lebih rendah dibandingkan
18
keadaan semestinya/keadaan inflamasi) dan aktivitas sel-sel imun
seluler, yaitu dengan fungsi sitotoksis yang rendah dan produksi
sitokin yang tinggi dalam cairan peritoneum. Penelitian ini pada
dasarnya hanya mendeteksi terjadinya penurunan jumlah (kuantitatif)
sel NK.30,32-36
2.4 Diagnosis
Endometriosis tidak dapat didiagnosis hanya berdasarkan gejala
klinis saja. Diduga endometriosis jika memiliki masalah tentang fertilitas,
nyeri haid, nyeri saat berhubungan dan nyeri pelvis kronis. Mekanisme
terjadinya nyeri pada endometriosis ini mungkin disebabkan oleh
peradangan lokal, infiltrasi yang dalam dengan kerusakan jaringan,
terlepasnya prostaglandin dan perlengketan.11,14,20,21,22
Perdarahan tidak teratur yang berhubungan dengan endometriosis
diperkirakan terjadi pada 11-34% penderita endometriosis. Hal ini
dikatakan diakibatkan oleh adanya kelainan pada ovarium yang luas
sehingga fungsi ovarium terganggu. Perdarahan ini juga dihubungkan
dengan terjadinya peningkatan kadar estrogen dan berkurangnya
progesteron yang mengakibatkan terganggunya keseimbangan eutopik
endometrium penderita endometriosis.10,23
Banyak juga wanita yang menderita endometriosis tetapi tidak
memiliki keluhan. Gold standard untuk diagnosis endometriosis memakai
pemeriksaan laparoskopi dengan atau tanpa biopsi untuk pemeriksaan
19
anamnesis, pemeriksaan fisik, pemeriksaan ginekologi, pemeriksaan
penunjang non invasif dan pemeriksaan serum CA 125 tetap diperlukan,
meskipun serum CA-125 mungkin dapat meningkat pada endometriosis
derajat sedang dan berat, ketentuan ini tidak dianjurkan sebagai
pemeriksaan rutin.12,14
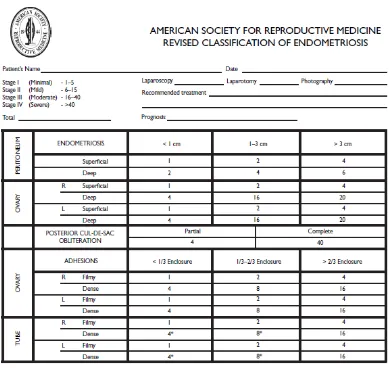
2.5 Klasifikasi
Sistem klasifikasi endometriosis oleh American Society for
Reproductive Medicine (ASRM) yang telah direvisi berdasarkan penilaian
terhadap lesi endometriosis pada peritoneum dan tuba menggunakan nilai
yang berhubungan dengan ukuran lesi. Penilaian ini juga didasarkan pada
perlengketan pada ovarium dan tuba fallopi. Dan juga terdapat penilaian
20
Gambar 2. Klasifikasi Endometriosis Berdasarkan The American Society
21
Penyakit yang minimal (stadium I) dengan skor: 1-5 dan ringan
(stadium II) dengan skor: 6-15 dikarakteristikkan dengan implan yang
menyebar kecuali pada daerah superfisial di tuba fallopi atau ovarium,
tanpa skar atau perlekatan pada anterior atau posterior permukaan
ovarium. Penyakit yang sedang (stadium III) dengan skor: 16-40,
dikarakteristikkan dengan implan multipel atau endometrioma berukuran
kecil (2 cm) meliputi satu atau kedua ovarium, minimal perlekatan
peritubular atau periovarium; tersebar dengan parut dan retraksi pada
struktur lainnya. Penyakit yang berat (stadium IV) dengan skor: > 40
dikarakteristikkan dengan endometrioma ovarium yang besar, perlekatan
tuba atau ovarium, obstruksi tuba, obliterasi cul-de-sac, meliputi
uterosakral dengan keterlibatan usus atau traktus urinarius yang nyata.1
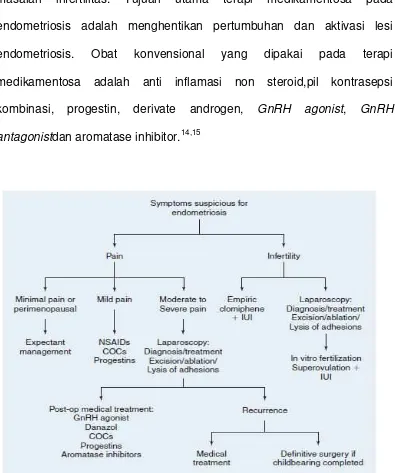
2.6 Penatalaksanaan
Penatalaksanaan endometriosis berdasarkan keluhan dari
penderita, lokasi lesi endometriosis, tujuan pengobatan dan keinginan
untuk hamil. Pada wanita penderita endometriosis derajat minimal sampai
ringan dapat dilakukan penatalaksanaan ekspektatif. Seperti Sutton dan
rekan (1997), endometriosis yang telah dikonfirmasi melalui laparoskopi
derajat minimal dan ringan yang dilakukan ekspektatif selama 1 tahun
29% pasien mengalami regresi penyakit, 42% tidak mengalami perubahan
dan 29% mengalami perkembangan penyakitnya. Penatalaksanaan
endomeriosis yaitu terapi medikamentosa, tetapi pembedahan dan
22
bermanfaat untuk mengatasi keluhan nyeri tetapi kurang bermanfaat untuk
masalah infertilitas. Tujuan utama terapi medikamentosa pada
endometriosis adalah menghentikan pertumbuhan dan aktivasi lesi
endometriosis. Obat konvensional yang dipakai pada terapi
medikamentosa adalah anti inflamasi non steroid,pil kontrasepsi
kombinasi, progestin, derivate androgen, GnRH agonist, GnRH
antagonistdan aromatase inhibitor.14,15
Gambar 3. Algoritma diagnostik dan penatalaksanaan pada wanita
dengan endometriosis. COCs = combination oral contraceptives; GnRH=
gonadotropin releasing hormone; IUI = intrauterine insemination; NSAIDs
23 2.7Reseptor Progesteron (PR)
Progesteron, adalah modulator kunci fungsi reproduksi normal,
termasuk ovulasi, uterus, perkembangan kelenjar mamae, dan ekspresi
respon seksual. Progesteron adalah hormon steroid yang sangat penting
sebagai regulator fisiologi reproduksi wanita normal. Progesteron
disekresikan terutama oleh korpus luteum yang berkembang di ovarium
setelah ovulasi. Progesteron mempengaruhi beberapa jaringan dan organ,
termasuk otak, payudara, uterus, ovarium, dan serviks. Hormon
progesteron ditemukan berperan dalam fungsi proliferasi atau memicu
diferensiasi terutama pada epitel. Progesteron menghalangi proliferasi
yang dipengaruhi oleh estrogen pada endometrium normal.Walaupun
begitu, peran progesteron terus diteliti karena estrogen dan progesteron
bersama-sama berperan dalam proliferasi sel stroma endometrium.7,22,37
Hormon steroid seperti estradiol dan progesteron yang lipofilik dan
masuk kedalam sel dan inti dengan menyebarkan melalui plasma dan
membran nuklir. Setelah di inti, steroid menemukan protein yang dikenal
sebagai reseptor karena mereka mengikat ligan yang sama dengan
afinitas tinggi dan spesifisitas.
Reseptor progesteron pada model mencit mengkonfirmasi perlunya
progesteron pada proses reproduksi yang normal dan pengaruhnya pada
berbagai jenis sel.21,22 Reseptor progesteron yang teraktivasi juga
memainkan peran penting dalam remodelling jaringan uterus. Uterus
terdiri dari tipe sel heterogen dengan gelombang sinkron dari proliferasi
24
estrogen dan progesteron.38-41
Pada manusia, efek progesteron yang dimediasi oleh PR dengan
inisiasi alternatif transkripsi dari dua promotor yang berbeda. Dua isoform
ini adalah PR-A dan PR-B yang dihasilkan gen tunggal dengan transkripsi
di dua promotor yang berbeda dan dengan inisiasi translasi di dua
promotor distal dan proksimal yang diduga untuk reseptor estrogen, yang
disebut Estrogen Response Element (ERE). PR-A dan PR-B memiliki
sekuensi yang identik, dimana PR-B lebih panjang dengan 164 asam
amino di N-terminal yang diapit oleh (Activation Factor) AF-1 dan AF-2,
PR-A dan PR-B secara fungsional mediator yang berbeda dari kerja
progesteron pada epitelium uterin. Fungsi kedua reseptor ini juga
berbeda. Studi pada tikus dengan ablasi selektif isoform PR
mengungkapkan bahwa PR-A diperlukan untuk ovulasi dan modulasi efek
antiproliferatif progesteron di dalam uterus, dan PR-B diperlukan untuk
perkembangan normal dan fungsi kelenjar mammae. Bukti terbaru telah
menyarankan adanya isoform reseptor progesteron ketiga fungsional,
yaitu PR-C, yang diyakini memainkan peran penting dalam persalinan.23
PR-C tidak memiliki N terminal PR dan satu finger zinc dari domain
ikatan DNA, sehingga inaktif dalam transkipsi tetapi dapat berikatan
dengan hormon, dimerisasi, dan terletak di nukleus. Beberapa laporan in
vitro menunjukkan bahwa PR-C dapat memicu atau menghambat aktivitas
PR-A/B melalui pengaruh dengan memberikan sinyal, namun belum
25
PR terdiri dari DNA Binding Domain (DBD) dan terminal karboksil
Ligand Binding Domain (LBD). Sebagai tambahan, PR memiliki beberapa
fungsi aktivasi dan inhibisi, yang akan memicu atau menghambat aktivasi
transkipsi oleh PR. Dalam keadaan inaktif, PR memiliki kompleks kaperon
multiprotein yang inaktif. Ikatan progestin pada PR akan menyebabkan
perubahan konformasional, disosiasi kaperon, dimerisasi, dan mengikat
ke elemen respon progestin pada promoter gen target, memicu sekresi
koaktivator spesifik dan faktor transkripsi. Selain itu, PR akan
mengaktifkan transkripsi melalui jalur non genomik seperti aktivasi sinyal
kaskade second messenger dan cross talk reseptor dengan sinyal faktor
pertumbuhan.22,42-45
Meskipun urutan LBD dari PR-A dan PR-B adalah identik, akan
tetapi kemampuan ligannya berbeda untuk dapat menginduksi perubahan
konformasi dalam PR.58Dari sudut pandang mekanisme kerja, dari
pengamatan dilihat bahwa PR-A saja sudah cukup untuk mendukung
ovulasi yang normal,yang menunjukkan bahwa interaksi antara
heterodimeric PR-A dan protein PR-B tidak diharuskan untuk dapat
mengatur progestin penting yang responsif terhadap gen target yang
terkait dengan ovulasi. Ekspresi rahim dari isoform PR-A diperlukan tidak
hanya untuk menghambat proliferasi estrogen-induced tetapi juga yang
disebabkan oleh progesteron yang bekerja melalui protein PR-B.46
Meskipun mekanisme yang tepat yang mendasari kegiatan
diferensial dari dua isoform PR manusia tidak sepenuhnya dipahami,
struktur-26
fungsi menunjukkan bahwa domain Activation Factor (AF3) terletak di
regio sekuensi upstream PR-B, yang tidak ditemukan dalam PR-A. PR-B
spesifik mengkode AF3 yang tidak dijumpai dari PR-A. Domain ini
berkontribusi terhadap aktivitas transkripsi dengan menekan aktivitas
domain inhibitor pada urutan umum untuk PR-A dan PR-B. Selain itu,
bukti menunjukkan bahwa kedua reseptor memiliki konformasi yang
berbeda dalam sel sehingga PR-A dan PR-B berinteraksi dengan
koregulator yang berbeda. 28,43-45
Rasio dari isoform individu bervariasi dalam jaringan reproduksi
sebagai akibat dari perkembangan dan status hormonal dan selama
karsinogenesis. Pengikatan progesteron untuk PR-B menghasilkan
rangsangan dari kegiatan proliferasi di dalam rahim; Namun, ekspresi
rahim dari isoform PR-A diperlukan untuk menginhibisi proliferasi
progesteron yang diinduksi melalui fungsi PR-B. Stimulasi PR-A dengan
progesteron juga telah dilaporkan untuk mempromosikan diferensiasi
sel-sel. Selamaovulasi in vivo, hilangnya aktivitas proliferasi tampaknya terkait
erat dengan regulasi LH dan fungsi sel cumulus. Fungsi sel cumulus
dibedakan, seperti produksi progesteron, sintesis Hyaluronan, atau
penutupan komunikasi gap junction, yang terlibat dalam pematangan
meiosis oosit kumulus tertutup. Dengan demikian, rasio tinggi PR-A untuk
ekspresi PR-B berperan penting dalam diferensiasi sel cumulus.46-48
Berbeda dengan ekspresi dominan dari salah satu isoform PR
sering diamati dalam jaringan hewan, dalam jaringan manusia normal in
27
memiliki tingkat PR-A dan PR-B yang sama. Penelitian Aupperlee dkk.
(2007) menunjukkan bahwa PR-A dan PR-B merupakan regulator
transkripsi fungsional yang unik, mampu mengatur transkripsi gen secara
berbeda dalam konteks promotor yang sama, dan mampu mengenali
promotor yang sama sekali berbeda.25
Conneely dkk. (2000), melakukan penelitian kultur jaringan yang
menunjukkan bahwa PR-A dan PR-B memiliki bahan transaktivasi
berbeda secara spesifik terhadap tipe sel dan promoternya. PR-B
ditemukan berfungsi sebagai aktivator transkipsi kuat dari beberapa target
gen PR yang diregulasi oleh kedua reseptor. Bila dalam 1 sel ada 2
isoform ini, PR-A yang meningkat dapat menekan PR-B dan sebaliknya.
Mekanisme ini dilakukan melalui terminal N dan komunikasi intramolekular
PR-A.26,27
Banyak ligan untuk PR telah disintesis dan digunakan dalam
pengobatan klinis dan penelitian. Ligan agonis maupun antagonis
berinteraksi dengan PR untuk mengaktifkan atau menekan ekspresi gen
dalam sel target. Agonis progesteron seperti progestin,
medroksiprogesteron asetat, noretindron asetat, megestrol asetat memiliki
kemampuan untuk menekan proliferasi endometrium yang diinduksi
estrogen. Sedangkan antiprogestin seperti onapriston, mifepriston,
28 2.8 Peran Reseptor Progesteron pada Endometriosis
Uterus terdiri dari tipe sel heterogen dengan gelombang sinkron
dari proliferasi dan diferensiasi sebagai respon terhadap perubahan siklik
pada kadar estrogen dan progesteron. Gelombang hormonal ini juga
mengatur rekruitmen sel inflamasi, apoptosis, hancurnya jaringan, dan
regenerasi. Hormon progesteron ditemukan berperan dalam fungsi
proliferasi atau memicu diferensiasi terutama pada epitel. Reseptor
progesteron yang teraktivasi juga memainkan peran penting dalam
remodelling jaringan uterus. 50-53
Endometrium adalah dinding uterus yang terus tumbuh, proliferasi,
regresi, dan mengulang siklus kembali dalam regulasi hormon steroid.
Diyakini bahwa hormon progesteron berada dalam kadar abnormal pada
kompartemen endometrium eutopik dan ektopik pada wanita dengan
endometriosis, tidak jelas apakah ini merupakan efek langsung, kausal
berkaitan dengan infertilitas, atau dapat digunakan untuk tujuan
diagnostik.42
Telah diketahui dengan jelas bahwa ada pengaruh estrogen pada
endometriosis. Namun, peran progesteron pada endometriosis belum
dipahami dengan jelas karena progesteron ditemukan protektif terhadap
kanker endometrium dan hanya sedikit pasien endometriosis yang
menerima manfaat dari terapi progestin. Walaupun begitu, peran
progesteron terus diteliti karena estrogen dan progesteron bersama-sama
berperan dalam proliferasi sel stroma endometrium.7,22,42
29
memiliki komplemen unik terhadap reseptor hormon steroid dibandingkan
dengan endometrium eutopik. Sebagai contoh, beberapa peneliti
melaporkan kadar ERβ yang tinggi dan kadar ERα yang rendah pada
jaringan endometriosis manusia dan sel-sel stroma primer jika
dibandingkan dengan jaringan dan sel endometrium eutopik.24,25
PR adalah gen target prototipe dari estrogen receptor (ERα) di
beberapa tipe sel termasuk sel epitel keganasan payudara. ERα
memediasi induksi PR oleh estradiol (E2).52,54Defisiensi ERα pada endometriosis dapat menyebabkan kegagalan E2 untuk menginduksi
ekspresi PR, mengakibatkan defisiensi PR sekunder dan resistensi
progesteron pada wanita dengan endometriosis. Observasi in vivo
menunjukkan bahwa E2 menginduksi ekspresi ERα pada jaringan uterus
tikus.30 Sangat mungkin bahwa E2 memiliki peran dalam meregulasi
ekspresi ERα pada sel stroma endometrium manusia. Tetapi, kuantitas besar E2 yang diproduksi melalui aktivitas aromatase lokal, ditambah
kadar ERβ yang tinggi pada sel stroma endometriosis, dapat mengganggu
regulasi dan menekan ekspresi ERα. Kadar ERβ yang tinggi menekan
ekspresi ERα dan respon terhadap estradiol di sel stroma endometriosis melalui ikatan dengan DNA nonklasik di promotor ERα yang digunakan bergantian. ERβ juga meregulasi proses siklus sel dan dapat berkontribusi
terhadap proliferasi sel-sel stroma endometriosis.55-58
Walaupun terdapat fakta bahwa kedua sekuens promoter PR yang
diajukan menunjukkan respon terhadap E2, keduanya tidak memiliki
30
di promoter PR manusia telah dilaporkan. Beberapa lokasi telah
menunjukkan ikatan ERα dan ERβ.28,41-49 Kadar kritis ERα perlu untuk
induksi PR yang tergantung E2 di sel-sel stroma endometrium.
Penempatan daerah promoter PR dengan berbagai rasio ERα dengan
ERβ mungkin diperlukan untuk menentukan efek E2 pada ekspresi PR.
Rasio ERα dan ERβ yang sangat rendah pada sel-sel stroma
endometriosis mungkin bertanggung jawab untuk pergeseran dari
stimulasi E2 menjadi inhibisi E2 terhadap ekspresi PR di sel-sel stroma
endometriosis.52-55
PR adalah salah satu gen yang responsif terhadap E2, dan E2
bekerja di jaringan endometrium eutopik dan sel stroma untuk
meningkatkan tingkat respon endometrium terhadap
progesteron.28 Sebagai kontras, mRNA dan kadar protein PR tidak
meningkat pada jaringan endometriosis yang dibiopsi setelah terekspos
kadar E2 tinggi saat fase proliferatif akhir ataupun pada jaringan
endometriosis yang telah diterapi dengan E2. Hal ini mengindikasikan
bahwa ekspresi PR yang diinduksi E2 mengalami gangguan pada
endometriosis.26
Pada penelitian terbaru mengenai kadar reseptor steroid pada sel
endometrium dan endometriosis, kadar mRNA ERα lebih rendah secara signifikan (tujuh kali lipat) pada sel-sel stroma endometriosis dibandingkan
sel stroma endometrium. Kadar mRNA ERβ lebih tinggi secara signifikan
(34 kali lipat) pada sel-sel stroma endometriosis, sedangkan kadarnya di
31
dijumpai. Rerata rasio ERα terhadap ERβ adalah 841 dan 21 pada sel-sel stroma endometriosis. Kadar mRNA total PR and PR-B pada sel stroma
endometriosis lebih rendah secara signifikan jika dibandingkan sel stroma
endometrium. Kadar protein ERα dan ERβ berbeda secara signifikan pada kedua kelompok, serupa dengan temuan terkait kadar mRNA. Sel-sel
stroma endometriosis mengandung ERβ yang amat tinggi dan ERα dan
PR yang lebih rendah secara signifikan jka dibandingkan dengan sel-sel
stroma endometrium.52,54
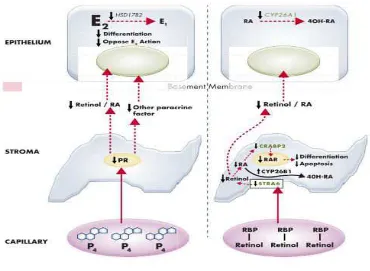
Di endometrium, mekanisme progesteron dilakukan melalui ikatan
pada PR di sel stroma. Pembuluh darah yang mengangkut progesteron
(P4) terletak dekat ke sel stroma. Kontak awal antara progesteron dan
aktivasi PR dimediasi oleh koaktivator reseptor steroid (SRC).35
Koaktivator akan berinteraksi dengan reseptor DBD dan LBD dengan
bantuan SNURF, GT198, dan HMG. DBD PR diperlukan untuk mengikat
urutan DNA elemen responsif tertentu, tapi lebih sedikit yang diketahui
tentang fungsi koaktivaktor nuklir yang mengikat DBD.40,56
PR stroma yang diaktivasi P4 di sirkulasi akan memproduksi
banyak faktor parakrin termasuk asam retinoat. Masuknya asam retinoat
dimediasi oleh uptake reseptor RBP-retinol dari sirkulasi. Retinol
dikonversi ke asam retinoat, yang kemudian ditransportasi oleh cellular
RA binding protein-2 (CRABP2) ke nukelus. RA-RAR akan menentukan
keseimbangan diferensiasi dan apoptosis pada sel stroma endometrium.
PR diketahui memicu STRA6 dan CRABP2 stroma serta menganggu
32
mediator parakrin lainnya kemudian akan memicu diferensiasi sel epitel
dan menghambat mekanisme proliferasi sel yang dipicu estradiol (E2).
Implantasi endometriosis berkaitan dengan aromatase dan 17β
-hydroxysteroid dehydrogenase tipe 1, yaitu suatu enzim yang merubah
androstenedione menjadi estron dan estron menjadi estradiol. Pada
keadaan ini didapati penurunan dari 17β-hydroxysteroid dehydrogenase
tipe 2 yang menonaktifkan estrogen.10-15 Asam retinoat dapat memicu
enzim 17βHSD2, yang mengubah E2 aktif ke estron yang lemah (E1) di sel epitel. Peningkatan PR akan meningkatkan proses ini dan
sebaliknya.37 Ekspresi dari PR dalam epitel, stroma dan kompartemen
seluler miometrium berada di bawah kendali estrogen dan progesteron.
Estrogen merangsang dan progesteron menghambat pertumbuhan
endometrium. Selain berikatan langsung dengan reseptor estrogen atau
progesteron, ada bukti kuat bahwa terdapat interaksi parakrin antara epitel
dan stroma sel di endometrium menjadi mediator resepon terhadap
33
Gambar 4. Mekanisme kerja reseptor progesteron59
Dalam endometrium manusia, tingkat ekspresi isoform PR diatur
secara berbeda selama siklus menstruasi yang normal. Profil ekspresi gen
dalam pemeriksaan microarray di endometrium wanita dengan
endometriosis menunjukkan bahwa sejumlah progesteron gen disregulasi
selama masa implantasi seperti glukodelin yang lebih rendah pada
jaringan endometriosis.56 Sebagai contoh, PR-A adalah isoform PR
dominan dan tetap dalam stroma, tapi ekspresi kedua isoform PR
menurun dalam sel epitel endometrium selama fase sekresi. Penelitian
Jericevic dkk. (2004) pada mencit dengan delesi PR-A/B menunjukkan
34
Penelitian Mangal dkk. (1997) menunjukkan 17β-estradiol menginduksi ekspresi PR-B lebih dari ekspresi PR-A di endometrium
manusia. Namun, belum ditetapkan mengapa ekspresi penurunan subtipe
PR terjadi pada endometriosis tergantung estrogen. Pada penelitian lain,
Mote dkk. dengan imunohistokimia menunjukkan bahwa ekspresi PR-A
dan PR-B meningkat selama fase proliferasi dan mencapai ekspresi
tertinggi selama fase akhir proliferasi. Kedua penelitian ini menunjukkan
bahwa ekspresi isoform PR bergantung hormonal dapat menentukan
inhibisi atau stimulasi kerja progestogen.40,41
Beberapa penelitian menunjukkan hasil yang kontroversial. Kao
dkk. (2003) melakukan pemeriksaan microarray pada sel dari biopsi
jaringan endometrium ektopik wanita endometriosis dan normal pada hari
8-10 setelah lonjakan LH pertengahan siklus. Burney dkk. (2007) yang
membandingkan ekspresi gen ektopik dan eutopik endometriosis pada
manusia menemukan penurunan respon progesteron normal. Berbeda
dengan penelitian, D’Amora dkk. (2009) menunjukkan bahwa terdapat peningkatan polimorfisme PROGINS (salah satu varian gen PR) pada
wanita endometriosis.3,9,14
Perbedaan ini dapat disebabkan karena PR memiliki dua isoform.
Pada keadaan patologis pada endometrium, kadar kedua isoform
biasanya tidak seimbang. Oleh karena itu, peneliti terus berupaya mencari
proses ketidakseimbangan kedua isoform PR. Terdapat variasi penelitian
walaupun kebanyakan penelitian menunjukkan downregulasi PR-B, sesuai
35
dan translasi sel.3,9 Terlebih lagi, kadar kedua isoform PR, terutama PR-B,
lebih rendah secara signifikan pada endometriosis jika dibandingkan
dengan endometrium eutopik.5,26
Penelitian Igarashi dkk. (2005) dilakukan untuk mengetahui
ekspresi PR-A dan PR-B di endometrium. Sampel adalah jaringan biopsi
endometrium ektopik dari seluruh sukarelawan wanita penderita
endometriosis yang telah menjalani pembedahan. Dilakukan analisis in
vivo ekspresi PR-A dan PR-B di endometrium dari wanita dengan
endometriosis dan kontrol. Hasil penelitian adalah ditemukan rasio
PR-B/PR-A lebih rendah pada jaringan endometrium dari wanita dengan
endometriosis dibandingkan dengan jaringan normal.10
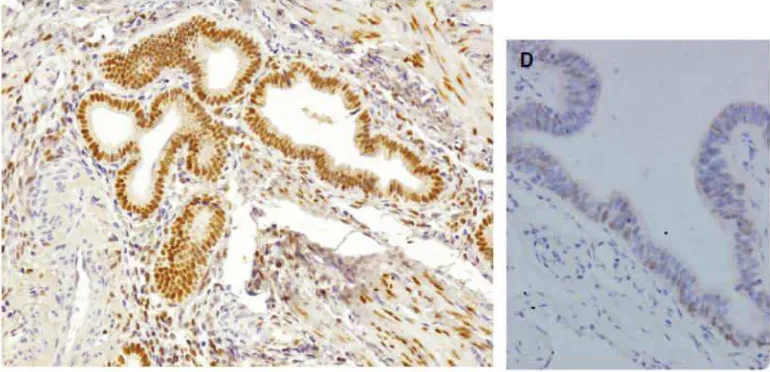
Attia dkk. (2000) melakukan penelitian untuk menentukan bahwa
aksi progesteron pada target gen endometriosis dilakukan oleh
homodimer PR-B, di mana PR-A berperan sebagai supresor fungsi PR-B.
Pada 18 jaringan endometrium normal, PR-B ditemukan pada 17 sampel
dengan kadar yang meningkat. PR-A ditemukan pada seluruh jaringan
endometriosis namun dalam kadar yang rendah. Pada 18 jaringan
endometriosis, tidak ditemukan PR-B namun ditemukan PR-A dalam
kadar yang rendah. Tidak adanya PR-B pada jaringan endometrium dapat
menjadi gambaran atas rendahnya rasio kelenjar/stroma dibandingkan
endometrium normal.43,56,60-63
Treloar dkk. (2005) pada 900 wanita tidak menemukan hubungan
antara endometriosis dengan peningkatan PR-B. Wu dkk. (2006) dan
36
ektopik, terjadi downregulasi PR-B, PR-A tetap, akibat hipermetilasi PR-B.
Selain itu, ditemukan bahwa PR-B menurun karena represi oleh reseptor
estrogen yang sangat tinggi dalam sel stroma.44-46
Akan tetapi, Fazleabas et al. (2003) menemukan tidak ada
perbedaan signfikan antara rasio PR-B/PR-A pada jaringan endometriosis
dibandingkan jaringan normal. Bahkan, Lee dkk. (2009) menunjukkan ada
peningkatan rasio PR-B/PR-A dan penurunan mRNA PR-A pada jaringan
endometrium ektopik model endometriosis mencit dibandingkan kontrol.
Wu dkk. (2006) telah menunjukkan bukti penurunan enzim 17HSD2 pada
peningkatan PR terutama PR-B dalam jaringan endometriosis. Berchuck
dkk. (2004) pertama sekali menyatakan bahwa penurunan risiko
endometriosis oleh karena adanya peningkatan PR-B.56,60,64
Berbagai penelitian juga menemukan adanya resistensi
progesteron pada endometriosis. Sel stroma endometriosis yang gagal
merespon progesteron akan mengalami penurunan rangsangan asam
retinoat. Hal ini juga akan menyebabkan turunnya enzim 17BetaHSD2 dan
kegagalan inaktivasi estradiol menjadi estron. Selain itu, juga ditemukan
berbagai defek pada jalur asam retinoat, kombinasi dengan tingkat
estradiol yang tinggi, defek progesteron ini akan lebih mempromosikan
endometriosis.49,64-67
Penelitian Attia dkk. (2000) menunjukkan bahwa resistensi
progesteron dapat dijelaskan akibat tidak ada satupun transkrip dari
protein PR-B dan adanya PR-A pada lesi ektopik. Resistensi progesteron
37
inflamasi kronik, tetapi juga akibat perubahan epigenetik kromatin yang
menentukan responsitivitas sel endometrial.43,56-58
Peneliti lain juga menunjukkan penurunan risiko endometriosis
karena peningkatan produksi PR-B. Semua hal ini berkontribusi dalam
patogenesis endometriosis.56-60
38 2.9 Kerangka Teori
Kolesterol Estradiol Estron
ENDOMETRIOSIS
Aromatase 17βHSD2
17βHSD1
Stroma Endometrium RANTES MCP-1 L-Selektin
39 2.10 Kerangka Konsep
VARIABEL INDEPENDEN VARIABEL DEPENDEN
Ekspresi reseptor