Study of Enzymatic Hydrolysis of Rice for Hydrogen
Production Using Mixed Cellulases
Nadiem Anwar, Arief Widjaya, Sugeng Winardi
2Jurusan Teknik Kimia-Fakultas Teknologi Industri-Institut Teknologi Sepuluh Nopember ITS Surabaya Indonesia 60111, email: [email protected]
Abstract
The objective of this work is to investigate the factors that affect the hydrolysis of rice straw, and compare the efectiveness of commercial enzyme with mixed crude enzyme from Trichoderma reesei and Aspergillus niger. The work is divided into two steps, the first is cellulase productions from rice and corn straw, and the second is enzymatic hydrolysis. Two activity unit of cellulase from
Trichoderma reesei and one unit of cellulase from A. niger were mixed. Enzymes were prepared by
solid fermentation. Five grams of 100 mesh rice straw were mixed with 25 mL nutrient solution containing (all in g/L except CMC) 1,4 (NH4)2SO4, 2 KH2PO4, 0,005 FeSO4, 0,5 ekstrak ragi, 1,5
pepton dan 5 mL 1% CMC solution. T. reesei dan A. niger were inoculated and then iccubated for 4, 6 dan 8 days. Before hydrolized, the rice straw is grinded and sieved to 80 – 100 and 120 – 140 mesh then heated at 85 oC with 2% sodium hydroxide for six hours. Hydrolyses was conducted in 300 mL beaker glass equipped by mechanical stirrer. Samples were analyzed by dinitrocalycilic acid and measured by spectrophotometer.
According to the study of enzyme production, it can be concluded that cellulases enzyme activity of rice straw is higher than corn straw, either using T. reesei strain or A. niger. The quality of T. reesei cellulase of rice straw is as good as A. niger cellulase of straw. After being incubated for four days with rice straw substrate, T. reesei gives higher enzyme productivity than A. niger.
Based on the study of hydrolysis, it can be concluded that the higher the enzyme ratio to substrate was, the higher the glucose concentration obtained, but enzyme eficiency become smaller. Straw particle for 120 – 140 mesh is optimum enough to be chosen for rice straw hydrolisys. Decreacing pH from 5,52 to 3,92 will decrease the time of optimum hydrolysis from 6 to 3 hours. For a smaller scale hydrolysis, the change of speed of stirring from 160 rpm to 250 rpm did not give a significant effect to glucose concentration obtained. Increasing hydrolysis temperature from 40 oC to 46 oC increased the glucose production to 15%. The highest glucose concentration that can be made by hydrolysis using cellulose enzyme from A. niger produced by Fluka Biochemika was obtained at temperature 46 oC, propeller speed of 160 rpm, enzyme ratio of 28 U/g straw, pH 3,92 and straw particle 120 – 140 mesh. Enzyme from T. reesesi dan A. niger that produced from mixture of Trichoderma reesei and Aspergillus niger with a ratio of 2 : 1 (unit/unit) was more effective to hydrolyze rice straw than using cellulose enzyme of Aspergillus niger from Fluka Biochemika production.
Keywords: cellulase, Trichoderma reesei, Aspergillus niger, rice straw, mixed enzymes, hydrolysis
I. PENDAHULUAN
Kebutuhan energi untuk menunjang kehidupan manusia meningkat terus secara eksponensial (Antonopoulou dkk., 2007), sementara persediaan bahan bakar fosil yang sampai saat ini merupakan sumber utama energi semakin menurun. Penggunaan bahan bakar fosil juga telah menurunkan kualitas lingkungan, oleh karena itu perlu dicarikan bahan bakar alternatif, salah satunya yang potensial adalah biohidrogen karena kandungan energinya tinggi, hampir tiga kali gasolin (Bossel, 2003). Pembakaran hidrogen juga hanya menghasilkan uap air sehingga hidrogen merupakan sumber energi yang ramah lingkungan (Ogino dkk., 2005). Hidrogen dapat diproduksi melalui sintesa kimia maupun secara biologis. Produksi hidrogen secara biologis memiliki kelebihan dibandingkan dengan sintesa kimia, karena dapat dilangsungkan pada suhu dan tekanan rendah sehingga lebih menguntungkan ditinjau dari sisi penggunaan energi.
Biohidrogen dapat diproduksi dari glukosa dengan perolehan berkisar 1,36 – 3,02 mol H2/mol glukosa (Pallazi
sukrosa dengan perolehan 1,3 – 4,8 mol H2/mol sukrosa yang setara 0,65 – 2,4 mol H2/mol glukosa (Sung dkk.,
2002; Lin dan Lay, 2004; Ogino dkk., 2005), dari pati ubi kayu dengan perolehan 1,1 – 1,5 mol H2/mol heksosa
(Mahakhan dkk., 2005; Lin dkk., 2008). Perolehan biohidrogen tertinggi dari glukosa murni sekitar 75 % perolehan teoritis, akan tetapi masih belum fisibel secara komersial karena harga bahan baku glukosa murni yang tinggi (Kapdan dan Kargi, 2006). Upaya meningkatkan fisibilitas ekonomi biohidrogen dimungkinkan dengan penggunaan bahan baku limbah yang dapat dikonversi menjadi glukosa, baik limbah cair (Reungsang dkk., 2004) maupun limbah padat (Raghavendra dkk., 2007). Salah satu limbah di Indonesia yang ketersediaannya melimpah adalah jerami padi yaitu 180 juta ton/tahun (Sabiham dan Mulyanto, 2005).
Jerami padi adalah limbah yang mengandung 37,7% selulosa; 22,0% hemiselulosa; 16,6% lignin (Dewi, 2002). Selulosa dalam jerami padi dapat dihidrolisis menghasilkan glukosa baik secara kimiawi (Xiang dkk., 2003), secara enzimatis (Dewi, 2002; de Vrije dkk., 2002; Raghavendra dkk., 2007) maupun fermentatif (Aderemi dkk., 2008). Hidrolisis enzimatik lebih menarik dari sisi penggunaan energi, karena dapat dilangsungkan pada temperatur rendah (de Vrije dkk., 2002) sedangkan hidrolisis kimiawi memerlukan temperatur tinggi (Xiang dkk., 2003). Hidrolisis enzimatis juga dapat menghasilkan konsentrasi glukosa yang lebih tinggi dibandingkan dengan hidrolisis fermentatif (Aderimi dkk., 2008), akan tetapi reaksi enzimatik memerlukan waktu yang lebih lama. Oleh karena itu perlu dilakukan upaya untuk mempercepat reaksi diantaranya dengan cara memperbesar rasio enszim dengan substrat.
Uraian di atas menunjukkan kemungkinkan memproduksi biohidrogen dari jerami padi melalui dua tahap proses terpisah yaitu hidrolisis enzimatik jerami padi menjadi glukosa yang dilanjutkan dengan fermentasi glukosa menjadi hidrogen. Penelitian ini bertujuan untuk mempelajari pengaruh berbagai kondisi operasi (temperatur, konsentrasi awal enzim, pH dan laju pengadukan) serta membandingkan efektivitas enzim selulase yang dibuat pada penelitian ini dengan selulase komersial.
II. DASAR TEORI
A. Jerami Padi
Jerami padi terdiri 37,7% selulosa, 22,0% hemiselulosa dan 16,6% lignin (Dewi, 2002) dengan kandungan nutrisi dalam basis kering 0,5 – 0,8% N; 0,16 – 0,27% P2O5; 1,4 – 2% K2O; 0,05 – 0,1% S dan 4 – 7% Si
(Dobermann dan Fairhurst, 2002). Selulosa berfungsi sebagai penguat pada batang tumbuhan, lignin untuk melindungi selulosa dari aksi kimiawi maupun biologis sedangkan hemiselulosa pengikat selulosa dengan lignin (Lee, 1992). Selulosa berupa polimer glukosa linier hidrofilik yang dihubungkan oleh ikatan glikosida. Banyaknya satuan glukosa dapat bervariasi antara 15 sampai lebih dari 10.000 per molekul selulosa. Polimer selulosa terdiri dari bagian kristalin dan amorf. Bagian amorf mudah dihidrolisis sedangkan bagian kristalin tidak mudah dihidrolisis baik secara kimiawi maupun enzimatik (Dahot dan Noomrio, 1996). Hemiselulosa adalah gabungan dari polimer-polimer dengan rantai relatif lebih pendek dan bercabang, yang terdiri dari monomer xylosa, arabinosa, glukosa, manosa dan galaktosa, dengan struktur amorf (Bailey dan Ollis, 1986). Meskipun dilindungi oleh lignin, jerami padi masih dapat dihidrolisis menjadi glukosa oleh mikroorganisma selulotik seperti Trichoderma ressei (Muthuvelayudham dan Viruthagiri, 2006) maupun Aspergillus niger (Aderemi dkk., 2008).
B. Selulosa
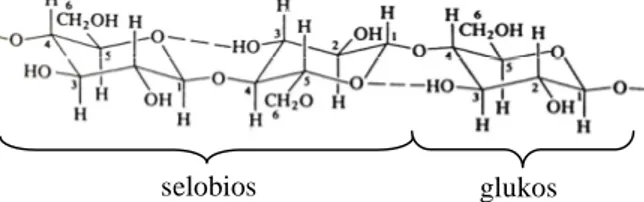
Selulosa adalah homopolisakarida yang tersusun atas satuan-satuan anhidroglukopiranosa yang berikatan dengan ikatan glikosida-β-(1,4) membentuk rantai molekul linier glukan. Selulosa secara empiris dapat ditulis sebagai (C6H10O5)n dengan n adalah derajat polimerisasinya yang menyatakan jumlah satuan glukosa yang
berikatan. Derajat polimerisasi selulosa tumbuhan berada pada kisaran 305 – 15300 (Widjaja, 2009). Kedudukan β dari gugus OH pada atom C nomor 1 (C1) membutuhkan pemutaran unit glukosa berikutnya
melalui sumbu C1 – C4 cincin piranosa. Satuan ulang dari rantai selulosa adalah satuan selobiosa dengan
panjang 1,03 nm.
Gambar 2.1 Struktur molekul selulosa
C. Glukosa
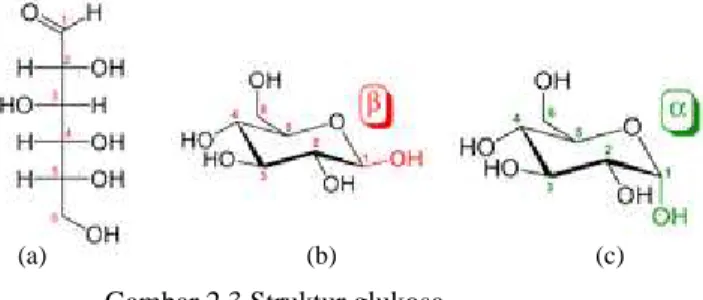
Glukosa adalah monomer penyusun selulosa yang dikenal sebagai gula darah. Glukosa ini merupakan sumber energi dalam makhluk hidup dan merupakan produk antara metabolisme. Glukosa merupakan karbohidrat produk utama fotosintesis yang penting. Glukosa dalam hewan dan fungi dihasilkan melalui proses pemecahan glikogen dan dalam tanaman dihasilkan berasal dari pemecahan pati. Glukosa dalam makhluk hidup umumnya adalah D-glukosa yang dalam larutan membentuk struktur ring piranosa yang terjadi karena ikatan antara aldehid C1
dengan hidroksil pada C5 seprti pada Gambar 2.3a. D(+)-glukosa adalah isomer optik glukosa yang memutar
bidang polarisasi ke arah (+) Keberadaan gugus-gugus OH yang terdapat pada ujung-ujung rantai glukosa menentukan sifat glukosa. Demikian pula posisi α dan β seperti yang dapat diihat pada Gambar 2.3b dan c, akan menentukan sifat senyawa yang dihasilkannya.
(a) (b) (c)
Gambar 2.3 Struktur glukosa
D. Enzim Selulase
Enzim adalah biokatalis yang diproduksi oleh jaringan hidup untuk mengkatalisis reaksi-reaksi yang terjadi dalam jaringan. Bila tidak ada enzim, reaksi-reaksi dalam jaringan akan berjalan terlalu lambat sehingga tidak dapat menopang kehidupan atau reaksi-reaksi tersebut memerlukan kondisi non fisiologis (Montgomery, 1993).
Selulase adalah campuran beberapa enzim yang komposisinya bervariasi, bergantung kepada mikroorganisme yang digunakan untuk memproduksi serta proses produksinya. Tiga komponen yang telah teridentifikasi dalam selulase adalah endoglukanase (endo-β-1,4-D-glukan-4-glukanohidrolase, E.C.3.2.1.4) yang memecah ikatan β -1,4 pada rantai selulosa secara acak, eksoglukanase (β-1,4-D-glukan-selobiohidrolase, E.C. 3.2.1.91) yang memecahkan satuan selobiosa dari ujung rantai dan β-glukosidase (E.C. 3.2.1.21) yang memecahkan selobiosa menjadi glukosa (Dahot dan Noomrio,1996).
Enzim selulase dapat dihasilkan oleh berbagai jenis mikroorganisme, tetapi hanya sedikit yang dapat menghasilkan selulase dalam jumlah yang cukup untuk menghidrolisis seluruh selulosa kristalin. Trichoderma
reesei telah dikenal mampu menghasilkan endoglukanase sampai 80% (Muthuvelayudham dan Viruthagiri, 2006),
tetapi β-glukosidasenya rendah (Martins dkk., 2008).
Kebanyakan sistem selulase yang dihasilkan oleh jamur selulotik, jumlah β-glukosidasenya lebih rendah dari yang dibutuhkan untuk hidrolisis selulosa menjadi glukosa secara efisien, sehingga produk utama hidrolisisnya bukan glukosa melainkan selobiosa (Juhasz dkk., 2003; Martins dkk., 2008; Ahamed dan Vermette, 2008), yang merupakan inhibitor kuat terhadap endo dan eksoglukanase. Persoalan ini dapat diatasi dengan menambahkan β -glukosidase dari luar (Juhasz dkk., 2003) atau memproduksi selulase dengan cara mengkombinasikan mikroorganisme yang kemampuan memproduksi endo dan eksoglukanasenya kuat seperti Trichoderma reesei dengan mikroorganisme yang kemampuan memproduksi β-glukosidasenya kuat seperti Aspergillus niger (Juhasz dkk., 2003).
E. Hidrolisis Selulosa Secara Enzimatik
Hidrolisis selulosa secara biologik dapat dilakukan baik menggunakan enzim selulase (Vrije dkk., 2002; Raghavendra dkk., 2007) maupun mikroorganisme penghasil selulase (Aderemi dkk., 2008). Hidrolisis selulosa dipengaruhi oleh jenis sumber subsrat (seperti serbuk gergaji, jerami padi, sabut sawit) dan ukuran partikel. Jerami padi lebih sulit dihidrolisis dibandingkan dengan serbuk gergaji maupun sabut sawit (Dewi, 2002). Raghavendra dkk. (2007) melaporkan bahwa diantara jerami padi, jerami gandum dan bagas tebu, yang paling mudah dihidrolisis adalah jerami gandum, diikuti bagas dan yang paling sulit dihidrolisis adalah jerami padi. Kesulitan hidrolisis substrat lignoselulosa karena adanya gangguan dari lignin yang menghambat kontak antara selulase dengan selulosa (Raghavendra dkk., 2007). Kesulitan tersebut dapat diperbaiki dengan cara melakukan pengolahan awal (Bak dkk., 2008; Vrije dkk., 2002; Dahot dan Noomrio, 1996).
Vrije dkk. (2002) melaporkan bahwa perolehan glukosa dari hidrolisis miscanthus tanpa perlakuan NaOH lebih kecil dari 20%, perolehan dari hasil penggilingan yang diberi perlakuan NaOH 12% pada suhu 70 oC selama 4 jam adalah 56% dan perolehan dari hasil ekstrusi yang diberi perlakuan NaOH adalah 71%. Bak dkk. (2008) melaporkan bahwa electron beam irradiation pretreatment meningkatkan perolehan glukosa dari jerami padi dari 5,1 – 22,6% menjadi 43,1 – 52,1%. Öhgren dkk. (2007) melaporkan bahwa perlakuan panas terhadap
corn stover dapat meningkatkan perolehan glukosa.
Mekanisme hidrolisis enzimatik selulosa menjadi glukosa adalah rumit seperti yang diperlihatkan pada Gambar 2.1, tetapi Aderemi dkk. (2008) melaporkan bahwa hidrolisis jerami padi mengikuti mekanisme reaksi enzimatik Michaelis-Menten:
S S S max , g g C K C r r + =
(1)
dengan CS adalah konsentrasi substrat, rg laju produksi glukosa, KS kanstanta konstanta penjenuhan, rg,max laju
produksi glukosa maksimum. Nilai rg,max adalah 1,5288 × 10-4 g/L.detik dan KS = 33,7 g/L, ditentukan
menggunkan metoda Lineweaver-Burk.
Li dkk. (2004) yang meneliti hidrolisis berbagai pulp, mengusulkan model kinetika hidrolisis enzimatik yang menunjukkan adanya inhibisi dari komponen lain selain substrat. Proses hidrolisis yang kompleks disederhanakan dan dianggap terdiri dari dua tahap reaksi seperti berikut:
(2)
Tahap pertama adalah reaksi heterogen antara substrat tidak larut S dengan enzim terlarut menghasilkan oligosakharida yang larut (O) oleh aksi sinergi eksoglukanase CBH dan dan endoglukanase EG yang dianggap sebagai pengendali hidrolisis keseluruhan. Tahap kedua adalah reaksi homogen pemecahan oligosakharida menjadi produk akhir glukosa (G) yang dikatalisis oleh βG dan mencapai kesetimbangan karena lebih cepat dari reaksi tahap pertama. Model kinetika yang diusulkan Li dkk. (2004) adalah sebagai berikut:
) S S ( 9 , 0 ] S ) K / 1 ( 1 [ K ) S S ( E k dt dS I M o − + + − = ∞ ∞
(3)
Konstanta kesetimbangan reaksi kedua pada (2): G S G O G K − = = (4)
dengan S adalah konsentrasi substrat, S∞ konsentrasi gula total akhir rata-rata, Eo konsentrasi enzim, G
konsentrasi glukosa, O konsentrasi oligosakharida, k konstanta laju, KM konstanta Michaelis-Menten dan KI
konstanta inhibisi kompetitive
III. METODE
Jerami padi diperoleh dari sekitar kampus ITS, dijemur 4 hari, dipotong kurang lebih 2 mm menggunakan pemotong kertas, digiling menggunakan penggiling biji-bijian kemudian diayak 80 – 100 mesh dan 120 – 140 mesh. Jerami kemudian didelignifikasi menggunakan NaOH 2% pada temperatur 85 oC selama 6 jam. Enzim yang digunakan adalah enzim komersial produk Fluka Biochemika. Lima gram serbuk jerami yang telah didelignifikasi dicampur dalam beaker glass 300 mL kemudian diaduk dengan kecepatan 160 rpm, temperatur hidrolisis 40 oC, pH 5,52; 4,75 dan 3,92. Ratio Enzim dengan jerami divariasikan 14, 18, 23 dan 28 U/g serta diameter partikel jerami 80 – 100 dan 120 – 140 mesh. Jumlah glukosa yang dihasilkan setiap selang waktu tertentu dianalisis menggunakan dinitrosalicylic acid (DNS), diukur menggunakan spektrofotometer pada panjang gelombang 540 nm.
IV. HASIL PENELITIAN
A. Pengaruh Rasio Enzim-Jerami Padi
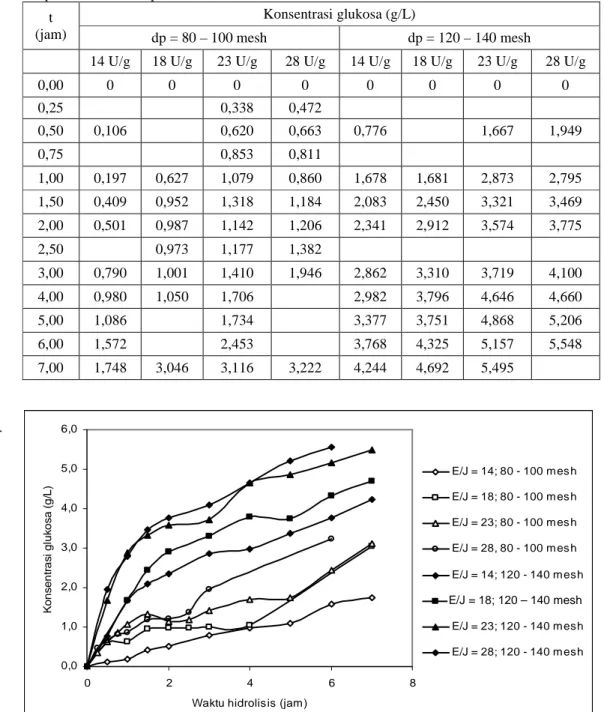
Sampel diambil 2 mL setiap sekali sampling untuk meminimalkan pengaruh pengurangan volume. Percobaan maksimal dilakukan selama tujuh jam karena lebih dari itu terbentuk campuran yang sulit diendapkan sehingga pengambilan sampel menjadi sulit pula. Hasilnya diperlihatkan pada Tabel 1 dan Gambar 1.
EG/CBH Oligo- βG sakarida (O) Selulosa (S) Glukosa (G)
0,0 1,0 2,0 3,0 4,0 5,0 6,0 0 2 4 6 8
Waktu hidrolis is (jam)
K o n s e n tr a s i g lu k o s a ( g /L ) E/J = 14; 80 - 100 mes h E/J = 18; 80 - 100 mes h E/J = 23; 80 - 100 mes h E/J = 28, 80 - 100 mes h E/J = 14; 120 - 140 mesh E/J = 23; 120 - 140 mesh E/J = 23; 120 - 140 mesh E/J = 28; 120 - 140 mesh E/J = 18; 120 – 140 mesh Tabel 1 Konsentrasi gula terhadap waktu untuk berbagai rasio enzim - jerami, pada 160 rpm, temperatur 40 oC dan pH 5,52.
t (jam)
Konsentrasi glukosa (g/L)
dp = 80 – 100 mesh dp = 120 – 140 mesh
14 U/g 18 U/g 23 U/g 28 U/g 14 U/g 18 U/g 23 U/g 28 U/g
0,00 0 0 0 0 0 0 0 0 0,25 0,338 0,472 0,50 0,106 0,620 0,663 0,776 1,667 1,949 0,75 0,853 0,811 1,00 0,197 0,627 1,079 0,860 1,678 1,681 2,873 2,795 1,50 0,409 0,952 1,318 1,184 2,083 2,450 3,321 3,469 2,00 0,501 0,987 1,142 1,206 2,341 2,912 3,574 3,775 2,50 0,973 1,177 1,382 3,00 0,790 1,001 1,410 1,946 2,862 3,310 3,719 4,100 4,00 0,980 1,050 1,706 2,982 3,796 4,646 4,660 5,00 1,086 1,734 3,377 3,751 4,868 5,206 6,00 1,572 2,453 3,768 4,325 5,157 5,548 7,00 1,748 3,046 3,116 3,222 4,244 4,692 5,495 .
Gambar 1 Konsentrasi gula terhadap waktu untuk berbagai rasio enzim terhadap jerami, pada 40 oC, 160 rpm, pH = 5,5
B. Pengaruh Diameter Partikel
Pengaruh diameter partikel terhadap laju hidrolisis yang diperlihatkan pada Tabel 4.3 dan Gambar 4.3, perubahan diameter partikel jerami dari 80 – 100 mesh ke 120 – 140 mesh meningkatkan produksi glukosa yang sangat berarti. Hal ini dimungkinkan karena ukuran partikel jerami makin kecil, makin besar luas permukaan kontak antara enzim dengan jerami. Akan tetapi ukuran ini tidak memungkinkan untuk diperkecil lagi karena kesulitan pada perlakuan awal secara kimiawi yaitu delignifikasi menggunakan NaOH 2%. Perolehan selulosa pada tahap perlakuan awal jerami 120 – 140 mesh ini cukup kecil, yaitu rata-rata 35% karena banyak yang hilang pada saat penyaringan. Perbedaan glukosa yang dihasilkan sangat signifikan, sehingga ukuran partikel 120 – 140 mesh sudah cukup baik.
0,0 0,1 0,1 0,2 0,2 0,3 0,3 0,4 0 2 4 6 8
Waktu hidrolisis (jam)
E fi s ie n s i e n z im ( g /L .U ) E/J = 14; 80 - 100 mesh E/J = 18; 80 - 100 mesh E/J = 23; 80 - 100 mesh E/J = 28, 80 - 100 mesh E/J = 14; 120 - 140 mesh E/J = 19; 120 - 140 mesh E/J = 23; 120 - 140 mesh E/J = 28; 120 - 140 mesh C. Efisiensi Enzim
Efisiensi enzim yang dimakud di sini adalah jumlah glukosa yang dihasilkan persatuan aktivitas enzim yang digunakan (Cg/E). Tabel 2 dan Gambar 2 menunjukkan bahwa efisiensi enzim tertinggi dihasilkan pada diameter
partikel yang lebih kecil yaitu 120 – 140 mesh. Tabel 2 juga menunjukkan bahwa pada percobaan ini, efisiensi enzim berbanding terbalik dengan rasio enzim substrat. Makin besar rasio enzim substrat makin kecil efisiensinya. Hal ini menunjukkan bahwa peningkatan jumlah enzim yang digunakan tidak sebanding dengan jumlah glukosa yang dihasilkan. Efisiensi tertinggi dihasilkan pada hidrolisis dengan diameter partikel 120 – 140 mesh dan rasio aktivitas enzim 14 U/g jerami. Gambar 1 juga menunjukkan bahwa laju produksi glukosa pada pada rasio enzim-substrat 28 U/g, kurang lebih dua kali laju produksi glukosa pada 14 U/g. Hal ini berarti, jika kita menginginkan waktu hidrolisis yang lebih singkat, dapat dilakukan dengan meningkatkan rasio enmim substrat dengan diameter partikel 120 – 140 mesh.
Tabel 2 Efisiensi enzim terhadap waktu untuk berbagai rasio enzim terhadap jerami, pada 160 rpm, temperatur 40 oC, pH = 5,5.
Gambar 2 Efisiensi enzim terhadap waktu untuk berbagai rasio enzim - jerami, pada 160 rpm, temperature 40 oC dan pH = 5,5.
t (jam)
Efisiensi enzim (g/L.U)
dp = 80 - 100 mesh dp = 120 - 140 mesh 14 U/g 18 U/g 23 U/g 28 U/g 14 U/g 18 U/g 23 U/g 28 U/g
0 0 0 0 0 0 0 0 0 0,25 0,015 0,017 0,5 0,008 0,027 0,024 0,056 0,072 0,070 0,75 0,037 0,029 1,0 0,014 0,034 0,046 0,031 0,120 0,090 0,124 0,100 1,5 0,029 0,051 0,057 0,042 0,149 0,132 0,143 0,124 2,0 0,036 0,053 0,049 0,043 0,168 0,157 0,154 0,135 2,5 0,052 0,051 0,050 3,0 0,057 0,054 0,061 0,070 0,205 0,178 0,160 0,147 4,0 0,070 0,056 0,073 0,214 0,204 0,200 0,167 5,0 0,078 0,075 0,242 0,202 0,209 0,187 6,0 0,113 0,106 0,270 0,233 0,222 0,199 7,0 0,125 0,164 0,134 0,115 0,304 0,252 0,236
0 1 2 3 4 5 6 7 0 1 2 3 4 5 6 7
Waktu hidrolisis (jam)
K o n s e n tr a s i g lu k o s a ( g /L ) pH = 5,52 pH = 4,75 pH = 3,92 D. Pengaruh pH
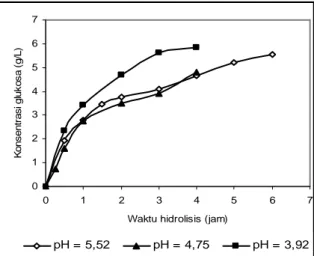
Glukosa yang dihasilkan ditujukan sebagai bahan baku produksi biohidrogen yang diperkirakan berlangsung optimum pada pH 6. Oleh karena itu hidrolisis yang paling baik menurut rencana penggunaan glukosanya adalah pH 6. Akan tetapi pada pH 6, konsentrasi glukosa yang dihasilkan sangat kecil, oleh karena itu dilakukan pengujian pengaruh pH di bawah 6. Tabel 4.5 menunjukkan bahwa penurunan pH dari 5,52 sampai 3,92 dapat mempersingkat waktu hidrolisis dari 6 jam menjadi sekitar 3 jam karena laju pembentukan glukosa 3 jam pertama pada pH 3,92 lebih cepat dari pH 6 meskipun konsentrasi glukosa akhirnya hampir sama, sedangkan pada pH 4,75 tidak jauh berbeda dengan pada pH 5,52.
Tabel 4.5 Konsentrasi glukosa terhadap waktu untuk berbagai pH pada 160 rpm, 120 – 140 mesh, 28 U/g jerami, temperatur 40 oC.
Gambar 4.5 Konsentrasi glukosa terhadap waktu hidrolisis, untuk berbagai rasio E/J, pada 160 rpm, pH = 5,5; dp = 120 – 140 mesh.
V. KESIMPULAN
1. Makin besar rasio enzim terhadap substrat, makin besar konsentrasi glukosa yang dihasilkan, tetapi efisiensi enzim menjadi lebih kecil. Peningkatan rasio enzim substrat dari 14 U/g sampai 28U/g meningkatkan konsontrasi glukosa yang dihasilkan selama 7 jam hidrolisis dari 1,748 g/L menjadi 3,222 g/L.
2. Ukuran partikel jerami 120 – 140 mesh cukup optimum untuk dipilih pada hidrolisis jerami padi dan dapat meningkatkan konsentrasi glukosa sekitar 72%.
3. Penurunan pH dari 5,52 sampai 3,92 dapat menurunkan waktu hidrolisis optimum dari 6 jam menjadi 3 jam. 4. Makin besar rasio enzim-substrat, efisiensi enzim makin kecil.
Saran
Hidrolisis yang dilakukan pada penelitian ini dilakukan dalam skala kecil, sehingga faktor-faktor hidrodinamika sepertinya tidak berpengaruh. Hidrolisis tersebut merupakan sistem heterogen yang dalam skala besar pengaruh faktor-faktor hidrodinamikanya sangat penting, oleh karena itu perlu dipelajari lebih lanjut tentang pengaruh faktor-faktor hidrodinamika dalam reaktor hidrolisis enzimatik jerami padi.
Ucapan Terima Kasih
Ucapan terima kasih kami sampaikan kepada DP2M DIKTI yang telah membiayai penelitian ini.
VI. PUSTAKA
1. B.O. Aderemi, B.O. , E. Abu, B. K. Highina, “The Kinetics of Glucose Production from Rice Straw by Aspergillus niger”, African Journal of Biotechnology Vol. 7 (11), pp. 1745-1752, June 2008.
2. G. Antonopoulou, I. Ntaikou, H.N. Gavala, I.V. Skiadas, K.Angelopoulos, G. Lyberados, “Biohydrogen Production from Sweet Sorgum Biomass Using Mixed Acidogenic Cultures and Pure Cultures of
Ruminococcus Albus”, Global NEST Journal, Vol 9, No 2, pp. 144-151, 2007.
t (jam) Konsentrasi glukosa (g/L) pH = 5,5 pH = 4,8 pH = 3,9 0 0 0 0 0,25 0,744 0,5 1,949 1,248 2,330 1,0 2,795 1,925 3,437 1,5 3,469 2,0 3,775 3,504 5,407 3,0 4,100 3,811 5,978 4,0 4,660 4,100 5,834 5,0 5,206 6,0 5,548
3. U. Bossel, U., “Well-to-Wheel Studies, Heating Values, and the Energy Conservation Principle”, European Fuel Cell Forum, 2003.
4. H.L. Chin, H.L., Z.S. Chen, C. P. Chou “Fedbatch Operation Using Clostridium acetobutylicum Suspension Culture as Biocatalyst for Enhancing Hydrogen” Production, Biotechnol. Prog., Vol. 19, 383 – 388, 2003. 5. H. Dewi, “Hidrolisis Limbah Hasil Pertanian Secara Enzimatik, Akta Agrosia”, Vol. 5, No. 2, pp. 67 – 71,
2002.
6. T. Juhasz, K. Kozma, Z. Szengyel, K. Reczey, “Production of β-Glucosidase in Mixed Culture of Aspergillus niger BKMF 1305 and Trichoderma reesei RUT C30”, Food Technol. Biotechnol. 41 (1), pp. 49–53, 2003.
7. I.K. Kapdan, F. Kargi, “Bio-hydrogen Production from Waste Materials”, Enzyme and Microbial Technology 38, 569–582, 2006.
8. S.M. Kotay, D. Das, “Microbial hydrogen production with Bacillus coagulans IIT-BT S1isolated from anaerobic sewage sludge”, Bioresource Technology, Elsevier, 2006.
9. P. Mahakhan, C. Chobvijuk, M. Ngmjarearnwong, S. Trakulnalermsai, C. Bucke, J. Svasti, W. Kanlayakrit, L. Chitradon, “Molecular Hydrogen Production by a Thermotolerant Rubrivivax gelatinosus using Raw Cassava Starch as an Electron Donor”, Science Asia, Vol. 31, pp. 415 – 424, 2005.
10. M. Morimoto,M. Atsuko, A.A.Y. Atif, M.A. Ngan, A. Fakhru’l-Razi, S.E. Iyukeb, A.M. Bakir (2004), “Biological Production of Hydrogen from Glucose by Natural Anaerobic Microflora”, International Journal of Hydrogen Energy 29, 7pp. 09-713, 2004.
11. H. Ogino, T. Miura, K. Ishimi, M. Seki, H. Yoshida, “Hydrogen Production from Glucose by Anaerobes”., Biotechnology Proggress, Vol. 21, pp. 1786–1788, 2005.
12. B. Raghavendra, Havnnavar, G.S., Geeta, “Pre-treatment of Agroresidues for Release of Maximum Reducing Sugar”, Karnataka J. Agric. Sci., 20 (4), pp. 771-772, 2007.
13. A. Reungsang, S. Sangyoka, T. Imai, P. Chaiprasert, “Biohydrogen Production from Cassava Starch Manufacturing Wastewater”, The Joint International Conference on “Sustainable Energy and Environment (SEE)”, pp. 319-327, 2004.
14. S. Sabiham and B. Mulyanto, “Biomass Utilization in Indonesia: Integration of Traditional and Modern Principles of Organic Matter Management”, Paper is presented in APECATC Workshop on Biomass Utilization held in Tokyo and Tsukuba Japan, pp. 19 – 21 January 2005.
15. S. Sung, L. Raskin, T. Duangmanee, S. Padmasiri, J. J. Simmons, “Hydrogen Production by Anaerobic Microbial Communities Exposed to Repeated Heat Treatment”, Proceedings of the 2002 U.S. DOE Hydrogen Program Review NREL/CP-610-32405, 2005,
16. T. de Vrije, G.G. de Haas, G.B. Tan, E.R.P. Keijsers, P.A.M. Claassen, “Pretreatment of Miscanthus for hydrogen production by Thermotoga elfii, International Journal of Hydrogen Energy, Vol. 27, pp. 1381 – 1390, 2002.
17. A. Widjaja, “Aplikasi Bioteknologi pada Industri Pulp dan Kertas”, itspress, pp. 4 – 5, 2009.
18. Q. Xiang, Y.Y. Lee, P.O. Pettersson, R.W. Torget, “Heterogeneous Aspects of Acid Hydrolysis of α-Cellulose”, Applied Biochemistry and Biotechnology, pp. 105–108, 2003.
19. H. Zhang, M.A. Bruns, B.E. Logan, “Biological Hydrogen Production by Clostridium asetobutylicum in an