Structure, expression and promoter activity of two polyubiquitin
genes from rice (
Oryza sati
6
a
L.)
Jianlin Wang, Junda Jiang, James H. Oard *
Department of Agronomy,Louisiana Agricultural Experiment Station,Louisiana State Uni6ersity Agricultural Center,Baton Rouge,
LA 70803, USA
Received 9 December 1999; received in revised form 14 March 2000; accepted 14 March 2000
Abstract
We have isolated two rice polyubiquitin genes designated as RUBQ1 and RUBQ2 by screening a Bacterial Artificial
Chromosome (BAC) genomic library with a32P-labeled ubiquitin cDNA probe. DNA sequence data revealed that both genes
contained an open reading frame encoding a hexameric precursor ubiquitin and an intron immediate upstream of the initiation codon. The deduced amino acid sequences of both genes were identical to each other and to other plant ubiquitin sequences. Several putative regulatory elements such as enhancer core and heat shock consensus sequences were found in the 5%-upstream regions of both genes. Northern blot analyses using the 3%-untranslated region as gene specific probes showed that both genes were
actively expressed in all rice plant tissues tested. Differential expression was observed in roots where RUBQ2 appeared to be predominantly expressed. Chimeric genes containing the 5%-upstream region including the intron of RUBQ1 or RUBQ2 and the
b-glucuronidase (GUS) coding region were constructed and transferred into rice suspension cells via particle bombardment. GUS activity from constructs containing RUBQ1 and RUBQ2 promoters in rice suspension cells was ten to 15-fold greater than those using the Cauliflower Mosaic Virus 35S (CaMV 35S) promoter, and two to threefold greater than constructs with the maize polyubiquitin Ubi1 promoter. The results demonstrate the potential usefulness of the two rice polyubiquitin promoters in rice or other monocot transformation systems. © 2000 Elsevier Science Ireland Ltd. All rights reserved.
Keywords:Polyubiquitin gene; Expression; Promoter activity;Oryza sati6aL.
www.elsevier.com/locate/plantsci
1. Introduction
Ubiquitin, a 76 amino acid protein, is present in all eukaryotic cells and is one of the most con-served proteins, differing in only three of 76 amino acids between higher plants and animals [1]. Ubiq-uitin is encoded by small multigenic families con-taining two types of genes: polyubiquitin and
ubiquitin extension/fusion genes. The
polyubiqui-tin gene consists of tandem repeats of a 228-bp ubiquitin coding region [2 – 4], while the ubiquitin
extension/fusion gene consists of a single ubiquitin
coding region linked to a sequence encoding ribo-somal proteins either 52 or 76 – 81 amino acids in length [5,6]. Both types of genes are translated as a polyprotein precursor, then protealytically pro-cessed to the ubiquitin monomers for ubiquitin function [1]. The best-characterized function of ubiquitin is its conjugation to target proteins as a recognition signal for protein degradation [7]. Ubiquitin is also involved in other cellular pro-cesses, such as ribosome biosynthesis, chromatin structure and cell cycle control [8 – 10].
Various ubiquitin genes have been isolated in a number of organisms. In yeast, a total of four ubiquitin genes have been characterized, one of which belongs to the polyubiquitin class [11]. In
Arabidopsis thaliana, 14 ubiquitin genes represent-ing the complement of the ubiquitin gene family have been isolated, five being polyubiquitin genes
Approved for publication by the Director of the Louisiana Agricultural Experiment Station as manuscript number 00-09-0122. * Corresponding author. Tel.: +1-225-3881301; fax: + 1-225-3881403.
E-mail address:[email protected] (J.H. Oard).
[12]. Ubiquitin extension/fusion genes in plants are expressed in meristematic tissues and their expres-sion is significantly reduced in mature tissues [5,6,13]. The polyubiquitin genes were reported to be constitutively expressed in all plant organs tested [3,14], with an increased level in young tissues [2,15]. Differential expression of ubiquitin genes has been documented in potato tuber tissues in response to injury and hormone treatment [16],
and in Nicotiana syl6erstris to heavy metal
treat-ment and pathogen infection [17]. Expression of ubiquitin genes in response to heat-shocked treat-ment was reported to be up-regulated [3,4], un-changed [14], or down-regulated [16]. Perturbation in the ubiquitin system of higher plants was found to elicit a hypersensitive response similar to that caused by pathogen infection [18 – 20].
Various promoters from ubiquitin genes have been tested for their potential use in plant
trans-formation, including those fromA. thaliana [5,21],
sunflower [22], N. tabacum [23], potato [6], and
maize [4]. The maize polyubiquitin Ubi1 promoter has been successfully used in maize and other monocot transformation systems [4,15,24].
We report here two polyubiquitin genes isolated from a rice genomic library, their expression in different rice plant tissues, and their response to heat shock treatment. The possible roles of the putative regulatory elements in gene expression are discussed. Furthermore, promoters from the two polyubiquitin genes have been tested for their potential use in rice transformation. Our results showed that the two rice polyubiquitin genes were actively expressed in all tissues tested and a high level of glucuronidase (GUS) expression under control of the two rice polyubiquitin promoters was observed in transformed rice cells, indicating potential usefulness of the two rice polyubiquitin promoters in rice and other monocot transforma-tion systems.
2. Materials and methods
2.1. Plant materials
Eight-month-old rice suspension cells (cv. Taipei 309, provided by Mr S. Yang, Department of Plant Pathology and Crop Physiology, Louisiana State University) were maintained in 200 ml flasks containing 50 ml R2 medium [25] on a gyratory
shaker (80 rpm). Cultures were grown in light at 25°C and subcultured once a week. Rice plants and seedlings (cv. Cypress) were grown in the greenhouse. For heat shock treatments, the 2-week-old seedlings were immersed in pre-heated distilled water for 20 min at 42°C and allowed to recover for up to 90 min.
2.2. Isolation of genomic clones
A bacterial artificial chromosome (BAC) rice genomic library, kindly provided by Dr P. Ronald, UC Davis, consisted of 11 000 clones with an average DNA insert size of 125 kb [26]. The 1.3-kb
region of sugarcane ubiquitin cDNA clone
Scubi561 [27] was used as a probe for library
screening. Filters were prehybridized in 5×
Den-hardt’s solution, 100 mg/ml denatured sperm
DNA, 6×SSC and 0.5% SDS for 6 h at 65°C.
Hybridizations were performed overnight at 65°C
in the same buffer containing 32P-labeled probe.
Filters were washed twice at room temperature in
2×SSC, 0.1% SDS and then once at 65°C for 20
min in 0.2×SSC, 0.1% SDS solution.
Autoradiog-raphy was carried out at −80°C with Kodak
Biomax MS film and one intensifying screen. Twenty-four positive clones were identified, and two of them, designated RUBQ1 and RUBQ2, were selected for further analysis because both clones showed strong hybridizing signals to the sugarcane ubiquitin cDNA probe (data not shown) but differed from each other based on restriction enzyme analysis.
2.3. DNA sequencing
HindIII restriction fragments encompassing
RUBQ1 and RUBQ2 were cloned into pBluescript
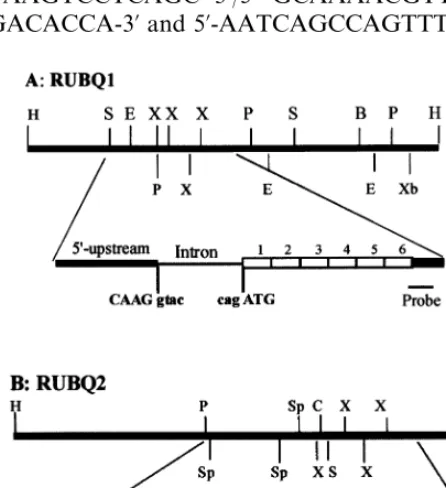
II SK+ (Stratagene) for restriction mapping (Fig.
1). When digested with XhoI or BglII restriction enzymes, both clones produced several fragments hybridizing to the sugarcane ubiquitin cDNA probe. These fragments were gel-purified and
sub-cloned into pBluescript SK II+ for DNA
se-quencing. Double stranded DNAs were prepared using plasmid mini preparation kit (Qiagen). DNA sequencing was performed on an ALF-win auto-matic DNA sequencer (Parmacia) using the
ALF-express autocycle sequencing kit (Parmacia)
and M13-reverse or specific oligodeoxyribonucle-otides as primers. The DNAsis (Hitachi, CA) soft-ware was used for data analysis. DNA sequences presented here corresponding to RUBQ1 and RUBQ2 have been entered into Genbank with accession numbers AF184279 and AF184280, respectively.
2.4. Northern and Southern bolt analyses
The 0.33 kb XhoI fragment of the RUBQ2 coding region (nucleotides 3513 – 3846 of RUBQ2) was used as a probe for Northern blot analysis.
Gene specific probes 3%-RUBQ1 (181 bp in length)
and 3%-RUBQ2 (218 bp in length) were produced
by polymerase chain reaction (PCR) using primers
flanking the 3%-untranlated regions of RUBQ1 and
RUBQ2. Two sets of primers used to amplify
3%-RUBQ1 and 3%-RUBQ2 were 5%
-GTGGCCAG-TAAGTCCTCAGC - 3%/5%-
GCAAAACGTTTCA-GACACCA-3%and 5%
-AATCAGCCAGTTTGGT-GGAG - 3%/5%-
TGCAAATGAGCAAATTGAGC-3%, respectively. The PCR thermocycle parameters
were 95°C/36 s, 50°C/36 s, and 72°C/84 s for 25
cycles. PCR products (3%-RUBQ1 and 3%-RUBQ2)
were gel purified and32P-labeled using the random
primed DNA labeling kit (Boehringer Mannheim) for Northern and Southern blotting.
Total RNA from different tissues of rice plants and heat-shocked versus control seedlings (cv.
Cy-press) were isolated with phenol/SDS extraction
and LiCl precipitation [28]. Total RNA (20 mg per
lane) were electrophoretically separated in 1.0% agarose – formaldehyde gels and blotted onto ny-lon membranes (Hybond-N, Amersham) [28]. Fil-ters were prehybridized for 6 h at 50°C in
5×SSC, 5×Denhards, 50 mg/ml salmon sperm,
and 50% formamide. Hybridization was per-formed overnight using the same conditions with
addition of 32P-labeled probe (specific activity of
1×106 cpm/ml). Filters were washed twice at
room temperature in 2×SSC and 0.1% SDS for
20 min followed by two 20 min washes at 50°C in
0.2×SSC and 0.1% SDS. Autoradiography
was carried out with Kodak Biomax MS film for 6 h. For Southern blots, DNAs were digested with restriction enzymes, fractionated by 1.0% agarose gel electrophoresis, and transferred onto nylon membranes (Hybond-N, Amersham) ac-cording to the Molecular Cloning Manual [29]. Hybridization and subsequent washing conditions were the same as described for the BAC library screening.
2.5. Chimeric gene constructions
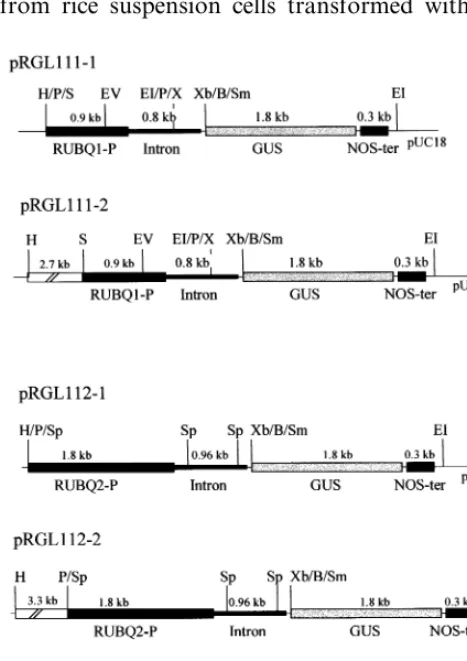
A 2.2 kb HindIII/EcoRI fragment containing
the GUS coding region and nopaline synthase (NOS) terminator was removed from pBI101 (Clontech) and ligated into the corresponding site of pUC18, generating pRGL110. Promoter re-gions to be tested were inserted into the polylinker of pRGL110 as a translational fusion to the GUS coding sequence.
For expression of the GUS gene under control of the rice ubiquitin promoters, the 1.7 kb SalI
/XhoI and 2.8 kb PstI/XhoI fragments, each
con-taining the 5%-upstream region and the intron of
RUBQ1 and RUBQ2, respectively, were ligated into the SalI site (compatible with XhoI site) of
the pRGL110, creating pRGL111-1 and
pRGL112-1 (Fig. 6). Constructs pRGL111-2 and
Fig. 1. Restriction maps and organization of two rice polyu-biquitin genes RUBQ1 (A) and RUBQ2 (B). Restriction maps of the insert of each genomic clone containing rice polyubiq-uitin genes are presented on the top, below is the organization of the two polyubiquitin genes based on sequence data. The 5%-upstream regions are indicated as a solid bar, followed by an intron defined by consensus sequences at 5%and 3% splice
sites. The six repeats of ubiquitin-coding region are indicated as open boxes, followed by 3%-untranslated regions. Gene
pRGL112-2 are the same as pRGL111-1 and pRGL112-1, except for inclusion of an additional
2.7 kb HindIII/ SalI or 3.3 kb HindIII/PstI
frag-ment at the 5%-upstream region of RUBQ1 and
RUBQ2, respectively (Fig. 6). All constructs en-coded a fusion protein at the N-terminus, consist-ing of a portion (15 amino acids) of ubiquitin followed by linker sequence of 11 amino acids upstream of the GUS protein.
For construction of the chimeric gene contain-ing the RUBQ2 promoter and the Luciferase (LUC) coding region, the SV 40 terminator in vector pGL3 (Promega, Madison, WI) was re-placed with the NOS terminator, creating pGL3-1. The RUBQ2 promoter, contained in a 2.8 kb
HindIII/ BamHI fragment from pRGL112-1, was
blunt-ended by treating with T4 DNA polymerase (New England Biolab) and ligated into the blunt-ended HindIII site of the pGL3-1. The resulting construct designated as pRUBQ2-LUC (Fig. 6) encoded a fusion protein with 29 amino acids at the N-terminus upstream of the LUC protein.
Two controls were used for rice transformation. The plasmid pAHC25, containing a maize polyu-biquitin Ubi1 promoter-GUS cassette, was kindly provided by Dr P. Quail (Plant Gene Expression Center, USDA, Albany, CA). Plasmid pBI121, containing a CaMV 35S promoter-GUS cassette, was obtained from Clontech (Palo Alto, CA).
2.6. Transient GUS and LUC assays
Approximately 200 mg rice suspension cells (Taipei 309) were evenly distributed over the sur-face of a 70 mm Whatman no. 2 filter paper in a
100×15 mm2 petri dish for particle
bombard-ment. Plasmids were precipitated onto gold
parti-cles (1.0 mm in diameter, Biorad) and introduced
into suspension cells via particle bombardment according to the method described by Christou et al. [30]. Two independent experiments were con-ducted as follows: Equimolar amounts of each of six constructs containing the chimeric GUS gene were co-transferred with pRUBQ2-LUC into sus-pension cells with four replications. The pRUBQ2-LUC served as internal control and inclusion of an internal control was reported to improve the reli-ability of transient expression studies by con-trolling the variations among replications [31]. After 48 h incubation of treated suspension cells at 25°C, cells were transferred into a 50 ml centrifuge
tube and homoginized with 300 ml 1×reporter
assay buffer (Promega). Cell homogenates were
centrifuged twice at 10 000×g for 10 min at 4°C,
then the supernatant was transferred to 1.5 ml
eppendorf tube and stored at −80°C until use for
both GUS and LUC assays. The means of GUS/
LUC relative activity were tested for significance with Tukey’s test using SAS version 6.12.
For the GUS enzyme assay, 20 ml of
superna-tant were assayed according to the method previ-ously described by Jefferson et al. [32] in a TKO 100 fluorimeter (Hoefer Co.). GUS activity was
expressed as pmol MU (4-methylumbelliferone)/
min. For the LUC enzyme assay, 20ml of
superna-tant were assayed in a scintillation analyzer (Packard) using a Luciferase assay system kit (Promega). Measurements began 2 min after
mix-ing the 20 ml supernatant and the luciferase assay
reagent, which took 12 s for each sample. The LUC activity was expressed as the number of counts per minute (CPM). Measurements based on light emission were standardized with defined
amounts of purified recombinant luciferase
(Promega) (1 light unit (L.U.) is equivalent to
2.8×105 CPM).
3. Results
3.1. Sequence and structure of RUBQ1 and RUBQ2
DNA sequences of the 5%-flanking region
includ-ing the first exon, an intron and the first 48 nucleotides of the ubiquitin-coding regions of RUBQ1 and RUBQ2 are presented in Fig. 2A and B, respectively.
Both genes contained an open reading frame of 1371 bp, arranged as six tandem, head to tail repeats of 228 bp, encoding a hexameric precursor ubiquitin (Fig. 1). The deduced amino acid se-quence of the 12 ubiquitin monomers from both genes was identical to each other and other
ubiqui-tin sequences in maize [4] and A. thaliana [2]. The
last (sixth) monomer of both RUBQ1 and RUBQ2 contained a glutamine extension. The ex-tension of one to several amino acid residues at the C-terminal of the last ubiquitin monomer is a common feature of the polyubiquitin genes [1].
Fig. 2. Nucleotide sequences of the 5%-upstream regions of rice RUBQ1 (A) and RUBQ2 (B). The heat shock consensus sequence
is boxed. The enhancer core consensus element is indicated with a black dotted line. The TATA box is underlined. Downstream of TATA box is the first exon (the transcription start site remains to be determined), followed by the intron indicated in lower case. The first 48 nucleotides of ubiquitin coding region of both genes are showed along with their deduced amino acid sequences. Restriction sites used to construct the ubiquitin promoter-based vectors are indicated.
showed high homology to other ubiquitin genes from Arabidopsis [2,12], sunflower [3], potato [6],
tomato [13], N. tobacum [23], and sugarcane [27]
and yeast [11]. For example, the coding sequences of RUBQ1 and RUBQ2 genes showed 88% iden-tity to the maize polyubiquitin gene (Genbank
accession no. S94464), 81% to the A. thaliana
ubiquitin gene (accession no. U84968), and 82% to
the sunflower ubiquitin gene (accession no.
X57004). The coding sequence of RUBQ1 showed 99% identity to the rice polyubiquitin cDNA clone rub1 [33] and Rubq1 [34], suggesting the genomic clone RUBQ1 reported here corresponded to these two previously isolated cDNA clones.
RUBQ1 and RUBQ2 genes (Figs. 1 and 2) based
on the consensus sequences CAAG/gtac at the
splice 5% site and cag/ATG at the splice 3% site,
which are invariant for all plant polyubiquitin genes characterized to date [3,4,14,21]. The posi-tion of the putative intron for RUBQ1 was ver-ified when sequence of RUBQ1 was compared to that of corresponding cDNA clones rub1 [33] and Rubq1 [34]. In contrast to the coding region, the RUBQ1 intron varied in length (782 bp) compared to the RUBQ2 intron (962 bp) and showed only 52% identity with each other.
The 5%-upstream regions of both genes showed
53% identity to each other. Putative regulatory
elements were found in the 5%-upstream regions of
both genes. For example, a putative TATA box was observed at position 811 for RUBQ1 gene and position 1712 for RUBQ2 gene (Fig. 2A and B). A
heat shock sequence similar to the Drosophila
consensus sequence 5%-CTGGAATnTTCTAGA
[35] was found twice at positions 630 (182 bases upstream of TATA box) and 652 (160 bases up-stream of TATA box) for RUBQ1, and once at position 1586 (126 bases upstream of the TATA box) for RUBQ2. It is interesting to note that an
enhancer core consensus sequence 5%
-GGTGTG-GAAA(or TTT)G-3% [36,37] was present at
posi-tion 286 (526 bp upstream of the TATA box) in the RUBQ1 and at position 1019 (694 bases up-stream of the TATA box) in the RUBQ2 (Fig. 2A and B).
3.2. Ubiquitin is encoded by a small gene family in rice
Southern blot analysis showed that multiple re-striction fragments from rice genomic DNA hy-bridized to the ubiquitin cDNA clone Scubi561 (Fig. 3A). Five major hybridization bands (14, 11, 8.3, 2.4 and 2.2 kb) were observed for DNA digested with HindIII enzyme and a comparative number of major bands were also found for DNAs digested with EcoRI or XbaI (Fig. 3A). These major bands appeared to represent fragments con-taining polyubiquitin genes and two of them (14 and 8.3 kb) were confirmed to contain RUBQ1 and RUBQ2 genes, respectively (see below). In addition to the major bands, several faint bands were observed, which may reflect fragments
carry-ing the ubiquitin extension/fusion genes with a
single repeat of ubiquitin monomer. These results indicate ubiquitin is encoded by a small gene family in rice, though the exact number of the ubiquitin genes remains to be determined.
A simple hybridization pattern was generated when the same filter was sequentially hybridized
by gene specific probe 3%-RUBQ1 and 3%-RUBQ2
(Fig. 3B and C). The 3%-RUBQ1 probe hybridized
to a 14 kb fragment in HindIII-digested genomic DNA, and the specific fragments in EcoRI or XbaI-digested genomic DNA were also recognized
by the 3%-RUBQ1 probe (Fig. 3B). Similarly, the
3%-RUBQ2 probe hybridized to a single 8.3 kb
fragment in HindIII digested genomic DNA, and it also recognized a single specific fragment in EcoRI or XbaI digested genomic DNA (Fig. 3C). The results suggest that RUBQ1 and RUBQ2 exist in the rice genome at a single locus and
demon-strate the utility of 3%-RUBQ1 and 3%-RUBQ2 as
gene-specific probes.
3.3. Expression of RUBQ1 and RUBQ2 in rice tissues
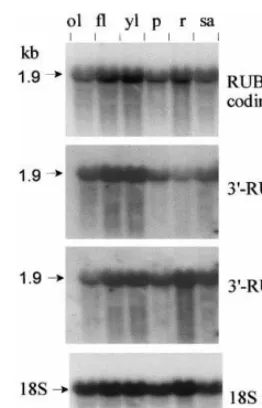
Total RNA isolated from different rice plant organs (cv. Cypress) was analyzed by Northern blot analysis using the ubiquitin coding region of
RUBQ2 and gene specific regions 3%-RUBQ1 and
3%-RUBQ2 as hybridization probes (Fig. 4).
Hy-bridization with the ubiquitin coding region (RUBQ2 coding) probe detected one predominant band with an estimated size of 1.9 kb. This 1.9 kb transcript were present in all tissues tested, which
Fig. 4. Northern blot analysis of ubiquitin expression in rice tissues. Total RNA (20mg per lane) was prepared from old leaves (ol), full-developed leaves (fl), young leaves (yl), panicle (p), roots (r) and shoot apex (sa). The filter was sequentially hybridized with three probes: ubiquitin coding region of
RUBQ2, gene specific probes 3%-RUBQ1 and 3%-RUBQ2.
Finally, a 24-base antisense oligonucleotide to rice 18S rRNA was labeled with [g-32P] ATP (Amersham) and T4
polynucle-otide kinase (NE Biolabs) and used as an internal control.
tested (Fig. 4). Differential expression of the two genes was observed in roots, where expression of RUBQ2 was substantially higher than that of RUBQ1.
3.4. Expression of RUBQ1 and RUBQ2 in response to heat shock
The effect of heat shock on expression of ubiq-uitin RNA was examined by Northern blot
analy-sis using the ubiquitin coding region
(RUBQ2-coding) as a probe. The level of 1.9 kb transcript was increased immediately after 20 min heat shock treatment at 42°C and accumulated over a 90-min period (Fig. 5). To test whether the increased abundance was due to higher transcript levels of RUBQ1 or RUBQ2 or both genes, Northern blots of heat shocked RNAs were
hy-bridized with gene specific probes 3%-RUBQ1 and
3%-RUBQ2. The results shown in Fig. 5 indicate
expression of both genes was up-regulated in re-sponse to heat shock, which was consistent with
the heat-shock sequence located in the 5%-upstream
regions of both genes
3.5. Transient expression of rice ubiquitin promoters in rice suspension cells
Fig. 6 shows schematic features of chimeric genes containing rice ubiquitin promoters. These constructs were co-transferred with pRUBQ2-LUC into rice suspension cells via particle
bom-bardment. Both GUS (pmol MU/min) and LUC
(light unit) activities were determined and their relative values are shown in Table 1. Results across two independent experiments were
consis-tent though the overall GUS/LUC relative
activi-ties were greater in experiment 2 than in
experiment 1.
As shown in Table 1, constructs pRGL111-1
and pRGL111-2 produced similar GUS/LUC
rela-tive activities in both experiments, which suggested
that the additional 5%-flanking region (2.7 kb) in
pRGL111-2 did not contribute to the promoter
activity of RUBQ1. However, construct
pRGL112-2 showed a reduced GUS/LUC relative
activity compared to that produced by construct
pRGL112-1. GUS/LUC relative activities
pro-duced by pRGL112-2 and pRGL112-1 were 2.58 and 3.82 in experiment 1, and 11.17 and 16.00 in experiment 2. These values were statistically
differ-Fig. 5. Northern blot analysis of rice ubiquitin expression in response to heat shock treatment. Two-week-old seedlings of rice variety Cypress were heat-shocked at 42°C for 20 min.
Total RNA (20 mg per lane) was prepared from unshocked
rice seedlings (lane 1), or heat-shocked seedlings allowed to recover for 0 min (lane 2), 30 min (lane 3) and 90 min (lane 4). The filter was sequentially hybridized with ubiquitin
cod-ing region of RUBQ2, gene specific probe 3%-RUBQ1 and
3%-RUBQ2. The filter was then probed with a 24-base
anti-sense oligonucleotide to rice 18S rRNA.
included old leaves, fully developed leaves, imma-ture leaves, young panicles, roots of seedlings, and vegetative shoot apex.
Gene specific probes 3%-RUBQ1 and 3%-RUBQ2
ent in both experiments, indicating the additional
5%-flanking region (3.3 kb) in pRGL112-2 may
reduce transformation efficiency or contain
nega-tive regulatory elements. Though means of GUS/
LUC relative activity were greater in experiment 2 than in experiment 1, the fold stimulation values related to pBI121 (35S-GUS) were relative
con-stant across two experiments, being 11.4
(pRGL112-2) and 16.9 (pRGL112-1) in
experi-ment 1, and 11.7 (pRGL112-2) and 16.8
(pRGL112-1) in experiment 2. Activity of the
RUBQ2 promoter was 1.5 times that of the
RUBQ1 promoter in rice suspension cells when
means of GUS/LUC relative activity produced by
pRGL111-1 and pRGL112-1 were compared. Higher levels of GUS expression were observed from rice suspension cells transformed with
con-Table 1
Expression of chimeric GUS genes transferred into rice sus-pension cells via particle bombardmenta
GUS/LUC ratios
Transferred DNA
Fold
Construct Experiment Fold Experiment
no. 1 no. 2
16.8 16.0093.26b
pRGL112-1 3.8290.82b 16.9
11.4 11.1792.38c 11.7
pRGL112-2 2.5890.26c
12.7 9.9292.28c 10.4
pRGL111-1 2.8690.14c
8.9 8.4592.25c
12.5
pRGL111-2 2.8490.56c
0.7990.06d
aEach construct was co-transferred with pRUBQ2-LUC
which served as internal control (see Section 2). The means and standard deviation of GUS/LUC ratios from four repli-cates are shown for two independent experiments. Data with different superscript letters (b–e) are significantly different from each other at the 5% level based on Tukey’s test.
Fig. 6. Schematic representation of constructs containing rice ubiquitin promoter and GUS or LUC chimeric genes. The
sizes of 5%-upstream regions (RUBQ1-P and RUBQ2-P) and
introns of RUBQ1 and RUBQ2 are indicated. Additional 5%-flanking regions are indicated as open boxes in plasmids pRGL111-2 and pRGL112-2. H, HindIII; P, PstI; S, SalI; EI, EcoRI; EV, EcoRV; X, XhoI; Xb, XbaI; B, BamHI; Sm, SmaI; Sp, SpeI; C, ClaI.
structs containing the rice ubiquitin promoter-GUS chimeric genes than those using the CaMV 35S or maize ubi1 promoter (Table 1). GUS activ-ity from constructs pRGL111-1 and pRGL112-1 in rice suspension cells was ten to 15-fold greater than the construct pBI121 (CaMV 35S-GUS), and two to threefold greater than pAHC25 (maize Ubi1-GUS), indicating the potential usefulness of the two rice polyubiquitin promoters in rice or other monocot transformation systems.
4. Discussion
We report here the isolation and characteriza-tion of two polyubiquitin genes from rice. Though
the 5%-upstream regions of the two rice
polyubiqui-tin genes showed only 53% sequence identity to each other, they consisted of the same putative regulatory elements, such as enhancer core and heat shock consensus elements. The enhancer core
elements found in the 5%-upstream regions of
RUBQ1 and RUBQ2 are GTTGTGGTTGG and
GTTGTGGTTTG, respectively, which are 9/11
and 10/11 identical to the enhancer core consensus
9/11 identical to the enhancer core consensus ele-ment. The maize adh1 promoter was most active in anaerobically induced root tissues [39], and a substitution mutation in the enhancer core element in the maize adh1 promoter contributed to a re-duced anaerobic response in roots of transgenic rice plants [40]. We speculate that the enhancer core element found in the two rice polyubiquitin genes could be also related to gene expression in roots. Furthermore, functions of the enhancer core element may not be restricted to roots. For exam-ple, part of the enhancer core element (TGTG-GTT) defined as a pollen box (PB) motif is present in the promoters of potato genes Lat52, Lat56, and Lat59, and a GG doublet in the PB core is critical for expression in pollen [41]. GUS expres-sion was also observed in the pollen of transgenic rice plants transformed with constructs containing the chimeric RUBQ1 or RUBQ2 promoter-GUS genes (Wang and Oard, unpublished data). It is likely that the enhancer core element in the two rice polyubiquitin genes serves as a global en-hancement of gene expression, though its exact
function(s) in plant genes remains to be
determined.
Expression of both rice polyubiquitin genes are induced in response to heat shock treatment, which is consistent with heat shock elements found
in the 5%-upstream regions of both rice
polyubiqui-tin genes. Heat-shock elements have also been
reported in the 5%-upstream regions of
polyubiqui-tin genes from maize [4], sunflower [3] and N.
tabacum[23]. The consensus heat shock element in
Drosophilawas defined as CTGGAATNTTCTAG [35]. Amin et al. [42] demonstrated that presence of a single module of an inverted repeat (nGAAn) was sufficient for heat shock induction, and devia-tions from the consensus sequence were tolerant if larger arrays of the repeats occurred. The heat shock element in the RUBQ2 fits the single mod-ule of an inverted repeat (nGAAn), while devia-tions from consensus sequence occur in the two heat shock elements found in the RUBQ1, which are similar to those in the maize polyubiquitin gene [4]. Little is known about the position effect of the heat shock element on the magnitude of response to heat shock treatment. The heat shock element is closer to the TATA box in the RUBQ2 than those in the RUBQ1, but comparison of the position effect is not possible since variation oc-curs in the sequences and the copy number of the
heat shock elements found in the promoter regions.
Activity of the RUBQ2 promoter was 1.5
times that of the RUBQ1 promoter in driving GUS expression in rice suspension cells. In addi-tion, the RUBQ2 promoter was also shown to be more efficient than the RUBQ1 promoter in di-recting reporter gene expression in transgenic rice plants (Wang and Oard, unpublished data). Sev-eral factors may contribute to the enhanced ex-pression of the RUBQ2 promoter. First, the
RUBQ2 promoter contains 1 kb longer 5%
-up-stream region than did the RUBQ1 promoter.
Second, sequence variations between the 5%
-up-stream regions including the introns of the two rice polyubiquitin genes may contribute to differ-ences in gene expression levels. In comparison, promoter regions of three polyubiquitin genes from A. thaliana were reported to be quantita-tively similar in driving transient GUS expression inA. thalianaleaves [21]. Their differential expres-sion could be revealed in transgenic plants since
differential expression ofA. thaliana polyubiquitin
genes were shown by Northern blot analysis [2].
Alternatively, the additional 5%-upstream regions
or 3%-untranslational regions that were not
in-cluded in the expression cassette were determi-nants of the differential expression [21].
The CaMV 35S promoter is widely used in both monocot and dicot transformation systems [43], while the maize Ubi1-promoter was shown to be more efficient than the CaMV 35S promoter in many monocot transformation systems including rice [15,24]. In this experiment, the transient GUS expression showed that constructs containing RUBQ1 and RUBQ2 promoters produced ten to 15-fold greater GUS activity than pBI121 (CaMV 35S-GUS), and two to threefold greater than pAHC25 (maize Ubi1-GUS), indicating the utility
of the 5%-flanking regions of the rice ubiquitin
direct comparisons of the rice ubiquitin promoters vs CaMV 35S or maize ubi1 promoters in rice and other plant transformation systems are needed to provide more information about their relative effi-ciency, the rice ubiquitin promoters reported here should be useful in plant transformation studies.
Acknowledgements
We thank Dr P. Ronald (UC Davis) for kindly providing the BAC rice genomic library, Dr H.
Albert (USDA/Agricultural Research Center,
Aiea, HI) for the sugarcane ubiquitin cDNA probe, Dr P. Quail (Plant Gene Expression Cen-ter, Albany, CA) for the plasmid pAHC25, Dr A.
Sylvester (USDA/Research Unit of Honey Bee
Breeding, Genetics and Physiology, Baton Rouge, LA) for use of ALF-win DNA sequencer, and J. Stelzer for technical assistance. The authors also thank Drs N. Murai and D. Shih for their critical readings of the manuscript.
References
[1] J. Callis, R.D. Vierstra, Ubiquitin and ubiquitin genes in higher plants, Oxford Surv. Plant Mol. Cell Biol. 6 (1989) 1 – 30.
[2] T. Burke, J. Callis, R.D. Vierstra, Characterization of a polyubiquitin gene fromArabidopsis thaliana, Mol. Gen. Genet. 213 (1988) 435 – 443.
[3] M.N. Binet, J.-H. Weil, L.-H. Tessier, Structure and expression of sunflower ubiquitin genes, Plant Mol. Biol. 17 (1991) 395 – 407.
[4] A.H. Christensen, R.A. Sharrok, P.H. Quail, Maize polyubiquitin genes: structure, thermal perturbation of expression and transcript splicing, and promoter activity following transfer to protoplasts by electroporation, Plant Mol. Biol. 18 (1992) 675 – 689.
[5] J. Callis, J. Raasch, R.D. Vierstra, Ubiquitin extension proteins of Arabidopsis thaliana: structure, localization and expression of their promoters in transgenic tobacco, J. Biol. Chem. 265 (1990) 12486 – 12493.
[6] J.E. Garbino, W.R. Belknap, Isolation of ubiquitin-ribo-somal protein of gene (ubi3) from potato and expression of its promoter in transgenic plants, Plant Mol. Biol. 24 (1994) 119 – 127.
[7] M. Hochstrasser, Ubiquitin-dependent protein degrada-tion, Annu. Rev. Genet. 30 (1996) 405 – 439.
[8] I.L. Goldknopf, H. Busch, Isopeptide linkage between non-histone and histone 2A polypeptides of chromoso-mal conjugate protein A24, Proc. Natl. Acad. Sci. USA 74 (1977) 864 – 868.
[9] D. Finley, B. Bartel, A. Varshavsky, The tail of ubiquitin precursors are ribosomal proteins whose fusion to
ubiq-uitin facilitates ribosome biogenesis, Nature 338 (1989) 394 – 401.
[10] M. Rechsteiner, Natural substrates of the ubiquitin pro-teolytic pathway, Cell 66 (1991) 615 – 618.
[11] E. Ozkaynak, D. Finley, M.J. Solomon, A. Varshvasky, The yeast ubiquitin genes: a family of natural gene fusions, EMBO J. 6 (1987) 1427 – 1439.
[12] J. Callis, T. Carpenter, C.-W. Sun, R.D. Vierstra, Struc-ture and evolution of genes encoding polyubiquitin and ubiquitin-like proteins in Arabidopsis thaliana ecotype Columbia, Genetics 139 (1995) 921 – 939.
[13] N.E. Hoffman, K. Ko, D. Milkowski, E. Pichersky, Isolation and characterization of tomato cDNA and genomic clones encoding the ubiquitin gene ubi3, Plant Mol. Biol. 17 (1991) 1189 – 1201.
[14] P. Kawalleck, I.E. Somssich, M. Feldbrugge, K.
Hahlbrock, B. Weisshaar, Polyubiquitin gene expression and structural properties of the ubi4-2 gene in Pet-roselium crispum, Plant Mol. Biol. 21 (1993) 673 – 684. [15] M.-J. Cornejo, D. Luth, K.M. Blankenship, O.D.
Ander-son, A. Blechl, Activity of a maize ubiquitin promoter in transgenic rice, Plant Mol. Biol. 23 (1993) 567 – 581. [16] J.E. Garbino, D.R. Rockhold, W.R. Belknap,
Expres-sion of stress-responsive ubiquitin genes in potato tu-bers, Plant Mol. Biol. 20 (1992) 235 – 244.
[17] P. Genschik, Y. Parmentier, A. Durr, J. Marbach, M.-C. Criqui, E. Jamet, J. Fleck, Ubiquitin genes are differen-tially regulated in protoplast-derived cultures of Nico-tiana syl6estris and in response to various stress, Plant
Mol. Biol. 20 (1992) 897 – 910.
[18] A. Bachmair, F. Becker, V. Masterson, J. Schell, Pertur-bation of the ubiquitin system causes leaf curling, vascu-lar tissue alterations, and necrotic lesions in a higher plant, EMBO J. 9 (1991) 4543 – 4549.
[19] F. Becker, E. Buschfeld, J. Schell, A. Bachmair, Altered response to viral infection by tobacco plants perturbed in ubiquitin system, Plant J. 3 (1993) 875 – 881.
[20] E.E. Karrer, R.N. Beachy, C.A. Holt, Cloning of to-bacco genes that elicit the hypersensitive response, Plant Mol. Biol. 36 (1998) 681 – 690.
[21] S.R. Norris, S.E. Meyer, J. Callis, The intron of Ara-bidopsis thaliana polyubiquitin genes is conserved in location and is a quantitative determinant of chimeric gene expression, Plant Mol. Biol. 21 (1993) 895 – 906. [22] M.N. Binet, M. Lepetit, J.-H. Weil, L.-H. Tessier,
Anal-ysis of a sunflower polyubiquitin promoter by transient expression, Plant Sci. 79 (1991) 87 – 94.
[23] P. Genschik, J. Marbach, M. Uze, M. Feuerman, B. Plesse, J. Fleck, Structure and promoter activity of a stress and developmentally regulated
polyubiquitin-en-coding gene of Nicotiana tabacum, Gene 148 (1994)
195 – 202.
[24] A.H. Christensen, P.H. Quail, Ubiquitin promoter-based vectors for high-level expression of selectable and/or screenable marker genes in monotyledonous plants, Transgenic Res. 5 (1996) 213 – 218.
[26] G. Wang, T.E. Holsten, W. Song, H. Wang, P. Ronald, Construction of a rice bacterial artificial chromosome library and identification of clones linked to Xa-21 disease resistant locus, Plant J. 7 (1995) 525 – 533. [27] H.H. Albert, B.C. James, P.H. Moore, Nucleotide
se-quence of a sugarcane polyubiquitin cDNA (accession no. L41658) (PRG95-045), Plant Physiol. 109 (1995) 337. [28] F.M. Ausubel, R. Brent, R.E. Kingston, D.D. Moore, J.G. Seidman, J.A. Smith, K. Struhl, Short Protocols on Molecular Biology, Wiley, New York, 1992, pp. 4.1 – 4.10.
[29] J. Sambrook, E.F. Fritsch, T. Maniatis, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989, pp. 9.1 – 9.58.
[30] P. Christou, T.L. Ford, M. Kofron, Production of trans-genic rice (Oryza sati6a L.) Plants from agronomically
important indica and japonica varieties via electric dis-charge particle-acceleration of exogenous DNA into
im-mature zygotic embryos, BioTechnology 9 (1991)
957 – 962.
[31] M. Lepetit, M. Ehling, C. Gigot, G. Hahne, An internal standard improves the reliability of transient expression studies in plant protoplasts, Plant Cell Rep. 10 (1991) 401 – 405.
[32] R.A. Jefferson, T.A. Kavanagh, M.W. Bevan, GUS fu-sions: b-glucuronidase as a sensitive and versatile gene fusion marker in higher plants, EMBO J. 6 (1987) 3901 – 3907.
[33] Y.-M. Kim, J.-K. Kim, Y.-S. Wang, Isolation and char-acterization of a full-length cDNA clone encoding a polyubiquitin, Plant Physiol. 106 (1994) 791 – 792. [34] E. Huq, L. Hirayama, M.A. Hossain, T.K. Hodges,
Characterization of a cDNA encoding a polyubiquitin gene in rice (accession no. U37687) (PRG97-004), Plant Physiol. 113 (1997) 305.
[35] H.R.H. Pelham, A regulatory upstream promoter ele-ment in Drosophila Hsp70 heat-shock gene, Cell 30 (1982) 517 – 528.
[36] H. Weiher, M. Konig, P. Gruss, Multiple point muta-tions affecting the simian virus 40 enhancer, Science 219 (1983) 626 – 631.
[37] G. Khoury, P. Gruss, Enhancer elements, Cell 33 (1983) 313 – 314.
[38] M.R. Olive, J.C. Walker, K. Singh, E.S. Dennis, W.J. Peacock, Functional properties of the anaerobic respon-sive element of the maize Adh1 gene, Plant Mol. Biol. 15 (1990) 593 – 604.
[39] J. Kyozaka, H. Fujimoto, T. Izawa, K. Shimamoto, Anaerobic induction and tissue- specific expression of maize Adh1 promoter in transgenic rice plants and their progeny, Mol. Gen. Genet. 228 (1991) 40 – 48.
[40] J. Kyozuka, M.R. Olive, W.J. Peacock, E.S. Dennis, K. Shimamoto, Promoter elements required for develop-mental expression of the maize adh1 gene in transgenic rice, Plant Cell 6 (1994) 799 – 810.
[41] D. Tweel, J. Yamaguchi, R.A. Wing, L. Ushida, S. McCormick, Promoter analysis of genes that are coordi-nately expressed during pollen development revealed pollen-specific enhancer sequences and shared regulatory elements, Gene Dev. 5 (1991) 496 – 507.
[42] J. Amin, J. Ananthan, R. Voellmy, Key features of heat shock regulatory elements, Mol. Cell Biol. 8 (1988) 3761 – 3769.
[43] P. Christou, Particle Bombardment for Genetic Engi-neering of Plants, R.G. Landes Company, Texas, 1996, pp. 43 – 116.
[44] K.R. Luehrsen, V. Walbot, Intron enhancement of gene expression and the splicing efficiency of introns in maize cells, Mol. Gen. Genet. 225 (1991) 81 – 93.