www.elsevier.comrlocaterapplanim
Deprivation of straw bedding alters PGF -induced
2anesting behaviour in female pigs
T.H.J. Burne
), P.J.E. Murfitt, C.L. Gilbert
MAFF Welfare and BehaÕiour Group, Department of Neurobiology, The Babraham Institute, Babraham,
Cambridge CB2 4AT, UK
Accepted 5 April 2000
Abstract
Sows are highly motivated to build a maternal nest on the day preceding parturition. A model for nest building has been established in pigs, in which exogenously administered prostaglandin
Ž .
F2a PGF2a may be used to elicit nesting behaviour in cyclic, pseudopregnant and pregnant pigs. The aim of this experiment was to examine the effects of deprivation of straw bedding on PGF -induced nest building in pseudopregnant Large White gilts. Oestradiol valerate injections2a
Ž5 mgrday were given on days 11–15 of the oestrous cycle to induce pseudopregnancy. The pigs.
Ž .
were housed individually in a pen 2.8=1.7 m and provided with 2-kg fresh straw each day. On the test day, on day 46 or 47 of pseudopregnancy, half of the pigs were deprived of straw Žsubstrate effect and they were injected intramuscularly with saline or 15 mg of PGF. 2aŽLutalyse,
. Ž .
Upjohn treatment effect allocated in a Latin-square design. Behaviour was recorded onto video tapes for 1 h either side of treatment for analysis using a computerised event recorder. PGF -treated pigs housed in bare or strawed pens showed significantly higher frequencies of2a
pawing and rooting, and stood for longer than saline-treated controls. This treatment effect has been previously shown to be comparable to pre-partum nest building. The removal of straw significantly reduced the frequency of pawing and the duration of rooting by PGF -treated pigs.2a
The results demonstrate that nesting behaviour can be initiated by exogenously administered PGF2a and is further modified by the provision of straw. This suggests that PGF -induced2a
nesting behaviour is subject to environmental feedback.q2000 Elsevier Science B.V. All rights
reserved.
Keywords: Pig, reproduction; Prostaglandin; Periparturient behaviour; Environmental enrichment; Nest
build-ing
)Corresponding author. Tel.:q44-1223-496518; fax:q44-1223-496028.
Ž .
E-mail address: [email protected] T.H.J. Burne .
0168-1591r00r$ - see front matterq2000 Elsevier Science B.V. All rights reserved. Ž .
1. Introduction
The use of straw bedding in pig housing has a number of benefits to pig welfare, providing floor comfort, the opportunity for recreation and addition to bulk in the diet ŽArey 1993 , and is recommended by the Ministry of Agriculture, Fisheries and Food in.
Ž .
the UK MAFF, 1983 . The requirement for straw is different for pigs of different ages and is dependent on their housing and husbandry conditions. Late pregnant and lactating
Ž sows use substrates, including straw, for nest building when housed in extensive Jensen,
. Ž .
1989 and intensive environments Lawrence et al., 1994 . There has been considerable research into the sows’ requirement for straw during the pre-partum period and this is seen as an important issue owing to the widespread commercial use of farrowing crate systems that restrict freedom of movement and do not provide substrates for nest
Ž .
building Arey, 1997 . The increased maternal stress that may result from the use of farrowing crates could be owing to either an inability to access suitable substrates or confinement or a combination of the two. The provision of straw or sawdust to confined
Ž
sows could reduce their frustrated attempts to nest build Cronin et al., 1994; Arey, .
1997 .
Under semi-natural conditions the pre-partum behaviour of sows involves distinct periods of wandering, nest site selection and nest building, culminating in the creation of
Ž . Ž a protected nest in which to give birth to piglets. According to Jensen 1988 later
.
developed in Jensen, 1989, and tested in Jensen, 1993 nest building is divided into two separate phases. The first phase of nest construction is regulated by internal factors and involves preparation of the site, characterised by vigorous nosing, rooting and pawing. The second phase of nest building involves gathering and arranging of nest material and is thought to be regulated by external factors, such as temperature, availability of
Ž .
suitable substrates and sensory feedback Jensen, 1993 .
Ž Sows induced to farrow by peripheral administration of prostaglandin F2a PGF ,2a
.
Lutalyse typically show an initial period of nest building behaviour. This is followed, some 12–24 h later, by a second more intense period of nest building, before the sow
Ž .
gives birth to her piglets 22–36 h after treatment Widowski and Curtis, 1989 . In the day preceding birth levels of the major metabolite of PGF , 13,14-dihydro-15-keto2a
Ž .
PGF2a rise in peripheral plasma Watts et al., 1988; Whitely et al., 1990 . Non-pregnant gilts injected intramuscularly with PGF2a display nest building for at least 1 h after
Ž .
treatment Blackshaw, 1983 . The effect of PGF2a on nesting behaviour is specific to post-pubertal females as PGF2a administered to boars results in displays of sexual
Ž .
behaviour Fonda et al., 1981 and PGF -treated juvenile pigs only show changes in2a
Ž .
activity and discomfort Widowski and Curtis, 1989 . We have previously shown that treatment of pseudopregnant gilts produces a consistent behavioural response on a stable
Ž .
endocrine background Boulton et al., 1997; Burne et al., 1999 . One advantage of using an artificial model of nest building is that the internal cue can be controlled to a much greater extent than using pregnant animals, in which litter size and hormone secretion patterns can vary markedly.
We have previously shown that PGF -induced nest building is altered following2a
environmental disturbance. For example, PGF -treated gilts confined in farrowing2a
Ž penned counterparts given access to straw, despite initial attempts to do so Boulton
.
et al., 1997; Burne et al., 1999 . It is possible that nest building seen using the PGF -induced nesting model is affected by the availability of suitable substrates, such2a
as an earth floor, straw, twigs and branches. However, PGF -treated gilts show similar2a
patterns of nesting behaviours, such as an increase in locomotion followed by increased pawing and rooting at straw, when housed on a concrete floor in a pen with straw ŽBurne et al., 1999, in press , as they do when housed outdoors with access to an earth.
Ž .
floor, straw and other vegetation Gilbert et al., 2000 . Therefore, it may be that the differences we have observed in PGF -induced nesting as a result of environmental2a
Ž .
disturbance confinement in a crate without straw are due to the complete absence of any nesting substrate.
The aim of this experiment is to investigate PGF -induced nesting behaviour in pigs2a
housed under the same conditions but with or without access to straw. We chose to use straw as a nesting substrate, rather than an earthen floor for example, because it is inexpensive, readily available and widely used in commercial husbandry systems. Moreover, because most late pregnant sows in the UK are housed on concrete floors, the provision of a wide range of substrates other than straw is simply not practical. A balanced design to test the hypothesis that straw has an effect on nesting behaviour would require selecting pigs that have either been housed for long periods on a bare floor or provided with daily access to straw. However, we restricted this study to pigs, which had been given daily access to straw. This experiment tested the hypothesis that acute removal of straw would alter the expression of PGF -induced nesting behaviour.2a
This would establish whether the PGF -induced nesting model is sensitive to environ-2a
mental feedback.
2. Methods
2.1. Animals and housing
Large White gilts were selected from the Babraham herd after their first oestrus cycle Ž
at ca. 6 months of age. Trials were run in three separate batches in April, August and
. Ž .
December of 1998 and animals housed in groups of 10 in a large covered pen 4=8 m Ž for the first 3 weeks of the experiment. Twenty-four gilts showing signs of oestrus see
.
below were selected and transferred in three sequential batches of eight to individual
Ž .
pens in a large enclosed fan-ventilated barn 40 days before the test day, see below .
Ž .
Each pen 2.8=1.7 m was constructed with solid walls and a concrete floor subdivided
Ž .
by a step into a raised lying area with under floor insulation 1.8=1.7 m draining onto
Ž .
a dunging area at the rear of the pen 1.0=1.7 m . Gilts were floor fed 1.5 kg pelleted
Ž .
ration Dalgety Agricultural Food, Bury St Edmunds, UK and given 2 kg fresh straw in
Ž .
the lying area once a day after their pen was cleaned 0800–0830 h . Water was available ad libitum from a bite drinker on the wall above the dunging area. Animals
Ž .
handled for 10 min a day while they were housed individually to permit familiarisation with the experimenters.
2.2. Pseudopregnancy
Ž .
Each gilt received an oral dose 20 mgrday of the synthetic progesterone
Al-Ž .
trenogest Regumate Porcine, Hoechst, Milton Keynes, UK for 18 days to synchronise oestrus. The gilts were then run with a teaser boar for 7 days to monitor for signs of oestrus. The first day of standing oestrus is referred to as day 0 of the oestrous cycle. Pseudopregnancy was induced with five daily intramuscular injections of 5 mg
oestra-Ž .
diol valerate Sigma, Poole, UK in 0.5 ml sesame oil on days 11–15 of the oestrous cycle. On days 39–41 of pseudopregnancy, each gilt was fitted with an indwelling jugular catheter as part of a larger experiment. At the end of the experiment, the gilts were given a lethal dose of sodium pentobarbitone into the catheter exactly 65 min after
Ž .
treatment with saline or PGF2a see below and the brains removed for future histochem-ical analysis. Post-mortem inspection on the test day revealed that two gilts had an unenlarged uterus and no corpora lutea and were deemed not to be pseudopregnant. In addition, one gilt had corpora albicana present and was deemed to have broken pseudopregnancy several days prior to the test day. However, the behavioural data from
Ž .
these animals ns1 PGF -treated, n2a s2 saline-treated did not vary significantly from that of the remaining animals and the full data set obtained from 24 animals was analysed together.
2.3. Experimental protocol
Ž . Ž .
Nest building was induced during the morning 1030 h or afternoon 1500 h on day 46 or 47 of pseudopregnancy and this day is referred to as the test day. The pens were cleaned out at 0900 or 1330 h. Half of the gilts had 2 kg fresh straw added to the pen lying area and the remaining gilts were left in a bare pen. This was the first day that gilts tested in a bare pen had been without straw in the experiment. All gilts received an
Ž .
intramuscular injection randomly into either the left or right side of the neck of 15 mg
Ž .
PGF2a Lutalyse, Upjohn, Crawley, UK or 3 ml saline at either 1030 or 1500 h. Each animal was tested once only. All treatments were allocated in a Latin-square design Žtreatment, substrate, time of day to give n. s6 animals in each of the following groups; saline-treated without straw, saline-treated with straw, PGF -treated without straw and2a
PGF -treated with straw.2a
2.4. BehaÕioural obserÕations
Ž .
The gilts behaviour was recorded for 1 h before pre-treatment to 1 h after treatment
Ž .
with saline or PGF2a post-treatment on the test day by video cameras attached to time-lapse video recorders recording at 24 framesrs. We decided to observe behaviour for only 1 h post-treatment as this period of time has been shown previously to include
Ž
peak levels of nesting activity in pigs given PGF2a Blackshaw, 1983; Widowski and .
Ž . previous studies, in which a single overhead camera was used e.g. Boulton et al., 1997 . The additional cameras at the corners of the pen permitted more detailed observation of the gilts’ behaviour. A microphone was also positioned above the pen to record the gilts’ vocalisations, which were scored as one of three different types of call: grunt, squeal or woof. The onset and duration of each behaviour was scored using the Observer
Ž .
computer package Noldus, 1991 and they are defined in Table 1.
2.5. Statistical analyses
The behavioural data were quantified using the elementary statistics option of the
Ž .
Observer computer package Noldus, 1991 . The behaviours were pooled into the following categories for analysis; stand, lie, change posture, inactive, root, paw, gather,
Ž .
scratch, essential and vocalise see Table 1 . Based on previous studies, the key nesting Ž
behaviours measured included root, paw and gather Widowski and Curtis, 1989; Jensen, .
1993; Burne et al., 1999 . We chose to analyse the post-treatment behavioural data in
Table 1
Behavioural ethogram Posture
Ž .
Stand duration : standing on all four legs
Ž .
Lie duration : lying on the belly
Ž .
Change posture frequency : moving between postures — sit, kneel, lie and stand Behaviour
Ž .
Inactive duration : no part of the pig is moving
Ž .
Root duration
Forage: oro-nasal contact with straw whilst chewing
Root straw: oro-nasal contact with straw and head-movement but without chewing Root floor: oro-nasal contact with floor and head-movement
Root wall: oro-nasal contact with wall and head-movement
Ž .
Chew fixture: chewing at any of the fixtures within the pen apart from the drinker
Ž .
Paw frequency
Paw straw: raking movement of either foreleg at an area of floor covered in straw Paw floor: raking movement of either foreleg at an area of floor devoid of straw Paw wall: raking movement of either foreleg at the walls of the pen
Ž .
Gather duration
Lift straw: grasping straw in the mouth and raising the head without locomotion Carry straw: grasping straw in the mouth and taking at least two steps
Ž .
Scratch frequency
Object scratch: scratch or rub any part of the body against another object
Rear leg scratch: scratch or attempt to scratch, any part of the body with the rear leg
detail. The data were non-normally distributed and non-parametric statistical procedures
Ž .
were used Siegel and Castellan, 1988 . The data sets for frequency per hour and Ž .
percentage duration were analysed using a Kruskal–Wallis test H and, where this indicated significant differences between groups, was followed by post-hoc Mann
Ž Whitney tests. Behaviours which were only possible in animals supplied with straw e.g.
.
gather were analysed with Mann Whitney tests alone.
Ž
Preliminary analysis of the data for two measures of activity proportion of time .
inactive and proportion of time standing in the hour pre-treatment indicated that the
Ž . Ž
time of the injection morning or afternoon was not a significant factor U)49,
. Ž
P)0.2 . Thus, the data were pooled for time of day ns12 tested in the morning and .
ns12 tested in the afternoon . Variation was seen in environmental temperature between the test day for each batch of pigs. The temperature within the barn was measured at the time of injection and there was a significant difference in the mean
Ž Ž .
temperature between the separate batches of pigs mean temperature"SEM 8C ; batch 1: 15.5"0.5; batch 2: 23.5"1.3; batch 3: 11.1"0.5; F2,23s48.8, P-0.001; one-way
.
analysis of variance . However, there was no overall significant difference between Ž
groups salinerno straw: 16.8"2.08C; salinerstraw: 16.8"2.98C; PGF rno straw:
2a
.
15.2"2.08C; PGF rstraw: 18.0"2.78C; F s1.3, Ps0.3 and so temperature was
2a 3,23
not included as a variable in the analysis.
wŽ . Ž . Ž . x Ž .
Fig. 1. Median scores for nesting behaviour, a gather, b paw and c root and duration of time inactive d by pigs housed in a bare or a strawed pen of the same size. The error bars indicate the interquartile range
Žlower bar: 25th percentile, upper bar: 75th percentile . Data are presented for the hour post-treatment with.
Ž . Ž . Ž .
3. Results
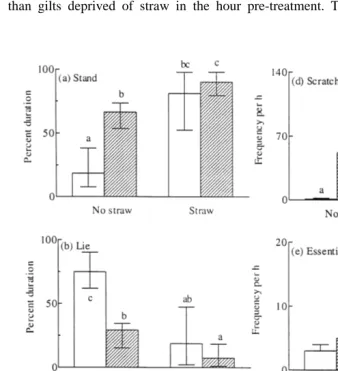
Acute deprivation of straw resulted in gilts vocalising more and lying down for longer periods of time than gilts that were given their usual ration of straw. As expected, gilts with access to straw continued to root at the straw and remained active for longer than gilts deprived of straw in the hour pre-treatment. This provided an appropriate
w Ž . Ž . Ž .x
Fig. 2. Median scores for postures, stand a , lie b and change posture c , and the frequencies of scratching
Ž .d , essential behaviours e and vocalising f by pigs housed in a bare or a strawed pen of the same size. TheŽ . Ž .
Ž .
error bars indicate the interquartile range lower bar: 25th percentile, upper bar: 75th percentile . Data are
Ž . Ž .
presented for the hour post-treatment with saline open bars or PGF2a hatched bars . Bars with different
Ž .
background with which to investigate the effects of PGF2a and saline on behaviour. There was a significant effect of straw and PGF -treatment on all nesting behaviours2a
measured in the hour post-treatment. PGF -treated gilts housed with straw had the2a
Ž . Ž
highest frequencies of gather Us1, Ps0.005, Fig. 1a , paw Hs19.2, Ps0.0002,
. Ž .
Fig. 1b and root Hs18.8, Ps0.0003, Fig. 1c . The scores for root and paw were higher in PGF -treated gilts with access to straw than PGF -treated gilts housed in a2a 2a
Ž .
bare pen Us5, Ps0.04 . PGF -treated pigs without access to straw rooted for a2a
Ž . Ž
longer duration Us4, Ps0.03 and had a higher frequency of pawing Us0, .
Ps0.002 than saline-treated pigs housed without straw. Saline-treated pigs housed in a
Ž . Ž .
straw pen had higher values for root Us0, Ps0.004 and paw Us3, Ps0.007 than saline-treated pigs housed in a bare pen. The duration of time gathering straw was
Ž .
higher in PGF -treated than saline-treated pigs2a Us1, Ps0.005 . There was a significant effect of treatment and housing condition on the amount of time spent
Ž .
inactive Hs13.7, Ps0.0003, Fig. 1d ; saline-treated pigs without access to straw
Ž .
were inactive for 10 times longer than pigs in the remaining groups Us2, P-0.01 . Saline-treated gilts housed in a bare pen were observed standing for the shortest
Ž . Ž
duration Hs12.0, Ps0.007, Fig. 2a and lying for the longest duration Hs12.0, .
Ps0.007, Fig. 2b . PGF -treatment resulted in significantly higher frequencies of2a
Ž . Ž .
postural change Hs12.3, Ps0.006 and scratching Hs17.6, Ps0.0005 , irre-spective of housing condition. There was no significant effect of treatment or straw on
Ž .
essential behaviours Hs2.6, Ps0.4 . There was a significant effect of both substrate
Ž .
and treatment on the frequency of vocalisations Hs8.4, Ps0.04 , in which saline-treated gilts housed in a bare pen vocalised the most and PGF -saline-treated gilts in a strawed2a
pen vocalised the least.
4. Discussion
This experiment demonstrates that depriving pigs of straw does not abolish PGF -in-2a
duced nesting behaviour. The two main components of nest building, pawing and rooting appear to be tightly controlled by internal cues. These observations are in
Ž
agreement with data obtained from pregnant sows Lawrence et al., 1994; Hutson, .
1992 , which have a low responsiveness to straw while they are motivated to nest build. However, the gilts in this study performed more pawing and rooting in a strawed pen, indicating that they will use substrates if they are available. Thus, acute removal of straw altered the expression of PGF -induced nesting behaviour and these data show2a
that the PGF -induced nesting model is sensitive to environmental feedback.2a
The sequence of events seen in PGF -induced nest building by pigs housed in a2a
farrowing crate without access to straw shares some similarities with that found in the present experiment for pigs housed in a bare pen. That is, pigs without access to straw in crates or pens do not display as much nest building behaviour when compared with
Ž .
penned counterparts given access to straw Boulton et al., 1997; Burne et al., 1999 . However, there are several differences between the nesting behaviours of PGF -treated2a
Ž .
Ž .
of treatment Burne et al., 1999 . By contrast, penned pigs continue to move around the pen and show higher levels of rooting. Pregnant sows housed in pens also show more
Ž .
nest building behaviour than sows housed in crates Lawrence et al., 1994 , although the levels seen in confined pseudopregnant pigs is even lower than in confined pregnant sows. Therefore, it seems likely that confined pseudopregnant gilts are less motivated to nest build than unrestricted pseudopregnant gilts. In the PGF -induced model, there-2a
Ž .
fore, a minimum amount of space at least enough room to turn around completely and
Ž .
an available substrate straw are key requirements for uninhibited expression of all aspects of nesting behaviour.
Although there are some differences between parturition-induced and PGF -induced2a
nest building, many of the behaviours are common to both. These include pawing, rooting and gathering nest material, as well as increases in locomotion, possibly as an
Ž
aid to isolation andror nest site selection Boulton et al., 1997; Burne et al., 1999; .
Gilbert et al., 2000 . However, the nesting behaviour of PGF -treated gilts occurs over2a
Ž .
a much shorter duration 2–4 h; Burne et al., 1999 when compared with pre-partum
Ž .
nest building 6–12 h; Widowski et al., 1990 , allowing for the possibility that some behaviours that are normally in sequence might overlap. One explanation for this mismatch is that PGF2a is involved in stimulating many of the components of nesting
Ž .
behaviour wandering, gathering, pawing and rooting but the bolus injection procedure used in the PGF2a model does not mimic pre-partum hormonal levels adequately. However, taken together these data suggest that PGF2a is a major underlying component of all stages of pre-partum nest building.
There was a significant variation in environmental temperature between batches of pigs. However, because of the balanced design used in this study, there were no differences in average temperature at test between the treatment groups. Jensen’s model ŽJensen, 1993 predicts that nest building behaviours would be differentially affected by. changes in ambient temperature, although this has not been tested systematically. As we did not systematically vary temperature in this experiment, we can provide no evidence for the effect of temperature on PGF -induced nesting behaviour. However, this is2a
currently being investigated.
Pigs choose to spend a large amount of time foraging if suitable substrates are available and the provision of straw to pigs can overcome problems of boredom and can
Ž .
reduce the incidence of stereotypes Arey, 1993; de Jong et al., 1998 . In the present study, gilts with access to straw spent much of their time engaged in foraging and rooting activities, whereas gilts deprived of straw had higher frequencies of vocalising, chewing at the fixtures, rooting the wall and floors, and lay down for longer periods of time. One limitation with this study is that the design was unbalanced in the sense that all pigs had access to straw prior to the test day. We did not observe animals that had been reared for long periods on bare concrete. To establish whether the PGF -induced2a
nesting model is sensitive to environmental feedback, one needs only to show that removal of straw alters the expression of PGF -induced nesting behaviour. A closer2a
estimation of the effects of straw manipulation on PGF -induced nesting behaviour2a
the pen walls and other pigs, whereas pigs in substrate-enriched housing spend more
Ž .
time exploring substrates de Jong et al., 1998 . Therefore, pigs redirect their exploratory
Ž .
behaviour towards the pen fixtures in the absence of straw after both the acute hours
Ž .
and chronic months absence of straw. Behavioural effects of PGF2a in animals with or without straw must therefore be viewed as being superimposed upon unstimulated behaviour, which differs between groups. It should also be noted that in the present experiment, to avoid confusion with movement and functional descriptors, no distinction
Ž
is made between different types of some behaviours e.g. forage, root straw and root .
floor so behaviours with potentially separate underlying motivations have been added together.
PGF2a has a wide range of effects, which appear unrelated to nesting behaviour, on a number of different tissues, such as blood vessels, cardiac and smooth muscle, gut, eye
Ž .
and brain Samuelsson et al., 1978 . Some of the observed behavioural responses, such as scratching, may have resulted from physiological changes mediated by PGF2a on these or other systems. It is clear that scratching, particularly at these high levels, is not part of ‘normal’ pre-partum behaviour, and may be as a result of supra-physiological doses following the bolus intramuscular injection procedure. It is, however, interesting
Ž .
to note that sows treated with 10 mg of PGF2a to induce parturition on day 112 of pregnancy display nesting behaviour but are not reported to show elevated levels of
Ž .
scratching Widowski and Curtis, 1989 . The reason for the discrepancy between pregnant and non-pregnant pigs is not known.
In conclusion, female pigs given PGF2a are highly motivated to perform nesting behaviours and if substrates are available they will use them. The underlying motivation for nest building does not appear to differ between a pre-partum sow and pseudopreg-nant gilt treated with PGF , suggesting that prostaglandins are directly involved in this2a
response. We suggest that allowing gilts access to straw would be beneficial for the sows’ welfare while they are motivated to nest build.
Acknowledgements
Support for this research was provided by the Ministry of Agriculture, Fisheries and
Ž .
Food commission AW0118 . The authors thank the Large Animal Unit staff for care of the pigs. They also thank Drs. A.N.B. Johnston and R.F. Parrott for valuable comments on the manuscript.
References
Arey, D.S., 1993. The effect of bedding on the behaviour and welfare of pigs. Anim. Welfare 2, 235–246. Arey, D.S., 1997. Behavioural observations of peri-parturient sows and the development of alternative
farrowing accommodation: a review. Anim. Welfare 6, 217–229.
Blackshaw, J.K., 1983. Prostaglandin F2a induced nest building behavior in the non-pregnant sow, and some welfare considerations. Int. J. Stud. Anim. Probl. 4, 299–304.
Burne, T.H.J., Murfitt, P.J.E., Goode, J.A., Boulton, M.I., Gilbert, C.L., 1999. Effects of oestrogen supplemen-tation and space restriction on PGF -induced nest-building in pseudopregnant gilts. Anim. Reprod. Sci.2a 55, 255–267.
Burne, T.H.J., Murfitt, P.J.E., Gilbert, C.L., 2000. Behavioural responses to intramuscular injections of
Ž .
prostaglandin F2ain female pigs. Pharmacol. Biochem. Behav. in press .
Cronin, G.M., Smith, J.A., Hodge, F.M., Hemsworth, P.H., 1994. The behaviour of primiparous sows around farrowing in response to restraint and straw bedding. Appl. Anim. Behav. Sci. 39, 269–280.
de Jong, I.C., Ekkel, E.D., Van de Burgwal, J.A., Lambooij, E., Korte, S.M., Ruis, M.A.W., Koolhaas, J.M., Blokhuis, H.J., 1998. Effects of strawbedding on physiological responses to stressors and behavior in growing pigs. Physiol. Behav. 64, 303–310.
Fonda, E.S., Diehl, J.R., Barb, C.R., Kiser, T.E., Kraeling, R.R., Rampacek, G.B., 1981. Serum luteinizing-hormone, testosterone, prolactin and cortisol concentrations after PGF2a in the boar. Prostaglandins 21, 933–943.
Gilbert, C.L., Murfitt, P.J.E., Boulton, M.I., Pain, J., Burne, T.H.J., 2000. Effects of prostaglandin F2a treatment on the behaviour of pseudopregnant pigs in an extensive environment. Horm. Behav. 37, 229–236.
Hutson, G.D., 1992. A comparison of operant responding by farrowing sows for food and nest-building materials. Appl. Anim. Behav. Sci. 34, 221–230.
Jensen, P., 1988. Maternal behavior and mother-young interactions during lactation in free-ranging domestic pigs. Appl. Anim. Behav. Sci. 20, 297–308.
Jensen, P., 1989. Nest site choice and nest building of free-ranging domestic pigs due to farrow. Appl. Anim. Behav. Sci. 22, 13–21.
Jensen, P., 1993. Nest building in domestic sows: the role of external stimuli. Anim. Behav. 45, 351–358. Lawrence, A.B., Petherick, J.C., McLean, K.A., Deans, L.A., Chirnside, J., Vaughan, A., Clutton, E.,
Terlouw, E.M.C., 1994. The effect of environment on behavior, plasma-cortisol and prolactin in parturient sows. Appl. Anim. Behav. Sci. 39, 313–330.
Ž .
Ministry of Agriculture, Fisheries and Food MAFF , 1983. Codes of Recommendations for the Welfare of Livestock Pigs. HMSO, London.
Noldus, L.P.J.J., 1991. The Observer: a software system for collection and analysis of observational data. Behav. Res. Methods Instrum. Comp. 23, 415–429.
Samuelsson, B., Goldyne, M., Granstrom, E., Hamberg, M., Hammarstrom, S., Malmsten, C., 1978.¨ ¨
Prostaglandins and thromboxanes. Annu. Rev. Biochem. 47, 997–1029.
Siegel, S., Castellan, N.J., 1988. Nonparametric Statistics for the Behavioral Sciences. 2nd edn. McGraw-Hill, New York.
Watts, A.D., Flint, A.P.F., Foxcroft, G.R., Porter, D.G., 1988. Plasma steroids, relaxin and dihydro-keto-prostaglandin F2achanges in the minipig in relation to myometrial electrical and mechanical activity in the prepartum period. J. Reprod. Fertil. 83, 553–564.
Whitely, J.L., Hartmann, P.E., Willcox, D.L., Bryant-Greenwood, G.D., Greenwood, F.C., 1990. Initiation of parturition and lactation in the sow: effects of delaying parturition with medroxyprogesterone acetate. J. Endocrinol. 124, 475–484.
Widowski, T.M., Curtis, S.E., 1989. Behavioral responses of periparturient sows and juvenile pigs to prostaglandin F2a. J. Anim. Sci. 67, 3266–3276.