Effects of soil compaction on the relationships between nematodes,
grass production and soil physical properties
L.A. Bouwman
a,∗, W.B.M. Arts
baAlterra, Green World Research, P.O. Box 47, 6700 AA Wageningen, The Netherlands
bDLO Research Institute for Agricultural and Environmental Engineering, P.O. Box 43, 6700 AA Wageningen, The Netherlands
Received 18 October 1997; received in revised form 24 February 2000; accepted 1 March 2000
Abstract
As farm machinery has become heavier, concern has grown about its direct effects on soil physical conditions and its indirect effects on crop yields and soil biota. To study the relationships between these parameters, non-grazed temporary grassland plots on a loamy sand soil were subjected to full-width load traffic with widely different loads (0, 4.5, 8.5 and 14.5 t) one to four times per year for a period of 5 years. Soil bulk density was monitored as an indicator of soil compaction. Grass yield was measured throughout the experimental period. Root distribution over the soil profile and nematodes populations were assessed during the final year of the experiment. Results indicate that a moderate degree of compaction (∼4.5 t load) gave the highest crop yield and that at higher degrees of compaction roots failed to penetrate into the deeper soil layers (>20 cm depth). Total numbers of nematodes were not affected by compaction, but their distribution over the various feeding types shifted towards a population with increased numbers of herbivores and decreased numbers of bacterivores and omnivores/predators. This change in the structure of the nematode assemblage is associated with poorer conditions for crop growth. © 2000 Elsevier Science B.V. All rights reserved.
Keywords: Grass production; Habitable pore space; Nematode trophic structure; Root distribution; Soil bulk density; Soil compaction
1. Introduction
Soil compaction is an important form of physical soil degradation (Van Ouwerkerk and Soane, 1994). The use of heavy load traffic (machinery) in modern agriculture causes substantial soil compaction, coun-teracted by soil tillage that loosens the soil. Higher soil bulk densities affect resistance to root penetration, soil pore volume and permeability to air, and thus, finally the pore space habitable for soil organisms. Through a combination of these factors, soil compaction will
∗Corresponding author. Tel.:+31-317-474352; fax:+31-317-424812.
E-mail address: [email protected] (L.A. Bouwman)
affect crop growth (Brussaard and Van Faassen, 1994; Soane and Van Ouwerkerk, 1994). Soil pores also ac-commodate biological processes such as organic mat-ter decomposition and nutrient mineralization (Elliott et al., 1980; Hassink et al., 1993). Also, herbivorous organisms feeding on living plant roots live in the interstitial spaces present in the soil. Conventionally, fields are regularly tilled for arable crop production and the use of heavy load machinery creates a plow layer with increased bulk density, usually below 25 cm depth, and with a decreased bulk density in the upper soil layers. The passage of heavy load traffic on peren-nial grassland also causes compaction of the upper soil layers but there are no tillage operations to counter-act this compcounter-action. In perennial and temporary (ley)
grasslands, compaction affects many physical and bio-logical soil properties and processes by various direct and indirect mechanisms. To unravel these, we should know the relationships between the load of the passing traffic and the resulting soil compaction, bulk density, root distribution over the soil profile, crop yield, and finally the structure of the community of soil organ-isms and its functioning. This knowledge may help farmers in their choice of machinery load, the width and the pressure of the tires and the frequency of traf-fic, depending on soil type, weather conditions, and acceptable degree of compaction (Arts et al., 1994).
Nematodes are involved in a wide range of activities and connections to other organisms in the soil food web, comprising species feeding on living roots (her-bivores (H)), on microorganisms (bacterivores (B); fungivores (F)), and species feeding on larger organ-isms (predators (P)) or on mixed diets (omnivores (O)). Nematode communities in permanent grasslands are relatively stable with respect to numbers and have a high species diversity. Numbers of specimens in trophic groups often decrease in the order B>F> H>O/P (Boström and Sohlenius, 1986). The various nematode trophic groups reflect important microbio-logical and phytopathogenic processes such as organic matter decomposition, nutrient mineralization and her-bivory. Although numbers of nematodes belonging to a particular feeding category are not always correlated to the amount of specific food, numbers are correlated with process rates, e.g. numbers of bacterivorous ne-matodes correlate with bacterial production (Clarholm et al., 1981) and N mineralization rate, in particular, in sandy soils with pore diameters between 30 and 90mm (Hassink et al., 1993), and herbivores correlate
with root growth, etc. Soil compaction affects nema-todes directly as well as indirectly. Vulnerable species can be damaged during passage of the traffic (Boag, 1985); decreased habitable pore space after passage of the traffic affects probably all taxa except those that are intimately connected with root development, and finally root dynamics in particular affects the numbers of herbivores and some root-generated microbivorous taxa (Griffiths et al., 1991; Bouwman et al., 1993). The effect of tillage and compaction regimes on total numbers and dynamics of nematodes can therefore not be generalized, but feeding categories and taxa should be examined individually (Fortnum and Karlen, 1985; Parmelee and Aston, 1986; Thomas, 1978).
Trophic differences among nematode populations were observed to be indicative of variations in crop yields (Edwards, 1988; Edwards and Kimpinski, 1997) and also of decomposition and, in particular, N mineralization processes (Hassink et al., 1993).
A field experiment was carried out to study the relationships between traffic load and physical, bio-logical and (crop) production characteristics of the soil. In 1988 the experiment (Arts et al., 1994) was established on a marine loamy sand soil in The Netherlands, where grass was sown on plots (strips) previously tilled to a depth of 50 cm and consequently subjected to different degrees of soil compaction over the whole surface of the plots. Soil physical characteristics as well as crop yields were measured throughout the 5-year trial period and finally, in the last year of the experiment (1992), the distribution of the roots over the soil profile and the nematode fauna were recorded in two plots with different degrees of compaction. It was hypothesized that compaction may have positive as well as negative effects on crop yield and thus, a site-specific optimum degree of soil compaction exists. Single season changes in soil management regimes have little lasting effect on nematode communities (Baird and Bernard, 1984; Sohlenius and Sandor, 1989), but the nematode fauna in the upper 10 cm of the grass-sod of the experimen-tal plots is assumed to be in a new equilibrium after 4 years of growth under experimental conditions.
2. Materials and methods
2.1. The experimental site
The research was carried out at the Oostwaardhoeve experimental farm (longitude 52◦18′N, latitude 4◦85′ E) in the Wieringermeer, a reclaimed polder, used for agricultural production since 1934. The soil is a cal-careous loamy (30% silt) marine sand (M50: 50–105), containing 5% organic matter (Arts et al., 1994).
2.2. The establishment of the field experiment
equal size, on which four plots (strips) were assigned (180 m×2 m) to four loading treatments: 0, 4.5, 8.5 and 14.5 t. For this purpose, a loading frame with a steel roller, 2 m wide and 1.2 m in diameter, carrying the various loads passed over the plots; the roller was drawn by a tractor with 3 m trail-width. After the first compaction treatment perennial ryegrass, Lolium
perenne (mixture BG3) was sown, and harvested up
to four times annually. After each harvest the specific traffic loads passed over the various plots. A load of 1 t exerts a pressure of 100–500 kg per dm2, depend-ing on the actual compactability of the upper soil layer (e.g. dry versus wet conditions).
The experiment was extended with different rates of nitrogen fertilization (0, 140, 280 and 420 kg N ha−1 per year) within various subplots.
2.3. Physical measurements
Over the period 1988–1992, soil porosity, perme-ability and penetrperme-ability were reduced. To characterize the soil condition, measurements were carried out at different depths, after each harvest: thickness of the initially loosened layer, soil bulk density, penetration resistance, and the soil moisture content (measured weekly).
To measure the thickness of the loosened layer, in each plot five tiles (30 cm×30 cm) were buried on top of the undisturbed subsoil, i.e. at a depth of 60 cm, after rotodigging in 1988. The thickness of the soil layer above these tiles was measured as well as the position of the tile with regard to a fixed reference point. This latter measurement was necessary to correct the first one for sinking of the tile.
Soil bulk density was measured in the 15–20 and 30–35 cm layers below the soil surface, in undisturbed ring samples containing 100 ml soil.
The penetration resistance was measured with a Bush Penetrometer conus type B, ASAE Standards, 1993. This parameter, expressed as average val-ues of 10 penetrations per event in the soil layers 0–5, 5–10, 10–15 cm, etc. indicates the pressures roots have to exert to overcome soil compaction, effective root-zone depth (Gregory, 1988), as well as soil porosity and compaction (Groenevelt et al., 1984).
Soil moisture contents (w/w) in different soil layers, including the 0–5 cm layer, were measured weekly in
mixed samples, originating from 10 subsamples, by overnight drying 20 g soil at 105◦C.
2.4. Crop measurements
Grass yield and distribution of grass roots over the soil profile were measured. Crop responses are likely to be directly related to properties such as pore size distribution, permeability, aeration, etc. and indirectly to bulk density and compaction.
Grass dry matter yield was determined after each harvest of 20 m×1.2 m strips. Annual grass yields are the sums of three or four harvests.
The distribution of the roots over the soil profile was measured once by the pin-board method (Böhm, 1979) in June 1992, in the 4.5 and 14.5 t treatments, fertilized with 280 kg N ha−1per year.
2.5. Nematode densities
Nematodes were isolated from the soil in 1992, the fifth year of the experiment, in March, June, September, and December, in the 4.5 and 14.5 t treatments (280 kg N). The organisms were isolated from 3×100 g fresh soil from the upper 10 cm with Oostenbrinks’s elutriation method (Jacob and Van Bezooyen, 1984). Total numbers were counted and expressed per 100 g dry soil; the figures presented are averages of three samples. Sixty specimens per sam-ple were identified to genus level and assigned to the following feeding types: bacterivores (Rhabditidae; Cephalobidae, including genera such as Cephalobus,
Acrobeloides, Acrobeles, Chiloplacus but also a small
number of non-cephalobid Teratocephalidae; Monhys-teridae including a small number of Prismatolaimidae; Plectidae; Panagrolaimidae; Alaimidae), fungivores (Aphelenchoididae including mainly Aphelenchoides and few Aphelenchus avenae; Tylenchidae; Psilenchi-dae), herbivores (Paratylenchidae; Pratylenchidae) and omnivores/predators (Dorylaimidae).
2.6. Statistical analysis
3. Results
3.1. Physical measurements
The thickness of the initially loosened layer of 60 cm depth had already stabilized during the first year (1988) of the experiment, and only fluctuated season-ally, due to moisture content in the subsequent years. Thickness decreased monotonously with load (from 0→14.5 t, it changed from 55→43 cm) (Fig. 1a).
Soil bulk density, not measured in the sod but in the 15–20 and 30–35 cm depth layers, increased with load treatment during the first 2 years, and sub-sequently stabilized; in the 0 t treatment (15–20 cm layer) bulk density only increased slightly in the last 2 years of the experiment (1.10→1.15 g cm−3). In the 4.5 t treatment the bulk density in the 15–20 cm layer increased from 1.10 g cm−3in the loosened soil in 1988 to 1.41 g cm−3in 1992.
The resistance to penetration in the various soil lay-ers varied between seasons along with soil moisture, with the highest values in summer (dry) and the lowest in winter (wet conditions). The resistance increased in all load treatments from 1988 to 1989 and then stabilized (Fig. 1b).The resistance also increased with soil depth but only in the upper soil layers. Resistance was highest in the 14.5 t treatment and lowest in the 0 t treatment. In the 0–5 cm layer the lowest values were measured in the 0 t treatment in winter 1991, (0.55 MPa) and the highest in the 14.5 t treatment in summer 1991 (4 MPa; Fig. 1b); the latter resistance was much too high to be broken by monocotyledons (Dexter, 1986).
Soil moisture content in the 0–5 cm layer showed a considerable increase parallel to the weight of the load-traffic (Fig. 1c). Moisture contents in the 14.5 t treatment were up to 30% higher than in the 0 t ment. In winter moisture contents in the 14.5 t treat-ment in the upper 5 cm soil layer approached to 50% (w/w), decreasing down to 10% in summer.
3.2. Crop measurements
Above ground dry matter yields as affected by N-application and load-treatment are presented in Fig. 2. The effect of the load-treatment was signif-icant (p<0.01) and the N-response/load-treatment interaction effect was also significant (p<0.01), but
not when values for 368 kg N and 0 t load were left out of the calculations; this means that in practice the loss of yield due to soil compaction was independent of the level of N application (Keen, 1993).
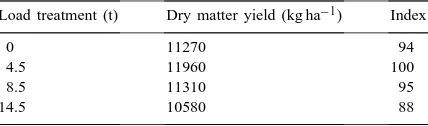
In all years, grass yield was highest in the 4.5 t treat-ment (Fig. 2), on average it was 12,000 kg dry matter per hectare (280 kg N plots) and it was 5, 6 and 12% less in the 8.5, 0 and 14.5 t treatments, respectively (Table 1).
The distribution of roots over the soil profile dif-fered significantly between the 4.5 and 14.5 t plots, although the total mass of root dry matter per hectare was the same in both treatments, 3.6 t (Fig. 3). In the most compacted soil (14.5 t) almost no roots had pen-etrated the soil below 20 cm depth, while in the 4.5 t treatment more than 20% of the total root mass was found between 20 and 50 cm depth. Consequently, root density in the upper soil layers was considerably lower in the less compacted soil.
3.3. Nematode densities
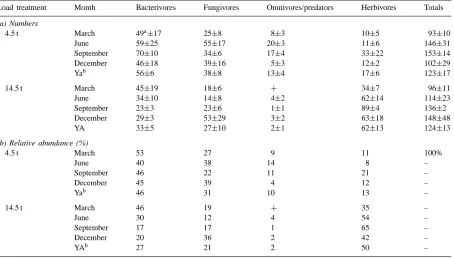
After 4 years of grass growth, total numbers of nematodes in the slightly (4.5 t), and in the heavily (14.5 t) compacted soil did not differ significantly throughout the year, and amounted to 123 and 124 ind. per gram fresh soil, respectively (Table 2). How-ever, numbers of bacterivores, herbivores and omni-vores/predators were significantly different under both compaction regimes at all sampling dates. The effects of compaction were tested by analysis of variance: numbers of bacterivores in heavily (14.5 t) compacted soil were significantly (p<0.035) lower than in the less (4.5 t) compacted soil, fungivores were not significantly lower in the 14.5 t treatment than in the 4.5 t treatment, herbivores were significantly (p<0.0001) higher in the 14.5 t treatment than in the 4.5 t treatment and finally omnivores/predators were significantly (p<0.0001) lower in the 14.5 t treatment than in the 4.5 t treatment. In the heavily compacted soil the average proportion of the herbivores (almost all Paratylenchus sp.) was ca. 50%, and in the slightly compacted soil 13%. Numbers and proportions of the other feeding types were much higher in the less compacted soil, in particular the omnivores/predators and the bacterivores.
Fig. 2. Effect of various load treatments on above ground dry matter yield of grass for four nitrogen fertilizer rates in 1992.
Table 1
Average annual dry matter yield (kg ha−1 per year) over 1988–
1992, under four load treatments (280 kg N ha−1per year)
Load treatment (t) Dry matter yield (kg ha−1) Index
0 11270 94
4.5 11960 100
8.5 11310 95
14.5 10580 88
heavily compacted soil, while the fungivorous
Aphe-lenchoides sp. was equally abundant in both plots. All
other taxa were more numerous in the less compacted soil, with the exception of Panagrolaimus sp.,
Aphe-lenchus avenae, PsiAphe-lenchus sp., and PratyAphe-lenchus sp.,
taxa that only occurred in very small numbers, less than two specimens per gram soil (Table 3).
4. Discussion
Passage of load traffic decreased the thickness of the loosened soil layer considerably and already after the first year of the experiment the soil had stabilized into a physical condition where loading and bearing ca-pacity were in equilibrium. Thickness of the loosened layer, bulk density and penetration resistance clearly depended on the load traffic. In all years the 4.5 t load-ing treatment gave the highest grass yields. Obviously,
a relatively low degree of soil compaction improved physical conditions for crop growth, e.g. created a bet-ter root–soil contact (Van Noordwijk and De Willigen, 1984), whereas at higher degrees of compaction roots failed to penetrate deeper soil layers and consequently exploited only the upper 20 cm of soil (Fig. 2b). As a result the optimum for crop yield was observed at the intermediate (4.5 t) treatment. The lower moisture content in the upper soil layers of the less compacted plots may have affected the organisms in that layer, in particular, in summer when moisture contents dropped to ca. 10% (w/w) (Bouwman and Zwart, 1994). How-ever, the nematodes in the dryest plots (4.5 t) were most numerous in June and September, and numbers were even higher than in the 14.5 t treated plots. Con-sequently, possible negative effects of drought do not seem to have been important. As the moisture con-tents in the lower soil layers do not significantly differ between differently compacted plots, effects on crop growth are assumed to be insignificant.
Fig. 3. Effect of two load treatments on cumulative dry matter weight of roots as a function of depth for one nitrogen fertilizer rate (280 kg N ha−1per year).
Table 2
Abundance of nematodes (feeding groups) in the upper 10 cm of the sod, in numbers per gram dry soil (a) and relative abundance (b), in the 4.5 and 14.5 t load treated plots (280 kg N), at four sampling dates in 1992
Load treatment Month Bacterivores Fungivores Omnivores/predators Herbivores Totals
(a) Numbers
4.5 t March 49a±17 25±8 8±3 10±5 93±10
June 59±25 55±17 20±3 11±6 146±31
September 70±10 34±6 17±4 33±22 153±14
December 46±18 39±16 5±3 12±2 102±29
Yab 56±6 38±8 13±4 17±6 123±17
14.5 t March 45±19 18±6 + 34±7 96±11
June 34±10 14±8 4±2 62±14 114±23
September 23±3 23±6 1±1 89±4 136±2
December 29±3 53±29 3±2 63±18 148±48
YA 33±5 27±10 2±1 62±13 124±13
(b) Relative abundance (%)
4.5 t March 53 27 9 11 100%
June 40 38 14 8 –
September 46 22 11 21 –
December 45 39 4 12 –
Yab 46 31 10 13 –
14.5 t March 46 19 + 35 –
June 30 12 4 54 –
September 17 17 1 65 –
December 20 36 2 42 –
YAb 27 21 2 50 –
Table 3
Average annual abundance’s of nematode taxa in numbers per gram dry soil in the 4.5 and 14.5 t load treated plots in 1992
Nematode taxa Load treatment
aSignificantly different numbers in the two plots (p<0.05).
high densities in widely different habitats (Yeates, 1979; Boström and Sohlenius, 1986; Dmowska, 1993). The identified taxa are very common in grass-lands, and most of them also in arable soils. Thus, the nematode faunas, developed under the compaction gradient, were not exceptional. The faunistic (feed-ing guilds) differences between the two investigated plots, however, were most striking. These differences resembled the differences due to soil tillage observed by Parmelee and Aston (1986) and Juma and Mishra (1988) who measured considerably higher densities of bacterivores and lower densities or proportions of herbivores in tilled versus non-tilled plots, while fungivores differed seasonally between the plots but not on an annual basis. Omnivores/predators are of-ten the most sensitive feeding category with respect to tillage, as found by Wasilewska (1979) comparing annual cropping systems with pastures. Comparison of the results of our experiment with the mentioned literature shows that with respect to the feeding guilds the less compact soil resembles the tilled arable plots and the heavily compacted soil resembles the non-tilled plots. It is often mentioned that soil texture, pH, water content, and crop rotation, show the best
numbers were considerably lower in the heavily compacted soil than in the slightly compacted soil. This is probably due to the third relevant factor: (3) The decrease of pore space habitable for nematodes, caused by increased soil bulk density. Nematodes living primarily in the soil (macro) pores (Jones and Thomasson, 1976) reacted negatively to the decreased habitable pore space. Bacterivorous Cephalobidae oc-cupy a position in both the rhizosphere and in the soil pores, and their numbers did not differ significantly between the 4.5 and 14.5 t treated plots.
It is evident that a certain degree of soil compaction is necessary to obtain optimum crop development and yield. If the soil is too loose (0 t) root contact with the soil particles may be hampered; if it is too com-pact (8.5 t and 14.5 t), root penetration into the lower soil layers is inhibited. Consequently, exploitation of the soil profile was suboptimal resulting in reduced crop yields The shift in the faunistic composition of the nematodes from microbivorous species associated with decomposition/mineralization processes to her-bivorous species associated with crop damage may also explain negative effects on crop yield. Densities of herbivorous Paratylenchus sp. of 30–90 specimens per gram dry soil could be harmful to the grass, as for example, carrots already experience damage at densi-ties of eight specimens per cm3soil (Weischer, 1959). The total numbers of nematodes not being affected by the degree of soil compaction, indicates that decreased habitable pore space for bacterivores was compensated for by the increased amount of feeding sites for herbivores.
Acknowledgements
The authors thank J. Bloem, P.C. de Ruiter and H.G. van Faassen for their help with the manuscript, G. Brouwer for the measurements of root distribution.
References
Arts, W.B.M., Verwijs, B.R., Van Maanen, J., 1994. The effect of traffic on the physical condition of a sandy soil and subsequent consequences for grass production. Rapp. 94-5, ISBN 90-5406-075-1, IMAG-DLO, Wageningen, The Netherlands.
Baird, S.M., Bernard, E.C., 1984. Nematode population and community dynamics in soybean–wheat cropping and tillage regimes. J. Nematol. 16, 379–386.
Boag, B., 1985. Effect of soil compaction on migratory plant-parasitic nematodes. Crop Res. (Hort. Res.) 25, 63–67. Böhm, W., 1979. Needleboard methods. In: Methods of Studying
Root Systems. Ecological Studies 33. Springer, Berlin, pp. 30–38, ISBN 3-540-09329x.
Boström, S., Sohlenius, B., 1986. Short-term dynamics of nematode communities in arable soil: influence of a perennial and an annual cropping system. Pedobiologia 29, 345–357. Bouwman, L.A., Hoenderboom, G.H.J., Van Klinken, A.C., De
Ruiter, P.C., 1993. Effect of growing crops and crop residues in arable fields on nematode production. In: Eysakkers, H.J.P., Hamers, T. (Eds.), Integrated Soil and Sediment Research: A Basis for Proper Protection. Kluwer, Dordrecht, The Netherlands, pp. 127–131.
Bouwman, L.A., Zwart, K.B., 1994. The ecology of bacterivorous protozoans and nematodes in arable soil. Agric. Ecosyst. Environ. 51, 145–160.
Brussaard, L., Van Faassen, H.G., 1994. Effects of compaction on soil biota and soil biological processes. In: Soane, B.D., van Ouwerkerk, C. (Eds.), Soil Compaction in Crop Production. Elsevier, Amsterdam, pp. 215–235.
Clarholm, M., Popovic, B., Rosswall, T., Söderström, B., Sohlenius, B., Staaf, H., Wiren, A., 1981. Biological aspects of nitrogen mineralization in humus from a pine forest podsol incubated under different moisture and temperature conditions. Oikos 37, 137–145.
De Goede, R.G.M., Bongers, T., 1994. Nematode community structure in relation to soil and vegetation characteristics. Appl. Soil Ecol. 1, 29–44.
Dexter, A.R., 1986. Model experiments on the behaviour of roots at the interface between a tilled seed-bed and a compacted sub-soil. I. Effects of seed-bed aggregate size and sub-soil strength on wheat roots. Plant Soil 95, 123–133.
Dmowska, E., 1993. Effects of long-term artificial rain on species range and diversity of soil nematodes. Eur. J. Soil Biol. 29, 97–107.
Edwards, L.M., 1988. The effects of slope position and cropping sequence on soil physical properties in Prince Edward Island. Can. J. Soil Sci. 68, 763–774.
Edwards, L.M., Kimpinski, J., 1997. Relationships between soil penetration resistance and soil nematode burden in barley on Prince Edward Island. Biol. Fertil. Soils 24, 13–17.
Elliott, E.T., Anderson, R.V., Coleman, D.C., Cole, C.V., 1980. Habitable pore space and microbial interactions. Oikos 35, 327– 335.
Fortnum, B.A., Karlen, D.L., 1985. Effect of tillage system and irrigation on population densities of plant nematodes in field corn. J. Nematol. 17, 25–28.
Gregory, P.J., 1988. Growth and functioning of plant roots. In: Wild, A. (Ed.), Russell’s Soil Condition and Plant Growth. Longman, UK, pp. 113–167.
Groenevelt, P.H., Kay, B.D., Grand, C.D., 1984. Physical assessment of a soil with respect to rooting potential. Geoderma 34, 101–114.
Hassink, J., Bouwman, L.A., Zwart, K.B., Brussaard, L., 1993. Relationships between habitable pore space, soil biota and mineralization rates in grassland soils. Soil Biol. Biochem. 25, 47–55.
Jacob, J.J., Van Bezooyen, J., 1984. Practical work in nematology. Agricultural University, Wageningen, The Netherlands. Jones, F.G.W., Thomasson, A.J., 1976. Bulk density as an indicator
of pore space in soils usable by nematodes. Nematologica 22, 133–137.
Juma, N.G., Mishra, C.C., 1988. Effect of an annual and a perennial crop on trophic group dynamics of nematodes. Can. J. Soil Sci. 68, 101–109.
Keen, A., 1993. Berijding grasland op zandgrond: statistische analyse jaaropbrengsten drogestof 1988 t/m 1992. IMAG-DLO, Wageningen, rapport 93–20, 6 pp.
Parmelee, R.W., Aston, D.G., 1986. Nematode trophic structure in conventional and no-tillage agroecosystems. J. Nematol. 18, 403–407.
Soane, B.D., Van Ouwerkerk, C., 1994. Soil compaction problems in world agriculture. In: Soane, B.D., van Ouwerkerk, C. (Eds.),
Soil Compaction in Crop Production. Elsevier, Amsterdam, pp. 1–21.
Sohlenius, B., Sandor, A., 1989. Ploughing of a perennial grass ley-effect on the nematode fauna. Pedobiologia 33, 199–210. Thomas, S.H., 1978. Population densities of nematodes under
seven tillage regimes. J. Nematol. 10, 24–27.
Van Noordwijk, M., De Willigen, P., 1984. Mathematical models on diffusion of oxygen to and within plant roots, with special emphasis on effects of soil-root contact. II. Applications. Plant Soil 77, 233–241.
Van Ouwerkerk, C., Soane, B.D., 1994. Conclusions and recommendations for further research on soil compaction in crop production. In: Soane, B.D., van Ouwerkerk, C.(Eds.), Soil Compaction in Crop Production. Elsevier, Amsterdam, pp. 627–642.
Wasilewska, L., 1979. The structure and function of soil nematode communities in natural ecosystems and agrocenoses. Pol. Ecol. Stud. 5, 97–115.
Weischer, B., 1959. Eine durch Nematoden verursachte Möhrenmüdigkeit. Verh. IV. Int. Pflanzenschutz Kongr. Hamburg, Braunschweig, Vol. 1, 1957, pp. 583–585. Yeates, G.W., 1979. Soil nematodes in terrestrial ecosystems. J.