TINJAUAN PUSTAKA
Sponge
Sponge merupakan organisme multiselluler yang dikelompokkan dalam Filum porifera (Kozloff 1990; Brusca dan Brusca 1990). Organisme ini tergolong hewan yang telah hidup jutaan tahun yang lalu, karena kemampuannya untuk bertahan dalam berbagai kondisi ekstrim. Beberapa spesies sponge telah hidup dari zaman Precambrian sekitar 600 juta tahun yang lalu (University of Stutgart Sponge Reef Project 2003). Hewan ini tidak bergerak, tetapi tinggal sepanjang hidupnya seperti tanaman, dan mampu menyaring air untuk memperoleh partikel makanan dari air dan substansi terlarut lainnya.
Diperkirakan terdapat lebih dari 7.000 spesies sponge yang tersebar di laut ataupun di perairan tawar, tetapi baru sebagian yang telah teridentifikasi dan diberi nama spesies (University of Stutgart Sponge Reef Project 2003). Secara garis besar terdapat 4 kelas dalam filum porifera ini, yaitu Calcarea, Hexactinellida, Demospongiae dan Sclerospongiae (Kozloff 1990; Brusca dan Brusca 1990). Keempat kelas ini dibedakan berdasarkan bentuk dari skeleton internal (spikula) dan masing- masing kelas mempunyai ciri-ciri tersendiri. Kelas Calcarea mempunyai spikula yang terdiri dari kalsium karbonat, bentuk spongenya relatif kecil dan sederhana, contoh kelas ini adalah Sycon dan Grantia. Kelas Sclerospongiae mempunyai spikula yang mengandung kalsium karbonat, calcite atau aragonite, mempunyai tekstur yang kuat dan umumnya hidup pada palung yang dalam. Kelas Hexactinellida atau dikenal dengan glass sponge mempunyai spikula yang terdiri dari silika denga n bentuk silindris yang simetrik dengan 6 sudut, hidup pada kedalaman lebih dari 50 m (Brusca dan Brusca 1990). Kelas Demospongiae merupakan kelas yang terbesar, meliputi 95% dari semua spesies sponge, spikula nya terdiri dari silika, umumnya tidak mempunyai bentuk yang teratur atau asimetrikal, termasuk dalam kelas ini adalah bath sponges, fresh water sponges dan boring sponges. Umumnya sponges yang banyak diteliti untuk diambil proteinnya atau kandungan metabolit sekunder adalah yang termasuk dalam kelas ini seperti Tethya, Axinella dan Suberitus.
Struktur Sponge
Sponge tidak mempunyai struktur tubuh yang jelas, karena tidak mempunyai batas jaringan yang nyata (Brusca dan Brusca 1990; Hawking dan Smith 1997). Sponge ada yang berbentuk sederhana seperti tabung dengan dinding tipis seperti yang dijumpai pada marga Leucosolenia, segumpal jaringan yang tidak menentu bentuknya, membuat kerak pada batu, cangkang, tonggak atau tumbuh-tumbuhan. Kelompok sponge lain mempunyai bentuk yang lebih teratur dan melekat pada dasar perairan melalui sekumpulan spikula. Jenis-jenis tertentu nampak berbulu getar karena spikulanya menyembul keluar dari badan.
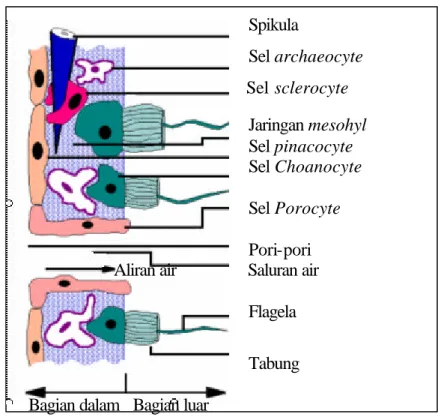
Selain bentuknya beragam, sponge juga mempunyai warna dan ukuran yang beragam. Warna sponge ada yang putih, abu-abu, kuning, orange, merah atau hijau. Sponge yang berwarna hijau umumnya disebabkan oleh adanya alga yang bersimbiotik (zoochlorellae) di dalam sponge. Sponge berukuran mulai dari sebesar kepala jarum pentul sampai berukuran diameter 0,9 m dan tebal 30,5 cm. Bagian tubuh sponge terdiri dari sistem saluran (canal), sistem kerangka dan sel somatik (Gambar 1). Sistem saluran ini bertindak seperti halnya sistem sirkulasi pada hewan tingkat tinggi. Sistem ini merupakan jalan untuk pemasukan makanan ke dalam tubuh dan untuk saluran pembuangan. Ada tiga macam sistem, yaitu askon, sikon dan ragon.
Semua sponge, kecuali yang termasuk ordo kecil Myxospongia, dilengkapi dengan kerangka. Kerangka ini ada yang terdiri dari kapur karbonat dan silika dalam bentuk spikula atau dari spongin dalam bentuk serat yang kurang lebih erat bersatu. Sponge tidak dapat berdiri tegak jika tidak ada spikula atau spongin yang menopang tubuhnya dan membuat saluran dan ruang-ruang bercambuk. Spikula tertimbun dalam sel-sel yang disebut scleroblast. Spikula silikon tersusun dari opal, suatu bentuk silika terhidrasi seperti kuarsa dalam struktur kimianya. Kapur karbonat dan silikon sebagai bahan dasar spikula diekstrak oleh sel-sel dari air di sekitarnya. Spongin adalah senyawa kimia yang menyerupai struktur sutera, yang dihasilkan oleh sel berbentuk toples yang disebut spongoblast.
Spikula Sel archaeocyte Sel sclerocyte Jaringan mesohyl Sel pinacocyte Sel Choanocyte Sel Porocyte Pori-pori Aliran air Saluran air
Flagela
Tabung
Bagian dalam Bagian luar
Sumber : http://www.ucmp.berkeley.edu/porifera/pororg.html
Gambar 1 Struktur Tubuh Sponge.
Sel- sel sponge mempunyai fungsi-fungsi khusus dan pembagian kerja yang jelas (Brusca dan Brusca 1990; Hawking dan Smith 1997). Sel-sel ini dipisahkan menjadi tiga kelompok, yaitu 1). Sel yang menyusun lapisan kulit, 2). Sel yang membentuk organ skeleton, dan 3) sel yang terletak di dalam jaringan mesohyl (sel amoeboid), yang berdiferensiasi dan mempunyai fungsi tertentu.
Sel yang termasuk kelompok pertama terdiri dari sel pinacocyte, porocyte, dan choanocyte. Sel pinacocyte merupakan sel penyusun lapisan permukaan sponge (seperti sel epitel pada hewan) yang membentuk pinacoderm. Sel ini berbentuk datar dan saling overlap, yang diselingi juga sel pinacocyte berbentuk T. Porocytes merupakan sel yang berbentuk silinder seperti tabung, dapat membuka dan menutup porinya untuk mengatur diameter ostial. Choanocytes merupakan sel yang berflagel yang berbaris membentuk lapisan choanoderm. Sel ini mempunyai colar (tabung) yang menjadi tempat masuknya air yang membawa makanan. Makanan disimpan dalam vakuola makanan dan dikirim ke sel di sampingnya (sel ameboid) tempat mencerna makanan (Brusca dan Brusca 1990) .
Sel yang membentuk organ skeleton terdiri dari sel sclerocytes, collencytes, lophocytes dan spongocytes. Sel sclerocytes bertanggung jawab
memproduksi spikula silika atau spikula karbonat (Brusca dan Brusca 1990). Pada sel ini juga terbentuk protein aksial yang kemudian dikenal dengan silicatein (Shimizu et al. 1998). Sel tersebut, merupakan sel yang aktif, mempunyai sejumlah besar mitokondria, retikulum endoplasma kasar dan vakuola. Diketahui kemudian bahwa vakuola dalam sel ini merupakan tempat untuk mendepositkan asam silika yang diambil dari perairan. Sel collencytes dan lophocytes bentuknya hampir sama dengan pinacocytes, berfungsi untuk menghasilkan kolagen, sedangkan sel spongocytes memproduksi serat seperti kolagen yang disebut spongin.
Sel yang terletak di jaringan mesohyl adalah myocytes, archaeocytes dan rhabdiferous. Sel myocytes merupakan sel kontraktil. Sel archaeocytes berukuran besar, sangat motil dan mempunyai peranan penting dalam mencerna dan mentranspor makanan. Sel ini mempunyai sejumlah enzim pencernaan (seperti acid phospatase, amilase, protease, lipase) dan menerima bahan phagocyt dari choanocytes. Sel rhabdiferous merupakan sel dalam mesohyl yang berukuran paling besar mengandung mucopolisakarida (Brusca dan Brusca 1990).
Struktur Biosilika
Sponge dan diatom serta beberapa organisme mensintesis giga ton silika pertahun dari asam silikat. Struktur biosilika yang dibuat mempunyai beragam struktur dengan ketepatan pengontrolan nanoarsitektur yang melebihi kemampuan manusia. Dalam sponge, biosilika ini terkumpul pada bagian yang bernama spikula.
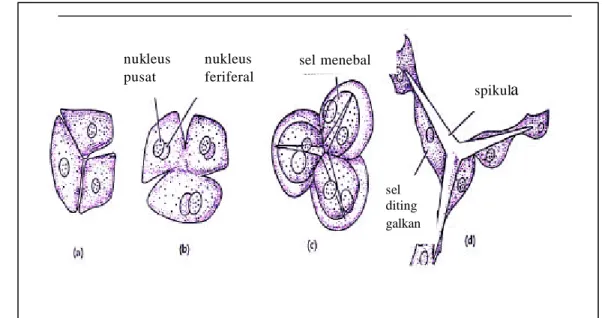
Spikula merupakan organ skeleton dari sel sponge yang menopang struktur jaringan dan tubuh sponge. Pada sponge, spikula ini tersusun atas kalsium karbonat (calcareous spicule) atau silika (silica spicule) dan sebagian kecil kolagen. Pada beberapa sponge, skeleton hanya terdiri dari kolagen saja. Spikula dibentuk oleh sel yang terletak di bagian mesohyl yang disebut sclerocytes (Gambar 1). Sel yang terdiri dari mitokondria, mikrofilamen sitoplasma dan vakuola kecil ini dapat menyimpan kalsium karbonat atau silika dan menyusunnya menjadi spikula atau silika spikula. Satu sel sclerocytes dapat
menghasilkan 1 silika spikula atau beberapa sel bekerja sama untuk menghasilkan 1 silika spikula (Gambar 2).
Sumber : Kozloff (1990)
Gambar 2 Pembentukan Spikula oleh Sel Sclerocytes.
Berdasarkan ukurannya spikula dapat dibedakan menjadi dua jenis, yaitu microscleres dan megacleres (Gambar 3). Pada sponge kelas Hexactinellida umumnya kedua tipe ini ada, dengan bentuk hexactinal. Pada sponge kelas Demospongiae terdapat spikula tipe megacleres dan terkadang bersamaan dengan tipe microscleres (Brusca dan Brusca 1990).
Sumber : Kozloff (1990) Sumber: Black (2003)
Gambar 3 Tipe Spikula. a. Megascleres dan b. Microscleres c. Bentuk Microscleres, Starburst pada T.aurantia.
c sel menebal spikula nukleus pusat nukleus feriferal sel diting galkan
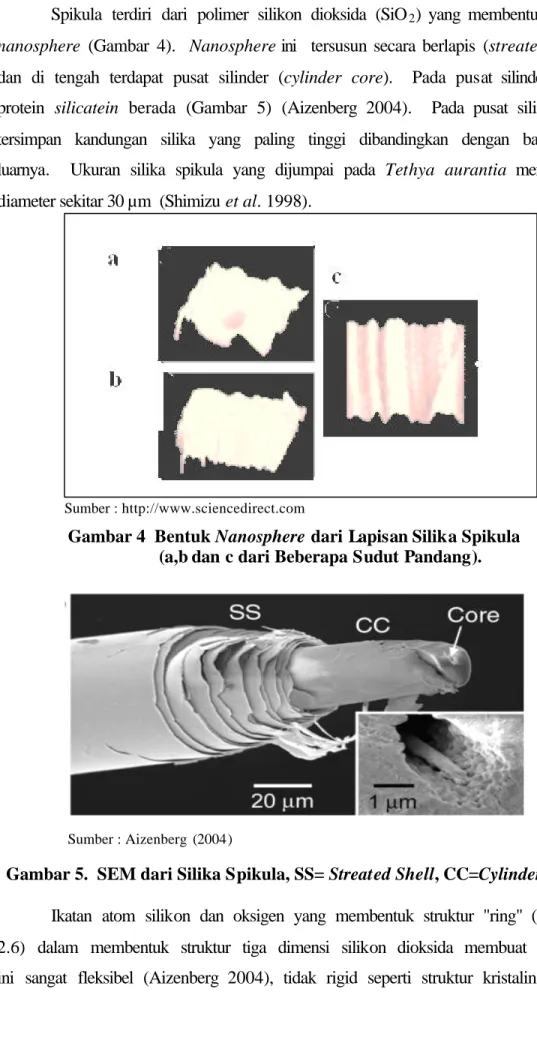
Spikula terdiri dari polimer silikon dioksida (SiO2) yang membentuk silika nanosphere (Gambar 4). Nanosphere ini tersusun secara berlapis (streated shell) dan di tengah terdapat pusat silinder (cylinder core). Pada pusat silinder inilah protein silicatein berada (Gambar 5) (Aizenberg 2004). Pada pusat silinder ini tersimpan kandungan silika yang paling tinggi dibandingkan dengan bagian di luarnya. Ukuran silika spikula yang dijumpai pada Tethya aurantia mempunyai diameter sekitar 30 µm (Shimizu et al. 1998).
Sumber : http://www.sciencedirect.com
Gambar 4 Bentuk Nanosphere dari Lapisan Silika Spikula (a,b dan c dari Beberapa Sudut Pandang).
Sumber : Aizenberg (2004)
Gambar 5. SEM dari Silika Spikula, SS= Streated Shell, CC=Cylinder Core
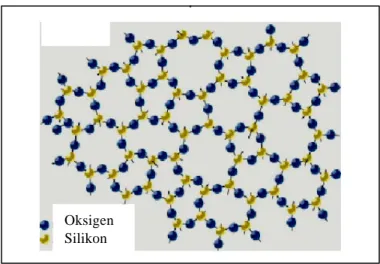
Ikatan atom silikon dan oksigen yang membentuk struktur "ring" (Gambar 2.6) dalam membentuk struktur tiga dimensi silikon dioksida membuat molekul ini sangat fleksibel (Aizenberg 2004), tidak rigid seperti struktur kristalin (ikatan
Si-Si). Struktur seperti ini dikenal dengan amorphous, dan struktur ini membuat beberapa ion anorganik penting, seperti Na+ dapat melewati lapisan silika.
.
Sumber: http://www.batnet.com/enigmatics/semiconductor_ processing/CVD_Fundamentals/films/SiO2_properties.html
Gambar 6 Jaringan Tiga Dimensi dari Silikon Dioksida.
Pembentukan biosilika dalam sponge dan diatom sangat dipengaruhi oleh keberadaan asam silikat, garam natrium dan adanya protein transporter. Krasko (2000) mempelajari ekspresi gen silicatein dengan meningkatkan konsentrasi asam silikat dari 1 µM menjadi sekitar 60 µM, dan ternyata menghasilkan ekspresi
gen yang sangat meningkat. Tidak adanya penambahan asam silikat dalam media, menyebabkan ekspresi gen yang sangat kecil. Begitupun halnya dengan ekspresi gen kolagen, gen yang sering dihubungkan dengan silicatein, meningkat dengan penambahan asam silikat tersebut.
Konsentrasi asam silikat di lautan berada pada level mikromolar per ml, sedangkan yang terdeposit dalam vakuola sponge terdapat 1000 kali lebih besar. Banyaknya asam silikat yang terdeposit ini diduga karena adanya protein transporter yang mengangkut asam silikat dari lingkungan ke dalam vakuola sel. Schroder et al. pada tahun 2004 mempelajari protein transporter yang mengangkut asam silikat ke dalam sponge Suberites domuncula; protein ini mirip dengan Na+ /HCO3– cotransporters. Coradin dan Lopez (2003) juga menemukan hal yang sama bahwa silika transporter mirip dengan Na+ /HCO3– cotransporters, karena sangat tergantung pada keberadaan natrium, dengan perbandingan antara silikat yang diangkut dengan natrium yang diperlukan (Si(OH)4 : Na+) sebesar 1:1.
Oksigen Silikon
Fabrikasi Silika
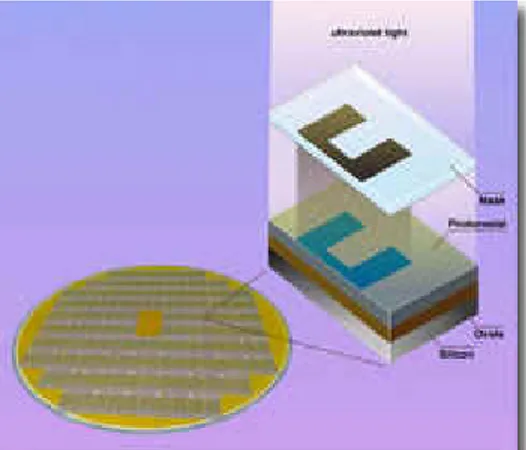
Silika merupakan bahan dasar untuk industri- industri besar. Produk intermediet silika yang umum digunakan adalah silica chips, fiber glass dan bahan keramik. Silica chips diaplikasikan untuk sparepart komputer, assay kit, dan lain-lain. Silica chips (Gambar 7) dibuat melalui beberapa langkah dan diawali dengan pembuatan lapisan- lapisan polisilikon (disebut wafer). Pembentukan wafer ini terdiri dari pembuatan polisilikon dari reaksi triklorosilane dengan hidrogen pada suhu 1000oC dan dikristalkan pada suhu 1200oC. Polikristalsilikon merupakan template untuk pembentukan lapisan-lapisan tipis silikon dioksida yang mampunyai ketebalan beberapa mikron. Melalui proses pemberian muatan dan pengemasan, wafer ini menjadi chips (Richardson 2001).
Sumber: Richardson (2001)
Gambar 7 Silica Chips.
Fiber glass merupakan bahan utama pembuatan peralatan medis, seperti statesteskop, alat pendeteksi, mikroskop serta peralatan elektronik (sebagai pelapis penghantar). Fiber glass dibuat dengan cara oksidasi termal dari silicon tetrachloride (SiCl4), pada suhu yang sangat tinggi (sekitar 2100°C), dengan formula: SiCl4 + O2 SiO2 + 2Cl2 (Fiber Tech 2004) .
Dalam sistem alami, pembentukan kristal lapisan tipis terjadi melalui perangkaian atom-atom yang diawali dengan penempelan ion pada permukaan bahan organik maupun anorganik sebagai template (Banfield 2000). Pembentukan lapisan tipis silika, seperti pada chips dan keramik, juga
memerlukan adanya template. Adamson (2004) menggambarkan proses kondensasi silika menjadi lapisan film pada industri keramik memerlukan adanya template (Gambar 8).
Sumber : Adamson (2004)
Gambar 8 Proses Kondensasi Slika dengan Menggunakan Template.
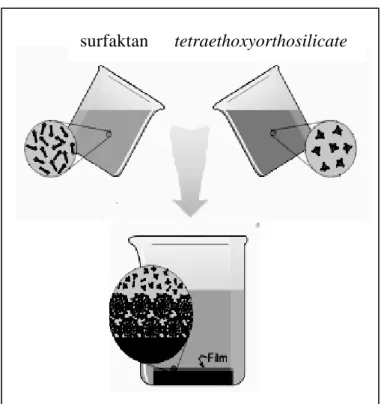
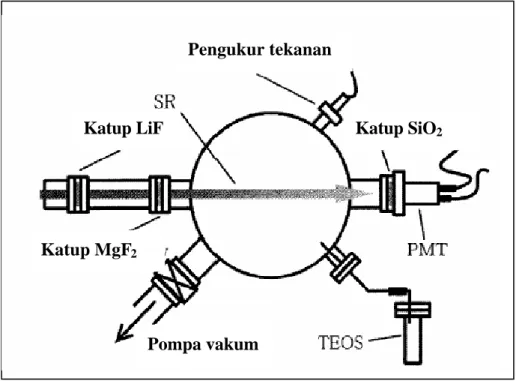
Dari uraian di atas, terdapat dua hal penting yang diperlukan dalam pembuatan polimer silika yaitu suhu yang sangat tinggi dan template. Para ilmuwan mulai memikirkan untuk mengganti cara pembuatan silika dengan cara yang lebih aman yaitu pada suhu rendah. Takezoe et al. (1999) mencoba melakukan pembuatan silika pada suhu kamar dengan menggunakan radiasi ultraviolet di bawah kondisi vakum dalam chamber yang terkontrol (Gambar 9). Proses ini menggunakan substrat murni tetraethoxyorthosilicate (TEOS) dengan katalis gas LiF dan MgF2.
Sumber: Takezoe et al (1999)
Gambar 9 Pembuatan Silika dengan Menggunakan Radiasi Ultraviolet.
Penggunaan katalis protein yang dapat melakukan kondensasi dan polimerisasi silika pada suhu rendah merupakan alternatif yang lebih aman. Shimizu et al. (1999) telah menunjukkan bahwa silicatein secara invitro dapat mengkatalisis hidrolisis tetraethoxyorthosilicate (TEOS) membentuk polimer silika pada pH netral, tekanan normal dan suhu kamar. Keistimewaan protein silicatein ini, selain berfungsi sebagai katalis juga dapat berfungsi sebagai template untuk penempelan silika.
Protein Silicatein
Tahun 1998, ditemukan protein yang menjadi katalis reaksi pembentukan biosilika dari sponge Tethya aurantia yang kemudian dikenal dengan silicatein oleh Shimizu et al. Protein silicatein diproduksi di dalam sel sclerocytes kemudian disekresikan ke membran vakuola tempat terdepositnya asam silikat. Di dalam vakuola protein terus memanjang dan dilapisi dengan silika yang terkondensasi dan terpolimerisasi olehnya, kemudian setelah membentuk spikula
Pompa vakum Katup MgF2
Katup LiF Katup SiO2
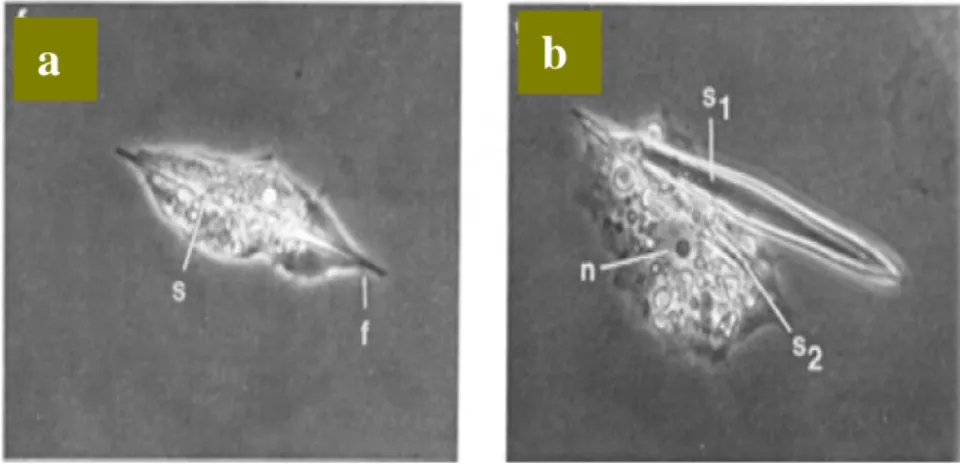
dengan panjang yang cukup protein dan spikula ini disekresikan ke luar sel. Sekresi protein ini dibuktikan dari foto mikroskop electron pada Gambar 10 (Aizenberg 2004). Pada Gambar 10a terlihat proses pemanjangan sel seiring dengan tumbuhnya protein dan silika spikula. Pada Gambar 10b, spikula pertama (s1) telah dikeluarkan dan sel siap membuat protein dan spikula berikutnya (s2). Dalam sel ini banyak ditemukan mitokondria yang mengindikasikan bahwa pembentukan protein ini memerlukan banyak energi.
Sumber : Aizenberg (2004)
Gambar 10 SEM dari Proses Sekresi Silika Spikula dengan Protein a. Protein dan Spikula Memanjang
b. Protein dan Spikula Keluar dari Sel.
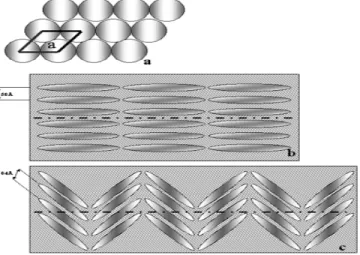
Susunan filamen protein di dalam silika spikula menjadi pertanyaan para peneliti. Croce (2004) meneliti hal ini dari spikula dua kelas sponge berbeda, yaitu sponge G. cydonium dan spikula dari Suberitus joubini dengan menggunakan X-ray fiber diffraction dengan SAXS beamline dari radiasi ELETTRA synchrotron. Digambarkan dalam dua dimensi, filamen protein ini menyusun hexagonal yang sangat teratur (Gambar 11a). Susunan filamen dari spikula G . cydonium, kelas Demospongiae (Gambar 11b) berbeda dengan model susunan filamen dari spikula S. joubini (Gambar 11c) dari kelas Hexactinellida.
Shimizu et al. (1998) yang mengisolasi protein ini, menemukan tiga pita protein dengan berat molekul 29, 28 dan 27 kDa yang kemudian menyebutnya sebagai 3 subunit α,β dan γ. Peneliti lain menyebutnya sebagai 3 protein isomer, karena satu subunit saja mampu melakukan reaksi katalisis sendiri terlepas dari sub unit lainnya. Ketiga protein ini mempunyai susunan asam amino yang hampir
sama. Ketiganya tersusun secara berulang membentuk protein filamen, yang diduga melalui ikatan nonionik dan nonkovalen, karena ikatan antar subunit mudah putus oleh penambahan SDS atau urea. Hasil analisis densitomer memperlihatkan bahwa silicatein α merupakan bagian terbesar yaitu sekitar 70% dengan perbandingan silicatein α,β dan γ =12:6:1 (Shimizu et al. 1998).
Sumber : Croce (2004)
Gambar 11 Susunan Filamen Protein dalam Silika Spikula a. Susunan Protein Secara Hexagonal
b. Susunan Protein pada Spikula G. cydonium c. Susunan Protein pada Spikula S. joubini.
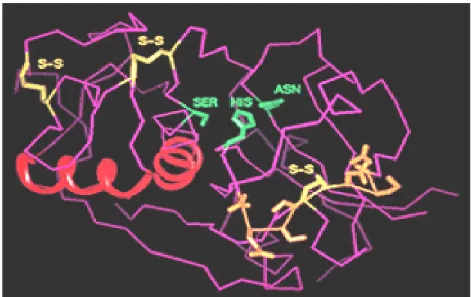
Shimizu et al. (1998) lebih lanjut mengemukakan bahwa sekuens dari silicatein α ini mempunyai similaritas yang tinggi dengan famili cathepsin-L dari grup cystein protease. Persamaan ini pertama terletak pada residu asam amino pada sisi aktif yaitu His dan Asn. Persamaan kedua, kedua protein ini mengandung 6 sistein yang membentuk jembatan disulfida; dan persamaan ketiga, pada kedua protein terdapat mekanisme pemotongan proprotein menjadi protein matang,. Berdasarkan kesamaan struktur ini diduga model tiga dimensi dari protein ini seperti pada Gambar 12.
Krasko et al. (2000) berhasil mengisolasi silicatein dari sponge Suberites domuncula dan menemukan bahwa ekspresi gen penyandi silicatein dikontrol secara positif oleh kehadiran substrat asam silikat. Protein ini mempunyai 79% similarity dengan silicatein dari T. aurantia dan tersusun dari 331 asam amino dengan berat molekul proprotein 36306 dalton dan 23125 dalton untuk protein
matang. Kecepatan sponge dalam membentuk spikula dari silika sangat tinggi, sekitar 5 µm per jam.
Sumber: Shimizu et al. (1998)
Gambar 12 Model Tiga Dimensi Silicatein.
Mekanisme pembentukan biosilika oleh protein ini belum banyak dimengerti. Diduga banyaknya asam amino hidroksil (serin, tirosin, dan treonin) pada silicatein berpengaruh besar pada proses biosilification. Hal ini didasarkan pada penelitian Perry dan Lu (1992) yang meneliti pembentukan silikon catekolat pada tanaman, yang melibatkan protein yang banyak mengandung gugus hidroksil. Begitu pula protein yang diduga pembentuk silika pada diatom mengandung sejumlah besar asam amino hidroksil (Kroger 1997).
Penelitian selanjutnya, pada tahun 1999 oleh Cha et al., menunjukkan bahwa protein ini secara invitro mampu mengkatalisis reaksi kondensasi dan polimerisasi silika dari substrat tetraethoxyorthosilicate dan siloxane walaupun belum diketahui mekanisme reaksinya secara pasti. Mekanisme reaksi katalisis dari protein ini diduga berdasarkan mekanisme reaksi pada serine protease atau cysteine protease.
Pembentukan biosilika pada sponge bersifat spesifik untuk setiap spesies diduga melibatkan protein sebagai katalis yang berbeda pula. Demikian pula
lingkungan yang berbeda diduga berpengaruh terhadap karakteristik proses katalisis dari protein tersebut.
Isolasi Protein dan Analisis Berat Molekul
Metode yang digunakan untuk isolasi protein tergantung dari sumber protein dan lokasi protein dalam sumber tersebut. Sumber protein dapat berasal dari hewan, tanaman atau bakteri. Sel hewan umumnya lebih mudah dipecah dibandingkan dengan sel tanaman, karena sel hewan tidak berdinding sel, sedangkan sel tanaman berdinding sel selulosa (Brummer dan Gunzer 1987). Lokasi protein mempengaruhi teknik pemisahan protein dari komponen lainnya. Ekstraksi dan isolasi protein ekstraseluler lebih mudah dibandingkan dengan protein intraseluler. Untuk isolasi protein ekstraseluler umumnya hanya dilakukan pemisahan berdasarkan sifat fisik (Thenawijaya 1989).
Masalah utama dalam isolasi protein adalah terjadinya denaturasi, kontaminasi protein oleh pirogen dan asam nukleat serta adanya proteolisis (Errson et al. 1998). Hal tersebut dapat diatasi dengan pemilihan larutan ekstraksi dan larutan penyangga yang tepat, waktu preparasi yang singkat dan suhu rendah. Faktor-faktor yang diperlukan dalam pemilihan larutan ekstraksi adalah pH, jenis larutan penyangga (anionik atau kationik), variasi pH dengan kekuatan ion atau suhu, reaktivitas, pengaruhnya terhadap aktivitas biologis protein yang diinginkan, kelarutan, pengaruh deterjen atau senyawa khaotropik, logam pengikat dan penghambatan proteolitik (Errson et al. 1998).
Protein dikarakterisasi berdasarkan beberapa sifat biokimianya. Sifat biokimia yang umum digunakan untuk mengkarakterisasi protein adalah berat molekul dan reaksi katalisis terhadap substrat tertentu. Berat molekul protein dapat dianalisis dengan menggunakan teknik elektroforesis. Elektroforesis merupakan teknik pemisahan fraksi- fraksi zat berdasarkan migrasi partikel bermuatan atau ion- ion makromolekul di bawah pengaruh medan listrik karena adanya perbedaan ukuran, bent uk, muatan atau sifat kimia molekul (Pomeranz dan Meloan 1980). Teknik ini dapat digunakan untuk menentukan berat molekul, mendeteksi kemurnian dan kerusakan protein, menetapkan titik isoelektrik protein
serta memisahkan spesies-spesies molekular yang berbeda secara kuantitatif dan kualitatif (Boyer 1986).
Pada elektroforesis harus digunakan media yang dapat mengurangi atau mencegah terjadinya konveksi dan tidak bereaksi dengan sampel atau menghambat pergerakan partikel sebagai akibat terjadinya ikatan antara sampel dengan matriks. Menurut Dunn (1989) elektroforesis dengan media gel poliakrilamida yang dikenal dengan PolyAcrilamide Gel Electrophoresis (PAGE) merupakan teknik yang paling banyak digunakan untuk memisahkan protein karena mempunyai kapasitas pemisahan yang tinggi. Gel poliakrilamida terbentuk melalui polimerisasi monomer akrilamida dan pembentukan ikatan silang kovalen antar rantai panjang akrilamida baik melalui reaksi kimia maupun fotokimia. Pembentukan gel melalui reaksi kimia dengan ammonium persulfat sebagai inisiator dan N,N,N’,N’-tetrametilendiamin (TEMED) sebagai katalis. Cross-linking agent yang digunakan adalah N,N’-metilen-bis- akrilamida (bis) (Perbal 1988)
Pembentukan ikatan silang antar rantai panjang akrilamida membuat struktur seperti pori-pori pada gel. Besar kecilnya pori-pori ini menentukan kecepatan pemisahan protein dalam gel. Ukuran pori-pori dapat diatur dengan mengubah persentase akrilamida. Persentasi akrilamida yang dibutuhkan untuk dapat memisahkan protein dengan berat molekul tertentu dapat dilihat pada Tabel 1.
Tabel 1 Hubungan Persentase Poliakrilamida dengan Berat Molekul Protein
% poliakrilamida Kisaran berat molekul
3-5 >100.000
5-12 20.000-150.000
15 10.000-80.000
>15 <150.000
Sumber :Dunn (1989)
SDS (Sodium Dedosyl Sulfate) merupakan detergen anionik bahan pendenaturasi protein bila campuran dipanaskan 100oC selama 3 menit. Penggunaan SDS bertujuan untuk memberikan muatan negatif pada protein yang akan dianalisa. Protein yang terdenaturasi sempurna akan mengikat SDS dalam
jumlah yang setara dengan berat molekul protein tersebut (Dunn 1989). Denaturasi protein dilakukan dengan merebus sampel dalam buffer yang mengandung β-merkaptoetanol (berfungsi untuk nereduksi ikatan disulfida), gliserol dan SDS. Muatan asli protein akan digantikan oleh muatan negatif dari anion yang terikat sehingga kompleks protein-SDS memiliki rasio muatan per berat molekul yang konstan. Kerapatan muatan listrik keseluruhan setelah terbentuk kompleks SDS-polipeptida menjadi sama sehingga bisa bergerak dalam gel (dengan porositas yang benar) menurut ukuran molekul (Hames 1987). Gel poliakrilamida akan memberi efek pemisahan berdasarkan perbedaan viskositas kompleks protein-SDS, yang juga meningkat secara eksponensial terhadap panjang rantai atau berat molekul protein (Mihalyi 1992).