1
AKTIVITAS PROMOTER KERATIN DAN HEAT SHOCK
PADA IKAN KOI Cyprinus carpio
DWI HANY YANTI
SKRIPSI
PROGRAM STUDI TEKNOLOGI DAN MANAJEMEN AKUAKULTUR FAKULTAS PERIKANAN DAN ILMU KELAUTAN
INSTITUT PERTANIAN BOGOR 2009
2
PERNYATAAN MENGENAI SKRIPSI DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa skripsi yang berjudul:
AKTIVITAS PROMOTER KERATIN DAN HEAT SHOCK PADA IKAN
KOI Cyprinus carpio
adalah benar merupakan hasil karya yang belum diajukan dalam bentuk apa pun kepada perguruan tinggi manapun. Semua sumber data dan informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Bogor, Januari 2009
DWI HANY YANTI C14104019
3
RINGKASAN
DWI HANY YANTI. Aktivitas Promoter Keratin dan Heat Shock pada Ikan Koi Cyprinus carpio. Dibimbing oleh KOMAR SUMANTADINATA dan
ALIMUDDIN.
Promoter merupakan bagian penting dari suatu konstruksi DNA, yang disambungkan dengan gen penyandi protein tertentu. Penelitian ini dilakukan untuk mengetahui aktivitas promoter keratin dan heat shock pada ikan koi Cyprinus carpio, yang disambungkan dengan gen Green Fluorescent Protein (GFP) sebagai penanda dalam bentuk konstruksi DNA keratin-GFP dan heat shock-GFP. Keaktifan promoter dapat dilihat dari ekspresi sementara (transient expression) gen penanda yang dihasilkan.
Perbanyakan konstruksi DNA dilakukan untuk mendapatkan konsentrasi larutan DNA sebanyak 50 ng/µl dalam 0,1 M KCl dan diinjeksikan ke dalam blastodisk telur ikan koi pada fase 1-2 sel. Jumlah telur yang diinjeksikan sebanyak 30 butir untuk setiap konstruksi DNA dan diulang sebanyak 3 kali. Telur diinkubasi pada akuarium dengan suhu air 28oC. Kemudian, dilakukan pengamatan ekspresi gen GFP menggunakan mikroskop fluoresen (Olympus SZX 16) setiap 2-6 jam setelah fertilisasi. Parameter yang diamati antara lain derajat kelangsungan hidup embrio (DKH-e), derajat penetasan (DP), persentase embrio yang mengekspresikan gen GFP (PEMG), persentase larva yang mengekspresikan gen GFP (PLMG), dan pola serta tingkat ekspresi gen GFP. DKH-e dihitung sebelum telur menetas (20 jam setelah fertilisasi), sedangkan DP dihitung ketika semua telur telah menetas. Pola ekspresi gen diamati pada jam ke-1 setelah fertilisasi setiap 2-6 jam hingga larva menetas. Tingkat ekspresi diamati pada fase embrio (jam ke-12 setelah fertilisasi).
Hasil penelitian memperlihatkan bahwa DKH-e (66,67±0,0%) dan DP (43,33±6,9%) kontrol lebih tinggi dibandingkan dengan perlakuan injeksi. DKH-e dan DP antara keratin-GFP dan heat shock-GFP (56,67±5,8% vs 55,56±9,6%; 36,67±8,8% vs 36,67±6,7%) relatif sama. Persentase embrio yang mengekspresikan gen heat shock-GFP (33,33±12,0%) lebih tinggi dibandingkan dengan keratin-GFP (15,56±8,4%). Larva yang mengekspresikan gen GFP terlihat hanya pada embrio yang diinjeksi gen heat shock-GFP (18,18±13,7%). Tingkat ekspresi gen heat shock-GFP lebih baik dibandingkan keratin-GFP dilihat dari ekspresi pendar hijau kurang terang (4,67±1,5 vs 2,00±1,0 butir), terang (3,00±1,0 vs 1,67±1,5 butir), dan sangat terang (2,33±1,5 vs 1,00±1,0 butir). Pola ekspresi gen keratin-GFP lebih cepat dari heat shock-GFP yang dilihat dari awal muncul ekspresi (jam ke-4 vs ke-6 setelah fertilisasi), puncak ekspresi (jam ke-12 vs jam ke-24 setelah fertilisasi), dan penurunan ekspresi keratin-GFP (jam ke-36 setelah fertilisasi) dan sudah tidak tampak lagi pada fase larva. Sedangkan ekspresi gen heat shock-GFP masih tetap ada hingga larva berumur 1 hari. Kesimpulannya adalah bahwa promoter heat shock ikan rainbow trout dapat mengendalikan ekspresi gen GFP lebih baik dibandingkan dengan promoter keratin ikan flounder Jepang pada ikan koi.
4
AKTIVITAS PROMOTER KERATIN DAN HEAT SHOCK
PADA IKAN KOI Cyprinus carpio
SKRIPSI
Sebagai salah satu syarat untuk memperoleh gelar Sarjana Perikanan pada Fakultas Perikanan dan Ilmu Kelautan
Institut Pertanian Bogor
Oleh: Dwi Hany Yanti
C14104019
PROGRAM STUDI TEKNOLOGI DAN MANAJEMEN AKUAKULTUR FAKULTAS PERIKANAN DAN ILMU KELAUTAN
INSTITUT PERTANIAN BOGOR 2009
5 Judul Skripsi : Aktivitas Promoter Keratin dan Heat Shock pada Ikan Koi
Cyprinus carpio Nama Mahasiswa : Dwi Hany Yanti
Nomor Pokok : C14104019
Disetujui,
Diketahui,
Dekan Fakultas Perikanan dan Ilmu Kelautan
Prof. Dr. Ir. Indra Jaya, M.Sc NIP. 131 578 799
Tanggal Lulus:
Pembimbing I Pembimbing II
Prof. Dr. Komar Sumantadinata NIP. 130 345 017
Dr. Alimuddin NIP. 132 133 953
6
KATA PENGANTAR
Puji dan syukur panjatkan kepada Allah SWT atas segala karunia-Nya sehingga skripsi ini dapat diselesaikan dengan baik. Judul yang dipilih dalam penelitian yang dilaksanakan pada bulan Agustus - September 2008 ini adalah Aktivitas Promoter Keratin dan Heat Shock pada Ikan Koi Cyprinus carpio.
Dalam menyelesaikan penelitian ini tidak semata didapatkan sendiri, melainkan dengan bantuan beberapa pihak. Untuk itu penulis mengucapkan terima kasih kepada:
1. Prof. Dr. Komar Sumantadinata selaku Pembimbing I dan Pembimbing Akademik yang telah membimbing dan mengarahkan penulis selama melakukan studi di IPB.
2. Dr. Alimuddin selaku Pembimbing II yang telah membimbing penulis selama melakukan penelitian hingga akhir penyusunan skripsi ini.
3. Dr. Kukuh Nirmala selaku dosen tamu yang telah memberikan kritik dan saran pada penyusunan skripsi ini.
4. Bapak Maskur, Bapak Adi Sucipto, Bapak Ayi Santika, dan Bapak Dian Hardiantho atas perijinan dan segala bantuan sehingga penelitian ini dapat dilaksanakan.
5. Ayahanda, Ibunda, kakak, dan adikku yang telah mencurahkan segala doa restu, kasih sayang, baik dukungan moril maupun materil.
6. Teman-teman BDP41, As-Syifa, Forsmile, FKM-C dan Ceria family atas kebersamaannya selama ini.
7. Lina Mulyani, Anna Octavera, Prihanik Marlina, Arief Eko Prasetiyo, Nafisah Ummatul Ukhroy, dan Ika Setiasih serta kepada semua orang yang tidak bisa disebutkan namanya satu per satu atas segala bantuan dan dukungan.
Semoga skripsi ini dapat bermanfaat bagi perikanan dan pengembangan teknologi transgenik di Indonesia.
Bogor, Januari 2009
7
RIWAYAT HIDUP
Penulis dilahirkan di Jakarta pada tanggal 9 Juli 1986 dari pasangan Bapak Sarjono dan Ibu Partilah. Penulis merupakan anak kedua dari tiga bersaudara.
Pendidikan formal yang dilalui Penulis adalah SDN 01 Ciangsana, Bogor tahun 1998; SLTPN 03 Gunung Putri, Bogor tahun 2001; dan SMUN 1 Cileungsi, Bogor tahun 2004. Pada tahun 2004 Penulis diterima di Program Studi Teknologi dan Manajemen Akuakultur, Departemen Budidaya Perairan, Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor melalui jalur Undangan Seleksi Masuk Institut Pertanian Bogor (USMI).
Selama mengikuti perkuliahan Penulis pernah mengikuti kegiatan magang di Sejahtera Lobster Farm, Desa Situ Daun, Bogor dan praktek lapang (PL) di Balai Riset Perikanan Budidaya Air Payau (BRPBAP) Maros, Sulawesi Selatan. Penulis juga pernah menjadi asisten mata kuliah Pendidikan Agama Islam pada 2006 - 2007 dan 2007 - 2008, asisten mata kuliah Nutrisi Ikan pada 2006-2007, dan asisten mata kuliah Dasar-Dasar Genetika Ikan pada 2007 - 2008. Selain itu, Penulis juga aktif menjadi staf Himpunan Mahasiswa Akuakultur (HIMAKUA) pada 2004 - 2005, staf Forum Keluarga Muslim FPIK pada 2004 – 2005 dan 2007 - 2008, staf Dewan Perwakilan Mahasiswa FPIK 2005-2006, dan staf Badan Eksekutif Mahasiswa FPIK pada 2006 - 2007.
Untuk menyelesaikan studi di Fakultas Perikanan dan Ilmu Kelautan, Penulis melakukan penelitian yang berjudul “Aktivitas Promoter Keratin dan
8
DAFTAR ISI
Halaman
DAFTAR TABEL ... x
DAFTAR GAMBAR ... xi
DAFTAR LAMPIRAN ... xii
I. PENDAHULUAN ... 1
1.1 Latar Belakang ... 1
1.2 Tujuan ... 3
II. TINJAUAN PUSTAKA ... 4
2.1 Perkembangan Telur Ikan Koi Cyprinus carpio ... 4
2.2 Promoter ... 4
2.2.1 Promoter Keratin ... 5
2.2.2 Promoter Heat Shock ... 6
2.3 Gen Green Fluorescent Protein (GFP) ... 7
2.4 Ekspresi Sementara (Transient Expression) ... 7
2.5 Mikroinjeksi ... 8
III. BAHAN DAN METODE ... 10
3.1 Waktu dan Tempat ... 10
3.2 Prosedur Kerja ... 10
3.2.1 Perbanyakan Konstruksi DNA ... 10
3.2.2 Pemeliharaan Induk, Pemijahan, dan Pengambilan Telur ... 10
3.2.3 Penghilangan Daya Rekat Telur ... 11
3.2.4 Pelaksanaan Mikroinjeksi ... 11
3.2.5 Penetasan Telur dan Pemeliharaan Larva ... 13
3.2.6 Pengamatan Ekspresi Gen Green Fluorescent Protein... 13
3.3 Pengamatan ... 14
3.3.1 Derajat Kelangsungan Hidup Embrio (DKH-e) ... 14
3.3.2 Derajat Penetasan (DP) ... 14
3.3.3 Persentase Embrio Mengekspresikan Gen GFP (PEMG) ... 14
3.3.4 Persentase Larva Mengekspresikan Gen GFP (PLMG) ... 14
9
3.4 Analisis Data ... 15
IV. HASIL DAN PEMBAHASAN ... 16
4.1 Hasil ... 16
4.1.1 Derajat Kelangsungan Hidup Embrio (DKH-e), Derajat Penetasan (DP), Persentase Embrio Mengekspresikan Gen GFP (PEMG), dan Persentase Larva Mengekspresikan Gen GFP (PLMG) ... 16
4.1.2 Tingkat dan Pola Ekspresi Gen Green Fluorescent Protein ... 17
4.2 Pembahasan ... 20 V. KESIMPULAN ... 24 5.1 Kesimpulan ... 24 5.2 Saran ... 24 DAFTAR PUSTAKA ... 25 LAMPIRAN ... 28
10
DAFTAR TABEL
Halaman
1. Derajat Kelangsungan Hidup Embrio, dan Derajat Penetasan Telur Ikan Koi Cyprinus carpio ... 16 2. Persentase Embrio dan Larva yang Mengekspresikan Gen Keratin-GFP dan
Heat Shock-GFP pada Ikan Koi Cyprinus carpio.. ... 17 3. Tingkat Ekspresi Gen Keratin-GFP dan Heat Shock-GFP pada Embrio Ikan
11
DAFTAR GAMBAR
Halaman
1. Sekuens Gen Keratin Ikan Flounder Jepang Paralichthys olivaceus (Yazawa
et al., 2005). ... 6
2. Cekungan Agarosa ... 11
3. Seperangkat Alat Mikroinjektor... 12
4. Injeksi pada Blastodisk Embrio Ikan Koi Fase Satu Sel ... 12
5. Seperangkat Alat Pengamatan Ekspresi Gen Green Fluorescent Protein ... 13
6. Kelompok Tingkat Ekspresi Gen Green Fluorescent Protein pada Embrio Ikan Koi Cyprinus carpio ... 17
7. Pola Ekspresi Gen Keratin-GFP dan Heat Shock-GFP pada Embrio Ikan Koi Cyprinus carpio ... 18
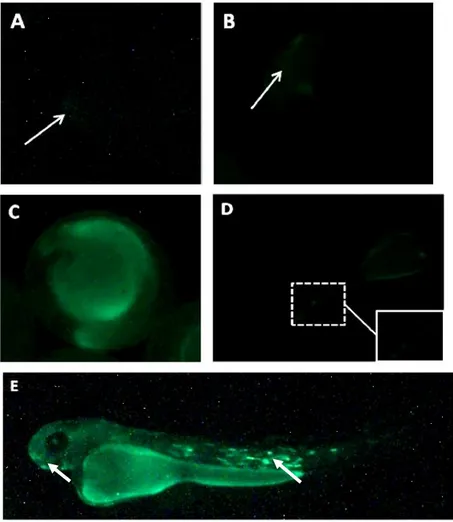
8. Ekspresi Gen Heat Shock-GFP pada Embrio Ikan Koi 6 Jam setelah Fertilisasi (A), 12 Jam setelah Fertilisasi (B), 24 Jam setelah Fertilisasi (C), pada Larva 30 Jam setelah Menetas di Sebagian Jaringan Tubuh (D) dan Hampir di Seluruh Tubuh (E) ... 19
9. Ekspresi Gen Keratin-GFP pada Embrio Ikan Koi 4 Jam setelah Fertilisasi (A), 12 Jam setelah Fertilisasi (B), 24 Jam setelah Fertilisasi (C) ... 19
12
DAFTAR LAMPIRAN
Halaman
1. Perkembangan Embriogenesis Telur Ikan Mas Cyprinus carpio (Woynarovich & Horvath, 1980) ... 28 2. Konstruksi Plasmid DNA Keratin-GFP (Yazawa et al., 2005) (A) dan Heat
Shock-GFP (Yamamoto dan Yoshizaki, belum dipublikasikan) (B) ... 30 3. Metode Kultur Cair Perbanyakan Bakteri dan Isolasi Plasmid DNA
Keratin-GFP dan Heat Shock-GFP ... 31 4. Proses Pemijahan Semi Alami Ikan Koi Cyprinus carpio ... 32
5. Derajat Kelangsungan Hidup Embrio, dan Derajat Penetasan Telur Ikan Koi Cyprinus carpio ... 33 6. Persentase Embrio yang Mengekspresikan Gen GFP, dan Persentase Larva
yang Mengekspresikan Gen GFP Ikan Koi Cyprinus carpio ... 34 7. Tingkat Ekspresi Gen Green Fluorescent Protein pada Embrio Ikan Koi
13
I. PENDAHULUAN
1.1Latar Belakang
Teknologi transgenesis menjadi sarana yang mulai diterapkan pada pengembangan akuakultur di Indonesia. Transgenesis dapat menghasilkan ikan transgenik yang memberikan beberapa manfaat antara lain peningkatan laju pertumbuhan (Nam et al., 2001; Kobayashi et al., 2007), peningkatan laju pemanfaatan pakan (Kobayashi et al., 2007), peningkatan kemampuan resisten ikan terhadap penyakit (Dunham et al., 2002; Sarmaşik, 2003), dan pembuatan ikan strain baru (Gong et al., 2003).
Transgenesis merupakan suatu proses mengintroduksikan DNA asing atau eksogenus (transgen) ke hewan uji dengan tujuan untuk memanipulasi struktur genetikanya (Glick dan Pasternak, 2003). Ada tiga tahap penting yang perlu dilakukan dalam menghasilkan ikan transgenik yaitu : 1) persiapan konstruksi DNA yang akan diintroduksikan, 2) pengintroduksian konstruksi DNA ke dalam embrio ikan melalui teknik transfer gen tertentu, dan 3) pengidentifikasian ikan yang membawa transgen (Hackett, 1993). Konstruksi DNA untuk transgenesis sebaiknya memiliki kemampuan dapat mengekspresikan suatu protein pada jaringan inang tertentu yang waktu dan tingkat ekspresinya dapat diatur sesuai dengan keinginan. Dalam hal ini, yang berperan penting dalam pengaturan ekspresi gen tersebut adalah elemen regulator (promoter).
Promoter merupakan sekuens DNA yang menginisiasi terjadinya proses transkripsi (Dunham, 2004), pengatur waktu, tempat, dan tingkat ekspresi suatu gen sehingga promoter dapat dianalogikan sebagai switch suatu gen (Glick dan Pasternak, 2003). Beberapa promoter telah berhasil diisolasi antara lain β-actin dari ikan medaka (Takagi et al., 1994), keratin8 dari ikan zebra (Gong et al., 2002),mylz2 (myosin light polypeptide 2), elaA (elastase A) dari ikan zebra (Gong et al., 2003), keratin dari ikan flounder Jepang (Yazawa et al., 2005), heat shock dari ikan rainbow trout (Kawamura dan Yoshizaki, tidak dipublikasikan), dan hsp27 dari ikan zebra (Wu et al., 2008). Promoter keratin dari ikan flounder Jepang merupakan promoter yang memiliki aktivitas tinggi pada jaringan epitel atau kulit yang merupakan pertahanan awal terhadap serangan penyakit (Yazawa
14 et al., 2005), sehingga dapat dimanfaatkan untuk mengekspresikan gen antiviral atau antibakteri pada ikan transgenik tahan penyakit. Promoter heat shock yang diisolasi dari ikan rainbow trout (Kawamura dan Yoshizaki, tidak dipublikasikan) diduga memiliki aktivitas yang sama dengan promoter hsp27 dari ikan zebra (Wu et al., 2008), yaitu memiliki aktivitas yang tinggi pada sel otot bahkan dapat aktif di seluruh jaringan jika dipicu dengan tekanan suhu (Wu et al., 2008), sehingga dapat dimanfaatkan salah satunya sebagai bioreaktor.
Pengujian aktivitas suatu promoter pun dilakukan setelah pengisolasian untuk mengetahui kemampuan suatu promoter dalam mengekspresikan gen penyandi protein tertentu. Promoter keratin ikan flounder Jepang dan heat shock ikan rainbow trout belum pernah diuji coba pada ikan koi. Ikan koi merupakan ikan hias air tawar yang memiliki nilai jual yang tinggi. Namun, terdapat hambatan dalam perkembangan usahanya yaitu serangan Koi Herpes Virus (KHV) dan belum ditemukan obat untuk mengatasinya. Berdasarkan hal ini diharapkan promoter keratin dari ikan flounder Jepang dan promoter heat shock dari ikan rainbow trout dapat dikembangkan untuk menghasilkan ikan koi transgenik tahan terhadap penyakit.
Untuk mengetahui aktivitas promoter, diperlukan adanya gen penanda (marker) yang disambungkan dengan promoter. Keaktifan promoter dapat dilihat dari ekspresi sementara (transient expression) yang dihasilkan gen penanda. Pola ekspresi yang dihasilkan pada embrio ikan menandakan keberadaan gen yang telah diatur oleh promoter (Yazawa et al., 2005). Berbagai macam gen penanda dapat digunakan untuk pengujian aktivitas promoter. Namun pada penelitian ini, gen penanda yang dipakai adalah gen Green Fluorescent Protein (GFP) dalam bentuk konstruksi DNA keratin-GFP dan heat shock-GFP. Gen GFP memiliki keunggulan yaitu tidak memerlukan substrat tambahan untuk ekspresinya, memiliki kandungan protein yang berpendar dan dapat divisualisasikan dengan menggunakan mikroskop fluoresen (Chalfie et al., 1994 dalam Iyengar et al., 1996). Gen GFP diisolasi dari ubur-ubur Aequorea victoria namun ada juga yang diisolasi dari anthozoa (soft coral) jenis Renilla reniformis yaitu gen hrGFP (Humanized Renilla reniformis Green Fluorescent Protein) (Felts et al., 2001).
15 Gen GFP juga berfungsi sebagai gen target seperti dalam pembuatan ikan hias berpendar yang berwarna-warni (Gong et al., 2003).
Umumnya pengujian aktivitas promoter dilakukan dengan metode mikroinjeksi yaitu menginjeksikan konstruksi DNA ke embrio dan mengamati ekspresi sementara yang dihasilkan gen penanda (Takagi et al., 1994; Higashijima et al., 1997; Hamada et al., 1998; Yazawa et al., 2005; Kato et al., 2007; Ath-thar, 2007; Purwanti, 2007). Pada penelitian ini konstruksi DNA keratin-GFP dan heat shock-GFP diinjeksikan ke blastodisk embrio ikan koi fase satu sampai dua sel dengan menggunakan mikroinjektor.
1.2Tujuan
Tujuan penelitian ini adalah untuk mengetahui aktivitas promoter keratin dan heat shock pada ikan koi dengan mengamati ekspresi sementara dari gen GFP (Green Fluorescent Protein) sebagai penanda (marker).
16
II. TINJAUAN PUSTAKA
2.1 Perkembangan Telur Ikan Koi Cyprinus carpio
Telur ikan koi yang telah dibuahi akan mengalami perubahan warna dan bentuk. Perkembangan telur ikan koi memiliki persamaan dengan perkembangan telur ikan mas yaitu dimulai dari proses pembelahan menjadi 2, 4, 8, 16, dan 32 sel (Lampiran 1). Pada fase pembelahan, telur akan terlihat seperti buah mulberry yang kemudian disebut sebagai fase morula (Woynarovich dan Horvath, 1980). Lapisan-lapisan sel yang disebut sebagai blastoderm akan mulai terbentuk, diawali dengan terbentuknya satu lapisan sel yang disebut blastomer. Fase morula merupakan fase yang sangat sensitif bagi perkembangan embrio. Pada fase morula, jumlah blastomer akan bertambah dan membuat ukuran sel semakin kecil. Jika terkena suatu gangguan, sel akan terguncang dan dapat mengakibatkan kematian pada embrio. Fase selanjutnya adalah fase blastula yaitu terbentuknya rongga yang membedakan antara kuning telur dengan sel. Sel mulai menutupi kuning telur secara bertahap dan hanya menyisakan satu lubang kecil yang disebut sebagai blastopor namun akhirnya blastopor pun tertutup sempurna. Pada fase blastula, terjadi transisi dari fase perkembangan awal germinal ke fase perkembangan embrio (organogenesis). Pada fase awal perkembangan organogenesis, massa sel akan membentuk cincin setengah lingkaran yang akan menjadi bakal kepala dan ekor di kedua ujungnya. Bintik mata mulai terlihat pada bagian kepala sebagai bakal mata dan bakal jantung mulai berkembang bahkan sudah berdetak. Pembuluh darah kemudian mulai terbentuk. Pada saat ekor telah terbentuk sempurna maka embrio mulai bergerak di dalam telur. Pergerakan tersebut yang akan membantu embrio keluar dari telur (menetas). Lama perkembangan telur pada ikan koi hingga menetas diduga sama seperti pada ikan mas yaitu selama 3,5-4 hari setelah fertilisasi pada suhu inkubasi 20-22oC.
2.2 Promoter
Promoter merupakan sekuens DNA yang menginisiasi terjadinya proses transkripsi (Dunham, 2004), pengatur waktu, tempat, dan tingkat ekspresi suatu gen sehingga promoter dapat dianalogikan sebagai switch suatu gen (Glick dan
17 Pasternak, 2003). Promoter menurut Hackett (1993) merupakan sekuens DNA yang terletak pada posisi upstream (terminal 5) dari lokasi dimulainya transkripsi. Sekuens ini dikenali oleh RNA polymerase yang kemudian menempel dan mengendalikan proses transkripsi (Hackett, 1993; Glick dan Pasternak, 2003).
Berbagai macam promoter dapat aktif pada sel ikan walaupun bukan dari ikan yang homolog, namun akan mempengaruhi tingkat ekspresi gen (Dunham, 2004). Tingkat ekspresi gen dipengaruhi oleh adanya interaksi antara elemen cis-regulator pada promoter dan elemen trans-regulator inang. Hackett (1993) menjelaskan bahwa cis-regulator akan mengatur tingkat transkripsi bergantung pada keberadaan protein trans-regulator. Cis-regulator akan berikatan dengan protein atau faktor trans-regulator lainnya yang kemudian akan meningkatkan atau menurunkan tingkat transkripsi. Kesesuaian antara elemen cis-regulator dan elemen trans-regulator akan menghasilkan ekspresi yang tinggi, sebaliknya, bila kurang atau tidak sesuai maka ekspresi yang dihasilkan rendah (Fletcher dan Davies, 1991 dalamAth-thar, 2007).
Promoter memiliki beberapa sifat yang terkait dengan aktivitas elemen-elemennya yaitu constitutive, ubiquitous dan house keeping (Liu, 1990 dalam Volckaert et al., 1994). Promoter constitutive yaitu promoter yang dapat aktif tanpa diberikan rangsangan dari luar seperti suhu dan hormon. Ubiquitous yaitu promoter yang dapat mengendalikan ekspresi gen di semua jenis jaringan/sel. Namun, ada juga promoter yang dapat bekerja pada jaringan spesifik. Sedangkan house keeping yaitu promoter tersebut dapat aktif kapan saja bila diperlukan. Jenis promoter yang memiliki semua sifat tersebut di atas antara lain β-aktin (Volckaert et al., 1994). Jenis promoter lainnya ada yang memiliki sifat dapat diinduksi (inducible) yaitu promoter yang memerlukan faktor pemicu, misalnya promoter metallotionin yang aktivitasnya dipengaruhi oleh keberadaan logam berat (Iyengar et al., 1996), dan promoter hsp27 yang memerlukan tekanan suhu untuk meningkatkan aktivitasnya (Wu et al., 2008).
2.2.1 Promoter Keratin
Promoter keratin diisolasi dari ikan flounder Jepang Paralichthys olivaceus dan memiliki panjang fragmen 1288 pasang basa (Yazawa et al., 2005).
18 Sekuens gen keratin dapat dilihat pada Gambar 1. Promoter keratin ikan flounder Jepang (endogenus) diketahui memiliki aktivitas hampir di seluruh jaringan, sedangkan yang diuji coba pada ikan zebra (eksogenus) memiliki aktivitas terkuat pada jaringan epitel dan hati. Yazawa et al. (2005) menjelaskan bahwa beberapa faktor transkripsi pada jaringan epitel embrio ikan zebra dapat berikatan dengan baik sehingga aktivitas promoter keratin ikan flounder Jepang dapat meningkat. Jenis promoter keratin lainnya yang telah digunakan dalam transgenesis ikan yaitu keratin8 yang diisolasi dari ikan zebra dan ditemukan aktif hanya pada kulit (Gong et al., 2002). Kulit merupakan alat pertahanan awal terhadap serangan penyakit (Yazawa et al., 2005), sehingga promoter keratin diduga dapat digunakan untuk menghasilkan ikan transgenik tahan penyakit.
Keterangan : Huruf kapital adalah ekson 1. Huruf kapital yang dicetak tebal adalah coding region dan deduced amino acid. Tanda bintang (*) merupakan daerah inisiasi transkripsi. Huruf yang digarisbawahi merupakan faktor transkripsi.
Gambar 1. Sekuens Gen Keratin Ikan Flounder Jepang Paralichthys olivaceus (Yazawa et al., 2005).
2.2.2 Promoter Heat Shock
Promoter heat shock diisolasi dari ikan rainbow trout (Kawamura dan Yoshizaki, belum dipublikasikan) dan diketahui termasuk ke dalam golongan heat shock protein. Promoter heat shock memiliki panjang fragmen 2759 pasang basa. Jenis promoter lainnya yang termasuk ke dalam golongan heat shock protein antara lain adalah promoter hsp27. Promoter hsp27 merupakan jenis promoter
19 yang bersifat dapat diinduksi (inducible) yaitu memerlukan faktor pemicu. Promoter hsp27 memiliki aktivitas yang tinggi pada sel otot bahkan dapat aktif di seluruh jaringan jika dipicu dengan tekanan suhu (Wu et al., 2008). Lebih lanjut dijelaskan oleh Wu et al. (2008) bahwa terdapat pengaruh waktu pemberian suhu terhadap munculnya ekspresi gen Green Fluorescent Protein. Semakin akhir stadia perkembangan embrio yang diberi induksi suhu, semakin lama waktu yang dibutuhkan (lag time) untuk menginisiasi ekspresi gen Green Fluorescent Protein. Aktivitas seperti itu diduga juga dimiliki oleh promoter heat shock ikan rainbow trout.
Protein heat shock dapat ditemukan di seluruh makhluk hidup untuk merespon adanya perubahan suhu dan menghindari kerusakan sel akibat panas. Pada kondisi normal, heat shock ditemukan dalam konsentrasi yang rendah. Konsentrasi tinggi diperoleh ketika terjadi perubahan suhu secara signifikan (Fang, 2003). Toyohara et al. (2005) juga menyatakan bahwa heat shock berperan sebagai respon terhadap perubahan kondisi suhu lingkungan.
2.3 Gen Green Fluorescent Protein (GFP)
Gen Green Fluorescent Protein (GFP) dimanfaatkan untuk mempelajari promoter dan ekspresinya (Dunham, 2004). Gen GFP diisolasi dari ubur-ubur Aequorea victoria dan memiliki keunggulan yaitu tidak memerlukan substrat tambahan untuk ekspresinya, memiliki kandungan protein yang berpendar serta dapat divisualisasikan dengan menggunakan mikroskop fluoresen (Chalfie et al., 1994 dalam Iyengar et al., 1996). Gen GFP ini selain berguna sebagai penanda (marker) juga berfungsi sebagai gen target seperti dalam pembuatan ikan hias berpendar yang berwarna warni (Gong et al., 2003). Jenis gen GFP lainnya yang juga digunakan sebagai gen penanda yaitu yang diisolasi dari Anthozoa (soft coral) jenis Renilla reniformis yaitu gen hrGFP (Humanized Renilla reniformis Green Fluorescent Protein) (Felts et al., 2001).
2.4 Ekspresi Sementara (Transient Expression)
Ekspresi sementara (transient expression) merupakan ekspresi dari gen target yang dikendalikan oleh promoter tertentu dan bersifat sementara, biasanya
20 ekspresi gen pada embrio yang diinjeksi dengan gen target tersebut. Ekspresi sementara ini memiliki pola yang awalnya rendah, meningkat dan akhirnya menurun hingga tak terlihat (Winkler et al., 1991; Hackett, 1993; Volckaert et al., 1994; Takagi et al., 1994; Iyengar et al., 1996). Umumnya waktu awal ekspresi gen terjadi pada fase mid blastula transition dimana terjadi proses transkripsi yang mengakibatkan akumulasi protein pada sitoplasma telur sehingga gen dapat terekspresi (Iyengar et al., 1996). Lebih lanjut dijelaskan bahwa waktu ekspresi berhubungan erat dengan keberadaan DNA yang diinjeksikan. Peningkatan ekspresi gen yang terjadi merupakan akumulasi dari replikasi DNA yang diinjeksikan pada fase pembelahan awal, dan dari enzim produk transkripsi pada fase mid blastula transition. Setelah itu, terjadi penurunan ekspresi gen yang diakibatkan oleh degradasi DNA. Hal ini juga diungkapkan oleh Winkler et al. (1991) bahwa peningkatan ekspresi gen yang terjadi dikarenakan adanya replikasi DNA yang diinjeksikan di dalam embrio. Hal ini bergantung pada jumlah plasmid DNA yang diinjeksikan dan umumnya terjadi pada fase gastrula. Pada perkembangan embrio selanjutnya diduga sejumlah DNA asing mengalami degradasi sehingga terjadi penurunan jumlah DNA yang diinjeksikan dan mengakibatkan penurunan tingkat ekspresi gen.
Ekspresi sementara ini dapat ditemukan di seluruh jaringan (ubiquitous) atau hanya di jaringan tertentu dan umumnya masih bersifat mozaik pada ikan transgenik keturunan nol (F0, founder) (Volckaert et al., 1994; Iyengar et al., 1996; Dunham, 2004). Hal ini diduga karena adanya replikasi DNA asing yang tidak tersebar merata di dalam sel embrio (Iyengar et al., 1996). Selain itu, diduga terjadi replikasi yang berlebihan pada jaringan tertentu seperti sel otot sehingga ditemukan tingkat ekspresi tertinggi pada jaringan tersebut (William et al., 1996 dalam Iyengar et al., 1996).
2.5 Mikroinjeksi
Mikroinjeksi merupakan teknik transfer gen yang umum digunakan pada transgenesis (Takagi et al., 1994; Volckaert et al., 1994; Hamada et al., 1998; Alimuddin et al., 2003; Kato et al., 2007). Konstruksi DNA diintroduksikan ke dalam sel embrio ikan dengan menggunakan jarum injeksi berukuran sangat kecil.
21 Introduksi dilakukan di bawah mikroskop dengan bantuan mikromanipulator yang mengatur posisi jarum suntik. Untuk memastikan material genetik masuk ke pronuklei, konsentrasi DNA yang tinggi (104-107 copy) biasanya diinjeksikan ke telur yang telah dibuahi. Meskipun injeksi dengan jumlah copy DNA yang tinggi meningkatkan integrasi transgen (DNA yang ditranfer), tetapi hal itu meningkatkan resiko kematian pada embrio. Integrasi transgen pada DNA inang umumnya tidak terjadi pada fase satu sel, sehingga tidak semua sel ikan membawa transgen (mozaik) (Zbikwoska, 2003).
Beberapa keuntungan dengan menggunakan telur ikan sebagai inang yaitu jumlah telur ikan yang relatif banyak sehingga memudahkan dalam penyediaan inang, dan fertilisasi terjadi secara eksternal sehingga memudahkan introduksi gen asing pengkode protein target. Selain itu, embrio ikan dapat dipelihara dalam air tanpa suplemen, karena untuk perkembangan embrio cukup mengandalkan nutrien dari kuning telur (Dunham, 2004).
22
III. BAHAN DAN METODE
3.1 Waktu dan Tempat
Penelitian ini terdiri dari tahap persiapan yaitu perbanyakan konstruksi DNA di Laboratorium Pengembangan dan Genetika Ikan, Departemen Budidaya Perairan, Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor, dan tahap utama di Laboratorium Pengembangbiakan Ikan Mas dan Laboratorium Kesehatan Ikan, Balai Besar Pengembangan Budidaya Air Tawar (BBPBAT) Sukabumi. Waktu pelaksanaan penelitian adalah bulan Agustus-September 2008.
3.2 Prosedur Kerja
3.2.1 Perbanyakan Konstruksi DNA
Bakteri Eschericia coli yang mengandung konstruksi plasmid DNA keratin-GFP dan heat shock-GFP (Lampiran 2) diperbanyak dengan metode kultur cair (Lampiran 3). Bakteri dipanen dan dikultur dalam media cair yang mengandung Triptone 1,6%, yeast extract 1%, NaCl 0,5%, dan antibiotik kanamisin, diinkubasi menggunakan shaker dengan kecepatan 250 rpm pada suhu 37oC, selama 16-18 jam. Kemudian, bakteri hasil kultur dimasukkan ke dalam microtube 1,5 ml, disentrifus pada kecepatan 12000 rpm selama 30 detik. Supernatan dibuang, pelet plasmid DNA yang terbentuk diisolasi dengan kit EZ 10 Spin Column Plasmid DNA (Lampiran 3). Konsentrasi larutan DNA awal dihitung dengan menggunakan GeneQuant, kemudian dibuat konsentrasi larutan DNA untuk mikroinjeksi sebesar 50 ng/µl (Kobayashi et al., 2007).
3.2.2 Pemeliharaan Induk, Pemijahan, dan Pengambilan Telur
Induk jantan dan betina ikan koi dipelihara di Balai Besar Pengembangan Budidaya Air Tawar (BBPBAT) Sukabumi pada bak semen. Pakan untuk induk adalah pelet yang diberikan 2 kali sehari secara satiasi. Sistem pemijahan dilakukan dengan sistem semi alami (Lampiran 4). Ovaprim dengan dosis 0,5 ml/kg induk disuntikkan ke induk jantan dan betina yang telah matang gonad. Pengurutan (stripping) dilakukan 12 jam setelah penyuntikan pada induk betina dan jantan untuk mendapatkan telur dan sperma. Sperma yang telah diperoleh
23 disimpan pada larutan fisiologis. Kemudian, telur dan sperma dicampur dan diaduk perlahan menggunakan bulu ayam. Telur dan sperma tersebut diberi air bersih sehingga pembuahan dapat terjadi (Woynarovich dan Horvath, 1980).
3.2.3 Penghilangan Daya Rekat Telur
Telur ikan koi yang telah dibuahi bersifat adesif, yaitu melekat pada substrat. Penghilangan daya rekat telur diperlukan untuk memudahkan proses mikroinjeksi. Untuk menghilangkan daya rekat telur, setelah pembuahan, telur direndam dengan larutan Tannin (0,5 gram Tannin/liter akuades) (Woynarovich dan Horvath, 1980) yang masih baru selama 3-5 detik kemudian segera dibilas dengan air bersih sebanyak 2 kali. Lalu, telur diambil dan diletakkan pada cekungan agarosa (Gambar 2) untuk selanjutnya dilakukan penginjeksian.
Gambar 2. Cekungan Agarosa
3.2.4 Pelaksanaan Mikroinjeksi
Larutan DNA sebanyak 4 µl dipipet menggunakan tip loading kemudian dimasukkan ke bagian ujung dalam jarum mikroinjeksi. Minyak mineral ditambahkan ke dalam jarum mikroinjeksi menggunakan jarum minyak mineral yang telah dipasang pada needle holder. Jarum minyak mineral dilepas dan jarum mikroinjeksi yang telah berisi larutan DNA dan minyak mineral disambungkan ke needle holder pada seperangkat alat mikroinjektor (Gambar 3).
Embrio ikan koi diletakkan dan diatur sedemikian rupa di dalam cekungan agarosa. Larutan DNA diinjeksikan ke blastodisk embrio ikan koi pada saat fase Cekungan (Tempat telur) Gel
24 pembelahan 1 (Gambar 4) sampai 2 sel secara perlahan dengan bantuan mikromanipulator (Gambar 3c) untuk menggerakkan jarum mikroinjeksi. Mikroinjeksi dilakukan di bawah mikroskop (Gambar 3a).
Gambar 3. Seperangkat Alat Mikroinjektor
Perlakuan injeksi pada penelitian ini terdiri dari injeksi gen keratin-GFP dan gen heat shock -GFP dilakukan sebanyak 3 kali ulangan. Jumlah embrio ikan koi yang diinjeksi sebanyak 30 butir tiap perlakuan dan ulangan.
Gambar 4. Injeksi pada Blastodisk Embrio Ikan Koi Fase Satu Sel. Jarum injeksi Keterangan : A. Mikroskop B. Needle holder C. Mikromanipulator D. Injektor dan magnetic stand Embrio Ikan Koi Blastodisk embrio
25
3.2.5 Penetasan Telur dan Pemeliharaan Larva
Telur-telur yang telah dibuahi dan diinjeksi dipindahkan ke dalam akuarium inkubasi yang telah diberi methylen blue dengan kepadatan 30 butir embrio tiap wadah dan suhu air berkisar 28oC. Embrio yang mati dan mengalami perubahan bentuk dibuang selama masa pengamatan.
3.2.6 Pengamatan Ekspresi Gen Green Fluorescent Protein
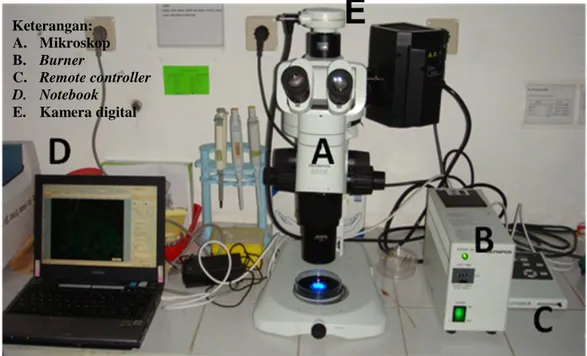
Pengamatan ekspresi gen Green Fluorescent Protein (GFP) dilakukan dengan menggunakan mikroskop fluoresen (Olympus SZX 16) yang dilengkapi filter GFP (Olympus SZX2-GFPHQ) (Gambar 5a) dan burner (Olympus) (Gambar 5b) pada perbesaran 1,25 kali. Pengamatan dilakukan tiap 2-6 jam setelah fertilisasi dan dimulai pada jam ke-1 setelah injeksi. Embrio dan larva difoto dengan menggunakan kamera digital High Speed Compact Color 2 megapiksel (DP20) (Gambar 5e) kemudian ditransfer ke notebook Toshiba (Gambar 5d) yang memiliki software Olympus DH2-BW melalui remote controller (Olympus DP-20) (Gambar 5c).
Gambar 5. Seperangkat Alat Pengamatan Ekspresi Gen Green Fluorescent Protein Keterangan: A. Mikroskop B. Burner C. Remote controller D. Notebook E. Kamera digital
26
3.3 Pengamatan
3.3.1 Derajat Kelangsungan Hidup Embrio (DKH-e)
Derajat kelangsungan hidup embrio adalah persentase jumlah embrio yang hidup dibandingkan jumlah embrio awal. Perhitungan dilakukan 20 jam setelah fertilisasi, dimana embrio belum menetas dengan rumus perhitungan sebagai berikut:
3.3.2 Derajat Penetasan (DP)
Derajat penetasan adalah persentase jumlah embrio yang menetas dibandingkan jumlah embrio awal. Perhitungan dilakukan ketika larva telah menetas secara keseluruhan dengan rumus perhitungan sebagai berikut :
3.3.3 Persentase Embrio Mengekspresikan Gen GFP (PEMG)
Persentase embrio mengekspresikan gen GFP diperoleh dari perbandingan jumlah embrio yang mengekspresikan gen GFP dengan jumlah total embrio yang diinjeksi. Perhitungan dilakukan pada jam ke-12 dengan rumus perhitungan sebagai berikut :
3.3.4 Persentase Larva Mengekspresikan Gen GFP (PLMG)
Persentase larva mengekspresikan gen GFP diperoleh dari perbandingan jumlah larva yang mengekspresikan gen GFP dibandingkan dengan jumlah total embrio diinjeksi yang menetas. Perhitungan dilakukan dengan rumus sebagai berikut :
27
3.3.5 Tingkat dan Pola Ekspresi Gen Green Fluorescent Protein
Tingkat ekspresi gen Green Fluorescent Protein (GFP) dikelompokkan secara kualitatif berdasarkan intensitas pendarannya menjadi 3 kelompok yaitu ekspresi pendar hijau kurang terang, ekspresi pendar hijau terang, dan ekspresi pendar hijau sangat terang. Ekspresi pendar hijau kurang terang yaitu pendaran berwarna hijau yang terlihat sedikit terang, ekspresi pendar hijau terang yaitu pendaran berwarna hijau yang sedikit lebih terang dibandingkan ekspresi pendar hijau kurang terang, dan ekspresi pendar hijau sangat terang yaitu pendaran berwarna hijau yang lebih terang dibandingkan lainnya. Pengamatan tingkat ekspresi gen GFP dilakukan pada fase embrio pada jam ke-12 setelah fertilisasi. Pengamatan pola ekspresi dilakukan pada jam ke-1 setelah fertilisasi setiap 2-6 jam hingga larva menetas.
3.4 Analisis Data
Data yang telah diperoleh, dianalisis secara deskriptif dan disajikan dalam bentuk tabel, grafik, serta gambar.
28
IV. HASIL DAN PEMBAHASAN
4.1 Hasil
4.1.1 Derajat Kelangsungan Hidup Embrio (DKH-e), Derajat Penetasan (DP), Persentase Embrio Mengekspresikan Gen GFP (PEMG), dan Persentase Larva Mengekspresikan Gen GFP (PLMG)
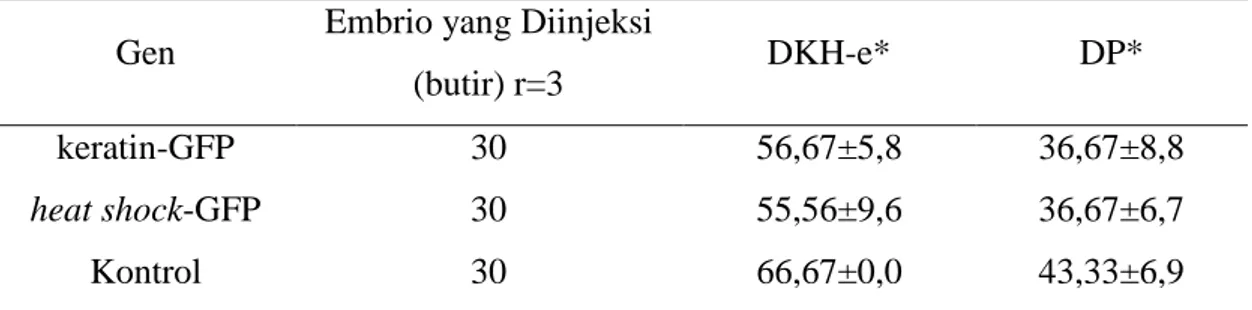
Derajat kelangsungan hidup embrio dan derajat penetasan pada perlakuan injeksi memiliki nilai yang lebih rendah dibandingkan kontrol/tidak diberikan perlakuan injeksi (Tabel 1). Derajat kelangsungan hidup embrio yang diinjeksi mempunyai nilai yang relatif sama baik pada gen keratin-GFP (56,67±5,8%) maupun heat shock-GFP (55,56±9,6%). Hal serupa juga terjadi pada derajat penetasan pada gen keratin-GFP dan heat shock-GFP yang memiliki nilai relatif sama, yaitu (36,67±8,8%) dan (36,67±6,7%). Data keseluruhan derajat kelangsungan hidup embrio dan derajat penetasan disajikan pada Lampiran 5.
Tabel 1. Derajat Kelangsungan Hidup Embrio, dan Derajat Penetasan Telur Ikan Koi Cyprinus carpio.
Gen Embrio yang Diinjeksi
(butir) r=3 DKH-e* DP*
keratin-GFP 30 56,67±5,8 36,67±8,8
heat shock-GFP 30 55,56±9,6 36,67±6,7
Kontrol 30 66,67±0,0 43,33±6,9
* r = Ulangan
DKH-e = Derajat kelangsungan hidup embrio (%, rataan ± standar deviasi)
DP = Derajat Penetasan (%, rataan ± standar deviasi)
Berdasarkan Tabel 2, persentase embrio yang mengekspresikan gen heat shock-GFP (33,33±12,0%) lebih tinggi dibandingkan dengan gen keratin-GFP (15,56±8,4%). Sementara itu, larva yang mengekspresikan gen GFP terlihat hanya pada embrio yang diinjeksi gen heat shock-GFP (18,18±13,7%). Data keseluruhan persentase embrio dan larva yang mengekspresikan gen GFP disajikan pada Lampiran 6.
29 Tabel 2. Persentase Embrio dan Larva yang Mengekspresikan Gen Keratin-GFP
dan Heat Shock-GFP pada Ikan Koi Cyprinus carpio. Gen Embrio yang Diinjeksi
(butir) r=3 PEMG* PLMG*
keratin-GFP 30 15,56±8,4 -
heat shock-GFP 30 33,33±12,0 18,18±13,7
* r = Ulangan
PEMG = Persentase embrio yang mengekspresikan gen GFP (%, rataan ± standar deviasi) PLMG = Persentase larva yang mengekspresikan gen GFP (%, rataan ± standar deviasi)
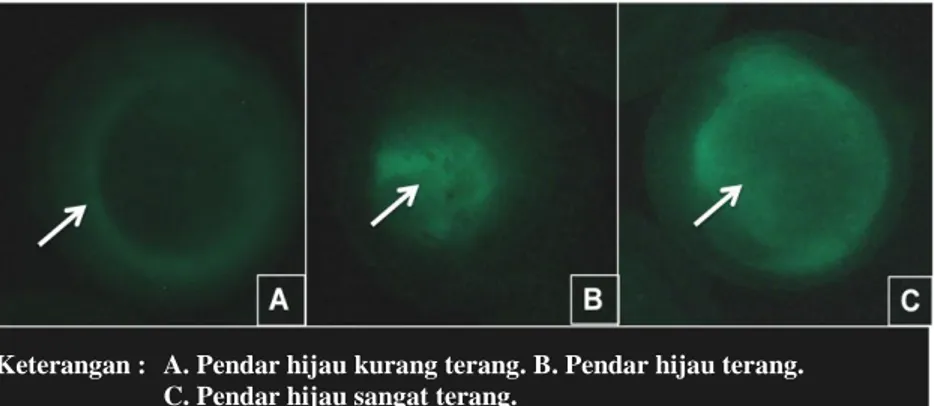
4.1.2 Tingkat dan Pola Ekspresi Gen Green Fluorescent Protein
Tingkat ekspresi yang dihasilkan dari perlakuan dibagi menjadi tiga kelompok dan disajikan pada Gambar 6. Perbandingan tingkat ekspresi antar konstruksi DNA diamati pada jam ke-12 dan disajikan pada Tabel 3.
Keterangan : A. Pendar hijau kurang terang. B. Pendar hijau terang.
C. Pendar C. Pendar hijau sangat terang.
Gambar 6. Kelompok Tingkat Ekspresi Gen Green Fluorescent Protein pada Embrio Ikan Koi Cyprinus carpio.
Tabel 3. Tingkat Ekspresi Gen Keratin-GFP dan Heat Shock-GFP pada Embrio Ikan Koi Cyprinus carpio.
Gen Tingkat Ekspresi Gen* Jumlah embrio yang
terekspresi*
1 2 3
keratin-GFP 2,00±1,0 1,67±1,5 1,00±1,0 4,67±2,5 heat shock-GFP 4,67±1,5 3,00±1,0 2,33±1,5 10,00±3,6 Keterangan : 1. Pendar hijau kurang terang. 2. Pendar hijau terang. 3. Pendar hijau sangat terang
*(jumlah telur ± standar deviasi)
Promoter heat shock mampu mengendalikan ekspresi gen GFP lebih baik dibandingkan promoter keratin berdasarkan Tabel 3. Hal ini dapat dilihat dari
30 rata-rata jumlah embrio yang mengekspresikan gen GFP dari masing-masing kelompok tingkat ekspresi gen. Heat shock-GFP memiliki pendar hijau kurang terang sebanyak 4,67±1,5 butir lebih tinggi dari keratin-GFP (2,00±1,0 butir). Pendar hijau terang yang dihasilkan heat shock-GFP sebanyak 3,00±1,0 butir lebih tinggi dari keratin-GFP (1,67±1,5 butir). Pendar hijau sangat terang yang dihasilkan heat shock-GFP sebanyak 2,33±1,5 butir lebih tinggi dari keratin-GFP (1,00±1,0 butir). Data keseluruhan tingkat ekspresi gen keratin-GFP dan heat shock-GFP disajikan pada Lampiran 7.
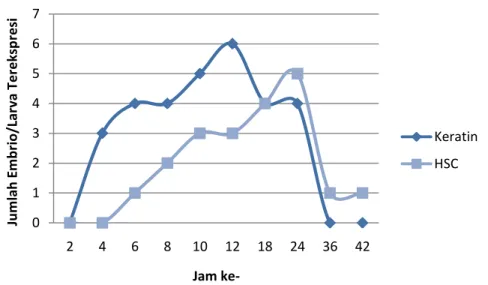
Gambar 7. Pola Ekspresi Gen Keratin-GFP dan Heat Shock-GFP pada Embrio Ikan Koi Cyprinus carpio.
Pola ekspresi gen keratin-GFP dan heat shock-GFP pada embrio ikan koi (Gambar 7, 8 dan 9) dapat diketahui bahwa ekspresi gen keratin-GFP mulai terlihat pada fase blastula (jam ke-4 setelah fertilisasi), hal serupa juga terjadi pada heat shock-GFP yang mulai terlihat pada fase blastula (jam ke-6 setelah fertilisasi). Ekspresi gen keratin-GFP terkuat dilihat dari banyaknya jumlah embrio yang mengekspresikan gen GFP yaitu pada fase gastrula (jam ke-12 setelah fertilisasi), sedangkan pada heat shock-GFP terkuat terlihat pada fase perkembangan organogenesis (jam ke-24 setelah fertilisasi). Ekspresi gen keratin-GFP terlihat mulai melemah pada fase perkembangan organogenesis (jam ke-36 setelah fertilisasi) dan sudah tidak tampak lagi pada fase larva. Sedangkan ekspresi gen heat shock-GFP masih tetap ada hingga larva berumur 1 hari.
0 1 2 3 4 5 6 7 2 4 6 8 10 12 18 24 36 42 Ju m lah E m b rio /La rva Te re ksp re si Jam ke-Keratin HSC
31 Gambar 8. Ekspresi Gen Heat Shock-GFP pada Embrio Ikan Koi 6 Jam setelah Fertilisasi (A), 12 Jam setelah Fertilisasi (B), 24 Jam setelah Fertilisasi (C), pada Larva 30 Jam setelah Menetas di Sebagian Jaringan Tubuh (D) dan Hampir di Seluruh Tubuh (E).
Gambar 9. Ekspresi Gen Keratin-GFP pada Embrio Ikan Koi 4 Jam setelah Fertilisasi (A), 12 Jam setelah Fertilisasi (B), 24 Jam setelah Fertilisasi (C).
32
4.2 Pembahasan
Telur ikan koi yang digunakan saat penelitian memiliki kualitas yang cukup bagus, dilihat dari nilai rata-rata derajat kelangsungan hidup dan derajat penetasan antara kontrol dengan perlakuan yang tidak berbeda jauh. Namun, nilai derajat kelangsungan hidup dan derajat penetasan dari perlakuan masih lebih rendah jika dibandingkan kontrol (tanpa perlakuan injeksi). Hal ini mungkin disebabkan karena kerusakan yang terjadi pada sel embrio setelah diinjeksi sehingga perkembangan embrio menjadi tidak normal dan kemudian mengalami kematian. Selain itu, juga diduga akibat tingginya volume larutan DNA yang diinjeksikan. Transfer gen dengan metode mikroinjeksi umumnya membutuhkan larutan DNA yang diinjeksikan dalam jumlah copy yang tinggi. Hal ini dilakukan untuk meningkatkan integrasi transgen ke dalam genom inang (Zbikwoska, 2003). Namun demikian, semakin tinggi jumlah copy DNA yang diinjeksikan juga akan meningkatkan mutagenesis atau meningkatkan jumlah partikel asing yang masuk dalam embrio, sehingga dapat mengganggu stabilitas embrio dan menyebabkan kematian (Hackett, 1993).
Persentase embrio yang mengekspresikan gen GFP pada heat shock-GFP lebih tinggi dibandingkan pada keratin-GFP, dan hanya pada heat shock-GFP yang mengekspresikan hingga fase larva. Ekspresi sementara (transient expression) yang dihasilkan menunjukkan bahwa promoter keratin dan heat shock dapat aktif mengendalikan ekspresi gen GFP dan menunjukkan pola ekspresi sementara (transient expression) gen GFP yang berbeda pada ikan koi, sehingga dapat digunakan dalam pembuatan ikan transgenik. Perbedaan persentase embrio yang mengekspresikan gen GFP juga terjadi pada penelitian Kato et al. (2007), bahwa konstruksi DNA (β-actin ikan kakap merah red sea bream + GFP + polyA β-actin ikan kakap merah red sea bream) yang terminatornya merupakan bagian dari gen β-actin itu sendiri, menghasilkan persentase embrio yang mengekspresikan gen GFP lebih tinggi dibandingkan konstruksi DNA yang terminatornya berupa polyA SV40 (polyadenylation simian virus 40). Pada pengamatan hari ketiga setelah injeksi, persentase embrio yang mengekspresikan gen GFP pada konstruksi DNA β-actin ikan kakap merah red sea bream + GFP + polyA β-actin ikan kakap merah red sea bream yaitu sebesar 17,1±9,8%,
33 sedangkan pada konstruksi DNA GFP+ polyA SV 40 sebesar 0,00±0,0%. Kato et al. (2007) menduga bahwa perbedaan persentase embrio yang mengekspresikan gen GFP disebabkan oleh adanya perbedaan pengaturan transkripsi dari masing-masing konstruksi. Perbedaan pengaturan transkripsi ini diduga juga terjadi pada konstruksi DNA keratin-GFP dan heat shock-GFP dilihat dari perbedaan persentase embrio dan larva yang mengekspresikan gen GFP. Keratin-GFP memiliki terminator berupa SV40 sedangkan heat shock-GFP memiliki terminator berupa 3’UTR heat shock yang merupakan bagian dari gen heat shock itu sendiri.
Pola ekspresi sementara yang dihasilkan gen keratin-GFP mulai terlihat pada jam ke-4 kemudian mencapai puncak pada jam ke-12 lalu menurun pada jam ke-36 hingga tidak terlihat lagi pada fase larva. Sedangkan, ekspresi sementara pada gen GFP di heat shock-GFP mulai terlihat pada jam ke-6, meningkat pada jam ke-24, dan masih ada hingga larva berumur 1 hari. Pola ekspresi sementara seperti ini umumnya terjadi pada banyak pengujian aktivitas promoter antara lain pada ikan medaka (Winkler et al., 1991; Takagi et al., 1994), ikan lele Afrika (Volckaert et al., 1994), ikan zebra (Higashijima et al., 1997), ikan kakap merah (Kato et al., 2007), ikan lele (Ath-thar, 2007), dan ikan mas (Purwanti, 2007) dengan menggunakan promoter yang berbeda pula. Pola ekspresi gen yang terbentuk umumnya hampir sama walaupun ada perbedaan waktu ekspresi gen antara satu promoter dengan promoter lainnya pada spesies ikan yang berbeda, yaitu pada awalnya rendah, meningkat, kemudian menurun hingga tidak terlihat lagi. Perbedaan waktu yang terjadi diduga karena tiap embrio memiliki kemampuan berkembang yang berbeda dimana dipengaruhi oleh laju transkripsi sel dalam embrio dan suhu inkubasi telur. Volckaert et al. (1994) menjelaskan bahwa pola waktu ekspresi gen asing bergantung pada pola perkembangan embrio. Woynarovich dan Horvath (1980) juga menambahkan bahwa laju perkembangan embrio bergantung pada suhu inkubasi. Hal ini dikarenakan di dalam embrio terdapat sejumlah enzim yang berperan terhadap perkembangannya. Ekspresi gen keratin-GFP dan heat shock-GFP mulai terlihat pada fase blastula yaitu pada fase terbentuknya rongga yang membedakan antara kuning telur dengan sel (Woynarovich dan Horvath, 1980). Umumnya waktu awal ekspresi gen terjadi pada fase mid blastula transition (MBT) dimana terjadi proses
34 transkripsi yang mengakibatkan akumulasi protein pada sitoplasma telur sehingga gen dapat terekspresi (Iyengar et al., 1996). Lebih lanjut dijelaskan bahwa waktu ekspresi berhubungan erat dengan keberadaan DNA yang diinjeksikan. Puncak ekspresi atau ekspresi terkuat yang dihasilkan dari kedua perlakuan diduga disebabkan terjadinya replikasi DNA yang diinjeksikan di dalam embrio pada fase perkembangan awal (Winkler et al., 1991). Sedangkan, perbedaan waktu puncak ekspresi antara gen keratin-GFP dan heat shock-GFP yang terjadi diduga dikarenakan terdapat perbedaan jumlah plasmid DNA yang diinjeksikan. Perbedaan jumlah plasmid DNA ini terjadi diduga karena adanya perbedaan ukuran bukaan jarum yang digunakan selama proses penginjeksian. Pada penelitian ini, tidak dilakukan kuantifikasi jumlah plasmid DNA yang diinjeksikan sehingga tidak diketahui secara pasti jumlah plasmid DNA yang masuk ke dalam blastodisk embrio ikan koi. Selain itu, gen heat shock-GFP yang dikendalikan oleh promoter heat shock memiliki kemampuan mengekspresikan suatu gen bergantung pada tekanan suhu (Wu et al., 2008) dan diduga ekspresi terkuat gen heat shock-GFP akan lebih cepat terlihat jika diikuti dengan peningkatan suhu inkubasi. Peningkatan ekspresi gen yang terjadi ditambahkan oleh Iyengar et al. (1996) merupakan akumulasi dari enzim produk transkripsi pada fase mid blastula transition. Ekspresi gen keratin-GFP terlihat mulai melemah pada fase perkembangan organogenesis (jam ke-36 setelah fertilisasi) dan sudah tidak tampak lagi pada fase larva. Sedangkan ekspresi gen heat shock -GFP masih tetap ada hingga larva berumur 1 hari walaupun intensitas pendarannya juga mengalami penurunan. Hal ini diduga dikarenakan sejumlah DNA yang diinjeksikan telah mengalami degradasi sehingga terjadi penurunan jumlah DNA dan mengakibatkan penurunan tingkat ekspresi gen.
Promoter heat shock mampu mengendalikan ekspresi gen GFP lebih baik dibandingkan promoter keratin. Hal ini dapat dilihat dari rata-rata jumlah embrio yang mengekspresikan gen GFP pada masing-masing kelompok tingkat ekspresi gen. Perbedaan tingkat ekspresi gen GFP yang terjadi diduga karena elemen cis -regulator heat shock dapat berikatan lebih baik atau lebih sesuai dengan elemen trans-regulator ikan koi. Perbedaan tingkat ekspresi dijelaskan oleh Dunham (2004) yaitu disebabkan karena promoter yang diintroduksikan bukan berasal dari
35 ikan yang homolog. Promoter yang bukan berasal dari ikan yang homolog memiliki interaksi antara elemen cis-regulator pada promoter dan elemen trans -regulator inang yang berbeda. Hackett (1993) juga menambahkan bahwa elemen cis-regulator akan berikatan dengan trans-regulator protein lainnya yang kemudian akan mengakibatkan peningkatan atau penurunan tingkat transkripsi. Fletcher dan Davies (1991) dalam Ath-thar (2007) menjelaskan bahwa tingkat ekspresi yang tinggi dipengaruhi oleh kesesuaian antara elemen cis-regulator dan trans-regulator.
36
V. KESIMPULAN
5.1 Kesimpulan
Promoter heat shock ikan rainbow trout dapat mengendalikan ekspresi gen Green Fluorescent Protein lebih baik dibandingkan dengan promoter keratin ikan flounder Jepang pada ikan koi.
5.2 Saran
Promoter heat shock ikan rainbow trout dianjurkan untuk digunakan pada pembuatan ikan koi transgenik tahan terhadap penyakit.
37
DAFTAR
PUSTAKA
Ath-thar MHF. 2007. Efektivitas promoter β-actin ikan medaka Oryzias latipes dengan penanda gen hrGFP (Humanized Renilla reniformis Green Fluorescent Protein) pada ikan lele Clarias sp. Keturunan F0. Skripsi. Departemen Budidaya Perairan. Perikanan dan Ilmu Kelautan. Institut Pertanian Bogor.
Alimuddin, Yoshizaki G, Carman O, dan Sumantadinata K. 2003. Aplikasi transfer gen dalam akuakultur. Jurnal Akuakultur Indonesia. 2(1) : 41-50. Dunham RA, Warr G, Nichols AJ, Duncan PL, Argue B, Middleton D, and Liu Z.
2002. Enhanced bacterial disease resistance of transgenic channel catfish, Ictalurus punctatus, possessing cecropin genes. Marine Biotechnology 4 : 338-344.
Dunham RA. 2004. Aquaculture and Fisheries Biotechnology: Genetic Approaches. Cambridge: CABI Publishing.
Fang F. 2003. Expression of heat shock genes HSP 16.6 and HTPG in the cyanobacterium Synechocystis sp. PCC 6803. Disertasion. Miami University.
Felts K, Rogers B, Chen K, Ji H, Sorge J, and Vaillancourt P. 2001. Recombinant Renilla reniformis GFP displays low toxicity. Stratagene 13: 85-87.
Glick BR and Pasternak JJ. 2003. Molecular Biotechnology: Principles and Application of Recombinant DNA. 3rd ed. Washington DC: ASM Press. Gong Z, Ju B, Wang X, He J, Wan H, Sudha PM, and Yan T. 2002. Green
fluorescent protein expression in germ-line transmitted transgenic zebrafish under a stratified epithelial promoter from keratin8. Developmental Dynamics 223: 204-215.
Gong Z, Wan H, Ju B, He J, Wang X, and Yan T. 2003. Generation of living color transgenic fish. In : Shimizu N, Aoki T, Hirono I, Takashima F. (eds) Aquatic Genomics: Steps Toward a Great Future. (pp.329-339). Springer-Verlag. New York.
38 Hackett PB. 1993. The molecular biology of transgenic fish. In: Hochachka and Mommesen (Eds.). Biochemistry and Molecular Biology of Fishes 2: 218-221.
Hamada K, Tamaki K, Sasado T, Watai Y, Kani S, Wakamatsu Y, Ozato K, Kinoshita M, Kohno R, Takagi S, and Kimura M. 1998. Usefulness of the medaka β-actin promoter investigated using a mutant GFP reporter gene in transgenic medaka Oryzias latipes. Molecular Marine Biology and Biotechnology 7: 173-180.
Higashijima S, Okamoto H, Ueno N, Hotta Y, and Eguchi G. 1997. High-frequency generation of transgenic zebrafish which reliably express GFP in whole muscle or the whole body by using promoters of zebrafish origin. Development Biology 192: 289-299.
Iyengar A, Muller F, and Maclean N. 1996. Regulation and expression of transgenes in fish-a review. Transgenic Research 5 : 147-166.
Kato K, Takagi M, Tamaru Y, Akiyama S, Konishi T, Murata O, and Kumai H. 2007. Construction of an expression vector containing a β-actin promoter region for gene transfer by microinjection in red sea bream Pagrus major. Fisheries Science 73 : 440-445.
Kobayashi SI, Alimuddin, Morita T, Miwa M, Lu J, Endo M, Takeuchi T, and Yoshizaki G. 2007. Transgenic Nile tilapia (Oreochromis niloticus) over-expressing growth hormone show reduced ammonia excretion. Aquaculture 270 : 427-435.
Nam YK, Noh JK, Cho YS, Cho HJ, Cho KN, Kim CG, and Kim DS. 2001. Dramatically accelerated growth and extraordinary gigantism of transgenic mud loachMisgurnus mizolepi.Transgenic Research 10: 353-362.
Purwanti LI. 2007. Uji aktivitas promoter β-actin ikan medaka (Oryzias latipes) dengan penanda gen hrGFP (Humanized Renilla reniformis Green Fluorescent Protein). Skripsi. Departemen Budidaya Perairan. Perikanan dan Ilmu Kelautan. Institut Pertanian Bogor
Sarmaşik A. 2003. Application of gene transfer technology for genetic improvement of fish-a review. Turkey Journal Zoology 27 : 1-6.
39 Takagi S, Sasado G, Tamiya G, Ozato K, Wakamatsu Y, Takeshita A, and Kimura M. 1994. An efficient expression vector for transgenic medaka construction. Molecular Marine Biology and Biotechnology 3: 192-199. Toyohara H, Hosoi M, Hayashi I, Kubota S, Hashimoto H, and Yokoyama Y.
2005. Expression of HSP70 in response to heat-shock and its cDNA cloning from Mediterranean blue mussel. Fisheries Science 71: 327-332. Volckaert FA, Hellemans BA, Galbusera P, and Ollevier F. 1994. Replication,
expression and fate of foreign DNA during embryonic and larval development of the African catfish Clarias gariepinus. Molecular Marine Biology and Biotechnology 3: 57-69.
Winkler C, Vielkind JR, and Schartl M. 1991. Transient expression of foreign DNA during embryonic and larval development of the medaka fish Oryzias latipes. Mol. Gen. Genet 226 : 129-140.
Woynarovich E and Horvath L. 1980. The Artificial Propagation of Warm Water Finfishes – A Manual for Extension. FAO Fisheries Technical Paper 201: 183p.
Wu YL, Pan X, Mudumana SP, Wang H, Kee PW, and Gong Z. 2008. Developmental of a heat shock inducible gfp transgenic zebrafish line by using the zebrafish hsp27 promoter. Gene 408: 85-94.
Yazawa R, Hirono I, and Aoki T. 2005. Characterization of promoter activities of four different Japanese flounder promoters in transgenic zebrafish. Marine Biotechnology 7: 625-633.
Zbikowska, HM. 2003. Fish can be First – Advances in Fish Transgenesis for Commercial Applications. Transgenic Research 12:379-389
40
LAMPIRAN
Lampiran 1. Perkembangan Embriogenesis Telur Ikan Mas Cyprinus carpio (Woynarovich dan Horvath, 1980).
Telur setelah dibuahi Pembelahan 2 sel Stadia 4 sel
Morula (awal) Morula (akhir) Blastula
Gastrula Penutupan blastopor
41 Lampiran 1. Perkembangan Embriogenesis Telur Ikan Mas Cyprinus carpio
42 Lampiran 2. Konstruksi Plasmid DNA Keratin-GFP (Yazawa et al., 2005) (A) dan Heat Shock-GFP (Yamamoto dan Yoshizaki, tidak dipublikasikan) (B).
Keterangan :
GFP = Green Fluorescent Protein SV40 = Simian virus 40
43 Lampiran 3. Metode Kultur Cair Perbanyakan Bakteri dan Isolasi Plasmid DNA
Keratin-GFP dan Heat Shock-GFP.
Keterangan :
= metode kultur cair
= metode isolasi plasmid
Endapan dibuang, sentrifuse 10000
44 Lampiran 4. Proses Pemijahan Semi Alami Ikan Koi Cyprinus carpio.
Keterangan :
1. Penyuntikan ovaprim pada induk ikan koi. 2. Penyatuan induk ikan koi jantan dan betina.
3. Pengejaran induk ikan koi betina oleh ikan koi jantan.
4. Pengeluaran telur oleh induk ikan koi betina disertai lompatan ke arah kakaban dan diikuti pengeluaran sperma oleh induk ikan koi jantan.
45 Lampiran 5. Derajat Kelangsungan Hidup Embrio, dan Derajat Penetasan Telur
Ikan Koi Cyprinus carpio.
Gen Ulangan n DKH-e DP
keratin-GFP 1 30 50.00 % 26.67 % 2 30 60.00 % 40.00 % 3 30 60.00 % 43.33 % Rata-rata 30 56.67 % 36.67 % Standar deviasi - 5.8 8.8 heat shock-GFP 1 30 50.00 % 36.67 % 2 30 66.67 % 30.00 % 3 30 50.00 % 43.33 % Rata-rata 30 55.56 % 36.67 % Standar deviasi - 9.6 6.7 Kontrol 1 30 66.67 % 36.67 % 2 30 66.67 % 40.00 % 3 30 66.67 % 50.00 % Rata-rata 30 66.67 % 42.22 % Standar deviasi - - 6.9 Keterangan :
n = jumlah embrio yang diinjeksi (butir) DKH-e = Derajat Kelangsungan Hidup Embrio (%) DP = Derajat Penetasan (%)
46 Lampiran 6. Persentase Embrio yang Mengekspresikan Gen GFP, dan Persentase
Larva yang Mengekspresikan Gen GFP Ikan Koi Cyprinus carpio.
Gen Ulangan n PEMG PLMG
keratin-GFP 1 30 16.67 % - 2 30 6.67 % - 3 30 23.33 % - Rata-rata 30 15.56 % - Standar deviasi - 8.4 - heat shock-GFP 1 30 43.33 % 27.00 % 2 30 36.67 % - 3 30 20.00 % 15.00 % Rata-rata 30 33.33 % 18.18 % Standar deviasi - 12.0 13.7 Keterangan :
n = jumlah embrio yang diinjeksi (butir)
PEMG = Persentase Embrio yang Mengekspresikan Gen GFP PLMG = Persentase Larva yang Mengekspresikan Gen GFP
47 Lampiran 7. Tingkat Ekspresi Gen Green Fluorescent Protein pada Embrio Ikan Koi Cyprinus carpio.
Keterangan : *1. Pendar hijau kurang terang. 2. Pendar hijau terang. 3. Pendar hijau sangat terang.
Gen Ulangan Tingkat Ekspresi* Jumlah embrio
yang terekspresi
1 2 3
keratin-GFP
1 3 butir 42.86 % 2 butir 28.57 % 2 butir 28.57 % 7 butir
2 2 butir 100.00 % - - - - 2 butir
3 1 butir 20.00 % 3 butir 60.00 % 1 butir 20.00 % 5 butir Rata-rata 2.00 butir 54.29 % 1.67 butir 29.52 % 1.00 butir 16.19 % 4.67 butir
Standar deviasi 1.0 41.21 1.5 30.01 1.0 14.66 2.5
heat shock-GFP
1 6 butir 46.15 % 3 butir 23.08 % 4 butir 30.77 % 13 butir 2 3 butir 50.00 % 2 butir 33.33 % 1 butir 16.67 % 6 butir 3 5 butir 45.45 % 4 butir 36.36 % 2 butir 18.18 % 11 butir Rata-rata 4.67 butir 47.20 % 3.00 butir 30.92 % 2.33 butir 21.87 % 10.00 butir