UJI PENDAHULUAN AKTIMTAS ANTICENDAWAN DARI
PROTEIN TOTAL AKAR BERAMBUT
Trichosanthes cucumerina
L.
AbstrakUji pendahuluan aktivitas anticendawan dari protein total yang diekstraksi dari akar berambut Trichosanthes cucumerina L. dilakukan secara in vitro. Protein total hasil ekstraksi
dari
akar berambut pada beberapa media perlakuan, diendapkan dengan ammonium sulfat, dikeringdinginkan lalu digunakan dalam pengujian. Protein yang diuji terdiri dari protein dari akar berambut hasil perlakuan kasein hidrolisat (0, 50, 100 dan 150 mg/l), sukrosa (20, 30, 40, dan 50 dl), sukrosa 40 g/l (hasil panen sub kultur I, 11, dan 111) dan 50 gA (hasil panen sub kultur I, 11, dan 111). Sehingga terdapat 14 sumber protein. Cendawan yang digunakan adalah cendawan patogen tanaman meliputi Fusarium sp. Sclerotium rolfsii, Rhizoctonia solani, dan Colletotricum gloeosporoides dan Colletotricum conyzoides..Metode pengujian dengan cara melarutkan 40, 70 dan 160 mg protein dalam 500 yl buffer fosfat dingin, kemudian larutan protein tersebut diaplikasikan sebanyak 20 y1 disekitar koloni cendawan yang berumur 1-2 hari melalui sebuah sumur berjarak 0.5 cm dari ujung koloni.
Hasil pengujian menunjukkan protein kasar belurn mampu menghambat perturnbuhan sebagian besar cendawan yang di uji. Indikasi pengharnbatan terlihat pada Fusarium sp. dengan protein asal perlakuan sukrosa 50 g/l panen sub kultur ke-I11 dan Sclerotium rolfsii dengan protein asal akar berambut hasil perlakuan sukrosa 40 g/l pada panen sub kultur ke-11. Narnun indikasi penghambatan pada Fusarium sp. tidak menunjukkan alur penghambatan yang spesifik terhadap pertumbuhan koloni cendawan tersebut. Sedangkan pada Sclerotium rolflsii pengaruh indikasi penghambatan terlihat lebih jelas namun belurn ditemukan secara konsisten pada semua ulangan.
Pendahuluan
Aka. tanaman di lapangan hampir selalu mengalami stress dari lingkungan antara lain dalam bentuk serangan berbagai patogen. Karena itu, akar baik secara terinduksi maupun konstitutif kemunglunan mempunyai mekanisme ketahanan tertentu terhadap patogen sehingga akar tetap dapat hidup dan menyokong pertumbuhan tanaman. Maeshima et al. (1995) dan Bowles et al. (1990) mengemukakan bahwa akar tanaman secara spesifik mensintesa dan mengakumulasi makromolekul seperti storage proteins dan protein yang berhubungan dengan mekanisme ketahanan seperti glucanohydralases, chitinase, glucanases dan sebagainya.
Akar berarnbut yang dihasilkan melalui infeksi tanaman dengan
Agrobacterium rhizogenes merupakan sarana yang ideal
untuk
mengelusidasi metabolisme spesifik pada akar tanaman. Hal ini disebabkan akar berambut secara stabil mengekspresikan lintasan biosintetik spesifik akar dan telah digunakan secara intensif untuk studi metabolit molekul rendah seperti alkaloid, flavonoid, polyacetilene dan sesquiterpen fitoaleksin (Savary dan Flores, 1994).Savary dan Flores (1994) telah melaporkan adanya protein bioaktif yang dihasilkan dari kultur akar berambut tanaman Trichosanthes kirilowii var.
japonicum. Sebanyak 5 protein tennasuk kedalam chitinase kelompok 111, 3 protein ditemukan dalam media kultur (ekstraselluler) dan 2 protein intraselluler. Protein yang dihasilkan tersebut bereaksi dengan antiserum dari Chitinase
kelompok I11 dari ketimun, yang menunjukkan identitasnya sebagai chitinase.
Protein lainnya yang dihasilkan termasuk ke dalam farnili Patogen Related Proteins (PRPs), bereaksi kuat dengan antiserum dari osmotin. Fraksi protein
protein ekstraseluler basic dan acidic yang mengandung chitinase dan permatin
menunjukkan aktivitas anticendawan yang kuat.
Berhubungan dengan adanya potensi aktivitas anticendawan dari protein yang dihasilkan akar berambut dari Trichosanthes sp., maka penelitian ini
dilakukan untuk mengetahui apakah protein kasar yang hekstraksi dari akar berambut Trichosanthes cucumerina mempunyai aktivitas anticendawan.
Bahan dan Metode Preparasi Protein
Protein diekstraksi dari akar berambut yang ditanam dalarn beberapa media perlakuan yaitu kasein hidrolisat (0, 50, 100, dan 150 mgll), sukrosa (20, 30, 40 dan 50 gll), dan sukrosa 40 dan 50 g/l dalam 3 kali sub kultur, sehingga terdapat 14 sumber protein. Protein Qekstrak dari akar berambut dengan menggerus contoh yang dianalisis (0.2 g bobot kering) dalam 10 ml buffer fosfat (NafI2P04 10 rnM, Na2HP04 15 mM, KC1 100 mM, Na2EDTA 2 rnM dan PVP 0.75%, pH 7.5) dingin dan disentrifugasi dengan kecepatan 8000 rpm selama 15 menit. Supernatan di ambil dan ke dalam endapan dtambahkan kembali 10 ml bdfer fosfat dan disentrifugasi kembali pada suhu 4°C dengan kecepatan 8000 rpm selama 15 menit.. Supemtan yang didapat digabungkan dan ditera hingga 25 ml dengan penambahan buffer fosfat. Protein diendapkan dengan penambahan ,ammonium sulfat kejenuhan 60%, lalu disentrifus pada 15 000 rpm, suhu 4OC , selama 15 menit. Pelet diambil dan dikeringdinginkan dengan fieeze dryer,
lcemudian Qsimpan dalarn deep freezer suhu 80°C sebelum digunakan untuk
Preparasi Kultur Cendawan
Cendawan yang digunakan dalam uji ahvitas protein terdiri dari
Fusarium sp. Sclerotium rolfsii, Rhizoctonia solani, Colletotrichum gloeosporoides dan Colletotrichum conyzoides. Cendawan tersebut ditanam dalam
media Potato Dekstrose Agar (PDA) yang ditambahkan 25 mg/l cloramphenicol.
Isolat cendawan ditanam dalam media PDA
dan
diinkubasikan dalam suhu ruangan selama 2 han dalam kondisi gelap. Pada umur dua hari, cendawan diambil pada bagian pinggir koloni dengan borer (diameter 5 mrn) dan ditanam dalam media PDA yang baru. Koloni cendawan berumur dua hari digunakan untuk pengujian.Pengujian Aktivitas Protein
Uji aktivitas anticendawan dari protein kasar dilakukan dengan plate disk methods (Cheong et al., 1997). Koloni umur 1-2 hari dengan ukuran diameter
koloni 1-1.5 cm dari masing-masing cendawan yang diuji digunakan dalam pengujian. Di sekitar koloni dibuat empat buah sumur berjarak 0.5-1 cm dan
ujung koloni. Sumur dibuat dengan borer berdiameter 5 mm. Metoda pengujian seperti terlihat pada Gambar 9.
Protein dari masing-masing swnber perlakuan akar berambut ditimbang sebanyak 40, 70, dan 160 mg. Protein dilarutkan dalam 500 pl buffer fosfat dingin. Sebanyak 20 yl larutan protein dari masing-masing konsentrasi dimasukkan ke dalam sumur yang telah disiapkan. Sebagai kontrol ke dalam sumur keempat dimasukkan 20 pl b a e r fosfat sedangkan kontrol lainnya adalah koloni cendawan tanpa aplikasi protein. Kultur cendawan diinkubasi dalam ruang gelap bersuhu 2527°C.
Pengujian dilakukan dua kali untuk masing-masing cendawan. Setiap ulangan terdiri dari 2-3 satuan percobaan. Pengamatan
&lllakukan
24 dan 48 jam setelah aplikasi protein dengan melihat ada tidaknya efek penghambatan dari perlakuan protein terhadap perturnbuhan koloni.Garnbar 9. Metode uji aktivitas anticendawan dari protein total akar berambut; a. sumur dengan kontrol buffer, b. sumur dengan protein 1.6 mg/20 p1, c.protein 3.2 mg/20 p1, d. protein 6.4 mg/20 pl, e. koloni cendawan berumur 1-2 hari dengan ukuran diameter koloni 1
-
1.5 cm.Hasil dan Pembahasan
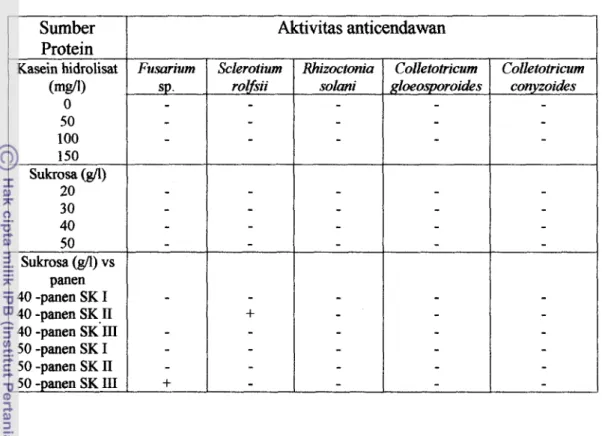
Hasil pengujian aktivitas anticendawan pada sebagian besar cendawan yang digunakan, menunjukkan hasil yang negatif (Tabel lo), kecuali pada
Fusarzum sp. dan Sclerotium rolfsslr terlihat adanya indikasi penghambatan seperti
terlihat pada Gambar 10. Narnun pada Fusarrum sp. tidak terlihat alur
pengharnbatan yang jelas pada pertumbuhan koloni cendawan.
Aktivitas penghambatan pada ScIerotrum ro&z terlihat dari terharnbatnya
pertumbuhan hifa dan pembentukan sklerotia, sedangkan pada kontrol ha1 tersebut tidak te rjadi. Hifa terlihat seperti mengalami kerusakan terutama pada bagian ujung yang sangat berperan bag1 perkembangan koloni cendawan. Penghambatan perturnbuhan hifa tidak mernbentuk alur yang spesifik, narnun intensitas hambatan terlihat kuat antara surnur b dan c, karena pengaruh protein dari kedua sumur tersebut.
Sclerotzum ro@n merupakan cendawan terbawa tanah, mempunyai inang lebih
dari
sekitar 500 spesies monokotil clan Qkotil, dm merupakan patogen perusak pada beberapa tanaman yang mempunyai nilai ekonomi pentingScIerotrum rolfsrr merusak tanaman diantaranya kacang tanah dengan gejala awal
cabang atau seluruh tajuk tanaman menjadi layu dan menguning, warm daun
menjadi coklat gelap, akar dan pangkal batang menjadi coklat dan busuk. Miselia cendawan turnbuh dengan cepat dalam kondsi hangat dan lembab. Cendawan tersebut menghasilkan sklerotia yang pada awalnya benvarna putih kemudian menjadi coklat (Benhamou dm Chet, 1996).
Tabel 10. Hasil uji aktivitas anticendawan pada beberapa sumber protein dari akar berambut Trichosunthes cucumerina L.
Dalam uji ahvitas protein ini, sklerotia pada Sclerotium rolfii terbentuk lebih awal dibanQngkan kontrol. Hal ini diduga sebagai mekanisme adaptasi
Sclerotium karena adanya stress akibat perlakuan protein. Sklerotia merupakan
alat untuk mempertahankan Qri bagi Sclerotium dalarn konQsi yang kurang menguntungkan dan segera kembali berkecambah dan tumbuh ketika keadaan lingkungan sesuai untuk pertumbuhannya.
Mekanisme penghambatan protein kasar terhadap pertumbuhan Sclerotium
rolfsii dalam uji in vitro belum dapat dipastikan, karena protein yang diuji masih
merupakan protein total. Namun beberapa protein anticendawan seperti chitinuse, dan /3 1,3 glucanase menyebabkan degradasi enzimatik dari chitin (polimer
karbohidrat) pada dinding sel cendawan (Yun et al. 1996).
Surnber Protein Kasein hidrolisat 0 50 100 150 Sukrosa (gA) 20 30 40 50 Sukrosa (gll) vs panen 40 -panen SK I 40 -panen SK I1 40 -panen SK'III 50 -panen SK I 50 -panen SK I1 50 -panen SK I11 Aktivitas anticendawan Fusarium sp . -
-
+
Sclerotium rolfsii-
-
+
Colletotricum gIoeosporoi&s-
-
- Rhizoctonia solmi-
-
-
-
-
- Colletotricum conyzoides --
-
-
GambarlD. Hasil uji aktivitas anticendawan dari protein total akar berambut Trichosanthes cucumerina L. A. Sclerotium rolfsii tanpa perlakuan protein (kontrol), B. Sclerotium rolfsii dengan perlakuan protein (a. sumur dengan kontrol buffer, b. protein 1.6 mg, c. protein
3.2 mg, c. protein 6.4 mg). C. Fusarium sp. tanpa perlakuan protein (kontrol), D. Fusarium sp.dengan perlakuan protein.
Mekanisme lain dari protein aktif anticendawan adalah dengan penipisan u j u q pertumbuhan cendawan. Hal tersebut dilaporkan dari Chitinase kelompok I
pertumbuhan fungi, diikuti oleh pembengkakan seperti balon dan b e r a b r dengan pecahnya hifa (Neuhaus dalam Datta dan Muthukrishnan,l999) . Cheong et al.
(1997) mengemukakan bahwa protein anticendawan PR-5 yang dihasilkan dari tanaman waluh yang juga dikenal sebagai thaumatin-likes protein menunjukkan aktivitas anticendawan dengan merusak membran fungi.
Adakalanya kombinasi antara chitinase dan
P
1,3 glucanase lebih efektif dalam menghambat pertumbuhan cendawan. Hal ini dilaporkan Jongedijk et al.(1995) dalam uji in vitro maupun in vivo pada tanaman tomat transgenik yang .mengekspresikan chitinase kelompok I dan
P
1,3 glucanase.Protein aktif kelompok RIPs merupakan N-glycosidase yang mampu memotong ikatan N-glycosidic dari adenin dalam sequens RNA ribosom yang spesifik. RIPs ada yang merupakan protein rantai tunggal (RIPs tipe I) dan ada yang mempunyai galactose-specrfic lectin domain yang menempel ke permukaan sel (RIP tipe 11) yang bersifat toksik (Stirpe, et al. 1992). Aktivitas anticendawan dari RIPS telah dilaporkan pada RIP 30 dari barley yang menghambat pertumbuhan Trichoderma reesei dan Fusarium sporotrichioides (Jensen et al.,
1999).
Berdasarkan uji aktivitas anticendawan dari protein kasar akar berambut
Ttrichosanthes cucumerina L. dalam penelitian ini, belum dapat disimpulkan apakah protein yang mempunyai aktivitas anticendawan terdapat dalam protein tersebut. Hal ini disebabkan hasil uji yang sebagian besar menunjukkan hasil yang negatif.
Berbagai faktor dapat mempengaruhi keberhasilan pengujian, mulai dari metoda penyiapan protein, kadar protein yang aktif dalarn protein total yang
mungkin sangat rendah, jenis patogen yang diuji (sensitif atau tidak terhadap protein yang diuji), metode pengujian, dan lain sebagainya.
Faktor-faktor yang berhubungan dengan aktivitas protein juga
P
mempengaruhi, seperti pH, suhu, ataupun garam-garam an organik. Menurut Marcus et al. (1997), aktivitas antimikroba dari senyawa antimicrobial peptide umumnya terjadi dalam kehadiran garam kalsium dan potasium.
Kesim pulan
Hasil uji aktivitas anticendawan clan protein kasar akar berambut belum menunjukkan hasil yang positif, kecuali sedikit indikasi penghambatan pada pertumbuhan Fusarium sp. dan Sclerotiurn rolfsii. Penghambatan pada Fusarium sp. tidak menunjukkan
d w
yang spesifik dan pada Sclerotium rolfsii penghambatan terlihat dengan rusaknya hifa dan munculnya sklerotia. Indikasi penghambatan belum konsisten pada beberapa ulangan.Berbagai faktor dapat mempengaruhi keberhasilan pengujian seperti preparasi protein, jenis patogen yang diuji, metode pengujian dan berbagai faktor yang mempengaruhi aktivitas yang belum dapat dikontrol dengan baik.
Daftar Pustaka
Benhamou N, Chet I. 1996. Parasitism of Sclerotium rolfsii by Trichoderma harzianum: ultrastructural and cytochemical aspects of the interaction. Biochem Cell Biol86;4:405-4 16.
Bowles DJ. 1990. Defense-related protein in higherplants. Annu Rev Biochem 59:873-907.
Cheong NE et al. 1997. Purification and characterization of an antifungal PR-5 protein from pumpkin leaves. Moll Cells 7:2 14-2 19.
Harlton CE, Levesque CA, Punja ZK. 1995. Genetic diversity in ScleroMum (Athelia) rolfsii and related species. Mol Plant Path01 85; 10: 1269- 1 28 1.
Jensen AB, Leah R, Chaudry B, Mundy J. 1999. Ribosome-Inactivating Proteim:structure, function and engineering. Di dalarn: Datta SK, Muthukrishnan S, editor. Pathogenesis-Related Proteins in Plants. Boca Raton, London, New York, Washington, D.C.:CRC Press. hlm 77-105. Jongedijk E et al. 1995 Synergistic activity of chitinases and
P
1,3 glucanasesenhances fungal resistance in transgenic tomato plants. Euphytica 85: 173- 180.
Maeshima M, Sasaki T, Asahi T. 1985. Characterization of major protein in sweet potato tuberous root. Phytochemistry 24: 1899- 1902.
Marcus JP, Goulter KC, Green JL, Harrison SJ, Manners JM. 1997. Purification, characterization and cDNA cIoning of an antimicrobial peptide from Macademia integrifoEia. Eur J Biochem 244:743-749
Neuhaus JM, 1999. Plant Chitinases (PR-3, PR-4, PR-8, PR-11) Di dalam: Datta SK, Muthukrishnan S, editor. Boca Raton, London, New York, Washington, D.C.: CRC Press. hlm 77-105
Savary BJ, Flores HE. 1994. Biosynthesis of defense-related protein in transformed root cultures of Trichosanthes kirilowii Maxim. Var. japonicum (Kitam). Plant Physiol106: 1 195- 1204
Stirpe F, Barbieri L, Batteli MG, Soria M, Lappi D. 1992. Ribosome-Inactivating Proteins from plants: present status and future prospects. Bioltech 10 :405- 412.
Yun DJ et al. 1996. Novel osmotically induced antifungal chitinases and bacterial expression of an active recombinant isoform. Plant Physiol 11 1:1219-